Introduction
The worldwide incidence of obesity associated with
metabolic syndrome continues to escalate, despite the increased
awareness and global efforts to understand its pathogenesis and
develop treatments (1). Dysregulated
energy homeostasis may result from a reduction in physical
activity, increased energy-dense food availability and
overconsumption, combined with genetic, social and economic
complicating factors (2). Among the
critical determinants for the development of obesity may be an
increase in the regional distribution of body fat (i.e., abdominal
obesity). Abdominal obesity may present with a number of
atherogenic risk factors, including hypertension, dyslipidemia,
alterations in coagulation and inflammatory cytokine profiles, and
hyperinsulinemic insulin resistance (3). As an expected consequence, there has
been an increase in morbidity and mortality due to cardiovascular
disease (4).
Prior studies have been conducted to elucidate the
association between increased adiposity and insulin resistance.
Adipokines (2) including leptin and
adiponectin, which are secreted by adipocytes, may modulate the
sensitivity of insulin, whose action activates multiple signaling
events following phosphorylation of insulin receptor, among other
molecules, in patients with type 2 diabetes (5–7).
A number of oral anti-diabetic medicines including
thiazolidinediones and metformin are currently used for improving
insulin resistance (8). However,
currently available medications for metabolic syndrome exhibit
various adverse effects and high rates of secondary failure
(9). Thus, there has been a growing
interest and focus on complementary and alternative approaches to
metabolic syndrome (10).
Conjugated linoleic acid (CLA) refers to a group of
isomers of linoleic acid (cis-9, cis-12 octadecadienoic acid). CLA
has been reported to possess certain anti-carcinogenic (11), anti-atherogenic (12) and immunomodulatory (13) activities, in addition to exhibiting
anti-obesity and anti-diabetic effects (14,15). In
addition, CLA treatment during adipocyte differentiation reduces
lipid accumulation and inhibits the expression of peroxisome
proliferator-activated receptor gamma, a nuclear receptor that
activates genes involved in lipid storage and metabolism (16,17). A
CLA dosage of 750 mg/kg has been shown to confer anti-obesity and
anti-diabetic effects on ob/ob obese mice (18).
The potato tuber (Solanum tuberosum) is the
source of potato proteinase inhibitor (PPI) II, which elicits a
satiety response (19) and delays
gastric emptying in humans (20).
While several methods to isolate and purify PPI II have been
developed for use in a laboratory setting, they are all laborious
and expensive (21–23). Considering the low yield and
complexity of PPI II isolation, crude PPI concentrate containing a
number of thermostable proteinase inhibitors, including PPI II, has
previously been developed (24).
This crude concentrate exhibited satiety-promoting activity in
vivo and enhanced the cholecystokinin (CCK) release in a
similar manner to PPI II (25).
Slendesta® Potato Extract 5% Powder (SLD) is a
standardized potato extract that contains 5% PPI II. The present
study was conducted to determine the anti-obesity and anti-diabetic
effects of SLD on ob/ob mice, which are
leptin-deficient and which is an established mouse model of
obesity. The highest dosage of SLD used was 300 mg/kg. In previous
clinical and in vivo studies (25,26),
which employed ~15 mg/kg PPI II, satiety response and anti-obesity
effects were observed.
The present study investigated the anti-obesity and
anti-diabetic effects of SLD, with the aim of determining its
potential as a complementary or alternative agent for the
management of metabolic syndrome. SLD dosages of 50, 150 and 300
mg/kg were administered orally once a day for 28 days to
ob/ob mice. Body weight, food consumption, organ
weight, serum chemistry and cytokines associated with obesity and
diabetes were measured.
Materials and methods
Materials
SLD, a light beige-colored powder, was provided by
Kemin Food L.C. (Des Moines, IA, USA). SLD was composed of 78.9%
carbohydrates, 15.3% protein and ~20 mg/g Na. CLA capsules
containing a light pink solution were purchased from RexGene
Biotech Co., Ltd., (Chungwon-gun, South Korea). SLD was stored at
4°C in a refrigerator, and CLA capsules were stored in a desiccator
to protect them from light and humidity until use. SLD and CLA were
dissolved or suspended in distilled water on the day of first
administration.
Animals
Five normoglycemic wild-type C57BL/6JJms mice
(5-weeks-old; Japan SLC, Inc., Shizuoka, Japan), and 25 male
genetically obese mice (C57BL/6JHam-ob/ob 5-weeks-old; SLC)
were used following acclimatization for 28 days. The mice (n=5 per
polycarbonate cage) were housed in a room controlled for
temperature (20–25°C), humidity (40–45%) and alternating 12-h
light/dark cycles. Standard rodent chow (Samyang Foods, Wonju,
Korea) and water were supplied ad libitum. SLD (50, 150 or
300 mg/kg) or CLA (750 mg/kg) solutions were administered by
gastric gavage once a day for 28 days. Equal volumes of distilled
water were orally administered in the wild-type and ob/ob
control mice. All mice were sacrificed on Day 28
post-administration by exsanguination following intramuscular
injection of Zoletil 50 (0.05 ml/kg; Virbac Korea Co. Ltd., Seoul,
Korea). The present study was approved by the Institutional Animal
Care and Use Committee of Daegu Hanny University (IRB approval no.
DHU2011-016; Gyeongsan, Republic of Korea).
Body weight
Body weight was measured once a day for 28 days,
from one day prior to the initiation of administration (Day −1)
using an automatic electronic balance (Precisa Gravimetrics AG,
Dietikon, Switzerland). At the initiation of administration (Day 0)
and at sacrifice (Day 28), all experimental animals were fasted
overnight (~12 h, with no restriction of water intake) to reduce
the differences from feeding.
Mean daily food consumption
An equal quantity of food (150 g) was supplied to
each cage, and the remaining food was weighed after 24 h using the
aforementioned automatic electronic balance. These measurements
were conducted on Days 1, 7, 14, 21, 25 and 27. The difference
between the supplied and remaining diet represented was the
individual mean daily food consumption of mice (g/day/mouse).
Organ weight
Following sacrifice, the weights of the pancreas and
epididymal fat pad were measured. To reduce the differences from
individual body weights, relative weights (%) were calculated using
the body weight at sacrifice.
Blood glucose levels
At sacrifice, blood was collected from the caudal
vena cava under anesthesia. The collected blood was deposited into
a sodium fluoride glucose vacuum tube (BD Biosciences, Franklin
Lakes, NJ, USA), and plasma was separated by centrifugation at
1,100 × g for 10 min at room temperature. Blood glucose levels were
detected using an automated blood analyzer (TBA-200FR; Toshiba,
Tokyo, Japan).
Plasma insulin levels
To detect plasma insulin levels, blood was collected
at sacrifice via the caudal vena cava under anesthesia, and the
collected blood was deposited in heparin pretreated vacuum tubes
(BD Biosciences) for plasma separation. Plasma insulin levels were
assayed with enzyme-linked immunosorbent assay kit (ELISA;
Boehringer Mannheim GmbH, Mannheim, Germany) as previously detailed
(27).
Serum biochemical parameters
At sacrifice, ~1 ml blood was collected from the
caudal vena cava under anesthesia. All blood samples were
centrifuged at 21,130 × g for 10 min at 4°C using a clotting
activated serum tube (Vacutte®; Greiner Bio-One GmbH,
Kremsmünster, Austria). Next, levels of serum total cholesterol
(TC), free fatty acid (FFA) and triglyceride were detected using an
Olympus AU400 automated blood analyzer (Olympus Corporation, Tokyo,
Japan).
Serum leptin and adiponectin
levels
To quantify serum levels of leptin and adiponectin,
serum was separated from the collected blood using a standard
method and ELISAs were performed as previously described (28). Briefly, serum leptin levels were
determined using a Mouse Leptin ELISA (cat. no. EZML-82K; EMD
Millipore, Billerica, MA, USA) and serum adiponectin levels were
detected using a Mouse/Rat adiponectin ELISA kit (Otsuka
Pharmaceutical Co., Ltd., Tokyo, Japan), according to the
manufacturer's protocols.
Histopathology
After measuring the organ weight, the splenic lobes
of pancreas, epididymal fat pad and dorsal abdominal fat pad
attached on the muscularis quadratus lumborum were sampled. Sampled
tissues were fixed in 10% neutral buffered formalin (Sigma-Aldrich,
St. Louis, MO, USA). After paraffin embedding (Leica Microsystems
GmbH, Wetzlar, Germany), 3–4 µm serial sections were prepared.
Representative sections were stained with hematoxylin and eosin
(H&E; Sigma-Aldrich) for light microscopy (Eclipse 80i; Nikon
Corporation, Tokyo, Japan) examination. The histological profiles
of individual organs were observed.
Histomorphometry
Based on the previous histomorphometrical analysis
of metabolic mice (29) or diabetic
rats (27,30), adipocyte deposition and hypertrophy,
percentages of pancreatic zymogen granule occupied regions,
pancreatic islet numbers, and occupied percentages were determined.
The mean diameters of epididymal and dorsal abdominal white
adipocytes were calculated as an indicator of adipocyte hypertrophy
in restricted view fields on a computer monitor using an automated
image analysis system (iSolution FL ver. 9.1; IMT iSolution Inc.,
Quebec, Canada). The mean diameter was expressed in µm and a
minimum of ten white adipocytes per each fat pad was measured.
Thickness of dorsal abdominal fat pad (mm) was measured using an
automated image analysis process. Mean area occupied by zymogen
granules were calculated as percentages between one field of liver
(%/mm2 of pancreatic parenchyma) using an automated
image analysis process. Mean number of pancreatic islets were
counted using an automated image analysis process located in
appropriate regions of pancreas (n/10 mm2 of pancreatic
parenchyma). Mean areas occupied by the pancreatic islets was
calculated as percentages between the appropriate pancreas field
(%/10 mm2 of pancreatic parenchyma) using an automated
image analysis process. The samples were analyzed by the same
histopathologist who was blinded to sample identity.
Immunohistochemistry
Additional prepared serial sectioned tissues were
immunostained using an immunohistochemical avidin-biotin-peroxidase
kit (Vectastain Elite ABC kit; cat. no. PK-6200; Vector
Laboratories, Inc., Burlingame, CA, USA), according to a previously
described method (31). Briefly, the
tissue sections were incubated with guinea pig anti-insulin (1:100;
cat. no. ab7842; Abcam, Cambridge, UK) or rabbit anti-glucagon
(1:50; cat. no. ab8055; Abcam) polyclonal antibodies. Based on
previous histomorphometrical analyses of endocrine cells of normal
ddN mice (32) and diabetic rats
(27,30), cells displaying >10%
immunoreactive density of insulin and glucagon were regarded as
positive immunoreactive (IR). The number of insulin- and
glucagon-IR cells and the ratio of insulin-IR to glucagon-IR cells
were determined. Mean numbers of insulin-IR and glucagon-IR cells
were counted in restricted view fields (mm2 of
pancreatic parenchyma) on a computer monitor using an automated
image analysis process. The aforementioned ratio was determined
based on these values. The samples were analyzed by the same
histopathologist who was blinded to sample identity.
Statistical analysis
All values are expressed as the mean ± standard
deviation. Statistical analyses were conducted using SPSS for
Windows, version 14.0K (SPSS, Inc., Chicago, IL, USA). Multiple
comparison tests for different dose groups were conducted. Variance
homogeneity was assessed using the Levene test. If the Levene test
indicated no significant deviations from variance homogeneity, the
obtain data were analyzed by one-way analysis of variance test
followed by the least-significant differences (LSD)
multi-comparison test. In cases where significant deviations from
variance homogeneity were observed with the Levene test, the
Kruskal-Wallis H test was conducted. When a significant difference
was observed in the Kruskal-Wallis H test, the Mann-Whitney
U-Wilcoxon Rank Sum W test was conducted to determine the specific
pairs of group comparison. The criterion for statistical
significance was set at either P<0.05 or P<0.0.1.
Results
Body weight change
Prior to the initiation of administration of the
study products, all ob/ob mice displayed significantly
(P<0.01) increased body weight compared with the wild-type mice.
Significantly decreased body weight was detected in mice treated
with CLA and SLD (150 and 300 mg/kg dose) from days 20, 22 and 23
compared with ob/ob control. Furthermore, the body weight
gain during the 28-day administration period was decreased in these
treatment groups compared with ob/ob control, and this
reduction was significant in the 150 and 300 mg/kg SLD groups
(Table I).
 | Table I.Body weight detected during 28 days
of oral treatment of SLD and CLA in ob/ob mice. |
Table I.
Body weight detected during 28 days
of oral treatment of SLD and CLA in ob/ob mice.
|
|
|
|
| SLD groups
(mg/kg) |
|---|
|
|
|
|
|
|
|---|
| Time point | Control | ob/ob
control | CLA | 50 | 150 | 300 |
|---|
| Day −1 |
24.40±1.03 |
42.94±4.71a |
42.58±2.91 |
42.74±4.00 |
42.96±2.28 |
42.60±4.36 |
| Day 0 |
21.22±0.94 |
39.60±4.50a |
39.90±2.73 |
40.10±4.63 |
39.50±2.56 |
39.70±4.65 |
| Day 7 |
25.38±1.06 |
46.30±4.16a |
43.42±2.98 |
44.16±3.13 |
43.70±2.98 |
41.88±4.33 |
| Day 14 |
25.44±1.34 |
46.88±4.07a |
41.56±2.93 |
44.82±1.37 |
43.24±3.30 |
41.14±4.45 |
| Day 20 |
26.20±1.16 |
50.22±4.52a |
42.72±2.94b |
48.50±1.51 |
43.82±2.73 |
42.40±4.53 |
| Day 21 |
25.66±1.20 |
49.84±4.78a |
42.08±2.47b |
48.98±1.00 |
43.46±2.69 |
42.06±4.59 |
| Day 22 |
26.04±1.25 |
50.74±5.01a |
43.12±3.27b |
50.06±0.92 |
44.16±2.44 |
41.88±5.17b |
| Day 23 |
25.86±1.26 |
50.26±4.63a |
42.42±2.90b |
49.58±1.47 |
43.58±2.64b |
41.52±5.16b |
| Day 28 |
23.82±1.24 |
48.46±4.03a |
40.08±2.75c |
47.96±1.38 |
41.82±2.29b |
39.50±5.23c |
|
Gaind |
2.60±0.40 |
8.50±1.48a |
0.18±1.74c |
7.86±4.52 |
2.32±0.72c |
−0.20±2.31c |
Mean daily food consumption
Significant (P<0.01) increases of mean daily food
consumption were detected in all ob/ob mice compared with
wild-type control. However, mice treated with CLA and 150 or 300
mg/kg SLD exhibited significant (P<0.01) decreases in mean daily
food consumption from Day 14 compared to the ob/ob control
(Table II).
| Table II.Mean daily food consumption during 27
days of oral treatment with SLD or CLA in ob/ob
mice. |
Table II.
Mean daily food consumption during 27
days of oral treatment with SLD or CLA in ob/ob
mice.
|
| Food consumption
(g/day/mouse) |
|---|
|
|
|
|---|
| Group | Day 1 | Day 7 | Day 14 | Day 21 | Day 25 | Day 27 |
|---|
| Controls |
|
|
Intact |
5.12±0.59 |
4.68±0.48 |
4.62±0.54 |
4.84±0.54 |
4.74±0.69 |
4.60±0.53 |
|
ob/ob |
7.36±0.60a |
6.84±0.40a |
6.94±0.50a |
7.18±0.60a |
6.88±0.47a |
6.66±0.46a |
|
CLA |
7.32±0.41a |
6.62±0.52a |
5.20±1.04b |
5.64±0.53b |
4.46±0.23b |
4.32±0.97b |
| SLD (mg/kg) |
|
| 50 |
7.14±0.26a |
6.86±0.29a |
6.78±0.76a |
7.18±0.91a |
6.74±0.55a |
6.48±0.28a |
|
150 |
7.06±0.59a |
6.50±0.55a |
5.74±0.47b,c |
5.80±0.73b,c |
5.54±0.38b,c |
5.16±0.11b |
|
300 |
7.06±0.47a |
6.42±0.44a |
5.62±0.35b,c |
5.18±0.84b |
4.36±0.79b |
4.00±0.70b |
Epididymal fat and pancreatic weight
change
Significant (P<0.01) increases in epididymal fat
pad weight were detected in ob/ob control mice compared with
the wild-type control at sacrifice. However, these increases were
significantly (P<0.01) decreased by treatment of CLA and 150 or
300 mg/kg SLD. Mice treated with 50 mg/kg SLD showed similar
epididymal fat pad weights as compared with ob/ob control
mice (Table III). Significant
(P<0.01) decreases in relative pancreas weight were detected in
all ob/ob mice regardless of treatment, compared with the
wild-type control. However, no meaningful changes in absolute
pancreas weight were evident in the CLA and SLD treated-mice
compared with the ob/ob control mice (Table III).
| Table III.Organ weight in ob/ob
mice orally administered SLD or CLA. |
Table III.
Organ weight in ob/ob
mice orally administered SLD or CLA.
|
| Absolute organ
weight (g) | Relative organ
weight (% of body weight) |
|---|
|
|
|
|
|---|
| Group | Epididymal
fats | Pancreas | Epididymal
fats | Pancreas |
|---|
| Controls |
|
|
|
|
|
Intact |
0.078±0.024 |
0.118±0.014 |
0.323±0.079 |
0.494±0.054 |
|
ob/ob |
1.145±0.287a |
0.138±0.042 |
2.353±0.477a |
0.287±0.095a |
|
CLA |
0.634±0.121a,b |
0.113±0.028 |
1.573±0.205a,b |
0.280±0.063a |
| SLD (mg/kg) |
|
|
|
|
| 50 |
1.083±0.176a |
0.139±0.036 |
2.260±0.376a |
0.289±0.070a |
|
150 |
0.637±0.117a,b |
0.132±0.024 |
1.520±0.248a,b |
0.314±0.042a |
|
300 |
0.545±0.105a,b |
0.126±0.011 |
1.391±0.265a,b |
0.324±0.059a |
Blood glucose
Significantly (P<0.01) increased blood glucose
levels were detected in ob/ob control mice compared to the
wild-type control. However, the blood glucose levels were
significantly (P<0.01) reduced by the treatment with CLA and SLD
(150 and 300 mg/kg) compared to the ob/ob control. Mice
treated with 50 mg/kg SLD displayed similar blood glucose levels as
the ob/ob control (Table
IV).
| Table IV.Serum biochemical parameters and
adiponectin levels in ob/ob mice orally administered
SLD or CLA. |
Table IV.
Serum biochemical parameters and
adiponectin levels in ob/ob mice orally administered
SLD or CLA.
| Group | Glucose
(mg/dl) | TC (mg/dl) | FFA (mmol/l) | Triglyceride
(mg/dl) | Adiponectin
(µg/ml) | Leptin (ng/ml) | Insulin (pM) |
|---|
| Controls |
|
|
Intact |
138.20±10.06 |
106.20±6.61 |
0.62±0.08 |
87.60±12.84 |
22.06±3.37 |
8.64±1.22 |
368.20±40.14 |
|
ob/ob |
598.00±48.98a |
289.00±18.22a |
1.77±0.17a |
131.40±8.44a |
8.98±1.46b |
1.30±0.17a |
5,043.00±350.75b |
|
CLA |
484.80±43.75a,c |
247.60±8.99a,c |
1.42±0.09a,c |
106.80±10.33a,c |
8.70±0.33b |
1.36±0.35a |
4,000.60±217.20b |
| SLD (mg/kg) |
|
| 50 |
602.40±67.01a |
280.60±14.01a |
1.73±0.16a |
128.60±6.66a |
9.39±1.03b |
1.23±0.32a |
5,168.00±265.60b |
|
150 |
428.20±43.42a,c |
247.80±10.18a,c |
1.35±0.11a,c |
101.00±9.96c,d |
12.22±1.44b,e |
1.26±0.31a |
3,651.80±836.52b,f |
|
300 |
383.00±54.30a,c |
232.20±21.02a,c |
1.18±0.15a,c |
94.60±9.13c |
13.53±1.12b,f |
1.26±0.17a |
3,082.60±122.49b,f |
Plasma insulin levels
Significantly (P<0.01) increased plasma insulin
levels were detected in ob/ob control compared with the
wild-type control. However, these increases were significantly
(P<0.01) inhibited by treatment with SLD (150 and 300 mg/kg)
compared to the ob/ob control. In mice treated with 50 mg/kg
SLD, the plasma insulin levels were similar to the levels in the
ob/ob control mice (Table
IV).
Serum TC, FFA and triglyceride
levels
Significant (P<0.01) increases of serum TC levels
were detected in ob/ob control compared to the wild-type
control. However, serum TC levels were significantly (P<0.01)
decreased in mice treated with CLA and 150 or 300 mg/kg SLD
compared to the ob/ob control (Table IV). Significant (P<0.01)
increases in serum FFA and triglyceride levels were detected in the
ob/ob control compared with the wild-type control. However,
these increases in serum FFA and triglyceride levels were
significantly (P<0.01) decreased by treatment with CLA and 150
or 300 mg/kg SLD compared with the ob/ob control (Table IV).
Serum leptin and adiponectin
levels
Significant (P<0.01) decreases of serum leptin
levels were detected in ob/ob control compared to the
wild-type control. However, similar serum leptin levels were
detected in mice treated with the three doses of SLD and CLA
compared with the ob/ob control (data not shown).
Significant (P<0.01) decreases of the serum adiponectin levels
were detected in ob/ob control mice compared to the
wild-type control. However, the serum adiponectin levels were
significantly (P<0.01) increased in mice treated with 150 and
300 mg/kg SLD compared to the ob/ob control. In mice treated
with CLA and 50 mg/kg SLD, the serum adiponectin levels were
similar to the levels in ob/ob control mice (Table IV).
Adipocyte diameters and the deposited
abdominal fat pad thickness
Significant (P<0.01) increases of epididymal and
abdominal adipocyte diameters were detected in the ob/ob
control compared to the wild-type control. However, diameters of
both adipocyte types were significantly (P<0.01) decreased in
mice treated with CLA and SLD (150 and 300 mg/kg) compared to the
ob/ob control. Mice treated with 50 mg/kg SLD showed a
similar trend compared to the ob/ob control (Table V and Fig.
1). Significant (P<0.01) increases of the deposited
abdominal fat pad thickness were detected in the ob/ob
control mice compared to the wild-type control. However, the
abdominal fat pad thickness was significantly (P<0.01) decreased
in mice with CLA and SLD (150 and 300 mg/kg) compared to the
ob/ob control. In mice treated with 50 mg/kg SLD, a similar
trend as the ob/ob control mice was observed (Table V and Fig.
1).
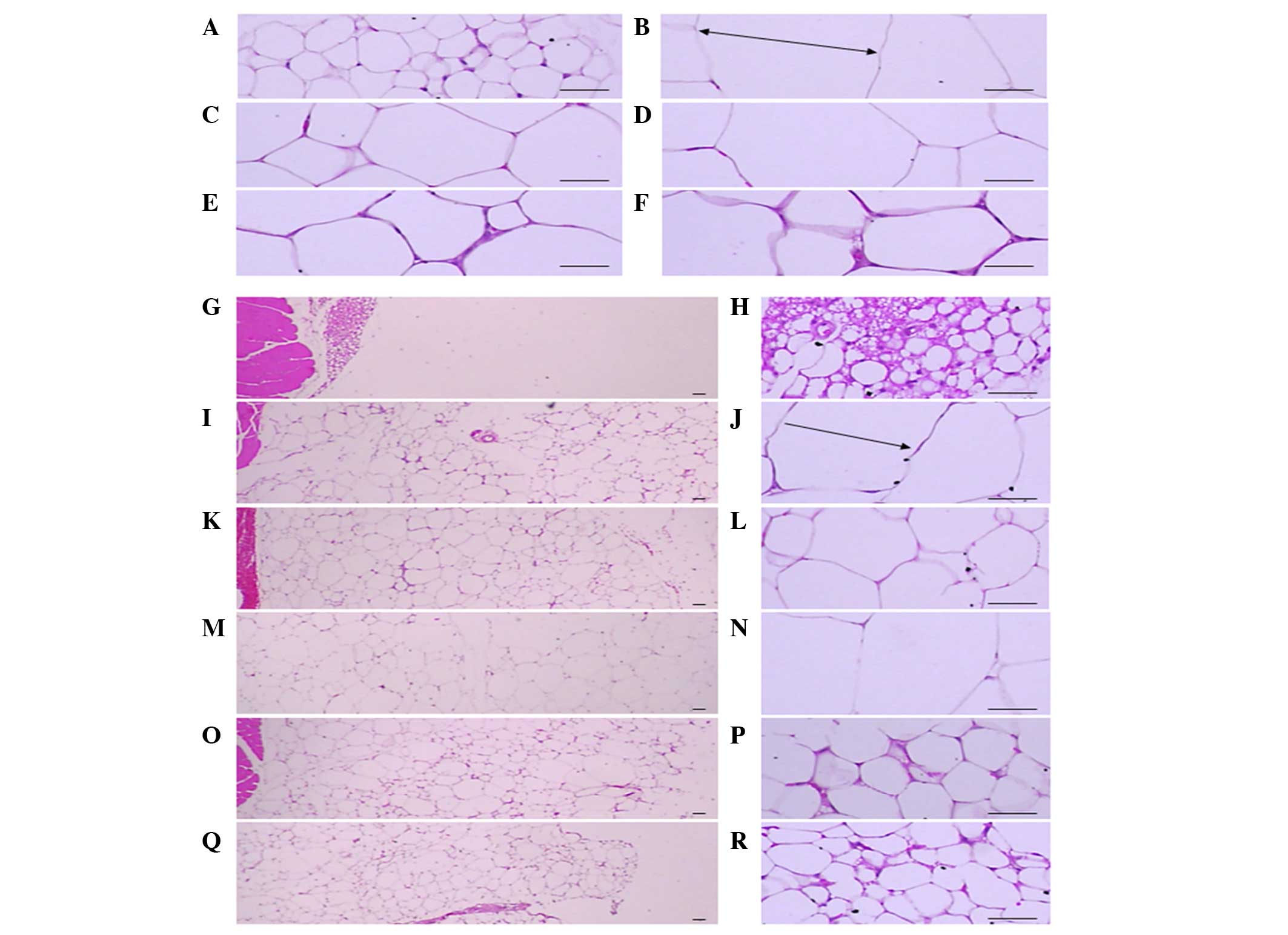 | Figure 1.Histological profiles of the
epididymal fat pad and abdominal fat pad (magnification, ×100 in
the left panels and ×400 in the right panels). Note that
significant increases of epididymal and abdominal adipocytes were
detected in the ob/ob control compared to wild-type
control. However, diameters of both adipocyte types were
significantly (P<0.01) decreased in treated with CLA and SLD
(150 and 300 mg/kg) compared to ob/ob control. (A, G
and H) Wild-type and (B, I and J) ob/ob control, (C,
K and L) CLA, (D, M and N) 50, (E, O and P) 150 and (F, Q and R)
300 mg/kg SLD-treated ob/ob mice (stain, hematoxylin
and eosin; scale bars, 80 µm). Arrows indicates the adipocyte
diameter measured. SLD, Slendesta™; CLA, conjugated linoleic acid
(750 mg/kg) treated group. |
| Table V.Adipocytes in ob/ob
mice orally administered SLD or CLA. |
Table V.
Adipocytes in ob/ob
mice orally administered SLD or CLA.
| Group | Epididymal
adipocytes diameter (µm) | Abdominal adipocyte
diameter (µm) | Abdominal fat pad
thickness (mm; mean fat pads from muscle) |
|---|
| Controls |
|
|
|
|
Intact |
88.59±13.03 |
46.27±8.53 |
1.25±0.28 |
|
ob/ob |
322.31±27.88a |
215.89±14.24a |
6.03±0.45a |
|
CLA |
218.25±37.98a,b |
168.30±23.87a,b |
4.73±0.59a,b |
| SLD (mg/kg) |
|
|
|
| 50 |
298.87±18.55a |
217.65±20.14a |
6.13±0.16a |
|
150 |
215.34±37.79a,b |
145.27±15.34a,b |
4.77±0.49a,b |
|
300 |
173.65±33.84a,b |
101.73±23.92a,b |
3.71±0.41a,b |
Exocrine pancreas zymogen granule
contents
Significant (P<0.01) decreases in the exocrine
pancreas zymogen granule contents (the percentages of exocrine
pancreas occupied by zymogen granules) were detected in
ob/ob control mice compared to the wild-type control mice.
However, the exocrine pancreas zymogen granule contents were
significantly (P<0.01 or P<0.05) increased in mice treated
with CLA and SLD compared to the ob/ob control, except for
mice treated with 50 mg/kg SLD (Table
VI).
| Table VI.Histomorphology of pancreas in
ob/ob mice orally administered SLD or CLA. |
Table VI.
Histomorphology of pancreas in
ob/ob mice orally administered SLD or CLA.
| Group | Zymogen granules
(%/mm2) | Pancreatic islets
(n/10 mm2) | Islet occupied
regions (%/10 mm2) |
|---|
| Controls |
|
|
|
|
Intact |
66.63±8.47 |
7.60±1.82 |
3.58±1.46 |
|
ob/ob |
31.90±5.85a |
27.20±1.92a |
24.34±4.55a |
|
CLA |
42.40±2.53a,b |
19.60±2.07a,b |
17.79±2.10a,b |
| SLD (mg/kg) |
|
|
|
| 50 |
32.56±5.94a |
26.40±3.36a |
23.00±2.65a |
|
150 |
42.19±4.46a,b |
22.40±1.67a,b |
18.09±2.07a,b |
|
300 |
59.67±3.90b |
19.20±1.92b |
13.31±1.45b |
Pancreatic islet hyperplasia and
expansions
Significant (P<0.01) increases of pancreatic
islet numbers and percentages of islet occupied regions were
detected in the ob/ob control compared to the wild-type
control, resulting from marked hyperplasia of the pancreatic islet
itself or component endocrine cells. However, these events were
significantly (P<0.01) reduced by treatment with CLA and SLD
(150 and 300 mg/kg), compared to the ob/ob control. Mice
treated with 50 mg/kg SLD displayed similar pancreatic islet
numbers and percentages of islet occupied regions compared with
ob/ob control (Table VI and
Fig. 2).
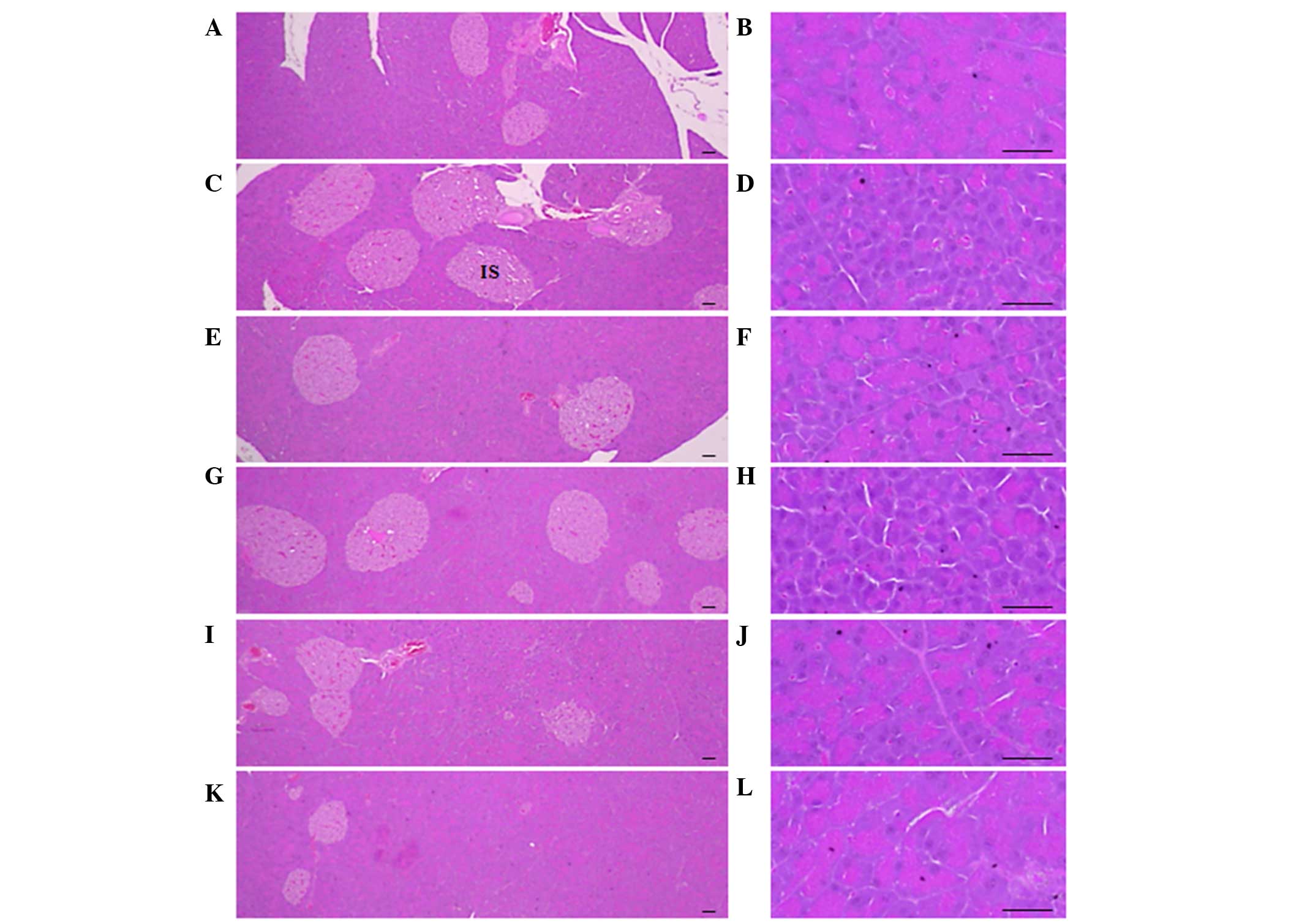 | Figure 2.Histological profiles of the pancreas
(magnification, ×100 in the left panels and ×400 in the right
panels). Marked decreases in exocrine pancreas zymogen granule
contents (the percentages of exocrine pancreas occupied by zymogen
granules) were detected in the ob/ob control compared
to wild-type control. However, the exocrine pancreas zymogen
granule contents were increased in mice treated with CLA (750
mg/kg) and SLD (150 or 300 mg/kg), compared to the
ob/ob control. In addition, increases in pancreatic
islet numbers and percentages of islet occupied regions were
detected in the ob/ob control compared to wild-type
control. However, these events were reduced by treatment with CLA
(750 mg/kg) and SLD (150 and 300 mg/kg) compared to the
ob/ob control. (A and B) Wild-type mice, (C and D)
ob/ob control mice, (E and F) CLA-treated
ob/ob mice and (G and H) 50, (I and J) 150 and (K and
L) 300 mg/kg SLD-treated ob/ob mice (stain,
hematoxylin and eosin; scale bars, 80 µm). SLD, Slendesta™; CLA,
conjugated linoleic acid; IS, pancreatic islets. |
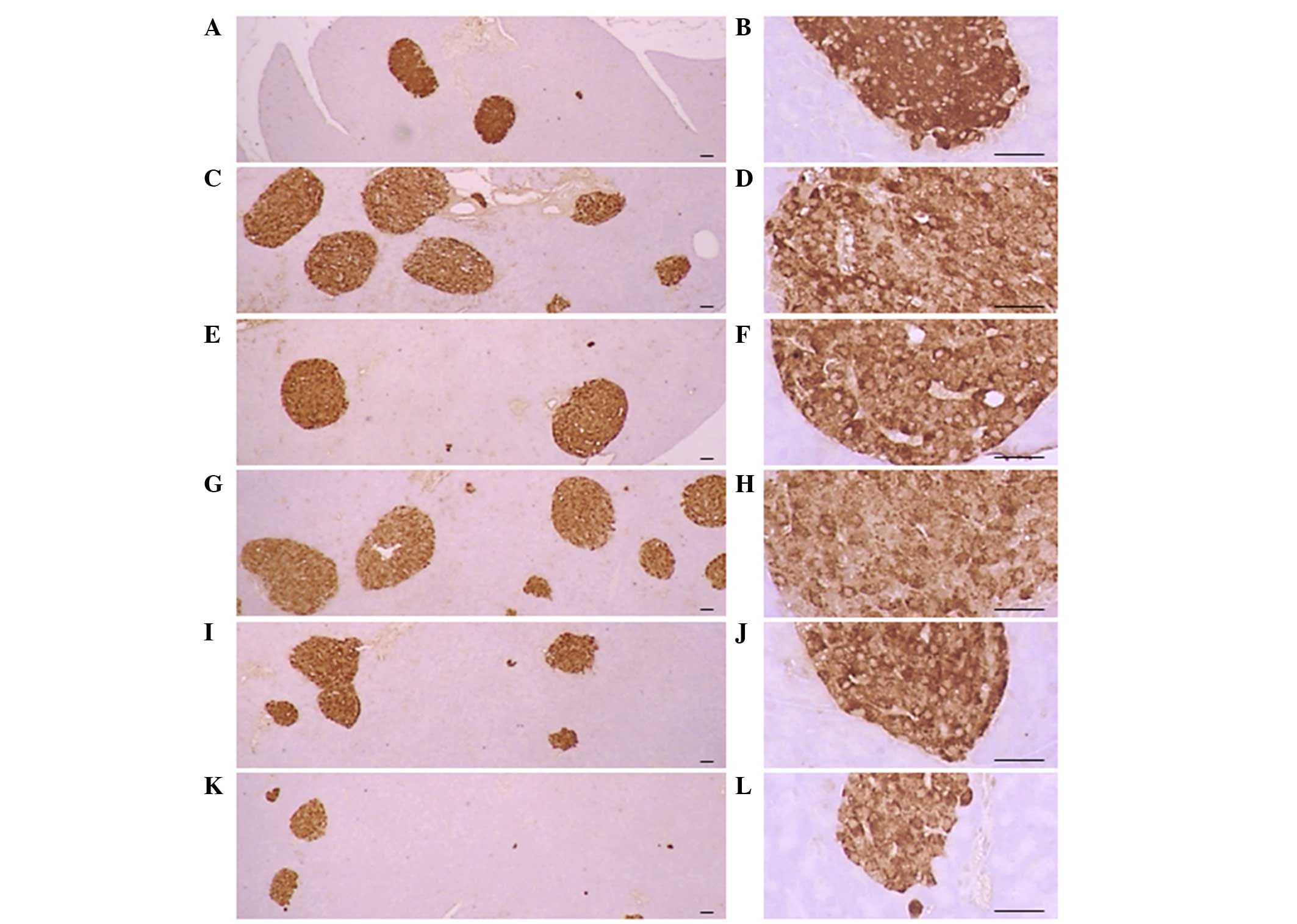
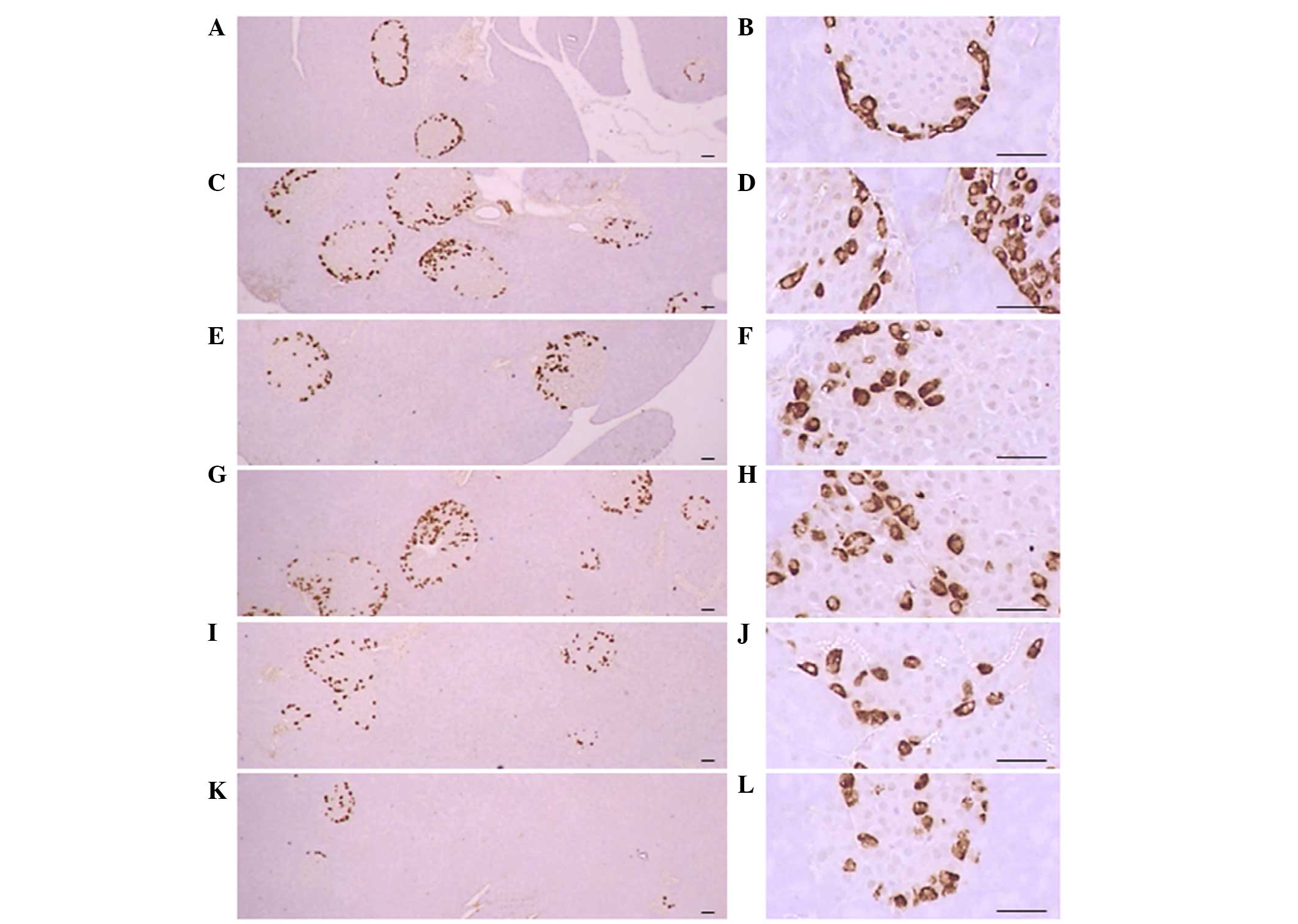
Pancreatic islet insulin-IR and
glucagon-IR cells
Significant (P<0.01) increases of insulin-IR and
glucagon-IR cells were detected in the ob/ob control
compared to the wild-type control, resulting from marked
hyperplasia of endocrine cells. However, these increases were
significantly (P<0.01) normalized by treatment of CLA and SLD
compared to the ob/ob control mice, except for mice treated
with 50 mg/kg SLD. No meaningful changes in insulin/glucagon-IR
cells were demonstrated in all ob/ob mice compared to the
wild-type control due to the simultaneous increases of the both
insulin-IR and glucagon-IR cells, regardless of treatment (Table VII and Figs. 3 and 4).
| Table VII.Insulin- and glucagon-producing cells
in ob/ob mice orally administered SLD or CLA. |
Table VII.
Insulin- and glucagon-producing cells
in ob/ob mice orally administered SLD or CLA.
| Group | Insulin-producing
cells (cells/mm2) | Glucagon-producing
cells (cells/mm2) |
Insulin/glucagon-producing cell ratio |
|---|
| Controls |
|
|
|
|
Intact |
610.40±95.00 |
247.40±59.33 |
2.51±0.30 |
|
ob/ob |
1,931.60±253.53a |
826.00±111.88a |
2.35±0.29 |
|
CLA |
1,242.40±133.11a,b |
522.80±65.76a,b |
2.39±0.29 |
| SLD (mg/kg) |
|
|
|
| 50 |
2,046.00±304.41a |
829.40±133.74a |
2.49±0.29 |
|
150 |
1,533.00±169.27a,b |
628.80±84.59a,c |
2.45±0.24 |
|
300 |
975.80±85.90b |
414.60±34.05a,b |
2.36±0.20 |
Discussion
Obesity has been a major public health concern for
decades, particularly in developed countries. Obesity is an
expensive burden on society, increasing medical care costs and
affecting the available labor pool (33). At the individual level, obesity is a
growing health issue; obesity-related health problems include
hyperlipidemia, diabetes mellitus and cardiovascular diseases
(34). Efforts to reduce obesity
have included research that has increased understanding of the
association between increased adiposity and insulin resistance
(2,7). Metabolic syndrome refers to the
simultaneous combination of obesity and related diseases, including
diabetes, hypertension, hypercholesterolemia and
hypertriglyceridemia (35).
Therefore, a complex disease animal model is required in order to
discover or develop novel drugs and/or alternative/complementary
therapies against metabolic syndrome.
In the present study, to understand the anti-obese
and anti-diabetic effects of SLD in genetically induced
obese-ob/ob mice, SLD was orally administered once a
day for 28 days. The effects were compared to untreated
ob/ob and normoglycemic wild-type mice. CLA was used
as a reference drug, which affects obesity and diabetes.
Increased mean daily food consumption was detected
in the ob/ob mice at each time point compared with
the wild-type mice. CLA presently decreased the daily food
consumption when compared to the ob/ob mice, and
showed anti-diabetic and anti-obesity effects, which is consistent
with a previous study (18). Similar
results were observed with SLD at 150 and 300 mg/kg dose, and lead
by the end of the study to a normalization of food consumption to a
level similar to that observed in wild-type mice. It is possible
that SLD operates by a similar mechanism of action as CLA. SLD is a
standardized potato extract containing 5% PPI II, which elicits a
satiety response (19) and delays
gastric emptying in humans (20).
This is peculated to occur via the increased release of CCK, one of
the most extensively studied peptides involved in the regulation of
food intake (25). However, the
possibility that other mechanisms may be involved cannot be
entirely excluded, as SLD also influences serum adiponectin levels,
while CLA does not affect serum adipokine. Further investigations
are therefore warranted.
In addition, an increased accumulation of adipose
tissues is a common feature in obesity, and adipocyte hypertrophy
has been observed in a previous histological examination (36). Adipose tissues are considered to be
an organ for energy storage, and also an endocrine and secretory
organ (37). Adipose tissues secrete
adipokines and changes in the expression, secretion and activity of
these adipokines as a result of obesity may be implicated in the
development of various diseases including insulin resistance
(2,37). Among these, leptin was initially
investigated as a satiety signal regulating food intake and energy
expenditure and was found to effectively reduce appetite and feed
intake (38). Therefore,
deficiencies in leptin signaling or functioning in the hypothalamus
are speculated to contribute to the development of obesity
(38). Adiponectin (also known as
Acrp30) is a novel adipokine that has been recently identified
(39). It is exclusively expressed
in adipose tissue (40) and is
abundantly released into the circulating blood (41). More recently, obesity-related
decreases in plasma adiponectin levels have been reported in humans
(42) and experimental animals
(43). Furthermore, it has been
shown that hypoadiponectinemia is closely related to insulin
resistance (2). In the present
study, 150 and 300 mg/kg SLD directly inhibited the deposition of
adipose tissues and the hypertrophy of adipocytes, while improving
serum adiponectin abnormal changes in ob/ob mice,
similar to CLA. These results are indicate that the favorable
effects on diabetes-related obesity may be induced by SLD
concentrations as low as 150 mg/kg. However, neither CLA nor SLD
had an influence on the serum leptin levels in ob/ob
mice, which are deficient in leptin.
Obesity leads to the development of pancreatic
steatosis, acinar cell atrophy and a diminution in the number of
zymogen granules (44,45). The increase of zymogen granules in
exocrine pancreatic acinar cells indicates the production of
digestive enzymes, particularly for lipid and protein digestion
(46). In the present study, the
decline in pancreatic zymogen granules was histopathologically
detected in ob/ob control mice compared to wild-type
control mice, which indicated a large release of pancreatic juice
to remove the lipids. Consequently, the contents of pancreatic
zymogen granules were markedly decreased. However, this decrease of
zymogen granules in the exocrine pancreas was markedly inhibited by
treatment with 150 and 300 mg/kg SLD, which was similar to the
effects of CLA. Therefore, the anti-obese effects of SLD and CLA
appear to mediate the inhibition of lipid digestion by decreasing
the pancreatic enzyme production. PPI II is active in eliciting a
satiety response (19) and delayed
gastric emptying in humans (41),
elevating the circulation of cholecystokinin (CCK) (25). We propose that the anti-obesity
effects of SLD may include the inhibition of lipid and protein
digestion by decreasing the release of pancreatic enzymes. However,
further investigation is warranted to elucidate a mechanism
underlying this phenomenon.
Hyperglycemia is the primary symptom of diabetes,
and it must be controlled to effectively treat diabetes (47). All ob/ob mice used in
the present study also showed hyperglycemia. Hyperglycemia was
markedly and significantly decreased by treatment with 150 and 300
mg/kg SLD, but not 50 mg/kg SLD, providing direct evidence that
>50 mg/kg SLD is necessary for a potent anti-diabetic effect.
Increased insulin secretion is in part related to pancreatic islet
hyperplasia in the progression of insulin-resistance in type-2
diabetes (48). Pancreatic islet
insulin-producing cells are increased in area and in number,
leading to increased insulin secretion to maintain glucose
homeostasis. Glucagon-producing cells also increase in similar
proportions to the increase in insulin cells (48). In the present study, a marked
increase in islet-occupied regions and insulin- and
glucagon-producing cells were observed in ob/ob
control mice with normal insulin/glucagon cell ratios. However,
treatment with CLA and SLD (150 and 300 mg/kg) brought the
endocrine pancreas secretion of insulin in the ob/ob
mice to normal levels. These results suggest that SLD exerts a
protective effect against insulin resistance. Progression of
chronic diabetes in ob/ob mice is associated with the
development of hyperlipemia (18).
The critical problem in hyperlipemia is elevated levels of serum
FFA, triglyceride and TC (49).
Similar favorable hypolipidemic, anti-obesity and anti-diabetic
effects were observed in mice treated with 150 mg/kg SLD, compared
with 750 mg/kg CLA in the present study.
The presents results suggest that 28 days of
continuous oral treatment of SLD effectively improved or normalized
the parameters associated with obesity and diabetes and their
related complications in ob/ob mice. Thus, SLD may be
a promising alternative therapeutic agent for the treatment of
metabolic syndrome, mediated by an already known mechanism, through
the food intake regulatory peptide CCK. However, further
investigations are required to identify the specific mechanism by
which the observed effects of SLD are mediated.
Acknowledgements
This study was supported by the National Research
Foundation of Korea grant funded by the Korea government (no.
2011-0030124). The authors would like to thank Ms. Samanta Maci
(Kemin) for providing assistance in reviewing the manuscript.
Glossary
Abbreviations
Abbreviations:
|
SLD
|
Slendesta
|
|
CLA
|
conjugated linoleic acid
|
|
ob/ob
|
C57BLKS/J-ob/ob
|
|
TC
|
total cholesterol
|
|
FFA
|
free fatty acid
|
|
PPI
|
potato proteinase inhibitor
|
|
CCK
|
cholecystokinin
|
|
H&E
|
hematoxylin and eosin
|
|
IR
|
immunoreactive
|
|
LSD
|
least-significant differences
|
|
MW test
|
Mann-Whitney U-Wilcoxon Rank Sum W
test
|
References
|
1
|
Yun JW: Possible anti-obesity therapeutics
from nature-a review. Phytochemistry. 71:1625–1641. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mitchell M, Armstrong DT, Robker RL and
Norman RJ: Adipokines: Implications for female fertility and
obesity. Reproduction. 130:583–597. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kunitomi M, Wada J, Takahashi K,
Tsuchiyama Y, Mimura Y, Hida K, Miyatake N, Fujii M, Kira S,
Shikata K and Maknio H: Relationship between reduced serum IGF-I
levels and accumulation of visceral fat in Japanese men. Int J Obes
Relat Metab Disord. 26:361–369. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hida K, Wada J, Eguchi J, Zhang H, Baba M,
Seida A, Hashimoto I, Okada T, Yasuhara A, Nakatsuka A, et al:
Visceral adipose tissue-derived serine protease inhibitor: A unique
insulin-sensitizing adipocytokine in obesity. Proc Natl Acad Sci
USA. 102:10610–10615. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wolf G, Chen S, Han DC and Ziyadeh FN:
Leptin and renal disease. Am J Kidney Dis. 39:1–11. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yamauchi T, Kamon J, Waki H, Imai Y,
Shimozawa N, Hioki K, Uchida S, Ito Y, Takakuwa K, Matsui J, et al:
Globular adiponectin protected ob/ob mice from diabetes and
ApoE-deficient mice from atherosclerosis. J Biol Chem.
278:2461–2468. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sakaue H, Nishizawa A, Ogawa W,
Teshigawara K, Mori T, Takashima Y, Noda T and Kasuga M:
Requirement for 3-phosphoinositide-kependent dinase-1 (PDK-1) in
insulin-induced glucose uptake in immortalized brown adipocytes. J
Biol Chem. 278:38870–38874. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Seufert J, Lübben G, Dietrich K and Bates
PC: A comparison of the effects of thiazolidinediones and metformin
on metabolic control in patients with type 2 diabetes mellitus.
Clin Ther. 26:805–818. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Inzucchi SE: Oral antihyperglycemic
therapy for type 2 diabetes: Scientific review. JAMA. 287:360–372.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sicińska P, Pytel E, Maćczak A and
Koter-Michalak M: The use of various diet supplements in metabolic
syndrome. Postepy Hig Med Dosw (Online). 69:25–33. 2015.(In
Polish). View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Parodi PW: Cows' milk fat components as
potential anticarcinogenic agents. J Nutr. 127:1055–1060.
1997.PubMed/NCBI
|
|
12
|
Lee KN, Kritchevsky D and Pariza MW:
Conjugated linoleic acid and atherosclerosis in rabbits.
Atherosclerosis. 108:19–25. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Parker J, Daniel LW and Waite M: Evidence
of protein kinase C involvement in phorbol diester-stimulated
arachidonic acid release and prostaglandin synthesis. J Biol Chem.
262:5385–5393. 1987.PubMed/NCBI
|
|
14
|
Banni S, Carta G, Angioni E, Murru E,
Scanu P, Melis MP, Bauman DE, Fischer SM and Ip C: Distribution of
conjugated linoleic acid and metabolites in different lipid
fractions in the rat liver. J Lipid Res. 42:1056–1061.
2001.PubMed/NCBI
|
|
15
|
Pariza MW, Park Y and Cook M: The
biologically active isomers of conjugated linoleic acid. Prog Lipid
Res. 40:283–298. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bocher V, Pineda-Torra I, Fruchart JC and
Staels B: PPARs: Transcription factors controlling lipid and
lipoprotein metabolism. Ann N Y Acad Sci. 967:7–18. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Granlund L, Pedersen JI and Nebb HI:
Impaired lipid accumulation by trans10, cis12 CLA during adipocyte
differentiation is dependent on timing and length of treatment.
Biochim Biophys Acta. 1687:11–22. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hue JJ, Lee KN, Jeong JH, Lee SH, Lee YH,
Jeong SW, Nam SY, Yun YW and Lee BJ: Anti-obesity activity of
diglyceride containing conjugated linoleic acid in C57BL/6J ob/ob
mice. J Vet Sci. 10:189–195. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hill AJ, Peikin SR, Ryan CA and Blundell
JE: Oral administration of proteinase inhibitor II from potatoes
reduces energy intake in man. Physiol Behav. 48:241–246. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schwartz JG, Guan D, Green GM and Phillips
WT: Treatment with an oral proteinase inhibitor slows gastric
emptying and acutely reduces glucose and insulin levels after a
liquid meal in type II diabetic patients. Diabetes Care.
17:255–262. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Levitt J: The isolation and preliminary
fractionation of proteins from dormant and growing potato tubers.
Plant Physiol. 26:59–65. 1951. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bryant J, Green TR, Gurusaddaiah T and
Ryan CA: Proteinase inhibitor II from potatoes: Isolation and
characterization of its protomer components. Biochemistry.
15:3418–3424. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim MH, Park SC, Kim JY, Lee SY, Lim HT,
Cheong H, Hahm KS and Park Y: Purification and characterization of
a heat-stable serine protease inhibitor from the tubers of new
potato variety ‘Golden Valley’. Biochem Biophys Res Commun.
346:681–686. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vlachojannis JE, Cameron M and Chrubasik
S: Medicinal use of potato-derived products: A systematic review.
Phytother Res. 24:159–162. 2010.PubMed/NCBI
|
|
25
|
Komarnytsky S, Cook A and Raskin I: Potato
protease inhibitors inhibit food intake and increase circulating
cholecystokinin levels by a trypsin-dependent mechanism. Int J Obes
(Lond). 35:236–243. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Peters HP, Foltz M, Kovacs EM, Mela DJ,
Schuring EA and Wiseman SA: The effect of protease inhibitors
derived from potato formulated in a minidrink on appetite, food
intake and plasma cholecystokinin levels in humans. Int J Obes
(Lond). 35:244–250. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim JD, Kang SM, Park MY, Jung TY, Choi HY
and Ku SK: Ameliorative anti-diabetic activity of dangnyosoko, a
Chinese herbal medicine, in diabetic rats. Biosci Biotechnol
Biochem. 71:1527–1534. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sahai A, Malladi P, Pan X, Paul R,
Melin-Aldana H, Green RM and Whitington PF: Obese and diabetic
db/db mice develop marked liver fibrosis in a model of nonalcoholic
steatohepatitis: Role of short-form leptin receptors and
osteopontin. Am J Physiol Gastrointest Liver Physiol.
287:G1035–G1043. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jung YM, Lee SH, Lee DS, You MJ, Chung IK,
Cheon WH, Kwon YS, Lee YJ and Ku SK: Fermented garlic protects
diabetic, obese mice when fed a high-fat diet by antioxidant
effects. Nutr Res. 31:387–396. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim JD, Kang SM, Seo BI, Choi HY, Choi HS
and Ku SK: Anti-diabetic activity of SMK001, a poly herbal formula
in streptozotocin induced diabetic rats: Therapeutic study. Biol
Pharm Bull. 29:477–482. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Edwin N and Leigh CM: Immunocytochemical
identification of islet cells containing calcitonin gene-related
peptide-like immunoreactivity in the plains rat pancreas
(Pseudomys australis). Singapore Med J. 40:528–530.
1999.PubMed/NCBI
|
|
32
|
Lee HS, Chang JH and Ku SK: An
immunohistochemical study of the pancreatic endocrine cells of the
ddN mouse. Folia Histochem Cytobiol. 48:387–393. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ng M, Fleming T, Robinson M, Thomson B,
Graetz N, Margono C, Mullany EC, Biryukov S, Abbafati C, Abera SF,
et al: Global, regional, and national prevalence of overweight and
obesity in children and adults during 1980–2013: A systematic
analysis for the Global Burden of Disease Study 2013. Lancet.
384:766–781. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liese AD, Mayer-Davis EJ and Haffner SM:
Development of the multiple metabolic syndrome: An epidemiologic
perspective. Epidemiol Rev. 20:157–172. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Deen D: Metabolic syndrome: Time for
action. Am Fam Physician. 69:2875–2882. 2004.PubMed/NCBI
|
|
36
|
Morange PE, Lijnen HR, Alessi MC, Kopp F,
Collen D and Juhan-Vague I: Influence of PAI-1 on adipose tissue
growth and metabolic parameters in a murine model of diet-induced
obesity. Arterioscler ThrombVasc Biol. 20:1150–1154. 2000.
View Article : Google Scholar
|
|
37
|
Fujita H, Fujishima H, Koshimura J, Hosoba
M, Yoshioka N, Shimotomai T, Morii T, Narita T, Kakei M and Ito S:
Effects of antidiabetic treatment with metformin and insulin on
serum and adipose tissue adiponectin levels in db/db mice. Endocr
J. 52:427–433. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bjørbaek C and Kahn BB: Leptin signaling
in the central nervous system and the periphery. Recent Prog Horm
Res. 59:305–331. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Scherer PE, Williams S, Fogliano M,
Baldini G and Lodish HF: A novel serum protein similar to C1q,
produced exclusively in adipocytes. J Biol Chem. 270:26746–26749.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Maeda K, Okubo K, Shimomura I, Funahashi
T, Matsuzawa Y and Matsubara K: CDNA cloning and expression of a
novel adipose specific collagen-like factor, apM1 (AdiPose Most
abundant Gene transcript 1). Biochem Biophys Res Commun.
221:286–289. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Arita Y, Kihara S, Ouchi N, Takahashi M,
Maeda K, Miyagawa J, Hotta K, Shimomura I, Nakamura T, Miyaoka K,
et al: Paradoxical decrease of an adipose-specific protein,
adiponectin, in obesity. Biochem Biophys Res Commun. 257:79–83.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Matsubara M: Plasma adiponectin decrease
in women with nonalcoholic Fatty liver. Endocr J. 51:587–593. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Maebuchi M, Machidori M, Urade R, Ogawa T
and Moriyama T: Low resistin levels in adipose tissues and serum in
high-fat fed mice and genetically obese mice: Development of an
ELISA system for quantification of resistin. Arch Biochem Biophys.
416:164–170. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tasso F, Clop J and Sarles H: The
interaction of ethanol, dietary lipids and proteins on the rat
pancreas. II. Ultrastructural study. Digestion. 4:23–34. 1971.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wilson JS, Korsten MA, Leo MA and Lieber
CS: Combined effects of protein deficiency and chronic ethanol
consumption on rat pancreas. Dig Dis Sci. 33:1250–1259. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hiatt JL and Gartner LP: Color textbook of
histology. WB Saunders Co. Philadelphia USA: 1–592. 2007.
|
|
47
|
Sathishsekar D and Subramanian S:
Beneficial effects of Momordica charantia seeds in the
treatment of STZ-induced diabetes in experimental rats. Biol Pharm
Bull. 28:978–983. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Terauchi Y, Takamoto I, Kubota N, Matsui
J, Suzuki R, Komeda K, Hara A, Toyoda Y, Miwa I, Aizawa S, et al:
Glucokinase and IRS-2 are required for compensatory beta cell
hyperplasia in response to high-fat diet-induced insulin
resistance. J Clin Invest. 117:246–257. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kamada T, Hata J, Kusunoki H, Ito M,
Tanaka S, Kawamura Y, Chayama K and Haruma K: Eradication of
Helicobacter pylori increases the incidence of
hyperlipidaemia and obesity in peptic ulcer patients. Dig Liver
Dis. 37:39–43. 2005. View Article : Google Scholar : PubMed/NCBI
|