Introduction
In terms of incidence, gastric cancer is the fourth
most common type of malignant tumor, with the second highest
mortality rate (1,2), as ~10% of patients with cancer succumb
to gastric cancer each year worldwide. Although novel
chemotherapeutic drugs have emerged to improve the treatment of
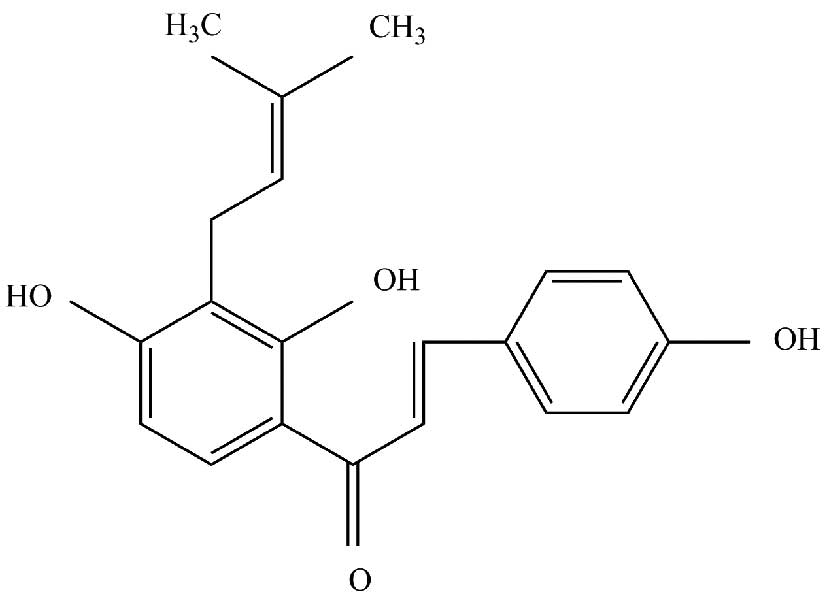
gastric cancer, the 5-year survival rate remains low (3). Isobavachalcone (IBC; Fig. 1) is an active ingredient isolated
from Psoralea corylifolia. This Chinese herbal medicine
belongs to the natural chalcones and as such has anti-inflammatory
and anti-fungal effects. Previous studies have demonstrated that
IBC is capable of inducing apoptosis in cancerous ovarian, breast,
lung, and other tumor cells, whereas it is non-toxic to normal
cells, including L-02 and human umbilical vascular endothelial
cells (HUVECs) (4,5). It has been suggested that IBC may be
used clinically as an efficient anti-tumor drug with low toxicity
(4); however, the exact molecular
mechanism has yet to be elucidated.
The main characteristic of malignant tumors is that
tumor cells can undergo unrestricted proliferation and apoptosis.
The B-cell lymphoma (Bcl)-2 gene family is an important group of
apoptosis-associated genes, which can encode anti-apoptotic
proteins (Bcl-2) and pro-apoptotic proteins [Bcl-2-associated X
protein (Bax)]. The expression of Bcl-2 protein is generally low in
normal cells, but abnormally high in tumor cells.
In order to expound the molecular mechanism of
IBC-induced apoptosis in gastric cancer cells and to explore
potential biological targets, various concentrations of IBC were
used to treat MGC803 cells in the present study. The effects on
apoptosis, migration and invasion, and the alterations in the
expression levels of apoptosis-related proteins in the Akt and
extracellular signal-regulated kinase (Erk) pathways were
investigated.
Materials and methods
Cell culture and treatment
MGC803 human gastric cancer cells were purchased
from American Type Culture Collection (Manassas, VA, USA) and
cultured in RPMI 1640 (Gibco; Thermo Fisher Scientific Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific Inc.) and 100 U/ml
penicillin/streptomycin (Nanjing KeyGen Biotech Co. Ltd., Nanjing,
China). Cells were maintained in the exponential phase of growth at
37°C in a humidified atmosphere containing 5% CO2. All
experiments were performed using cells in the log phase. IBC
(2′,4′,4-trihydroxy-3′-[3′-methylbut-3′-ethyl]chalcone) was a
generous gift from the Dalian Institute Of Chemical Physics, also
known as the Chinese Academy Of Sciences (Dalian, China). IBC was
dissolved in dimethyl sulfoxide (DMSO) to generate a 100 mM stock
solution, and was diluted to a final concentration of 0.1% in
culture media.
Cell proliferation analysis
Cell viability was determined using a
methylthiazolyltetrazolium (MTT) assay (Sigma-Aldrich, St. Louis,
MI, USA). A total of 6×103 MGC803 cells/well were seeded
into 96-well cell culture plates with complete growth medium and
maintained for 24 h. The treatment groups were treated with 10–60
µM IBC, and the control groups were incubated with RPMI 1640
culture media, and were cultured for 24 or 48 h at 37°C. The
control groups included a blank control, treated with corresponding
cell culture medium, and a negative control, treated with DMSO.
Cells were incubated with 20 µl MTT solution (5 mg/ml) for 4 h at
37°C in an atmosphere containing 5% CO2. The formazan
crystals were subsequently dissolved in 200 µl DMSO. The optical
density (OD) was detected by measuring absorbance at 490 nm using a
microplate reader (xMark; Bio-Rad Laboratories Inc., Hercules, CA,
USA). Survival rate was determined using the following calculation:
Survival rate = (experimental group OD value - negative control
group OD value)/(blank control group OD value - negative control
group OD value) × 100%. The half-maximal inhibitory concentration
(IC50) values of IBC were calculated using SPSS version
16.0 for Windows (SPSS, Inc., Chicago, IL, USA). All experiments
were repeated in triplicate, the mean values were calculated and
the cell proliferation curves were generated.
Cell morphological analysis
A total of 5×105 MGC803 cells/well were
seeded in 6-well plates and incubated overnight at 37°C. Following
a 48 h reaction with 40 µM IBC, cells from the negative control and
treatment groups were collected and centrifuged at 300 × g for 5
min. Cell smears were subsequently prepared and stained with
Wright-Giemsa solution (Sigma-Aldrich) for 10 min, washed with
running water and dried. Cell morphological changes were observed
and images were captured using a CKX31 light microscope (Olympus
Corp., Tokyo, Japan).
Cell apoptosis analysis
A total of 5×105 MGC803 cells/well were
cultured in 6-well plates overnight and treated with various
concentrations (0, 20 and 40 µM) of IBC for 48 h. Following
treatment, the cells were washed twice with phosphate-buffered
saline (PBS) and resuspended in 200 µl binding buffer.
Subsequently, the cell suspension was stained with Annexin
V-fluorescein isothiocyanate (FITC) and propidium iodide (Nanjing
KeyGen Biotech Co., Ltd.) according to the manufacturer's protocol,
prior to incubation for 10 min at 37°C in the dark. A FACSVerse
flow cytometer (BD Biosciences, San Jose, CA, USA) was used to
analyze the population of Annexin V-FITC positive cells.
Wound healing analysis
A total of 8×105 MGC803 cells/well were
seeded in 6-well plates and cultured in complete medium at 37°C.
Upon confluent monolayers, the cells were scratched with a sterile
20 µl pipette tip to create a denuded zone through the central axis
of the plate. Subsequently, the cells were incubated with
serum-free medium supplemented with various concentrations (0, 20
and 40 µM) of IBC at 37°C. The distances of wound healing were
observed and images were captured (CKX31 microscope) at 0 and 48 h
following treatment. Cell relative motility was calculated using
the following formula: Cell relative motility = (average area of
original wound - average area of 48 h wound)/average area of
original wound × 100%.
Cell invasion analysis
Cell invasion analysis was conducted using 24-well
Transwell® chambers [unit pore size, 8 µm with polyvinylidene
fluoride (PVDF)] and Matrigel™ (BD Biosciences). A total of 60 µl
Matrigel™ diluted 1:5 in serum-free medium was added to each well 4
h prior to cell seeding onto the upper chamber. MGC803 cells were
cultured in serum-free medium with various concentrations (0, 20
and 40 µM) of IBC in the upper chamber; whereas the lower chamber
was filled with medium containing 10% FBS. Following 48 h, the
cells that had migrated through the PVDF membrane and Matrigel™ to
the lower surface were fixed with 4% formaldehyde and stained with
0.5% crystal violet. Cells were visually counted in five random
fields using an IX51 inverted microscope (Olympus Corp.).
Western blot analysis
A total of 1×106 MGC803 cells/well were
cultured in 6-well plates overnight and treated with 0–40 µM IBC
for 48 h. Following this, the cells were lysed using lysis buffer
[50 mM Tris; 150 mM NaCl; 1% Triton X-100; 1% deoxycholate; 0.1%
sodium dodecyl sulfate (SDS); and 2 mM PMSF; pH 7.4] and total
cellular proteins were prepared. Protein concentrations were
determined using a bicinchoninic acid protein assay (Nanjing KeyGen
Biotech Co., Ltd.), using bovine serum albumin as a standard. Equal
amounts of protein (20 µg) were separated by 12% SDS-polyacrylamide
gel electrophoresis and transferred to PVDF membranes (GE
Healthcare, Amersham, UK). Following 1-h incubation at 37°C in a
blocking buffer of PBS with Tween-20 (10 mM Tris-HCl, 150 mM NaCl
and 0.1% Tween-20). and 5% nonfat dry milk, the membranes were
incubated with the following primary antibodies: Akt (1:200; cat.
no. sc-8312); phosphorylated (p)-Akt (1:200; cat. no. sc-135650);
Erk (1:200; cat. no. sc-94); p-Erk (1:200; cat. no. sc-16982);
Bcl-2 (1:300; cat. no. sc-492); Bax (1:300; cat. no. sc-493);
caspase-3 (1:200; cat. no. sc-7184; used for active caspase-3 and
pro-caspase-3); and β-actin (1:1,000; cat. no. sc-1616; all from
Santa Cruz Biotechnology, Inc., Dallas, TX, USA), overnight at 4°C.
The membranes were then treated with horseradish
peroxidase-conjugated secondary antibody (1:800; cat. nos. sc-2954
and sc-2955; Santa Cruz Biotechnology, Inc.) for 2 h. The blots
were incubated with a chemiluminescence substrate (Santa Cruz
Biotechnology, Inc.) and the data were corrected for loading using
the values for β-actin. Immunoreactive bands were quantified using
ImageJ2X software (National Institutes of Health, Bethesda, MD,
USA).
Statistical analysis
Three independent experiments were performed, and
statistical analyses were performed using SPSS version 16.0
software. Quantitative variables were analyzed by one-way analysis
of variance and expressed as the mean ± standard deviation.
P<0.05 was considered to indicate a statistically significant
difference.
Results
IBC inhibits the proliferation of
MGC803 cells
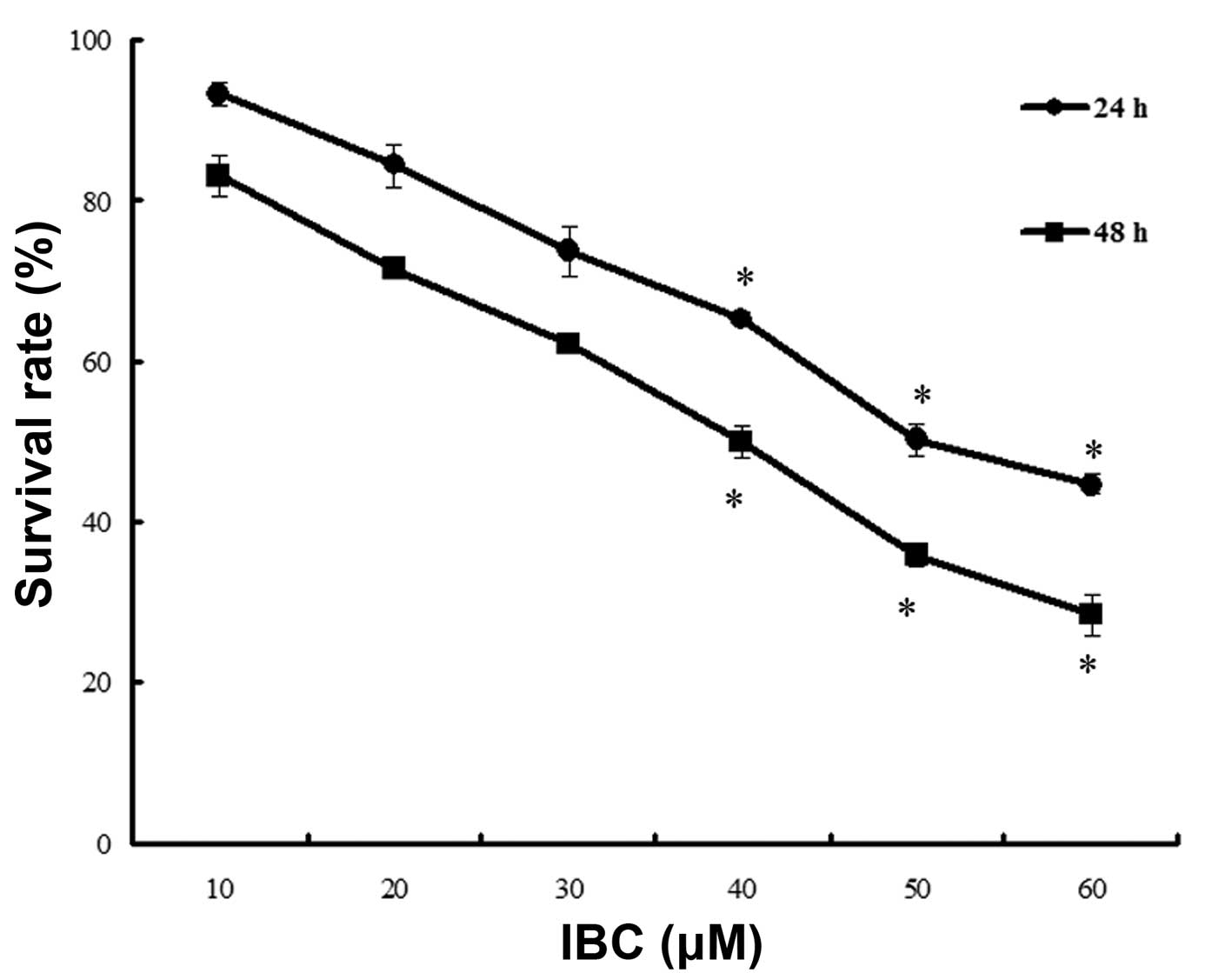
MGC803 cells were cultured in 0–60 µM IBC for 24 and
48 h. The results of the MTT assay demonstrated that IBC inhibited
the proliferation of MGC803 cells in a concentration- and
time-dependent manner (Fig. 2). The
IC50 values at 24 and 48 h were 49.68±4.26 and
39.96±3.78 µM, respectively.
IBC induces apoptosis in MGC803
cells
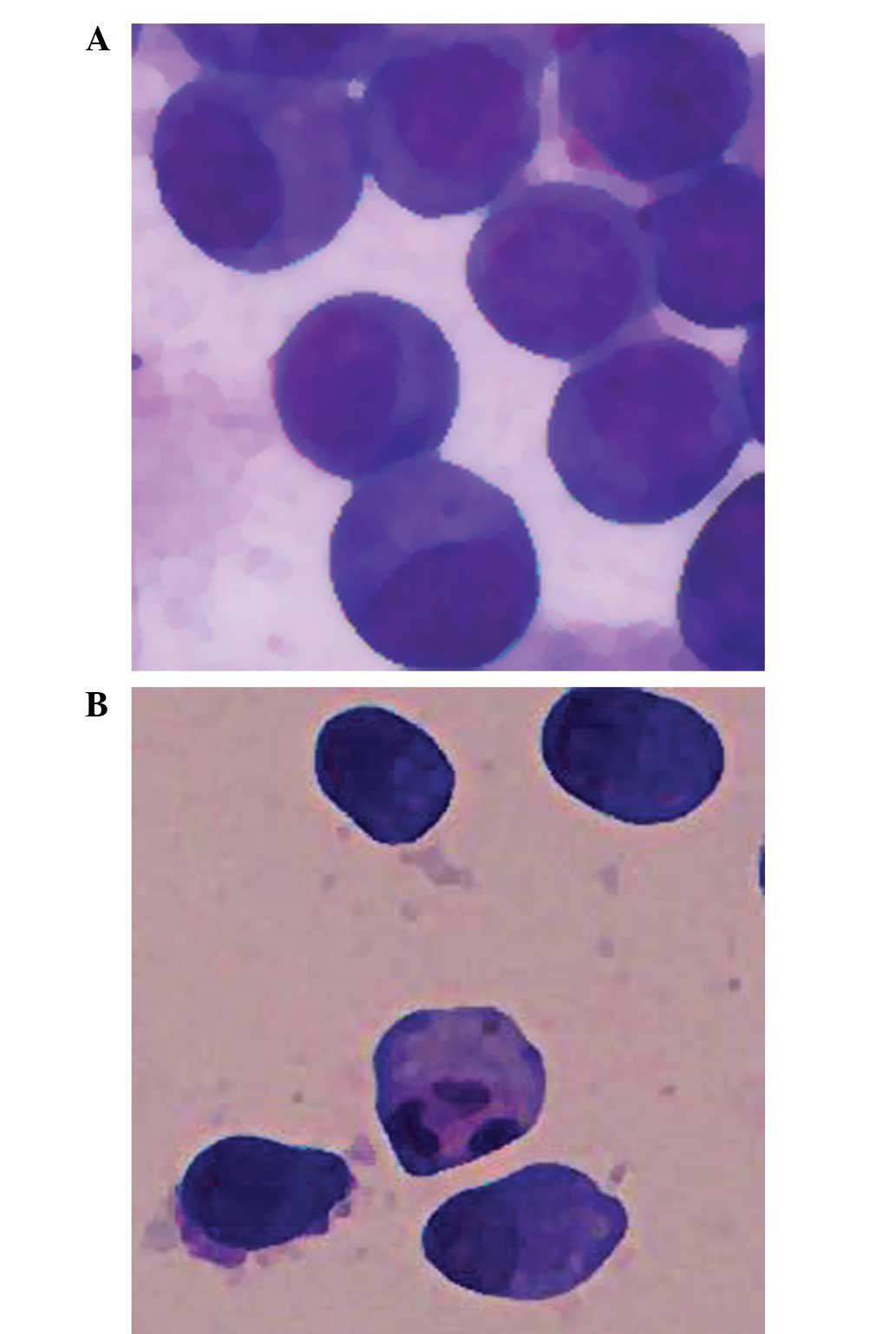
MGC803 cells were cultured in 40 µM IBC for 48 h and
the typical morphological features of apoptosis were detected using
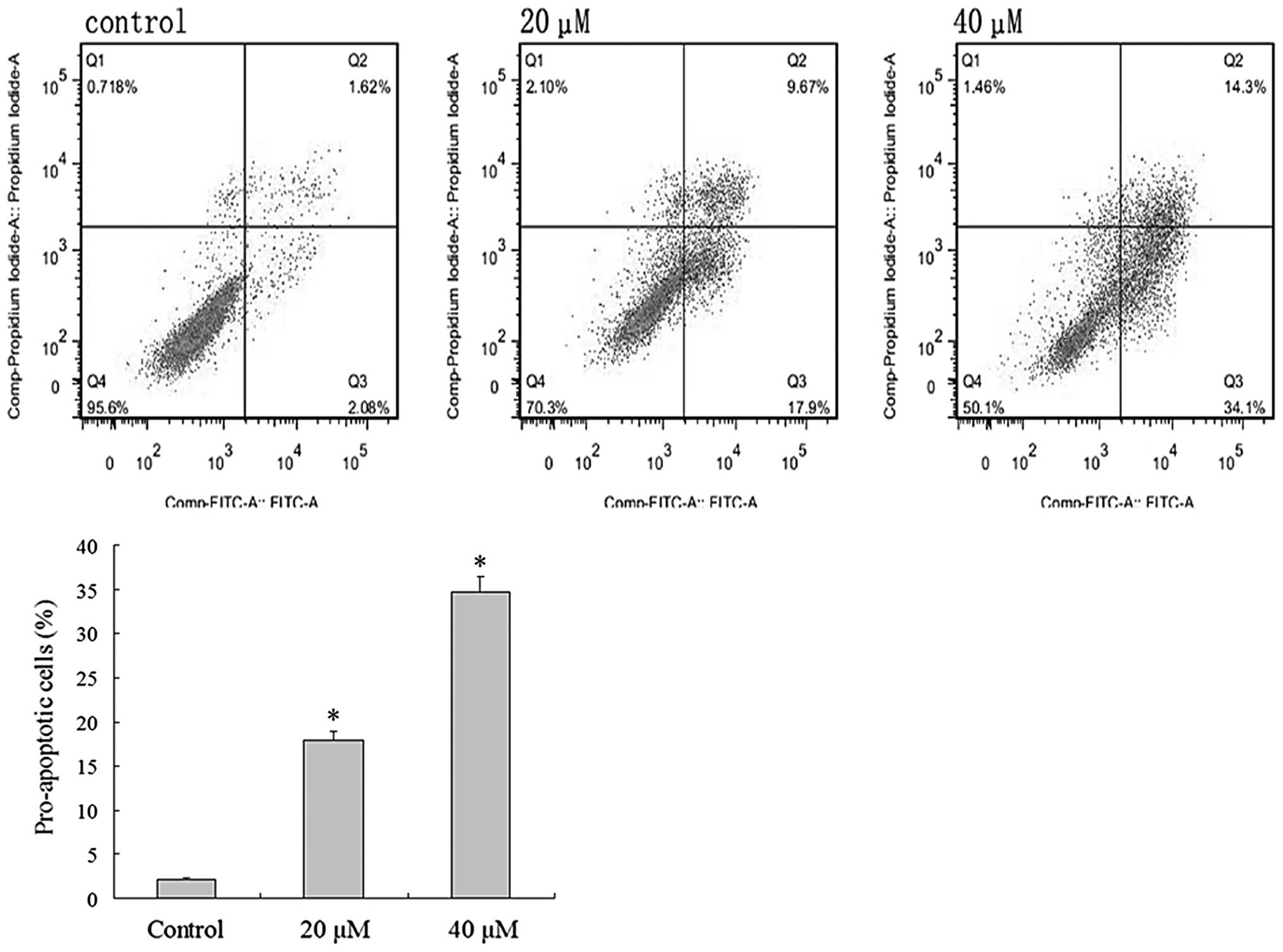
light microscopy, including apoptotic bodies (Fig. 3). Flow cytometric analysis was
undertaken to verify the effect of IBC on the apoptosis of MGC803
cells. Intact cells were used as a control group. Following 48 h,
2.08±0.46% of the counted cells in the control group had become
pro-apoptotic (Fig. 4). Following 48
h of treatment with 20 and 40 µM IBC, the percentage of
pro-apoptotic cells was 17.9±2.65 and 34.1±3.40%, respectively,
significantly greater than that of the control groups (n=3,
P<0.05). These results indicate that IBC treatment may induce
apoptosis in MGC803 cells in a concentration-dependent manner.
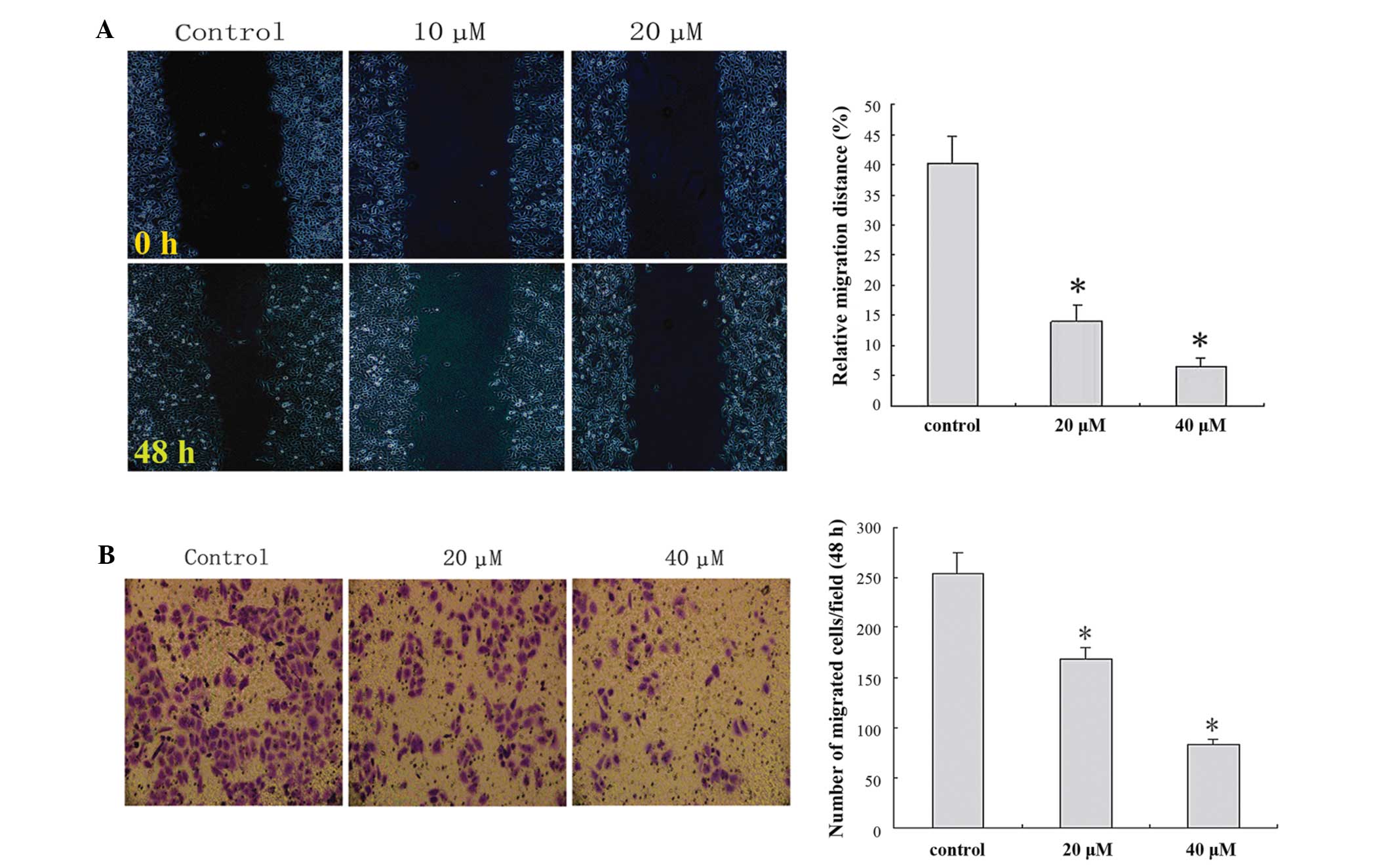
IBC decreases migration and invasion
of MGC803 cells
In order to clarify the important role of IBC in the
migration and invasion of MGC803 cells, wound healing and Matrigel™
Transwell® invasion assays were performed. IBC administration
decreased cell migration and invasion in a concentration-dependent
manner (Fig. 5).
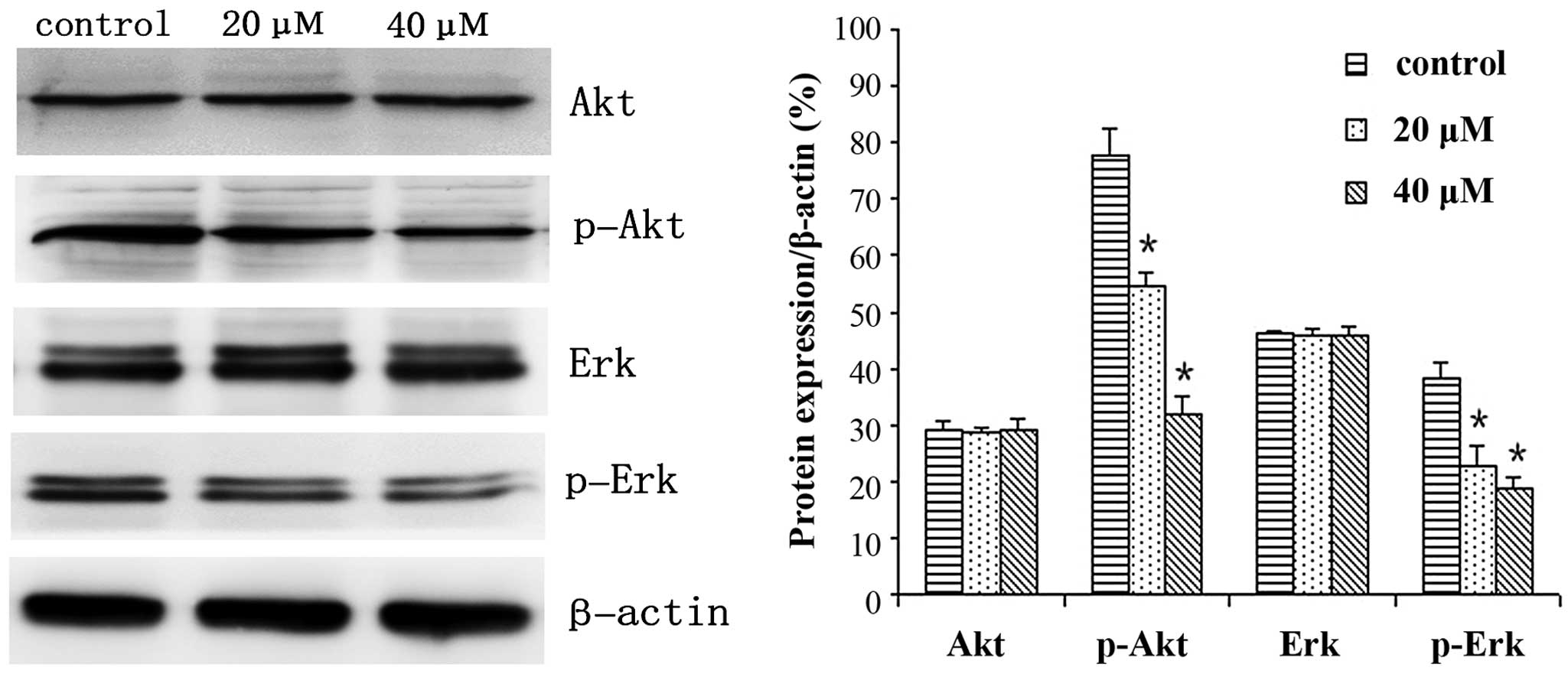
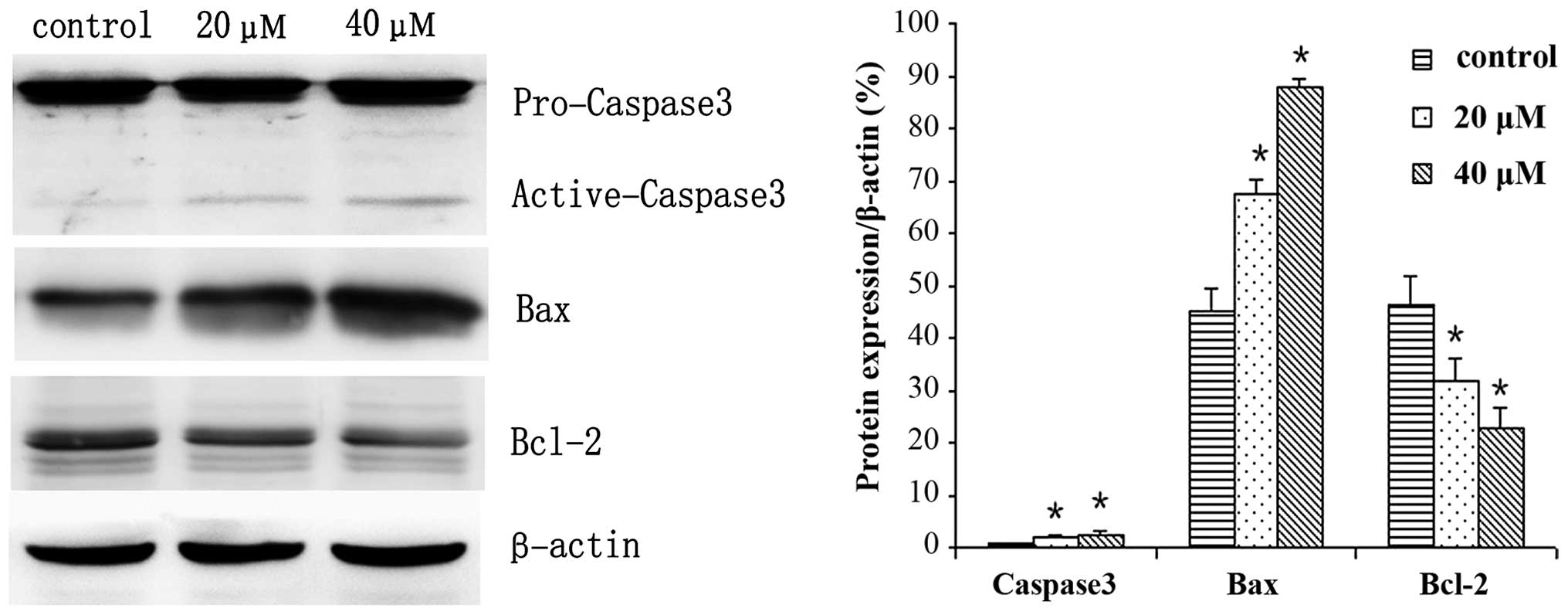
IBC regulates the expression levels of
proteins in MGC803 cells
MGC803 cells were treated with 0, 20 or 40 µM IBC
for 48 h and subjected to western blot analysis. IBC decreased
Bcl-2, p-Akt and p-Erk expression levels, whereas Bax and active
caspase-3 protein expression levels were increased in a
concentration-dependent manner (Figs.
6 and 7). As compared with the
control, the Bax/Bcl-2 ratio was 2.17 and 3.96, respectively
(P<0.05). This result was to be expected as Bcl-2 is an
anti-apoptotic protein, whereas Bax is an apoptosis-promoting
protein. Furthermore, following prolonged treatment with IBC, the
expression levels of active caspase-3 increased (Fig. 7).
Discussion
IBC was first isolated from Psoralea
corylifolia in 1968 by Bhalla et al (6). IBC is a chalcone, which possesses
various features, including anti-inflammatory, anti-fungal and
anti-reverse transcriptase activity (5). Previous studies have demonstrated that
IBC also has good anti-tumor activity (7–9).
Furthermore, a previous study demonstrated that IBC is capable of
inducing apoptosis in human neuroblastoma cells, whereas no
significant cytotoxicity was detected in normal rat cerebellar
granule cells following treatment with the same dose of IBC
(10). By observing various solid
tumor cells and normal cell lines, including liver cells, L-02
cells and HUVECs, Jing et al (4) demonstrated that IBC had an obvious
destructive effect on tumor cells, and notably, no toxic side
effects were demonstrated in normal cells. These results indicated
that IBC administration may be a highly efficient treatment for
cancer patients with low toxicity.
The present study observed the effects of IBC on the
proliferation of MGC803 gastric carcinoma cells, and the results
demonstrated that IBC inhibited the proliferation of MGC803 cells
in a concentration- and time-dependent manner. To verify whether
the reduction in cell proliferation was associated with apoptosis,
Wright-Giemsa staining was conducted to observe the morphology.
MGC803 cells were cultured in 20 µM IBC for 48 h. Light microscopy
detected the typical morphological features of apoptosis, including
cytoplasmic condensation, pyknosis and the presence of apoptotic
bodies. In order to validate the role of IBC in the promotion of
apoptosis in gastric cancer cells, flow cytometric analysis was
conducted in the present study. Flow cytometric analysis of early
apoptosis demonstrated that the apoptotic rate of MGC803 cells was
gradually increased with increasing IBC concentration. These
results demonstrated that IBC may promote the apoptosis of MGC803
cells.
Invasion and metastasis are the most lethal features
of cancer, accounting for >90% of cancer-associated mortality
(11). Distant organ or tissue
metastasis is a marker of poor prognosis in patients with gastric
cancer. The results of the present study demonstrated that various
concentrations of IBC were capable of inhibiting the migratory and
invasive ability of MGC803 cells, in a dose-dependent manner.
However, the present study was inadequate in that only the
phenomenon of IBC-induced inhibition of gastric cancer cell
migration and invasion was observed, without further exploration of
the specific molecular mechanisms; therefore, these investigations
are required in future studies.
Apoptosis is a defensive mechanism of the body that
eliminates malignant cells, and has a crucial role in the
prevention of cancer. Notably, the predominant function of numerous
antitumor drugs is the induction of apoptosis in tumor cells, via
the regulation of various apoptosis-related signaling pathways
(12–15).
The Bcl-2 family includes anti-apoptotic proteins,
such as Bcl-2, and proapoptotic proteins, such as Bax and
Bcl-2-associated death promoter (16). The Bcl-2 protein family has an
important regulatory role in the release of cytochrome c
from mitochondria, which activates endogenous apoptosis pathways.
Bax was the first protein to be identified that interacts with
Bcl-2, and is capable of inhibiting the apoptosis-resisting ability
of Bcl-2 (17,18). The present study demonstrated that
IBC decreased the protein expression levels of Bcl-2 and increased
the protein expression levels of Bax to induce the apoptosis of
gastric cancer cells.
The Akt signaling pathways have an important role in
the survival of mammalian cells and resistance to apoptosis
(19). As multicellular organisms,
mammals maintain stable development through mutual checks and
balances of proliferation and apoptosis, and if this balance is
broken, it may lead to cancer. Previous studies have demonstrated
that excessive activation of Akt affects the activity of each of
the downstream effectors, thereby leading to resistance to
apoptosis, and promoting the proliferation of tumor cells (20,21).
Various cellular stimulations are capable of activating Erk1/2,
also known as p44/p42 mitogen activated protein kinase (MAPK); some
may lead to the activation of transcription factors and others may
activate cell proliferation, differentiation, cell cycle regulation
and cell survival-associated serine/threonine protein kinase
(15). Previous large phase I/II
clinical trials have demonstrated that MAPK/Erk inhibitors are
capable of prolonging the survival of patients with various types
of cancer (22,23). The results of the present study
demonstrated that various concentrations of IBC successfully
reduced the protein expression levels of p-Akt and p-Erk in MGC803
cells. Jing et al (4)
demonstrated that IBC may induce apoptosis in various tumor cells
by inhibiting the activity of Akt signaling pathways, which was
consistent with the results of the present study. Therefore, IBC
may induce apoptosis in MGC803 cells via the Akt and Erk signaling
pathways.
In conclusion, the results of the present study
demonstrated that IBC was capable of inhibiting the proliferation
of gastric cancer cells and the induction of apoptosis in a
concentration- and time-dependent manner. Furthermore, IBC may
inhibit the migratory and invasive ability of MGC803 cells. In
terms of the underlying mechanisms, the results of the present
study demonstrated that IBC induced apoptosis in gastric cancer
cells by regulating apoptosis-related proteins and inhibiting Akt
and Erk signaling pathways.
Acknowledgements
The present study was supported by The First
Affiliated Hospital of Liaoning Medical University (Jinzhou,
China). The authors would like to thank Professor Zhi-tu Zhu for
valuable support.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Guggenheim DE and Shah MA: Gastric cancer
epidemiology and risk factors. J Surg Oncol. 107:230–236. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cervantes A, Roda D, Tarazona N, Roselló S
and Pérez-Fidalgo JA: Current questions for the treatment of
advanced gastric cancer. Cancer Treat Rev. 39:60–67. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jing H, Zhou X, Dong X, Cao J, Zhu H, Lou
J, Hu Y, He Q and Yang B: Abrogation of Akt signaling by
Isobavachalcone contributes to its anti-proliferative effects
towards human cancer cells. Cancer Lett. 294:167–177. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Akihisa T, Tokuda H, Hasegawa D, Ukiya M,
Kimura Y, Enjo F, Suzuki T and Nishino H: Chalcones and other
compounds from the exudates of Angelica keiskei and their cancer
chemopreventive effects. J Nat Prod. 69:38–42. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bhalla VX, Nayak UR and Dev S: Some new
flavonoids from Psoralea corylifolia. Tetrahedron Lett.
20:2401–2406. 1968. View Article : Google Scholar
|
|
7
|
Szliszka E, Jaworska D, Ksek M, Czuba ZP
and Król W: Targeting death receptor TRAIL-R2 by chalcones for
TRAIL-induced apoptosis in cancer cells. Int J Mol Sci.
13:15343–15359. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Szliszka E, Czuba ZP, Mazur B, Sedek L,
Paradysz A and Krol W: Chalcones enhance TRAIL-induced apoptosis in
prostate cancer cells. Int J Mol Sci. 11:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li W, Li S, Higai K, Sasaki T, Asada Y,
Ohshima S and Koike K: Evaluation of licorice flavonoids as protein
tyrosine phosphatase 1B inhibitors. Bioorg Med Chem Lett.
23:5836–5839. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nishimura R, Tabata K, Arakawa M, Ito Y,
Kimura Y, Akihisa T, Nagai H, Sakuma A, Kohno H and Suzuki T:
Isobavachalcone, a chalcone constituent of Angelica keiskei,
induces apoptosis in neuroblastoma. Biol Pharm Bull. 30:1878–1883.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jin X, Zhu Z and Shi Y: Metastasis
mechanism and gene/protein expression in gastric cancer with
distant organs metastasis. Bull Cancer. 101:E1–E12. 2014.PubMed/NCBI
|
|
12
|
Khan KH, Blanco-Codesido M and Molife LR:
Cancer therapeutics, Targeting the apoptotic pathway. Crit Rev
Oncol Hematol. 90:6200–219. 2014. View Article : Google Scholar
|
|
13
|
Westin JR: Status of PI/3K/Akt/mTOR
pathway inhibitors in lymphoma. Clin Lymphoma Myeloma Leuk.
14:335–342. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Koehler BC, Jäger D and Schulze-Bergkamen
H: Targeting cell death signaling in colorectal cancer Current
strategies and future perspectives. World J Gastroenterol.
20:1923–1934. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aranda F, Vacchelli E, Eggermont A, Galon
J, Fridman WH, Zitvogel L, Kroemer G and Galluzzi L: Trial Watch,
Immunostimulatory monoclonal antibodies in cancer therapy.
Oncoimmunology. 3:e272972014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Park JW, Choi YJ, Suh SI, Baek WK, Suh MH,
Jin IN, Min DS, Woo JH, Chang JS, Passaniti A, Lee YH and Kwon TK:
Bcl-2 overexpression attenuates resveratrol-induced apoptosis in
U937 cells by inhibition of caspase-3 activity. Carcinogenesis.
22:1633–1639. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sharawat SK, Bakhshi R, Vishnubhatla S,
Gupta R and Bakhshi S: BAX/BCL2 RMFI ratio predicts better
induction response in pediatric patients with acute myeloid
leukemia. Pediatr Blood Cancer. 60:E63–E66. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sjöström J, Blomqvist C, von Boguslawski
K, Bengtsson NO, Mjaaland I, Malmström P, Ostenstadt B, Wist E,
Valvere V, Takayama S, Reed JC and Saksela E: The predictive value
of bcl-2, bax, bcl-xL, bag-1, fas and fasL for chemotherapy
response in advanced breast cancer. Clin Cancer Res. 8:811–816.
2002.PubMed/NCBI
|
|
19
|
Dent P: Crosstalk between ERK AKT and cell
survival. Cancer Biol Ther. 15:245–246. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Briest F and Grabowski P:
PI3K-AKT-mTOR-signaling and beyond: The complex network in
gastroenteropancreatic neuroendocrine neoplasms. Theranostics.
4:336–365. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Polivka J: Jr andJ anku F: Molecular
targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol
Ther. 142:164–175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
McCubrey JA, Steelman LS, Kempf CR,
Chappell WH, Abrams SL, Stivala F, Malaponte G, Nicoletti F, Libra
M, Bäsecke J, et al: Therapeutic resistance resulting from
mutations in Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR signaling pathways.
J Cell Physiol. 226:2762–2781. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Infante JR, Fecher LA, Falchook GS,
Nallapareddy S, Gordon MS, Becerra C, DeMarini DJ, Cox DS, Xu Y,
Morris SR, et al: Safety, pharmacokinetic, pharmacodynamic, and
efficacy data for the oral MEK inhibitor trametinib: A phase 1
dose-escalation trial. Lancet Oncol. 13:773–781. 2012. View Article : Google Scholar : PubMed/NCBI
|