Introduction
Ibuprofen is an over-the-counter non-steroidal
anti-inflammatory drug (NSAID) (1),
which is predominantly used as an analgesic, anti-inflammatory and
antipyretic compound (2). Ibuprofen
has previously been demonstrated to limit the production of
pro-inflammatory cytokines and to exert neuroprotective effects
(1). Furthermore, ibuprofen has
demonstrated effectiveness in the treatment of Parkinson's disease
(PD) (3), neuroinflammation
associated with β-amyloid (Aβ) deposition (4), and motor deficits in hepatic
encephalopathy (5), due to its
anti-inflammatory and antioxidant properties. In addition, it may
reduce the risk of developing Alzheimer's disease (AD), as
increased expression levels of inflammatory cytokines have
previously been associated with AD (6).
The pathogenesis of various neurodegenerative
disorders is attributed to neurotoxicity, which is associated with
various factors, including aluminum (Al) (7). Previous studies have demonstrated a
role for Al in dementia (8), PD
(9) and AD (10). Furthermore, an aluminum chloride
(AlCl3)-induced model has previously been established
for the investigation of neurotoxicity (11), AD (12), dementia (13), and neuroinflammation (14).
Neuronal pentraxins (NPs), which have key roles in
neuroinflammation (15), belong to a
family of proteins that are homologous to immune system proteins,
including C-reactive and acute phase proteins, and are believed to
be involved in synaptic functions (16). The NPs, including NP1, NP2 and NP
receptor (NPR) (17), are proteins
that were initially identified due to their ability to bind to the
snake poison taipoxin (18) and
mediate its internalization, leading to cell death. As taipoxin
blocks neuromuscular transmission at the pre-synaptic junction
(19), it has been suggested that
NPs may have a role in the neuronal uptake process (18). NP2 is a secreted glycoprotein that
has been demonstrated to enhance the growth of neuronal dendrites
(20), and does not bind to
taipoxin; thus suggesting that it has a distinct function from NP1
(20).
NP1 is expressed in the neurons of the cerebellum,
hippocampus and cerebral cortex (21), whereas NP2 is widely expressed in the
testis, liver, pancreas, skeletal muscles, heart and brain
(20). Furthermore, the expression
of NP2 has been demonstrated to be upregulated in response to
neuronal activity, upon which it may mediate clustering of
2-amino-3-(3-hydroxy-5-methyl-isoxazol-4-yl) propanoic acid
receptors (22). NP1, NP2 and NPR
have 50% identity with each other and their C-terminal halves have
20–30% identity with immune system pentraxins, including the serum
amyloid protein (23), C-reactive
protein (24) and long pentraxin 3
(25).
Previous studies have suggested that NPs may
contribute to the pathogenesis of neurodegenerative disorders,
including AD (26) and PD (16). A predominant characteristic of the AD
brain is accumulated Aβ protein, which interferes with synaptic
functions and induces neuronal cell death (27). Aβ toxicity has been demonstrated to
initiate the overexpression of NP1, which is an apoptotic protein
(28), resulting in the activation
of programmed cell death due to loss of neuronal functions
(26). In addition, NP2 has been
demonstrated to be markedly upregulated in PD (16); thus suggesting an important role for
NPs in neuroinflammatory pathways. The present study aimed to
analyze the effects of ibuprofen on the expression levels of NPs,
and inflammation in an AlCl3-induced mouse model of
neurotoxicity.
Materials and methods
Agents
Aluminum chloride hexahydrate (AL0770) was obtained
from Sigma-Aldrich (St. Louis, MO, USA). Sodium chloride (SO0225)
was purchased from Scharlab, S.L. (Barcelona, Spain). Ibuprofen
tablets (600 mg; RN: 007850) were purchased from Abbot
Laboratories, Ltd. (Karachi, Pakistan). Chloroform (24216) was
obtained from Riedel-de Haën (Seelze, Germany). RNA was extracted
using TRIzol® reagent obtained from Invitrogen Life Technologies
(Carlsbad, CA, USA), and reverse transcription-polymerase chain
reaction (RT-PCR) was performed using Taq polymerase, dNTPs
and reverse transcriptase, all obtained from Thermo Fisher
Scientific, Inc. (Pittsburgh, PA, USA).
Mice
Male BALB/c mice (age, 3 months; weight, 35–40 g;
n=10 per group) were purchased from the National Institute of
Health (Islamabad, Pakistan). All experiments performed complied
with the guidelines outlined by the Institute of Laboratory Animal
Research, Division on Earth and Life Sciences, National Institute
of Health, USA (Guide for the Care and Use of Laboratory Animals:
Eighth Edition, 2011). The protocol for the present study was
approved by the Ethical Committee for Research on Animals Internal
Review Board at the Atta-ur-Rahman School of Applied Biosciences,
National University of Sciences and Technology (Islamabad,
Pakistan). The mice were maintained in a controlled environment
(22–25°C) within an animal house, with a natural day and night
cycle. The animals were administered tap water and fed a standard
diet. Mice were divided in three groups (n=10 per group), as
follows: Control, treated with normal saline intraperitoneally
(i.p.); AlCl3-treated, which received 150 mg/kg/day
AlCl3 (i.p.); and ibuprofen-treated, which received 100
mg/kg/day (i.p.).
Administration of agents
Ibuprofen (100 mg/kg/day) and AlCl3 (150
mg/kg/day) were administered for 12 consecutive days.
AlCl3 was dissolved in distilled water and injected
(i.p.) into the AlCl3-treated group mice (29). Normal saline (0.9%) was administered
to the control group according to body weight. Ibuprofen was
administered orally via feed-stuff to the ibuprofen-treated group
mice.
Morris water maze test
The procedure used was identical to the method
outlined by Ahmed and Gilani (30).
The experimental apparatus consisted of a circular tub containing
an invisible platform submerged 1.5 cm below the surface of water.
The temperature of the water was maintained at 20–23°C. External
cues, including pictures and objects, were present and the position
of the cues remained constant throughout the study.
The Morris water maze test was conducted for 7
consecutive days. Each mouse received two trials per day between
10:00 AM and 5:00 PM. The mice were randomly placed in one of four
quadrants of the tub, facing towards the wall of the tub, and were
allowed to swim until they reached the platform. The time taken to
reach the platform was recorded, and once a mouse reached the
platform it was allowed to rest for 15 sec. If a mouse failed to
locate the platform within 90 sec, it was placed on the platform
and allowed to rest for 15 sec.
Open field test
The open field test procedure was based on the
method outlined by Choleris et al (31), with minor alterations. Briefly, the
mice were placed into a square chamber and their activity was
recorded using a SteadyShot DSC-W610 camera (Sony Corporation,
Tokyo, Japan) fixed in a tripod stand at the top of the chamber.
The box was divided into central and peripheral areas. A minor
alteration of this method compared with Choleris et al was
that the peripheral area was ≤2 cm from the walls of the chamber.
The test duration was 30 min and the time spent by the animal in
the central and peripheral areas was recorded.
Gene expression analysis
The gene expression analysis was conducted according
to the procedure outlined by Ahmed et al (32), with minor alterations. The mice were
sacrificed under diethyl ether anesthesia (676845; Sigma-Aldrich),
after which the brains were removed in order to extract the cortex,
hippocampus and amygdala. RNA was extracted using TRIzol® reagent.
The quantity and quality of the RNA was determined using agarose
gel electrophoresis. RNA (1 µg in 40 µl reaction volume) was
reverse transcribed into cDNA, which was used as a template for PCR
with specific primers (Eurofins Scientific, Luxembourg), which are
listed in Table I. PCR cycling was
performed using a 2720 Thermal Cycler (Applied Biosystems, Inc.,
Waltham, MA, USA). cDNA (3 µl) was used for the PCR reactions in
the following 25-µl reaction mixture: 10 µM each primer, 25 µM
MgCl2, 10 µM dNTPs final concentration and 0.625 U Taq
Polymerase (Thermo Fisher Scientific, Waltham, MA, USA). PCR
cycling was conducted as follows: Initial denaturation at 95°C for
5 min, followed by denaturation at 94°C for 30 sec, annealing (at
temperatures given in Table I) for
30 sec and extension at 72°C for 30 sec for the number of cycles
given for each gene in Table I. This
was followed by a final extension step at 72°C for 10 min. The PCR
products were separated by 2% agarose gel electrophoresis, and
visualized with ethidium bromide. The intensity of each PCR product
band was quantified using ImageJ software (National Institutes of
Health, Bethesda, MA, USA), and β-actin was used as an internal
control.
 | Table I.List of primers and PCR
conditions |
Table I.
List of primers and PCR
conditions
| Primer no. | Gene | Primer sequence
(5′-3′) | Annealing
temperatures (°C) | Number of
cycles |
|---|
| 1 | Actin |
F=GCCTTCCTTCTTGGGTATGG | 55 | 32 |
|
|
|
R=CAGCTCAGTAACAGTCCGC |
|
|
| 2 | NP1 |
F=CAGGACACTCTAGGTGGAGG | 55 | 32 |
|
|
|
R=GAGGGGAGAAGAGAGACGAT |
|
|
| 3 | NP2 |
F=CATCAACGACAAGGTCGCAC | 55 | 32 |
|
|
|
R=CTCTTCACAGGTCTCCACAGG |
|
|
| 4 | NPR |
F=CAAGAGCAGGATACCTTGGG | 55 | 32 |
|
|
|
R=GAAGTGGGAGGTATAGCCAG |
|
|
| 5 | IL-1β |
F=TGAAGAAGAGCCCATCCTCTG | 55 | 32 |
|
|
|
R=GGGTGTGCCGTCTTTCATTA |
|
|
Statistical analysis
The data are presented as the mean ± standard error
of the mean. The results were analyzed by two way analysis of
variance and the Bonferroni post hoc test using GraphPad Prism®
software (GraphPad, Inc., La Jolla, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of ibuprofen on learning and
memory in an AlCl3-induced neurotoxicity mouse
model
In order to analyze the pharmacological effects of
ibuprofen on the learning and memory abilities of
AlCl3-treated mice, ibuprofen (100 mg/kg/day) and
AlCl3 (150 mg/kg/day) were administered to the
appropriate groups for 12 days, after which the mice were subjected
to memory testing using the Morris water maze test for the final 7
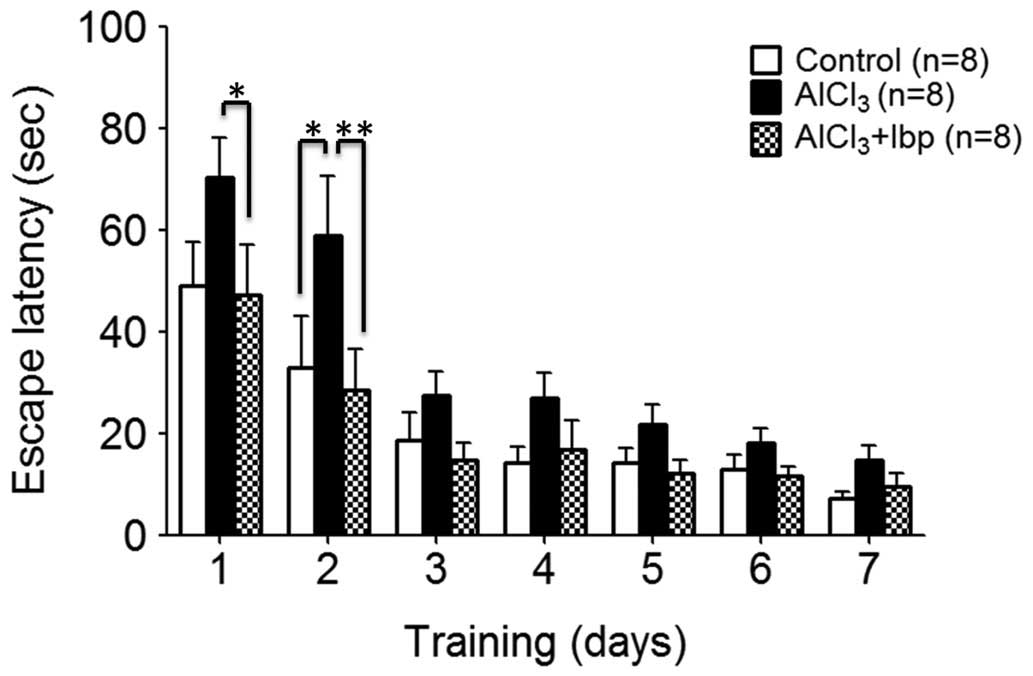
days of treatment, prior to sacrifice. On day two, the control
group demonstrated a significant decrease in escape latency time
(32.94±10.18), as compared with the AlCl3-treated group
(58.88±11.70; P<0.05; Fig.
1).
Treatment with ibuprofen significantly improved the
spatial reference memory of mice in the treated group (47.25±9.93)
on day one, as compared with the AlCl3-treated group
(70.38±7.75; P<0.05). On the second day, the ibuprofen-treated
group exhibited significantly improved learning and memory
abilities (28.63±7.94), as compared with the
AlCl3-treated group on the same day (58.88±11.7;
P<0.01; Fig. 1). However, on all
other days, no significant differences between the three groups
were observed (P>0.05).
Effects of ibuprofen on anxiety and
locomotion in AlCl3-induced neurotoxicity model
In order to investigate the therapeutic effects of
ibuprofen on anxiety and locomotion, open field tests were
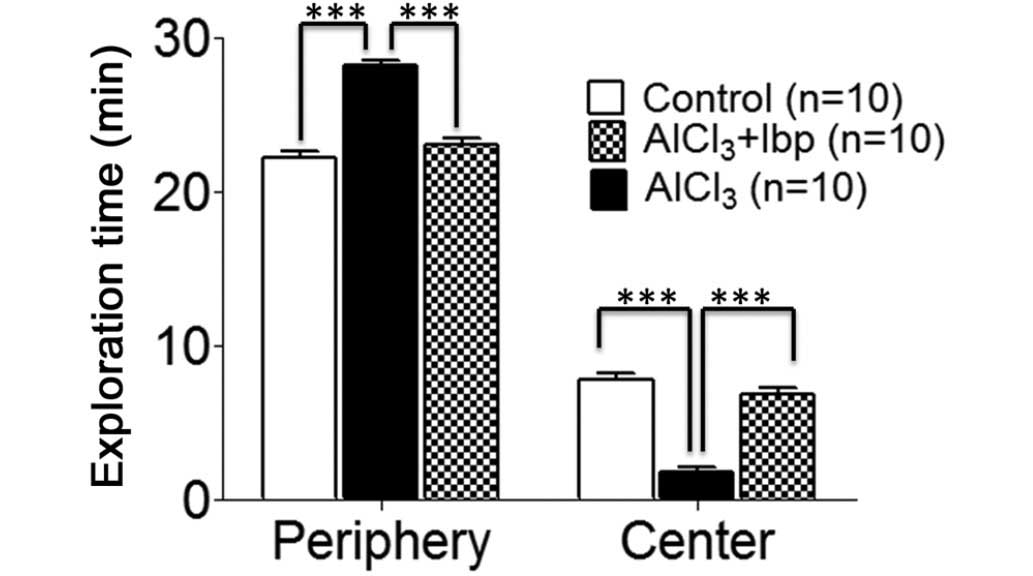
conducted on day 12 of treatment. The control group spent less time
(22.20±0.41 min) in the peripheral area of the chamber, as compared
with the AlCl3-treated group (28.2±0.29 min; Fig. 2), whereas the time spent by the
control group mice in the center (7.80±0.41 min) was greater than
the AlCl3-treated group (1.80±0.29 min); thus suggesting
that mice in the AlCl3-treated group had elevated levels
of anxiety (Fig. 2).
Following treatment with ibuprofen, a decrease in
the time spent in the periphery (23.10±0.34 min) was observed in
the ibuprofen-treated group, as compared with the
AlCl3-treated group (28.20±0.29 min; Fig. 2), whereas, the time spent in the
center (6.90±0.34 min) was significantly increased in the
ibuprofen-treated group, as compared with the
AlCl3-treated group (1.80±0.29 min; P<0.001; Fig. 2).
Effects of ibuprofen on the expression
levels of NPs in the hippocampus, cortex and amygdala
In order to evaluate the effects of ibuprofen on the
expression levels of NPs, the mice were sacrificed at the end of
treatment and RNA was extracted for analysis. Briefly, RNA was
reverse transcribed into cDNA, which was used as a template for
PCR. Ibuprofen treatment significantly decreased the expression
levels of NP1 (1.32±0.47) in the hippocampus of ibuprofen-treated
mice, as compared with the AlCl3-treated group
(8.62±1.54; P<0.001; Fig. 3A). No
significant decreases in the expression levels of NP2 and NPR were
observed among the three groups; however, a trend of decreasing
expression was detected in the ibuprofen-treated group, as compared
with the AlCl3-treated group (Fig. 3A).
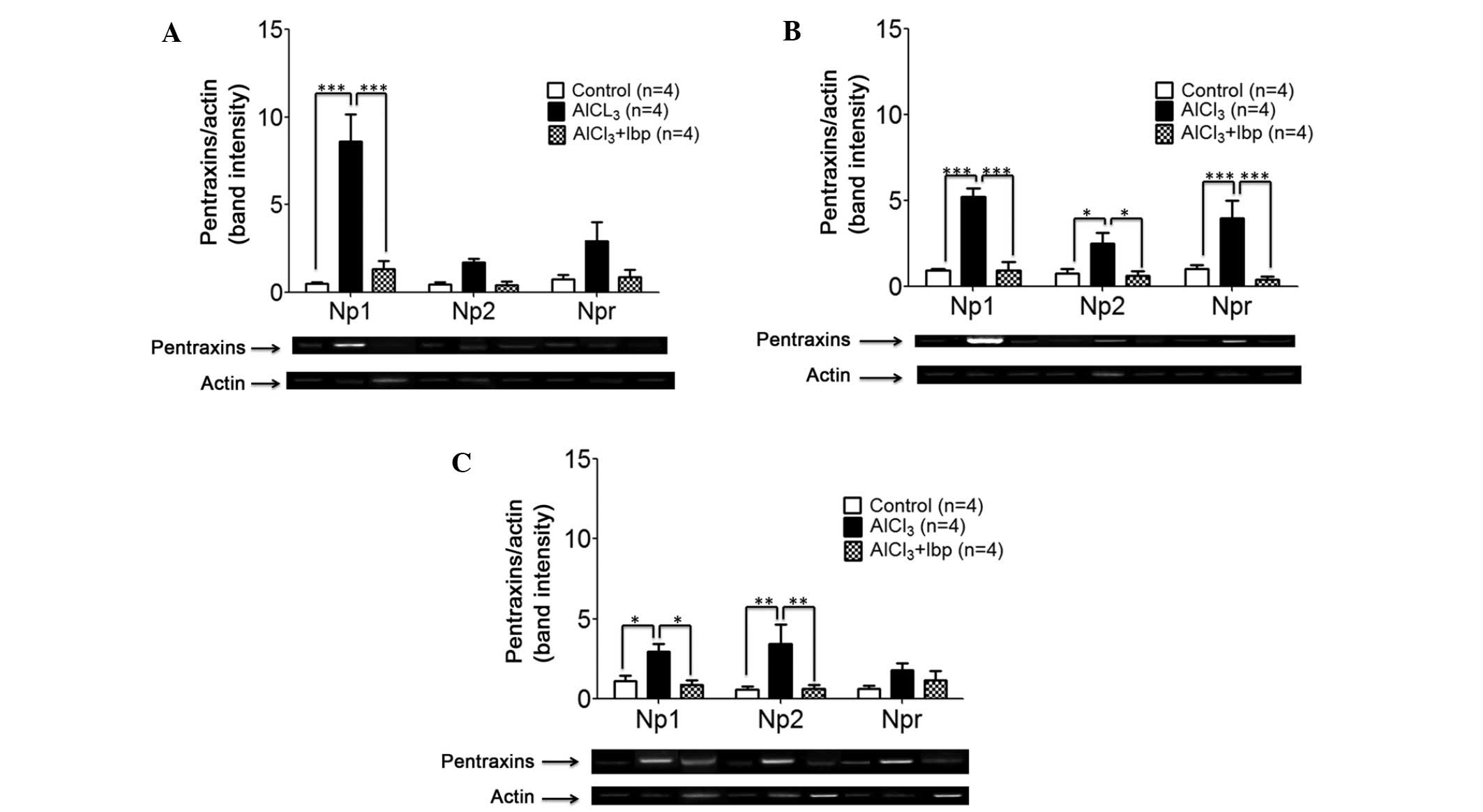 | Figure 3.Effects of ibuprofen treatment on the
expression levels of neuronal pentraxins in the various brain
structures of an AlCl3-induced mouse model of
neurotoxicity. (A) Ibuprofen treatment significantly decreased the
expression levels of NP1 in the hippocampus, as compared with the
AlCl3-treated group. (B) Ibuprofen significantly
decreased the expression levels of NP1, NP2 and NPR in the cortex,
as compared with the AlCl3-treated group. (C) Ibuprofen
significantly decreased the expression levels of NP1 and NP2 in the
amygdala, as compared with the AlCl3-treated group.
Reverse transcription-polymerase chain reaction and agarose gel
electrophoresis were used to analyze the expression levels of NP1,
NP2 and NPR in the various brain compartments. Data are presented
as the mean ± standard error of the mean (n=4). *P<0.05 vs. the
AlCl3-treated group; **P<0.01 vs. the
AlCl3-treated group; ***P<0.001 vs. the
AlCl3-treated group. NP1, neuronal pentraxin-1; NP2,
neuronal pentraxin-2; NPR, neuronal pentraxin receptor; Ibp,
ibuprofen; AlCl3, aluminum chloride. |
In the cortex, ibuprofen treatment was associated
with a significant decrease in the expression levels of NP1
(0.94±0.48; P<0.001), NP2 (0.62±0.28; P<0.05) and NPR
(0.43±0.14; P<0.001), as compared with the expression levels of
NP1 (5.21±0.51), NP2 (2.50±0.65) and NPR (4.00±1.01) in the
AlCl3-treated group (Fig.
3B).
In the amygdala, ibuprofen treatment was associated
with a significant decrease in the expression levels of NP1
(0.90±0.26; P<0.05) and NP2 (0.62±0.26; P<0.01), as compared
with the expression levels of NP1 (2.95±0.47) and NP2 (3.43±1.22)
in the AlCl3-treated group; however, no significant
difference in the expression levels of NPR were detected among the
groups (P>0.05; Fig. 3C).
Effects of ibuprofen on the expression
levels of IL-1β in the hippocampus, cortex and amygdala
In order to analyze the effects of ibuprofen on
inflammation in the AlCl3-treated group, IL-1β
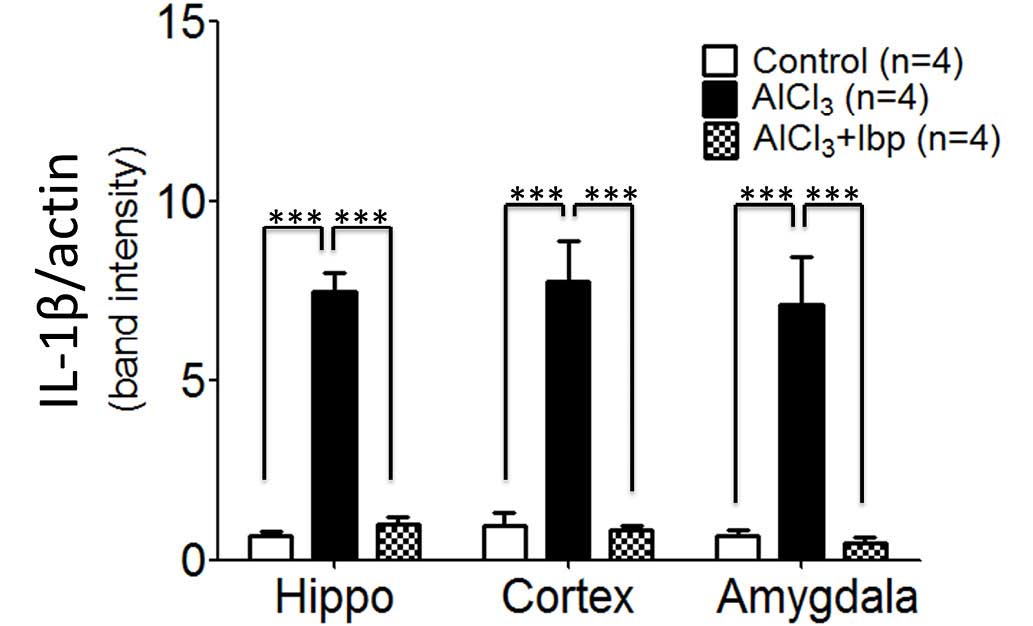
expression levels were analyzed. The control group exhibited
significantly reduced levels of IL-1β expression in the hippocampus
(0.69±0.10), cortex (0.98±0.33) and amygdala (0.67±0.18), as
compared with the levels observed in the hippocampus (7.47±0.53),
cortex (7.75±1.13) and amygdala (7.45±0.17) of the
AlCl3-treated group (P<0.001; Fig. 4). Furthermore, ibuprofen treatment
significantly decreased the expression levels of IL-1β in the
hippocampus (0.99±0.21), cortex (0.86±0.09) and amygdala
(0.49±0.13), as compared with the hippocampus, cortex and amygdala
of AlCl3-treated mice (P<0.001; Fig. 4).
Discussion
It has previously been suggested that NSAIDs exert
neuroprotective effects, in addition to their established
anti-inflammatory functions (33).
Ibuprofen is an NSAID that has been demonstrated to readily cross
the blood-brain barrier (34) and
exert memory-enhancing effects (29). The present study investigated the
neuroprotective effects of ibuprofen (100 mg/kg/day) in an
AlCl3-induced mouse model of neurotoxicity. In order to
assess hippocampus-dependent spatial and reference memory (35), a Morris water maze test was
conducted. The results of the present study demonstrated that
learning and memory were impaired in AlCl3-treated mice,
and that treatment with ibuprofen was able to reverse these
effects; thus suggesting that ibuprofen is able to exert
memory-enhancing effects. However, the effects were not significant
in the later training days.
In order to investigate the effects of ibuprofen on
the levels of anxiety, locomotion, and exploratory behavior in the
AlCl3-treated mice, an open field test was conducted in
the present study (36). Blanchard
et al (37) previously
suggested that the reluctance of an animal to move from one place
to another, or into the central area in the test, is indicative of
elevated levels of anxiety in the mice. The results of the present
study demonstrated an increase in the anxiety levels of
AlCl3-treated mice, as compared with the control group.
Treatment with ibuprofen reversed the effects of
AlCl3-treatment and was associated with a decrease in
the levels of anxiety in ibuprofen-treated mice. Anxiety has been
reported to be a predominant clinical symptom of neurodegenerative
disorders (38); therefore,
ibuprofen may have an additional therapeutic effect in the
treatment of neurodegenerative disorder-associated anxiety.
In order to investigate the underlying mechanisms of
ibuprofen, the effects of ibuprofen on the expression levels of NPs
and IL-1β in the AlCl3-treated mice were analyzed.
Previous studies have detected an association between the
accumulation of Aβ and the upregulation of NP1, which is an
apoptotic protein that promotes the death of damaged neurons
(26). The results of the present
study indicated that treatment with AlCl3 led to a
significant increase in the expression levels of NP1 in the
hippocampus; thus suggesting a possible role of NP1 in
AlCl3-induced neurotoxicity, which is consistent with a
previous report (26). The increase
in NP2 and NPR expression levels in the hippocampus was not
significant; however minor upregulation was observed in the
AlCl3-treated group, as compared with the control group.
Increased expression levels of NP1, NP2 and NPR were observed in
the cortex, whereas NP1 and NP2 expression levels were increased in
the amygdala, indicating that NPs have a role in
AlCl3-induced neurodegeneration, dementia and learning
and memory. Notably, treatment with ibuprofen was able to attenuate
memory impairment. Inflammation is a hallmark of AD pathogenesis
(39,40) and IL-1β, a pro-inflammatory cytokine,
has a role in inflammation (41). It
has previously been suggested that IL-1β overexpression is
associated with the formation of plaques (42). The results of the present study
demonstrated a significant increase in IL-1β expression levels in
the hippocampus, cortex, and amygdala of AlCl3-treated
mice, supporting previous data that IL-1β is elevated in a
degenerating brain (43).
Furthermore, treatment with ibuprofen was able to significantly
decrease the expression levels of IL-1β in the various brain
compartments, and the memory improvement observed may be due to
this, as it has previously been demonstrated that IL-1β has a
prominent role in memory impairment and aging (44). In addition, significant decreases in
the expression levels of NPs following treatment with ibuprofen
illustrated that ibuprofen may have a role in the expression of
NPs, as well as interleukins; thus suggesting that there may be a
possible link between NPs and neuroinflammation. NPs may promote
the upregulation of interleukins; however, the underlying mechanism
by which NPs are associated with inflammation remains unknown. The
present study identified novel ibuprofen targets and demonstrated
that treatment with ibuprofen may reduce the risk of developing
neurodegeneration. Therefore, ibuprofen may be considered a
potential therapeutic option for the treatment of patients with
neurodegenerative disorders, including AD.
In conclusion, the present study analyzed the
therapeutic effects of ibuprofen on the performance of mice in
various behavioral tests, and on the expression levels of NPs.
Ibuprofen increased learning and memory, and decreased anxiety in
mice. Furthermore, ibuprofen was able to reduce inflammation, which
is a hallmark of neurodegeneration and AD pathogenesis. Ibuprofen
also decreased the expression levels of NPs, which are associated
with neuropathology. Future studies should endeavor to elucidate
the mechanisms by which NPs are involved in inflammation and the
expression of IL-1β.
Acknowledgements
The present study was supported by The
Atta-ur-Rahman School of Applied Biosciences, National University
of Sciences and Technology, Islamabad, Pakistan.
References
|
1
|
Dokmeci D: Ibuprofen andA lzheimer's
disease. Folia Med (Plovdiv). 46:5–10. 2004.PubMed/NCBI
|
|
2
|
Bushra R and Aslam N: An overview of
clinical pharmacology of Ibuprofen. Oman Med J. 25:155–1661.
2010.PubMed/NCBI
|
|
3
|
Zaminelli T, Gradowski RW, Bassani TB,
Barbiero JK, Santiago RM, Maria-Ferreira D, Baggio CH and Vital MA:
Antidepressant and antioxidative effect of Ibuprofen in the
rotenone model of Parkinson's disease. Neurotox Res. 26:351–362.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zurita MP, Muñoz G, Sepúlveda FJ, Gómez P,
Castillo C, Burgos CF, Fuentealba J, Opazo C and Aguayo LG:
Ibuprofen inhibits the synaptic failure induced by the amyloid-β
peptide in hippocampal neurons. J Alzheimers Dis. 35:463–473.
2013.PubMed/NCBI
|
|
5
|
Cauli O, Rodrigo R, Piedrafita B, Llansola
M, Mansouri MT and Felipo V: Neuroinflammation contributes to
hypokinesia in rats with hepatic encephalopathy, Ibuprofen restores
its motor activity. J Neurosci Res. 87:1369–1374. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lim GP, Yang F, Chu T, Chen P, Beech W,
Teter B, Tran T, Ubeda O, Ashe KH, Frautschy SA and Cole GM:
Ibuprofen suppresses plaque pathology and inflammation in a mouse
model for Alzheimer's disease. J Neurosci. 20:5709–5714.
2000.PubMed/NCBI
|
|
7
|
Zawilla NH, Taha FM, Kishk NA, Farahat SA,
Farghaly M and Hussein M: Occupational exposure to aluminum and its
amyloidogenic link with cognitive functions. J Inorg Biochem.
139:57–64. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Khan KA, Kumar N, Nayak PG, Nampoothiri M,
Shenoy RR, Krishnadas N, Rao CM and Mudgal J: Impact of caffeic
acid on aluminium chloride-induced dementia in rats. J Pharm
Pharmacol. 65:1745–1752. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ahmed SS and Santosh W: Metallomic
profiling and linkage map analysis of early Parkinson's disease: A
new insight to aluminum marker for the possible diagnosis. PLoS
One. 5:e112522010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kawahara M and Kato-Negishi M: Link
between aluminum and the pathogenesis of Alzheimer's disease: The
integration of the aluminum and amyloid cascade hypotheses. Int J
Alzheimers Dis. 2011:2763932011.PubMed/NCBI
|
|
11
|
He BP and Strong MJ: Motor neuronal death
in sporadic amyotrophic lateral sclerosis (ALS) is not apoptotic.
Histopathology. Neuropathol Appl Neurobiol. 26:150–160. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pan R, Qiu S, Lu DX and Dong J: Curcumin
improves learning and memory ability and its neuroprotective
mechanism in mice. Chin Med J (Engl). 121:832–839. 2008.PubMed/NCBI
|
|
13
|
Shen LX, Jin LQ, Zhang DS and Xue GP:
Effect of osthol on memory impairment of mice in AlCl3-induced
acute senile model. Yao Xue Xue Bao. 37:178–180. 2002.PubMed/NCBI
|
|
14
|
Zaky A, Mohammad B, Moftah M, Kandeel KM
and Bassiouny AR: Apurinic/apyrimidinic endonuclease 1 is a key
modulator of aluminum-induced neuroinflammation. BMC Neurosci.
14:262013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Veerhuis R: Histological and direct
evidence for the role of complement in the neuroinflammation of AD.
Curr Alzheimer Res. 8:34–58. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Moran LB, Hickey L, Michael GJ, Derkacs M,
Christian LM, Kalaitzakis ME, Pearce RK and Graeber MB: Neuronal
pentraxin II is highly upregulated in Parkinson's disease and a
novel component of Lewy bodies. Acta Neuropathol. 115:471–478.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bjartmar L, Huberman AD, Ullian EM,
Rentería RC, Liu X, Xu W, Prezioso J, Susman MW, Stellwagen D,
Stokes CC, et al: Neuronal pentraxins mediate synaptic refinement
in the developing visual system. J Neurosci. 26:6269–6281. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schlimgen AK, Helms JA, Vogel H and Perin
MS: Neuronal pentraxin, a secreted protein with homology to acute
phase proteins of the immune system. Neuron. 14:519–526. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Goodman AR, Cardozo T, Abagyan R, Altmeyer
A, Wisniewski HG and Vilcek J: Long pentraxins, An emerging group
of proteins with diverse functions. Cytokine Growth Factor Rev.
7:191–202. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tsui CC, Copeland NG, Gilbert DJ, Jenkins
NA, Barnes C and Worley PF: Narp, a novel member of the pentraxin
family, promotes neurite outgrowth and is dynamically regulated by
neuronal activity. J Neurosci. 16:2463–2478. 1996.PubMed/NCBI
|
|
21
|
Kamenskaya MA and Thesleff S: The
neuromuscular blocking action of an isolated toxin from the elapid
(Oxyuranus scutellactus). Acta Physiol Scand. 90:716–724.
1974. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu D, Hopf C, Reddy R, Cho RW, Guo L,
Lanahan A, Petralia RS, Wenthold RJ, O'Brien RJ and Worley P: Narp
and NP1 form heterocomplexes that function in developmental and
activity-dependent synaptic plasticity. Neuron. 39:513–528. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dowton SB and McGrew SD: Rat serum amyloid
P component. Histopathology. Biochem J. 270:553–556. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Whitehead AS, Zahedi K, Rits M, Mortensen
RF and Lelias JM: Mouse C-reactive protein. Histopathology. Biochem
J. 266:283–290. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Breviario F, d'Aniello EM, Golay J, Peri
G, Bottazzi B, Bairoch A, Saccone S, Marzella R, Predazzi V, Rocchi
M, et al: Interleukin-1-inducible genes in endothelial cells.
Cloning of a new gene related to C-reactive protein and serum
amyloid P component. J Biol Chem. 267:22190–22197. 1992.PubMed/NCBI
|
|
26
|
Abad MA, Enguita M, DeGregorio-Rocasolano
N, Ferrer I and Trullas R: Neuronal pentraxin 1 contributes to the
neuronal damage evoked by amyloid-beta and is overexpressed in
dystrophic neurites in Alzheimer's brain. J Neurosci.
26:12735–12747. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Walsh DM and Selkoe DJ: Deciphering the
molecular basis of memory failure in Alzheimer's disease. Neuron.
44:181–193. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
DeGregorio-Rocasolano N, Gasull T and
Trullas R: Overexpression of neuronal pentraxin 1 is involved in
neuronal death evoked by low K(+) in cerebellar granule
cells. J Biol Chem. 276:796–803. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hashmi AN, Yaqinuddin A and Ahmed T:
Pharmacological effects of Ibuprofen on learning and memory,
muscarinic receptors gene expression and APP isoforms level in
pre-frontal cortex of AlCl3-induced toxicity mouse model. Int J
Neurosci. 125:277–287. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ahmed T and Gilani AH: Inhibitory effect
of curcuminoids on acetylcholinesterase activity and attenuation of
scopolamine-induced amnesia may explain medicinal use of turmeric
in Alzheimer's disease. Pharmacol Biochem Behav. 91:554–559. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Choleris E, Thomas AW, Kavaliers M and
Prato FS: A detailed ethological analysis of the mouse open field
test, Effects of diazepam, chlordiazepoxide and an extremely low
frequency pulsed magnetic field. Neurosci Biobehav Rev. 25:235–260.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ahmed T, Enam SA and Gilani AH:
Curcuminoids enhance memory in an amyloid-infused rat model of
Alzheimer's disease. Neuroscience. 169:1296–1306. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Asanuma M, Miyazaki I and Ogawa N:
Neuroprotective effects of nonsteroidal anti-inflammatory drugs on
neurodegenerative diseases. Curr Pharm Des. 10:695–700. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Parepally JM, Mandula H and Smith QR:
Brain uptake of nonsteroidal anti-inflammatory drugs, Ibuprofen,
flurbiprofen, and indomethacin. Pharm Res. 23:873–881. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bromley-Brits K, Deng Y and Song W: Morris
water maze test for learning and memory deficits in Alzheimer's
disease model mice. J Vis Exp. (53)2920:2011.
|
|
36
|
Walsh RN and Cummins RA: The Open-Field
Test: A critical review. Psychol Bull. 83:482–504. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Blanchard DC, Griebel G and Blanchard RJ:
Mouse defensive behaviors, Pharmacological and behavioral assays
for anxiety and panic. Neurosci Biobehav Rev. 25:205–218. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ferretti L, McCurry SM, Logsdon R, Gibbons
L and Teri L: Anxiety and Alzheimer's disease. J Geriatr Psychiatry
Neurol. 14:52–58. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Akiyama H, Barger S, Barnum S, Bradt B,
Bauer J, Cole GM, Cooper NR, Eikelenboom P, Emmerling M, Fiebich
BL, et al: Inflammation and Alzheimer's disease. Neurobiol Aging.
21:383–421. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tuppo EE and Arias HR: The role of
inflammation in Alzheimer's disease. Int J Biochem Cell Biol.
37:289–305. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ren K and Torres R: Role of
interleukin-1beta during pain and inflammation. Brain Res Rev.
60:57–64. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mrak RE and Griffin WS: Interleukin-1,
neuroinflammation and Alzheimer's disease. Neurobiol Aging.
22:903–908. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cacabelos R, Alvarez XA, Fernández-Novoa
L, Franco A, Mangues R, Pellicer A and Nishimura T: Brain
interleukin-1 beta in Alzheimer's disease and vascular dementia.
Methods Find Exp Clin Pharmacol. 16:141–151. 1994.PubMed/NCBI
|
|
44
|
Ahmed T and Gilani AH: A comparative study
of curcuminoids to measure their effect on inflammatory and
apoptotic gene expression in an Aβ plus ibotenic acid-infused rat
model of Alzheimer's disease. Brain Res. 1400:1–18. 2011.
View Article : Google Scholar : PubMed/NCBI
|