Introduction
Breast cancer is the most common malignant disease
in women (1). Meanwhile, 90% of
cancer-related mortality is caused by metastasis formed at distant
anatomic sites by disseminated primary tumor cells (2). Breast cancer patients frequently
develop metastases at multiple sites. Primary breast cancer cells
metastasize through the blood vessels to various distant organs,
preferentially, to the lung, liver and bones (3). Surgery, radiation therapy and
chemotherapy were the effective strategies used for treating
primary tumor; however, these treatments have limited utility in
curbing the metastatic spread of cancer cells, resulting in
metastasis (4). Thus, novel therapy
targets are required for treating breast cancer metastasis.
microRNAs (miRs) are short non-coding RNAs, 18–22
nucleotides long, which were discovered in mammals as a large class
of evolutionarily conserved small noncoding RNAs (5). miRNAs regulate gene expression at the
post-transcriptional level by targeting the 3′-untranslated region
(UTR) of mRNA transcripts (6).
Furthermore, miRNAs have emerged as powerful regulators of a wide
range of biological processes through influencing protein
translation. It was quickly recognized that miRNAs can be
efficiently inhibited by antisense technologies for prolonged
periods, which has increased the interest of researchers in the
inhibition of specific miRNAs as a feasible therapeutic option for
several diseases (7,8).
A previous study has demonstrated that miR-376b
regulates angiogenesis in middle cerebral artery occlusion
(9). Meanwhile, miR-376-5p mediates
the M3 subtype of muscarinic acetylcholine receptor-inducing
cardioprotection (10). However, the
role of miR-376b in regulating breast cancer metastasis remains
elusive. In the present study, miR-376b expression in normal breast
tissue (NBT), JC and 4T1 breast cancer cells was determined by
qPCR. Furthermore, in vitro and in vivo experiments
were performed to determine the effect of miR-376b on breast cancer
metastasis. The direct target of miR-376b was determined by the
luciferase assay and western blotting. The results indicated that
silencing of miR-376b significantly inhibited 4T1 cell migration
and invasion in vitro. Lung metastasis was also evidently
decreased following silencing of miR-376b in 4T1 cells. Moreover,
luciferase assay and western blotting identified that Hoxd10 is the
direct target of miR-376b during the regulation of breast cancer
metastasis. To the best of our knowledge, the present study was the
first to demonstrate a role of miR-376b in promoting breast cancer
metastasis. Therefore, it may be a novel therapy target and
prognostic biomarker for breast cancer.
Materials and methods
Cell culture and transfection
The JC and 4T1 breast cancer cell lines were
obtained from American Type Culture Collection (Manassas, VA, USA).
The cells were cultured in Dulbecco's modified Eagle's medium
(DMEM) containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Grand Island, NY, USA). All the cells were
maintained in a humidified atmosphere containing 5% CO2
at 37°C. Cell transfection was performed using FuGENE HD
Transfection Reagent (Roche Diagnostics, Indianapolis, IN, USA)
according to the manufacturer's instructions. Briefly, 4T1 cells
were seeded in 6-well plates at a density of 2×105
cells/well and cultured for 24 h to reach 70–80% confluence. A
total of 2 µg plasmids (encoding miR-376b-mimic, miR-376b and
miR-NC) were diluted in 100 µl DMEM without serum and 5 µl FuGENE
HD Transfection Reagent was added to the tubes containing diluted
DNA. These were then mixed and the transfection complex was
incubated for 15 min at room temperature, and then added to the
6-well plates.
Vector construction
The miR-376b and miR-376b-mimic expression plasmids
and the negative control (miR-NC) were purchased from the Origene
Company (Rockville, MD, USA). Plasmids were extracted using
EndoFree Plasmid Giga kits (#12362; Qiagen GmbH, Hilden, Germany)
from DH5α (Genewiz, Suzhou, China) Escherichia coli
transformants and stored at −20°C until further use. The
concentration was determined by measuring the A260/A280 ratio using
an ND 2000 spectrophotometer (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA).
Target prediction
The miRWalk database (http://www.ma.uni-heidelberg.de/apps/zmf/mirwalk/) and
other programs (miRanda, Sanger miRDB, RNAhybrid and Targetscan)
were used variously for target prediction The online tool miRWalk
2.0 (http://www.umm.uni-heidelberg.de/apps/zmf/mirwalk/predictedmirnagene.html)
was used to predict potential target mRNAs of miR-376b.
Quantitative polymerase chain reaction
(qPCR)
The total RNA was extracted from each experimental
group using TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Inc., Carlsbad, CA, USA) according to the manufacturer's
instructions. The RNA concentration was assessed
spectrophotometrically at 260 nm (ND 2000; Thermo Fisher
Scientific, Inc.). In addition, reverse transcription was performed
on the isolated total RNA using a Reverse Transcription kit
(#RR047A; Takara Bio, Inc., Otsu, Japan), and PCR was performed
using a SYBR Premix Ex Taq kit (#RR820A; Takara Bio, Inc.),
according to the manufacturer's instructions. gDNA eraser (1.0 µl),
5X gDNA eraser buffer (2.0 µl) and mRNA template (2.0 µg) were
added into one well. Then RNAase-free H2O was added to
the final volume (10.0 µl). The well was incubated at room for 5
min. Reverse transcription was performed at 65°C for 5 min, 30°C
for 10 min, 42°C for 10–30 min and 2°C for 3 min. The PCR
conditions were as follows: Denaturation at 94°C for 2 min;
amplification for 30 cycles at 94°C for 0.5 min, annealing at 58°C
for 0.5 min and extension at 72°C for 1 min; followed by a terminal
elongation step at 72°C for 10 min. The reaction was performed on a
CFX96 thermal cycler (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). U6 was amplified as an internal control and the Ct value of
each PCR product was calculated, and the fold change analyzed. The
m-miR-376b and m-U6 primers were supplied by Ribo Biotechnology
(Guangzhou, China) but the sequences were not supplied due to the
rules of the company. The results was analyzed using the software
that supplied in the CFX-96 (Bio-Rad Laboratories, Inc.)
Luciferase assays
The miR-376b binding site was synthesized and cloned
into an pMIR-REPORT vector (Ambion; Thermo Fisher Scientific, Inc.)
to generate pMiRluc-376b 3′-UTRs of Hoxd10 containing miR-376b
binding sites. These were then amplified and cloned into the same
vector to generate pMiRluc-Hoxd10. The reporter was co-transfected
with a cytomegalovirus β-galactosidase vector using FuGENE HD
(Promega Corporation, Madison, WI, USA). The luciferase activity
was measured 4 h later using the luciferase reporter assay (#E1500;
Promega Corporation). Values were normalized against
β-galactosidase activity.
Western blot analysis
4T1 cells were transfected with m-miR-376b-mimic and
miR-NC, and 48 h later the total protein was collected. Cells were
lysed on ice for 30 min with radioimmunoprecipitation assay lysis
buffer (Beyotime Institute of Biotechnology, Shanghai, China). The
proteins (20 µg) were separated by 10% SDS-PAGE and electronically
transferred onto a polyvinylidene difluoride membrane (EMD
Millipore, Billerica, MA, USA). Following blocking in 5% milk in
TBS/T buffer for 1 h at 37°C, the membranes were incubated with the
recommended dilution of primary antibodies against Hoxd10
(#ab172865; rabbit polyclonal; 1:800 for 1 h at 37°C; Abcam,
Cambridge, MA, USA), and GAPDH (sc-25778; 1:5,000; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) for 1 h at 37°C. This was
followed by incubation with goat anti-rabbit (#ab6721; 1:2,000) and
anti-mouse (#ab6789; 1:2,000) horseradish peroxidase-conjugated
secondary antibodies (Abcam) at 1 h for 37°C. Peroxidase-labeled
bands were visualized using an enhanced chemiluminescence kit
(#32106; Pierce Protein Biology; Thermo Fisher Scientific, Inc.,
Rockford, IL, USA). The ratio of Hoxd10/GAPDH was then calculated
using densitometry, and values were normalized by dividing with the
ratio of the blank sample. Protein expression was evaluated using a
bicinchoninic acid assay kit (Beyotime, Beijing, China).
Invasion assay
The invasion of 4T1 cells was evaluated using
Transwell-24 units (pore size, 8 µm; EMD Millipore), as described
previously with some modification (11). Briefly, the filter of the Transwell
plate was covered with 50 µl matrigel (BD Biosciences, Franklin
Lakes, NJ, USA). Following matrigel polymerization, 500 µl DMEM
containing 10% FBS was placed in the lower chamber and 100 µl 4T1
suspension (2×104 cells/well in DMEM without 10% FBS)
was placed in the upper chamber. After 48 h incubation, cells on
the back surface were fixed in 100% methanol, stained with 0.05%
crystal violet, counted in at least six randomly chosen fields and
photographed.
Migration assay
The migration of 4T1 cells was evaluated using
Transwell-24 units, as described previously and with some
modification. Briefly, 500 µl DMEM containing 10% FBS was placed in
the lower chamber and a 100 µl 4T1 suspension (2×104
cells/well; in DMEM without 10% FBS) was placed in the upper
chamber. After a 48 h incubation, cells on the back surface were
fixed in 100% methanol, stained with 0.05% crystal violet, counted
in at least six randomly chosen fields and photographed (BX600;
Olympus Corporation, Tokyo, Japan).
Animal study
Female BALB/c mice (age, 5–6 weeks) were purchased
from the Animal Center of Sichuan University (Chengdu, China), with
six mice per group. All animal research was approved by the Sichuan
Provincial People's Hospital Committee on Animal Research. The mice
were housed at 26°C under a 12-h light/dark cycle, with ad
libitum access to food and water. The 4T1 cell stable
transfection with miR-NC and miR-376b-mimic (2×106
cells) were injected into the mouse tail vein to establish the 4T1
lung metastasis model. The survival percentage of the mice were
analyzed. At 20 days after tumor cell injection, the mice were
anesthetized using diethyl ether (100 mg/kg; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) then sacrificed by cervical dislocation.
The lungs were subsequetly dissected for further study.
H&E staining
Paraffin-embedded lung sections (3–5 µm) were
mounted on 3-aminopropyl triethoxysilane-coated glass slides.
Sections were deparaffinized in xylene, treated with a graded
series of alcohol [100, 95 and 80% ethanol/double-distilled
H2O (v/v)], and rehydrated in PBS (pH 7.4). Following
the PBS washes, the slides were stained with hematoxylin for 3 min
at room temperature followed by washing with
H2O2 for 10–15 min. Next, the slides were
stained with eosin for 2–3 min at room temperature followed by
washing with H2O2 for 3–5 min. The slides
were then coated with neutral balsam after drying with a graded
series of alcohol. All specimens were evaluated using a BX600
microscope (Olympus Corporation) and Spot Fiex camera.
Statistical analysis
Statistical comparisons of all the results were
analyzed using a one-way analysis of variance. Statistical analyses
were performed using SPSS version 19.0 (IBM SPSS, Armonk, NY, USA).
The values are expressed as the mean ± standard error of the mean.
P<0.05 was used to indicate a statistically significant
difference.
Results
Silencing of miR-376b inhibits 4T1
cell migration and invasion in vitro
In order to further determine the role of miR-376b
in breast cancer, NBT from BALB/c mice, and mouse breast cancer JC
and 4T1 cells were collected for detection of miR-376b expression
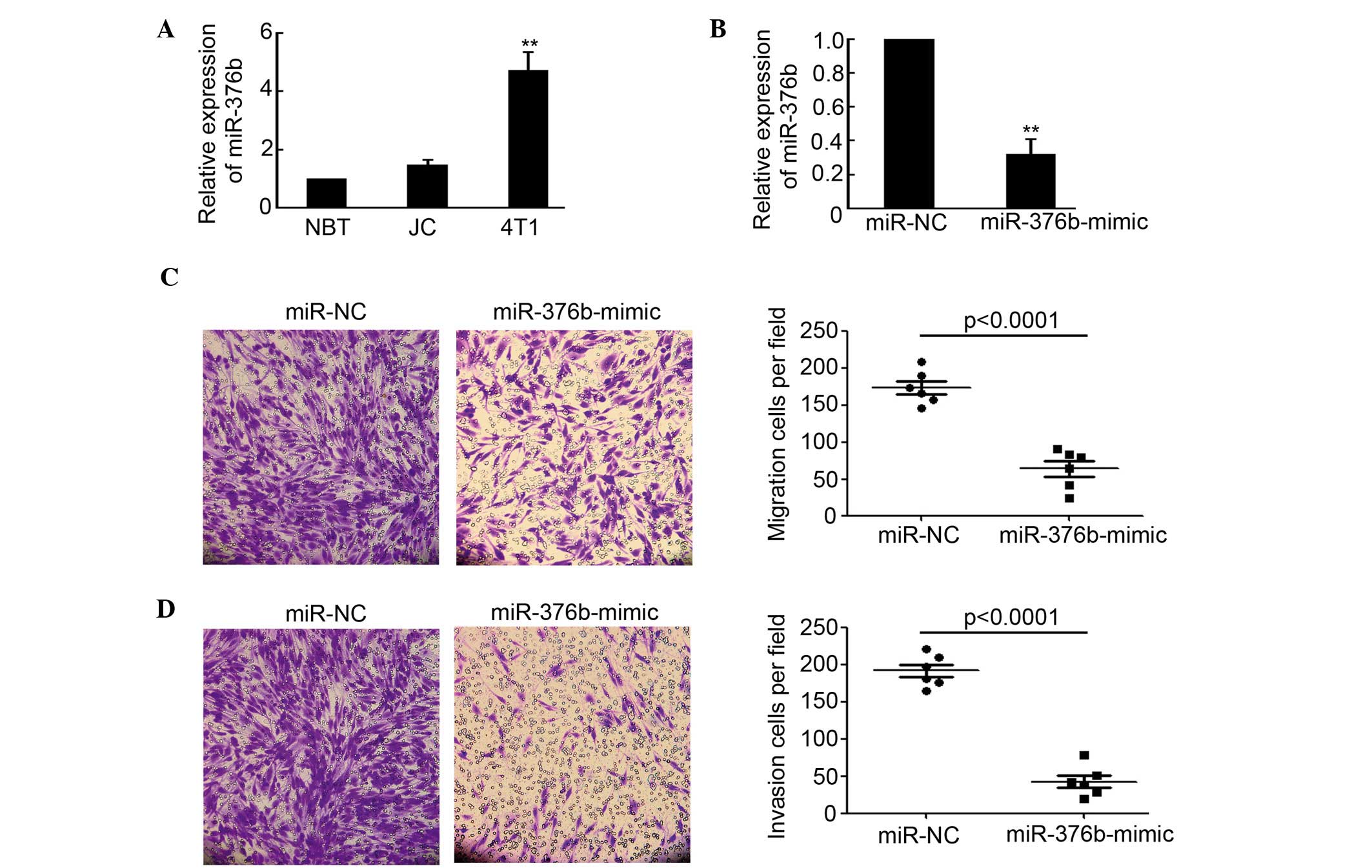
by qPCR. As shown in Fig. 1A,
miR-376b expression in 4T1 cells, which get a high capacity of
migration and invasion, was significantly increased compared with
NBT (P<0.01). However, there was no evident difference between
NBT and JC on miR-376b expression. The miR-376b-mimic was employed
to inhibit the activity of miR-376b. The results indicated that
miR-376b expression in 4T1 cells was evidently decreased after
being transfected with the miR-376b-mimic (Fig. 1B). Moreover, the migration and
invasion capacity of 4T1 cells that were transfected with miR-NC or
the miR-376b-mimic were determined by transwell migration assay and
transwell-based matrigel invasion assay. The results shown in
Fig. 1C and D indicated that
silencing of miR-376b significantly inhibited 4T1 cell migration
and invasion. Overall, the results of the present study
demonstrated that miR-376b was important in regulating breast
cancer migration and invasion in vitro.
Hoxd10 is a direct target of
miR-376b
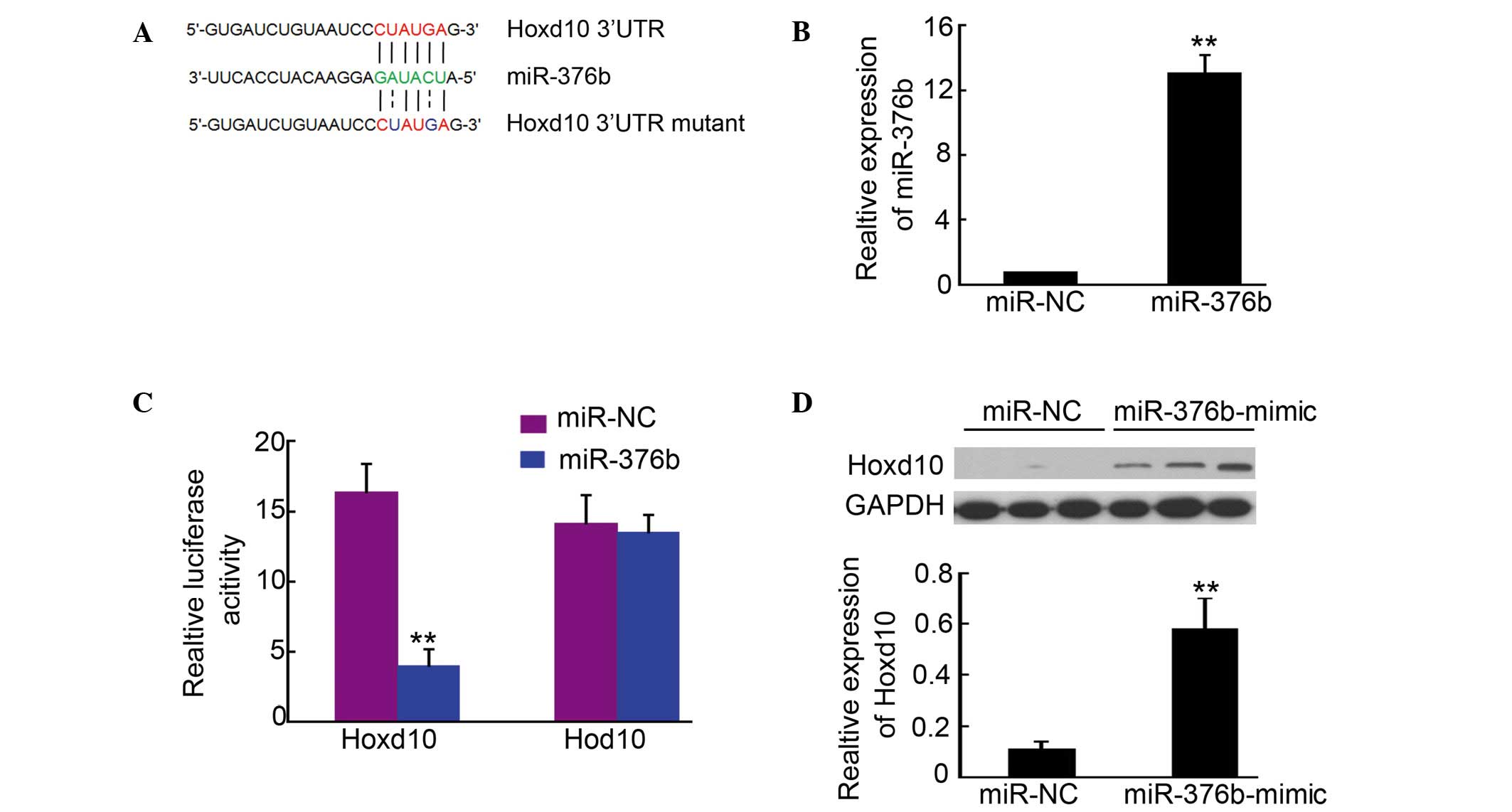
In order to determine the targets of miR-376b, a
large number of potential target proteins were screened in a
database library to identify promising miRNA binding seed sequences
within the 3′-UTR. Using bioinformatics analysis tools (Targetscan.org, RNA.org and microRNA
Seq), Hoxd10 was identified as a candidate target for miR-376b
(Fig. 2A). In order to verify this
result, Hoxd10-3′UTR containing an miR-376b binding site was cloned
downstream of the luciferase open reading frame. Meanwhile, a
Hoxd10-3′UTR mutant, which contained a mutated miR-376b binding
site, was also introduced into the luciferase construct. The
plasmid expressing miR-376b was transfected in the 293 cells, and
puromycin was used to select the stable expression cells. qPCR
analysis confirmed that miR-376b was upregulated in the cells
(Fig. 2B). Next, the
luciferase-Hoxd10-3′UTR and luciferase-Hoxd10-3′UTR mutant
constructs were transfected into 293 cells for stable miR-376b
expression. After 4–6 h, luciferase expression in Hoxd10-3′UTR
constructs was significantly reduced compared with the control
(Fig. 2C). Moreover, a consistent
reduction of luciferase expression was observed in cells
transfected with miRNA binding site mutant plasmids (Fig. 2C). Further western blotting indicated
that Hoxd10 expression in 4T1 cells was significantly increased
after being transfected with the miR-376b-mimic (Fig. 2D). The results described above
demonstrated that Hoxd10 is a direct target of miR-376b.
Silencing of miR-376b inhibits 4T1
cell metastasis in mice
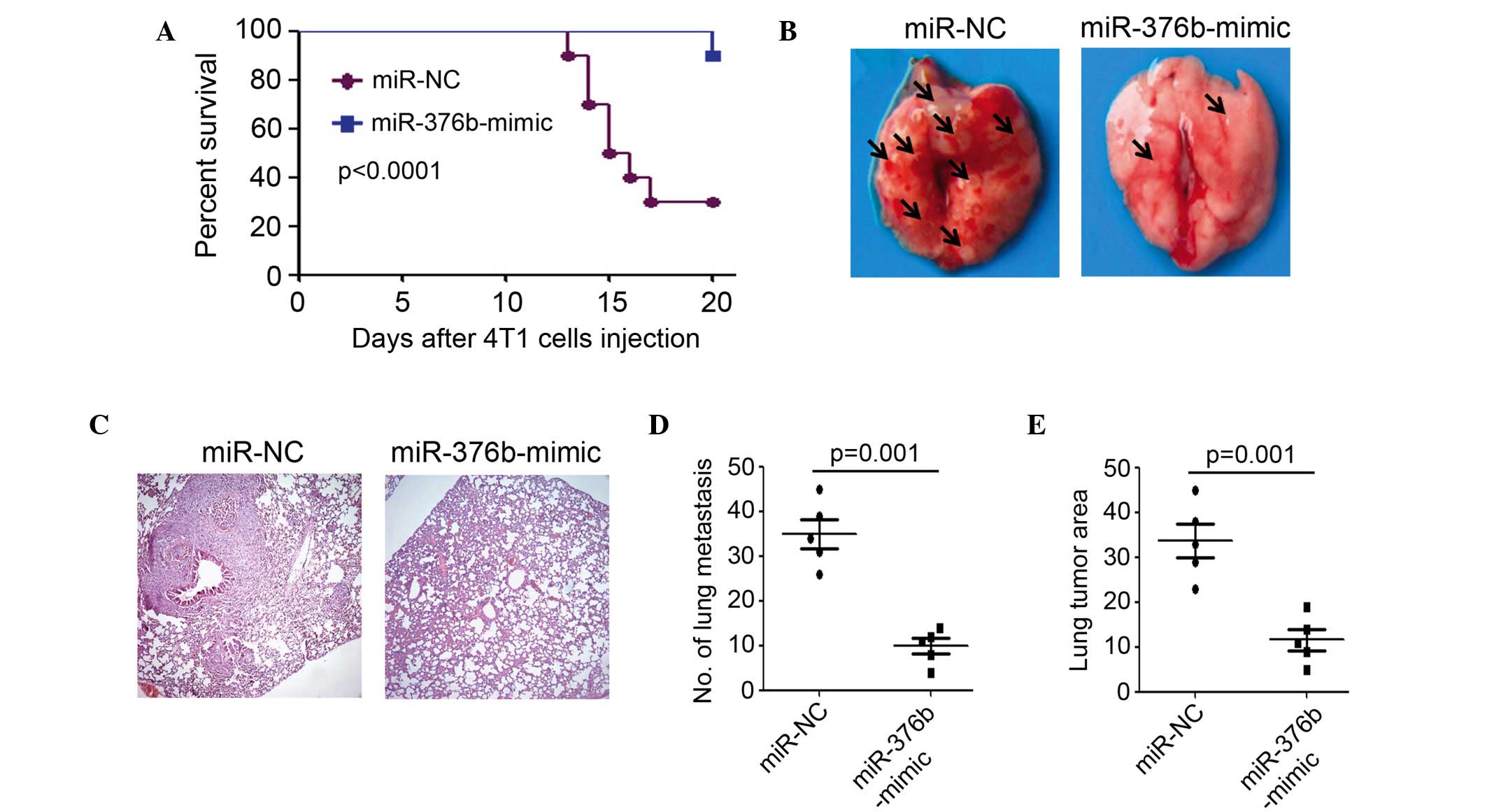
In order to establish a breast cancer metastasis
model, 2×106 4T1 cells that were stably transfected with
miR-NC or the miR-376b-mimic, were intravenously injected into
Balb/c mice separately. The results of the present study indicated
that silencing of miR-376b after transfection with the
miR-376b-mimic significantly increased the survival percentage,
compared with the miR-NC transfected group (Fig. 3A). In total, 7 out of 10 mice died in
the miR-NC transfected group, whereas only 1 out of 10 mice died in
the miR-376b-mimic transfected group at 20 days post cancer cell
injection (Fig. 3A). Bright-field
microscopy and H&E staining demonstrated that less lung
metastasis was found in the miR-376b-mimic transfected group
compared to the miR-NC transfected group (Fig. 3B and C). In total, a 69% decrease in
the number of macroscopically visible pulmonary metastasis was
achieved through silencing of miR-376b (P=0.001) (Fig. 3D). Moreover, a smaller tumor area
(tumor area/total lung area) was observed in the miR-376b-mimic
compared to the miR-NC transfected group (Fig. 3E). Overall, 4T1 cell metastasis in
mice was significantly inhibited after silencing miR-376b.
Silencing of miR-376b and induction of
Hoxd10 in the lungs of mice
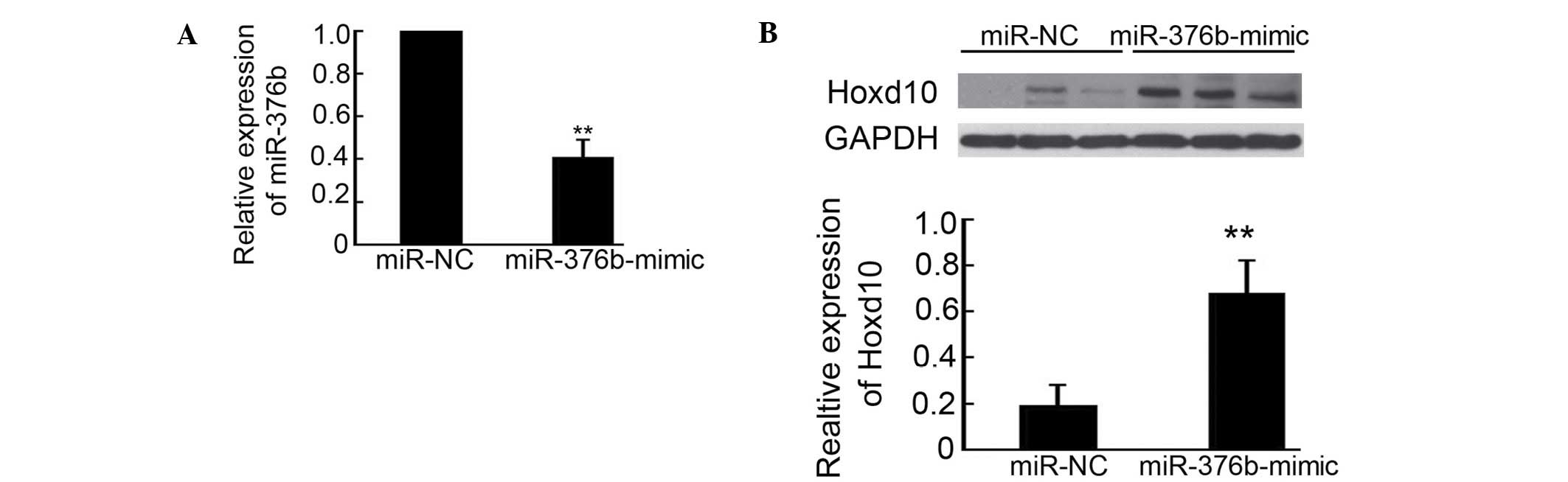
Relative to the effects of the miR-376b-mimic on
decreasing 4T1 metastasis in mice, miR-376b and Hoxd10 expression
in the lungs after 4T1 cancer cell injection was determined by qPCR
and western blotting. As shown in Fig.
4A, the mice injected with the miR-376b-mimic transfected 4T1
cells exhibited a 61% reduction of miR-376b expression in the
lungs, compared with miR-NC transfected group. Meanwhile, Hoxd10
expression was upregulated 3.6-fold after silencing miR-376b
(Fig. 4B). These results are
consistent with the observation that silencing of miR-376b
inhibited 4T1 cell metastasis in mice.
Discussion
The present study initially examined the expression
of miR-376b in NBT, JC and 4T1 breast cancer cells by qPCR. The
results indicated that miR-376b was highly expressed in 4T1 cells,
which gets the high capacity of migration and invasion, compared
with normal mouse breast tissue (8,12).
Further migration and invasion assays demonstrated that silencing
of miR-376b significantly inhibited 4T1 cell migration and invasion
in vitro. Lung metastasis was also evidently decreased after
silencing of miR-376b in 4T1 cells. Luciferase assay and western
blotting identified that Hoxd10 is the direct target of miR-376b
during the regulation of breast cancer metastasis. To the best of
our knowledge, the present study was the first to demonstrate the
promoting breast cancer metastasis role of miR-376b.
Hoxd10 belongs to the type I class homeobox (Hox)
family, and is important for suppressing angiogenesis and
maintaining a quiescent, differentiated phenotype in endothelial
cells (13). In particular, Hoxd10
suppresses expression of genes that directly affect remodeling of
the extracellular matrix and cell migration during angiogenesis,
including α3 integrin, matrix metalloproteinase 14 and
urokinase-type plasminogen activator receptor (14). A previous study has indicated that
Hoxd10 inhibits the development and progression of head and neck
squamous cell carcinoma (15).
Meanwhile, various studies have demonstrated that Hoxd10 was
involved in the metastasis of breast, bladder, gastric and
hepatocellular cancer (16–19). In the present study, a luciferase
assay was performed that indicated that Hoxd10 is the direct target
of miR-376b. Silencing of miR-376b in 4T1 cells significantly
induced Hoxd10 expression both in vitro and in vivo.
Meanwhile, inducing expression of Hoxd10 by miR-376b-mimic
significantly inhibited 4T1 cell migration and invasion in
vitro and lung metastasis in vivo.
The 4T1 cell line was originally derived from a
spontaneous mouse mammary carcinoma from the BALB/c strain
(20). A previous study has reported
that 4T1 cells get a high capacity of migration and invasion
(8,12). The intravenous injection of 4T1 cells
into the BALB/c mice could form metastasis in the lungs. Therefore,
a 4T1 breast mouse model is appropriate to mimic human breast
cancer metastasis in vivo, and for investigating the effect
of miR-376b on metastasis. In the present study, the miR-NC or
miR-376b-mimic transfected 4T1 cells was injected into the mouse
tail vein. At 20 days post 4T1 cell injection, the survival
percentage, lung metastasis formation number and lung metastasis
area were analyzed. The results indicated the metastasis capacity
of miR-376b in vivo. It is consistent with the results in
the migration and invasion assays. Moreover, lower expression of
miR-376b and higher expression of Hoxd10 were also observed in the
lungs from the miR-376b-mimic transfected group.
In summary, the present study initially demonstrated
the role of miR-376b in regulating breast cancer metastasis in
vitro and in vivo. Moreover, the luciferase assay
indicated that Hoxd10 is the direct target of miR-376b during the
regulation of breast cancer metastasis. Thus, these results may
provide a novel therapy target and prognosis biomarker for breast
cancer.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fidler IJ: The pathogenesis of cancer
metastasis: The ‘seed and soil’ hypothesis revisited. Nat Rev
Cancer. 3:453–458. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee YT: Breast carcinoma: Pattern of
metastasis at autopsy. J Surg Oncol. 23:175–180. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Steeg PS: Tumor metastasis: Mechanistic
insights and clinical challenges. Nat Med. 12:895–904. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baek D, Villén J, Shin C, Camargo FD, Gygi
SP and Bartel DP: The impact of microRNAs on protein output.
Nature. 455:64–71. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van Rooij E, Marshall WS and Olson EN:
Toward microRNA-based therapeutics for heart disease: The sense in
antisense. Circ Res. 103:919–928. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ma L, Reinhardt F, Pan E, Soutschek J,
Bhat B, Marcusson EG, Teruya-Feldstein J, Bell GW and Weinberg RA:
Therapeutic silencing of miR-10b inhibits metastasis in a mouse
mammary tumor model. Nat Biotechnol. 28:341–347. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li LJ, Huang Q, Zhang N, Wang GB and Liu
YH: miR-376b-5p regulates angiogenesis in cerebral ischemia. Mol
Med Rep. 10:527–535. 2014.PubMed/NCBI
|
|
10
|
Pan Z, Guo Y, Qi H, Fan K, Wang S, Zhao H,
Fan Y, Xie J, Guo F, Hou Y, et al: M3 subtype of muscarinic
acetylcholine receptor promotes cardioprotection via the
suppression of miR-376b-5p. PLoS One. 7:e325712012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dai L, Cui X, Zhang X, Cheng L, Liu Y,
Yang Y, Fan P, Wang Q, Lin Y, Zhang J, et al: SARI inhibits
angiogenesis and tumour growth of human colon cancer through
directly targeting ceruloplasmin. Nat Commun. 7:119962016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cronin PA, Wang JH and Redmond HP: Hypoxia
increases the metastatic ability of breast cancer cells via
upregulation of CXCR4. BMC Cancer. 10:2252010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Myers C, Charboneau A, Cheung I, Hanks D
and Boudreau N: Sustained expression of homeobox D10 inhibits
angiogenesis. Am J Pathol. 161:2099–2109. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sekar P, Bharti JN, Nigam JS, Sharma A and
Soni PB: Evaluation of p53, HoxD10 and E-Cadherin status in breast
cancer and correlation with histological grade and other prognostic
factors. J Oncol. 2014:7025272014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hakami F, Darda L, Stafford P, Woll P,
Lambert DW and Hunter KD: The roles of HOXD10 in the development
and progression of head and neck squamous cell carcinoma (HNSCC).
Br J Cancer. 111:807–816. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li Q, Ding C, Chen C, Zhang Z, Xiao H, Xie
F, Lei L, Chen Y, Mao B, Jiang M, et al: miR-224 promotion of cell
migration and invasion by targeting Homeobox D 10 gene in human
hepatocellular carcinoma. J Gastroenterol Hepatol. 29:835–842.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu Z, Zhu J, Cao H, Ren H and Fang X:
miR-10b promotes cell invasion through RhoC-AKT signaling pathway
by targeting HOXD10 in gastric cancer. Int J Oncol. 40:1553–1560.
2012.PubMed/NCBI
|
|
18
|
Vardhini NV, Rao PJ, Murthy PB and
Sudhakar G: HOXD10 expression in human breast cancer. Tumour Biol.
35:10855–10860. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xiao H, Li H, Yu G, Xiao W, Hu J, Tang K,
Zeng J, He W, Zeng G, Ye Z and Xu H: MicroRNA-10b promotes
migration and invasion through KLF4 and HOXD10 in human bladder
cancer. Oncol Rep. 31:1832–1838. 2014.PubMed/NCBI
|
|
20
|
Pulaski BA, Terman DS, Khan S, Muller E
and Ostrand-Rosenberg S: Cooperativity of staphylococcal aureus
enterotoxin B superantigen, major histocompatibility complex class
II, and CD80 for immunotherapy of advanced spontaneous metastases
in a clinically relevant postoperative mouse breast cancer model.
Cancer Res. 60:2710–2715. 2000.PubMed/NCBI
|