Introduction
Gastrointestinal homeostasis is one of several
factors for longevity and human health. Although the common causes
of gastritis are related to non-steroidal anti-inflammatory drugs,
burns, brain injury, autoimmunity, and Helicobacter pylori
infection (1), modern lifestyles and
dietary habits, such as non-exercise related obesity, cigarette
smoking, heavy alcohol intake, and fast food consumption, are also
major driving forces of gastric disorders (2). There are several mechanisms involved in
the pathogenesis of gastritis, depending on its cause. However, the
pathological outcomes are similar, including inflammation, gastric
mucosal cell alterations (degeneration, apoptosis, and necrosis),
barrier damage, and hemorrhage. Interestingly, endoplasmic
reticulum (ER) stress has been associated with the pathogenicity of
gastritis through apoptotic pathways (3). Moreover, oxidative-induced gastric
disorders play a crucial role in gastric mucosal cell injury,
particularly from exogenous stimuli such as diet and drugs
(4). Parietal cell, acid secreting
cell, and chief cell, pepsinogen secreting cell, are affected
during ethanol-induced gastritis (5)
that is characterized by narrowing of the mucosal membrane and
atrophy of secretory cells in the glands. Morphological
identification of these cells is important for new therapeutic
modalities (6). Unfortunately, there
are no reports describing the ultrastructural integrity of rough ER
(RER) in parietal and chief cells in association with
ethanol-induced gastritis.
Recently, several natural products have been
introduced to alleviate gastrointestinal disorders with their
anti-oxidative property (7,8). Nevertheless, omeprazole, a basic
medication on the World Health Organization's list of essential
medicine, is now globally available for treatment of some
gastro-intestinal diseases. Omeprazole has the ability to inhibit a
proton pump to block acid secretion in the stomach. Importantly, it
also modulates endogenous oxidative stress and prevents
proinflammatory cytokines release (9). However, it is doubtful that whether
omeprazole can preserve the RER integrity in gastric mucosal cells
related to gastrointestinal disorders.
The present study aimed to demonstrate the
anti-oxidative property and epithelial protective effect of
omeprazole in an ethanol-induced gastritis rat model. Microscopic
and fine morphological structures of the cells in gastric-mucosal,
-submucosal, and -muscular layers were compared in rats with or
without omeprazole-treatment. Anti-oxidative stress marker,
super-oxide dismutase (SOD), and aquaporin (AQP)-4, acid
suppression (10) and
re-epithelialized marker (11,12),
were also examined using immunohistochemical study. The results of
the present study may provide a better understanding of the
mechanical action of omeprazole correlates to the pathogenesis of
gastritis, together with for the benefit of a new therapeutic
approach.
Materials and methods
Ethic statement
All activities related to the animal studies were
performed in accordance with the Ethical Principles and Guidelines
for the Use of Animals, National Research Council of Thailand, and
approved by National Laboratory Animal Center-Animal Care and Use
Committee (ACUC), Mahidol University. Sprague-Dawley rats were
provided by National Laboratory Animal Center, Mahidol University
and housed in the barrier system with temperature, ventilation, and
humidity control. Animal were provided with standard diet (Perfect
Companion Co., Ltd., Bangkok, Thailand) and chlorinated water ad
libitum and a 12/12 h light/dark cycle.
Omeprazole preparation
Omeprazole was purchased from the Government
Pharmaceutical Organization, Thailand. Omeprazole granules were
removed from the capsule and ground into a powder. To dissolve the
powder, 8.4% (w/v) of sodium bicarbonate (8.4 g in 100 ml deionized
water) was made as a diluent. An omeprazole suspension was prepared
by dissolving the powder in a diluent with strong mixing. The
suspension was kept at room temperature for no more than 15
days.
Induction gastritis rats
Thirty rats were included to the study and equally
divided into two groups with or without omeprazole treatment. In
each group, five rats were randomly allocated into each of three
subgroups according to induction period: 4, 7, and 14 days. To
induce gastritis, all rats were fasted overnight with free access
to water. Fasted rats were orally administrated a single dose of 5
ml/kg absolute ethanol. Therefore, 20 mg/kg of omeprazole and water
were administered daily to treated and untreated groups,
respectively (13). Routine clinical
observations were performed under attending veterinarian
control.
Specimen collection
All rats in each subgroup were humanely euthanized
with overdose of carbon dioxide (CO2) inhalation on 4,
7, and 14 days post-induction. Grandular stomach was removed and
cut into three parts, one was fixed in 10% neutral buffer formalin
for 48 h and subjected to the histopathological examination, the
second part was separated for PGE-2 activity study, and the
remaining part was fixed in 2.5% glutaraldehyde in 0.1 M sucrose
phosphate buffer (SPB), pH 7.4 for 1 h and attended to the electron
microscopic studies.
PGE-2 activity assessment
Collected grandular stomachs were homogenized in
1.15% potassium chloride with 1:5 ratio (w/v) and then centrifuged
at 10,000 g, 4°C for 10 min. The supernatant was collected for
PGE-2 activity using commercial enzyme-linked immunosorbent assay
(ELISA) kit (Cayman Chemical, Ann Arbor, MI, USA). The level of
PGE-2 in grandular stomach was measured based on the conjugate
reaction of PGE-2 and PGE-2 acetylcholinesterase (AChE) which was
obviously detected at 412 nm absorbance by ELISA reader.
Electron microscopic studies
Scanning electron microscopy
(SEM)
Fixed stomachs were washed three times with 0.1 M
SPB for 10 min each and secondarily fixed with 1% osmium tetroxide
in 0.1 M SPB for 1 h. After three consecutive wash, the stomachs
were dehydrated with graded ethyl-alcohol and dried in liquid
CO2 using critical point dryer (HCP-2; HITACHI, Ltd.,
Tokyo, Japan). Then, the stomachs were mounted on an aluminum stub
using double-side carbon tape and coated with gold film to 20 nm
thickness using a sputter coater (K550; EMITECH, UK). All specimens
were examined under a scanning electron microscope (JSM-6610LV;
JEOL, Tokyo, Japan) with a 15 kV acceleration voltage.
Transmission electron microscopy
(TEM)
Fixed stomachs were washed and secondary fixed as
mentioned above. The specimens were then dehydrated in graded
ethanol, infiltrated with LR White resin (EMS, Hatfield, PA, USA),
embedded in capsule beams, and finally polymerized at 65°C for 48
h. Then the specimens were cut in 90–100 nm thickness and stained
with uranyl acetate and lead citrate. The ultra-thin sections were
investigated under a transmission electron microscope (HT7700;
HITACHI, Ltd.).
Ultrastructural changes in the chief and parietal
cells were evaluated by focusing on the RER and mitochondrial
alterations, and then semi-quantified by the H-score, a
multiplication between percentage of the cells with an RER or
mitochondrial alterations (0–100%) and severity score (four grades;
0=no changing, 1=low severity, 2=moderate severity, and 3=severe
changing). At least 50 chief and parietal cells each were counted
per animal.
Histopathological studies
Fixed stomachs were dehydrated in graded ethanol,
infiltrated and embedded in paraffin, sectioned to 4-µm thickness,
and then stained with hematoxylin and eosin (H&E). All
histopathological changes in gastric-mucosa, -submucosa, and
-muscular layer were examined under light microscope and scored by
H-score, as a multiplication between the severity score (0–3) and
the histological changing area (0–100%). Briefly, the severity was
scored in four grades (0=no changing, 1=mild severity, 2=moderate
severity or progressive stage, and 3=severe or end stage). In
association with the distribution of each histological change,
percentage of affected area per section of each animal was
simultaneously estimated.
The gross hemorrhagic area was quantified by an
imaging analysis program (ImageJ® v.1.36; National
Institutes of Health; Bethesda, MD, USA). Briefly, stomach images
were acquired in color. The non-hemorrhagic area was adjusted to
white by the replace mode. The adjusted images were transformed to
gray scale and then the hemorrhagic area was localized by the
threshold mode. A line was drawn over the area of glandular portion
and finally the hemorrhagic area was measured as the percentage
hemorrhagic area/grandular portion.
Immunohistochemical studies
SOD and AQP-4 expression on gastric mucosal layer
were used to demonstrate anti-oxidative property and acid
suppression/re-epithelialized effect of omeprazole by
immunohistochemical technique. Polyclonal rabbit anti-AQP-4 (D2408;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) and polyclonal
rabbit anti-Mn-SOD (2683863; Millipore, Billerica, MA, USA) were
used as primary antibodies in cooperation with rabbit anti-IgG
(Santa Cruz Biotechnology, Inc.) as a negative control for the
staining system validation. In addition, rat brain and liver were
used as a positive control for AQP-4 and SOD labeling,
respectively. Sections were deparaffinized in xylene, rehydrated in
graded ethanol, unmasked antigens in heated citrate buffer pH 6.0,
and peroxidase/non-specific binding blocked with EnVision FLEX/HRP
blocking reagent (K8002; DAKO; Agilent Technologies, Inc., Santa
Clara, CA, USA). The sections were incubated with primary antibody
for 1 h and labeled polymer HRP anti-mouse/rabbit (K8002; DAKO) for
20 min, then visualization with diaminobenzidine; DAB (K8002;
DAKO). Finally, the sections were counterstained in hematoxylin and
mounted with Permount®.
In each animal (belong to any subgroup; 4 and 7 days
post induction), ten color images of mucosal layer were randomly
acquired by a light microscope (BX51; Olympus Corp., Tokyo, Japan)
and digital camera (DP70; Olympus Corp.) at ×400 magnifications.
Therefore fifty images were included to the analysis per subgroup.
SOD and AQP-4 expression were evaluated using the H-score.
Percentage area of expression/field (0–100%) and intensity score
(three grades; 0=negative staining, 1=low intensity staining,
2=moderate intensity staining, and 3=strong intensity staining)
were computed. The area of expression was measured by an imaging
analysis program as described elsewhere (14). Briefly, color images were converted
to gray scale. The expression areas were then located by threshold
adjustment and measured as percentage area of the
expression/field.
Immunogold electron microscopic
studies
For aldehyde blocking, gastric sections were
incubated in 50 mM glycine in phosphate buffer (PBS), pH 7.4. A
section's non-specific binding was consequently blocked in 5%
bovine serum albumin (BSA, 25557; EMS) in PBS, pH 7.4. The sections
were washed with 0.1% BSA in PBS (incubated buffer). The sections
were incubated in polyclonal rabbit anti-AQP-4 (D2408; Santa Cruz
Biotechnology, Inc.) or polyclonal rabbit anti-Mn-SOD (2683863;
Millipore) for 1 h and then incubated in goat anti-rabbit IgG
conjugated with 10 nm gold (G7402-.4ML; Sigma Chemical Co., St.
Louis, MO, USA) for 1 h. After that the sections were rigorously
washed with incubated buffer and distilled water. To enhance gold
signalling, silver enhancement kit (Aurion R-Gent SE-EM kit, 25521;
EMS) was used followed by the manufacture guideline. Finally, the
sections were then stained by uranyl acetate and lead citrate. To
quantify the AQP-4 and SOD expression, gold labelled particle in
gastric cell (n=10 cells/animal) was counted under transmission
electron microscope (HT7700, HITACHI, Ltd.).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism® v.5. The non-parametric t-test was used to
compare the difference in histopathological/ultrastructural changes
between subgroups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Histopathology and electron
microscopy
Gross appearance and SEM of the gastric
mucosa
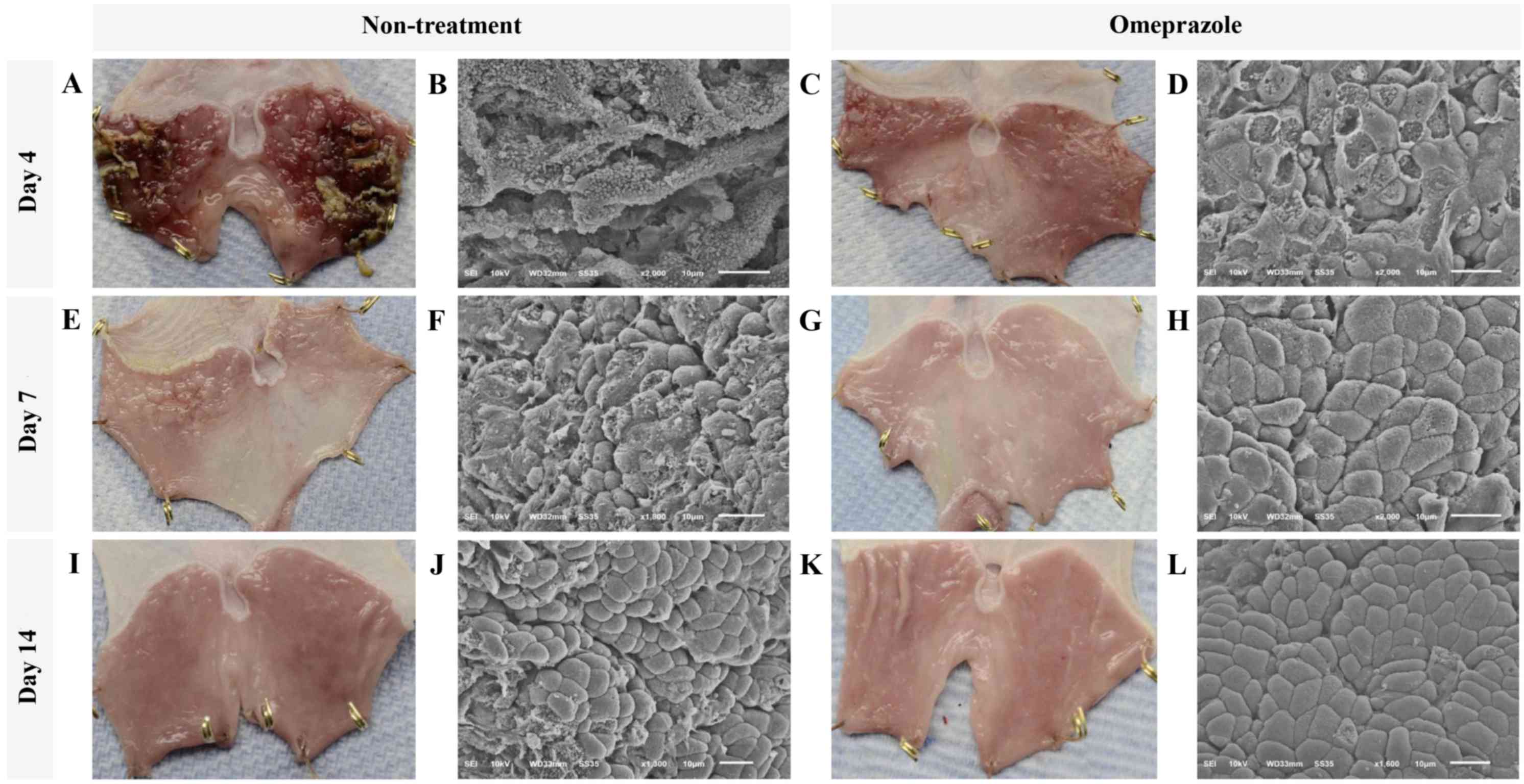
At gross level by 4 days post induction, a single
dose of 5 ml/kg absolute ethanol (Fig.
1) induced severe generalized hemorrhages and necrotizing
membrane deposit on the mucosal surface of glandular stomach in all
rats without treatment whereas only pinpoint hemorrhages were
observed in omeprazole treated rats (Fig. 1A and C). In addition, ultrastructure
of gastric mucosal areas in untreated rats was highly affected as
shown by extensive epithelial sloughing and a completely lost,
stripped and folded membrane, loss of gastric pits, and cellular
debris accumulation (Fig. 1B). In
contrast to the treated rats, epithelial cells were better
preserved. Although many epitheliums were flattened and ruptured in
treated rats, they were not transformed as presented in the
untreated rats. However, the cellular arrangement was not well
formed, especially in gastric pit and cellular interface (Fig. 1D).
The improvement of gross lesions on gastric surface
was rapidly resolved in all rats with or without treatment
(Fig. 1A, E & I and C, G &
K) at 7 and 14 days post induction. Nevertheless, scarce
hemorrhagic areas and irregular membranes remained in all rats
without treatment (Fig. 1E). At 4
days post induction, the gross hemorrhagic area in untreated rats
was significantly higher than that in omeprazole treated rats
(Table I). However, the severity of
bleeding was intensely decreased in both groups at 4 days after
induction. Corresponding to the ultrastructural appearance, the
cellular arrangement was almost completely regenerated by 7 days
post induction in omeprazole treated rats. Moreover, cell to cell
interfaces and gastric pits were clearly localized and almost
reformed (Fig. 1F-J). Meanwhile, the
cellular arrangement of untreated rats was closed to intact at 14
days post induction (Fig. 1L).
 | Table I.The comparison of histopathological,
PGE-2 activity, and ultrastructural changes in glandular stomach of
omeprazole-treated and untreated rats at 4, 7, and 14 days post
gastritis induction. |
Table I.
The comparison of histopathological,
PGE-2 activity, and ultrastructural changes in glandular stomach of
omeprazole-treated and untreated rats at 4, 7, and 14 days post
gastritis induction.
|
| 4 Day PI | 7 Day PI | 14 Day PI |
|---|
|
|
|
|
|
|---|
|
Pathology/parameter | Omeprazole (n=5
rats) | Non-treatment (n=5
rats) | P-value | Omeprazole (n=5
rats) | Non-treatment (n=5
rats) | P-value | Omeprazole (n=5
rats) | Non-treatment (n=5
rats) | P-value |
|---|
| Mucosa |
|
|
|
|
|
|
|
|
|
| Glandular
atrophy/dilatation |
52.5±35.4 |
97.5±35.4 | 0.404 |
0.0±0.0 |
30.0±30.0 | 0.374 |
0.0±0.0 |
0.0±0.0 | 1.000 |
|
Necrosis |
45.5±35.7 |
202.5±70.7 | 0.094 |
0.0±0.0 |
30.0±30.0 | 0.374 |
0.0±0.0 |
0.0±0.0 | 1.000 |
| Tissue
debris |
22.5±14.3 |
150.0±71.4 | 0.131 |
0.0±0.0 |
0.0±0.0 | 1.000 |
0.0±0.0 |
0.0±0.0 | 1.000 |
|
Inflammation |
15.0±15.0 |
262.5±22.5 | 0.000 |
0.0±0.0 |
50.0±50.0 | 0.374 |
0.0±0.0 |
0.0±0.0 | 1.000 |
|
Hemorrhage |
30.0±17.3 |
210.7±71.4 | 0.049 |
0.0±0.0 |
0.0±0.0 | 1.000 |
0.0±0.0 |
0.0±0.0 | 1.000 |
|
Congestion |
77.5±31.1 |
37.5±10.3 | 0.269 |
5.0±2.8 |
30.0±0.0 | 0.001 |
5.0±2.8 |
3.3±1.6 | 1.000 |
| Submucosa |
|
|
|
|
|
|
|
|
|
|
Edema |
215.0±31.22 |
187.5±71.8 | 0.737 |
0.0±0.0 |
30.0±30.0 | 0.394 |
0.0±0.0 |
0.0±0.0 | 0.589 |
|
Inflammation |
250.0±50.0 |
225.0±75.0 | 0.791 |
0.0±0.0 |
163.3±74.4 | 0.093 |
0.0±0.0 |
0.0±0.0 | 1.000 |
|
Hemorrhage |
97.5±59.2 |
127.5±43.0 | 0.696 |
0.0±0.0 |
0.0±0.0 | 1.000 |
0.0±0.0 |
0.0±0.0 | 1.000 |
|
Congestion |
22.5±16.0 |
22.5±7.5 | 1.000 |
3.3±1.6 |
40.0±10.0 | 0.022 |
1.6±1.6 |
1.6±1.6 | 1.000 |
| Fatty
degeneration |
82.5±51.0 |
67.5±25.6 | 0.802 |
0.0±0.0 |
16.6±8.8 | 0.132 |
0.0±0.0 |
0.0±0.0 | 1.000 |
| Muscularis |
|
|
|
|
|
|
|
|
|
|
Vacuolated degeneration |
90.0±37.6 |
87.5±55.5 | 0.972 |
0.0±0.0 |
90.0±56.8 | 0.189 |
0.0±0.0 |
0.0±0.0 | 1.000 |
|
Inflammation |
0.0±0.0 |
22.5±13.1 | 0.138 |
0.0±0.0 |
0.0±0.0 | 1.000 |
0.0±0.0 |
0.0±0.0 | 1.000 |
|
Hemorrhage |
0.0±0.0 |
22.5±14.3 | 0.168 |
0.0±0.0 |
0.0±0.0 | 1.000 |
0.0±0.0 |
0.0±0.0 | 1.000 |
|
Congestion |
0.0±0.0 |
7.5±2.5 | 0.024 |
0.0±0.0 |
6.6±1.6 | 0.016 |
0.0±0.0 |
0.0±0.0 | 1.000 |
|
Grandular PGE-2 (pg/ml) |
2,277.1±291.7 |
1,643.1±89.89 | 0.047 |
3,580.4±714.7 |
1,263.0±20.44 | 0.017 |
1,408.7±13.41 |
986.5±19.21 | 0.000 |
| Gross
hemorrhagic area |
14.0±7.2 |
28.2±10.2 | 0.040 |
0.1±0.1 |
1.2±0.5 | 0.985 |
0.0±0.0 |
0.0±0.0 | 1.000 |
|
Defected RER in chief
cells |
5.2±0.6 |
174.0±8.7 | 0.000 |
5.6±0.8 |
250.0±15.1 | 0.000 |
6.8±0.8 |
6.4±1.0 | 0.899 |
|
Defected RER in parietal
cells |
5.7±0.8 |
4.7±1.0 | 0.483 |
6.2.0±1.1 |
5.5±0.6 | 0.580 |
6.2±1.2 |
6.5±1.9 | 0.890 |
|
Swelling Mito in chief
cells |
24.0±6.1 |
26.0±7.0 | 0.796 |
87.3±2.8 |
84.6±9.2 | 0.654 |
15.0±1.1 |
11.0±1.1 | 0.07 |
|
Swelling Mito in parietal
cells |
36.3±2.9 |
24.0±6.4 | 0.156 |
47.3±2.7 |
28.3±21.6 | 0.840 |
44.0±4.7 |
32.6±6.9 | 0.248 |
Pathological changes in the glandular
stomach
Pathological changes of ethanol-induced gastritis in
each gastric layer are shown in Table
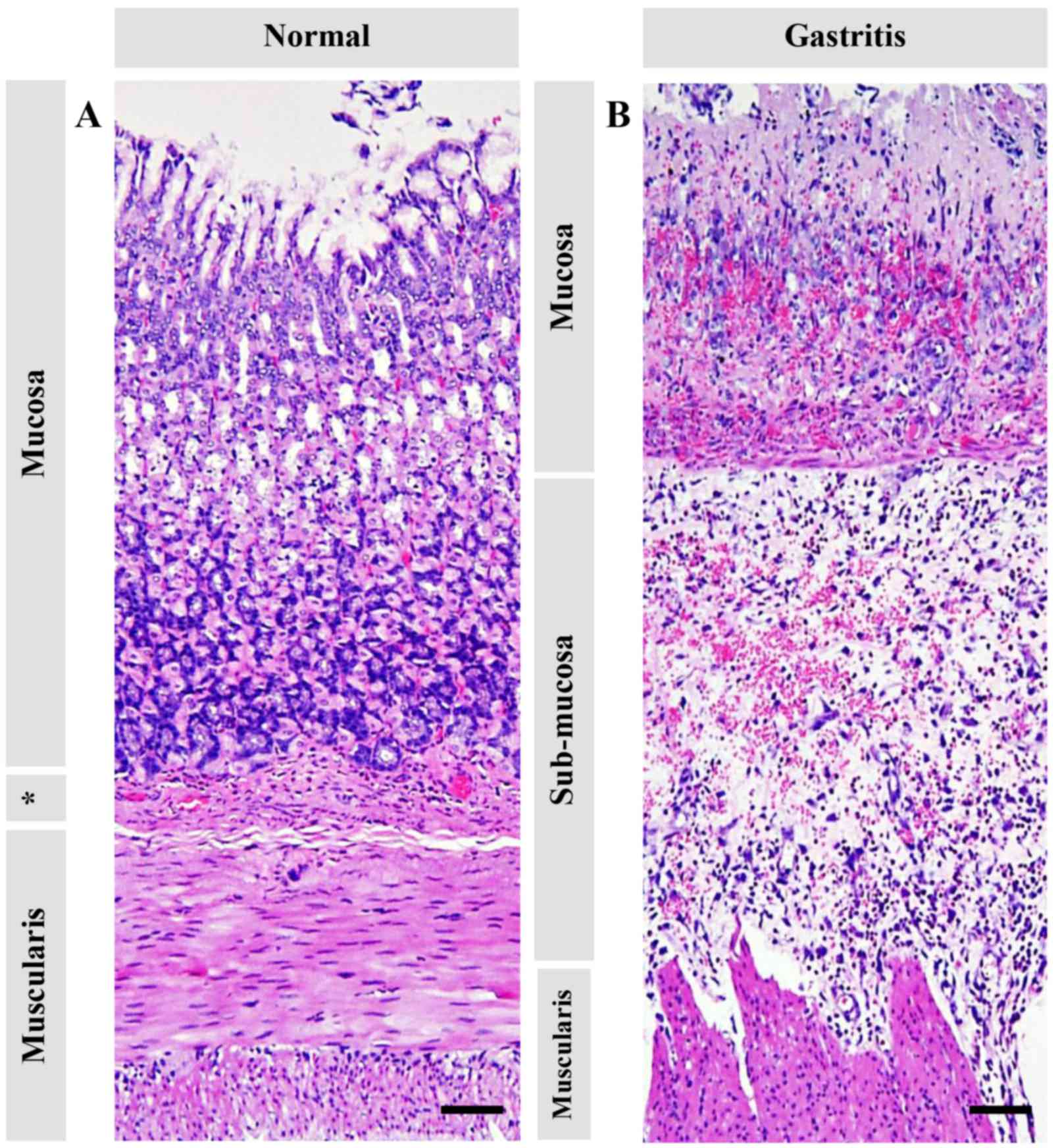
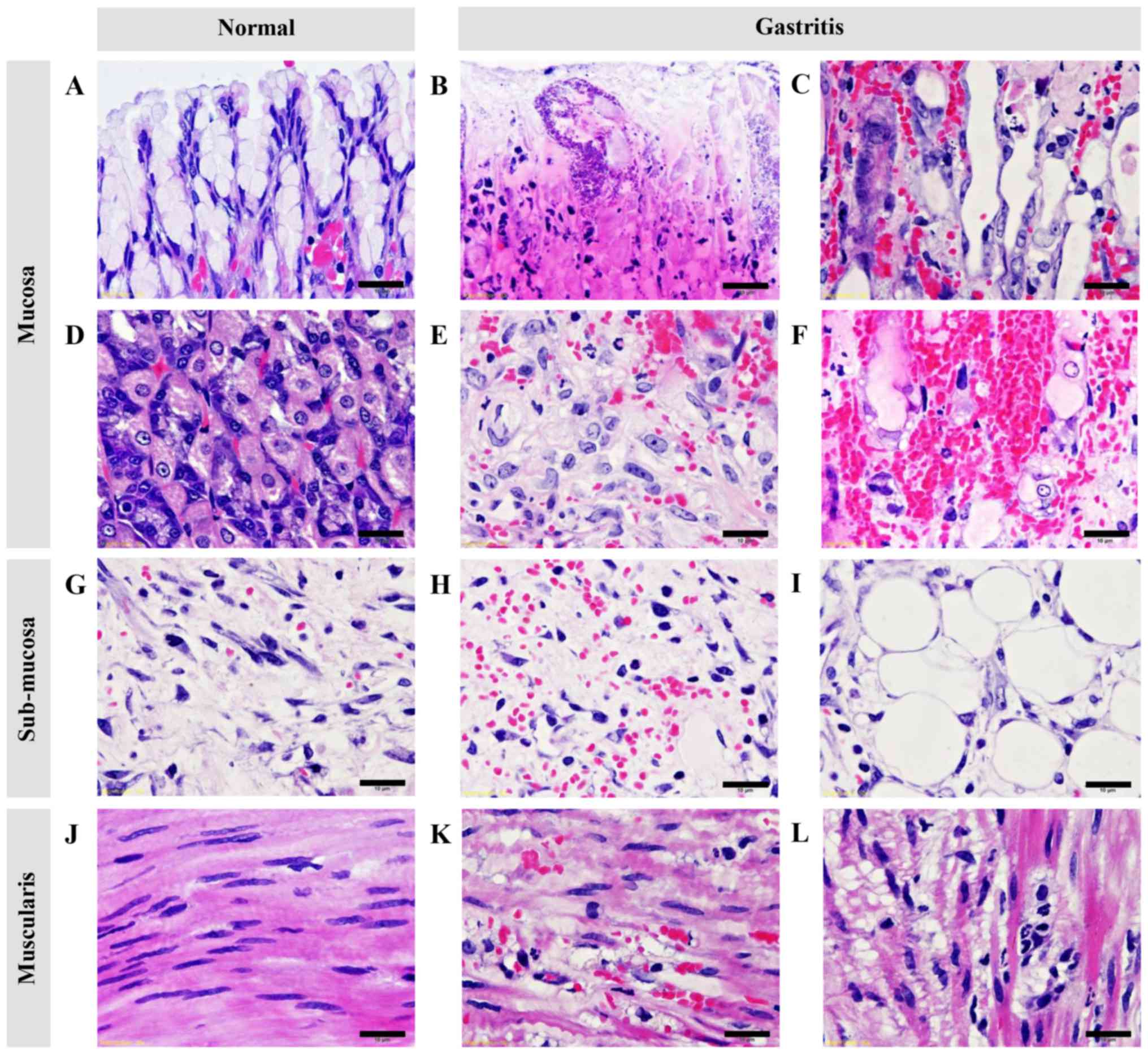
I, and Figs. 2 and 3. The mucosal layer exhibited a shortening
in thickness (Fig. 2), severe
hemorrhage (Fig. 3F), acute
inflammation (Fig. 3B),
cellular-swelling, -degeneration, and -necrosis (Fig. 3B), and glandular atrophy/dilattion
(Fig. 3C). Predominant changes in
submucosal layer composed of edema, inflammation, hemorrhage
(Figs. 2 and 3H), and fatty degeneration (Fig. 3I). Interestingly, vacuolated
degeneration was found in the muscular layer accompanied by
inflammation (Fig. 3K and L).
Similar to the other studies (7,13), at 4
days post induction, mucosal inflammation and hemorrhage in
untreated rats were significantly higher than those in treated
rats. Although glandular atrophy/dilatation and necrosis in mucosal
layer were similar in both groups, untreated rats showed a higher
trended than omeprazole treated rats. Subsequently, at 7 and 14
days post induction, the main histopathological changes in each
gastric layer were similar in both groups.
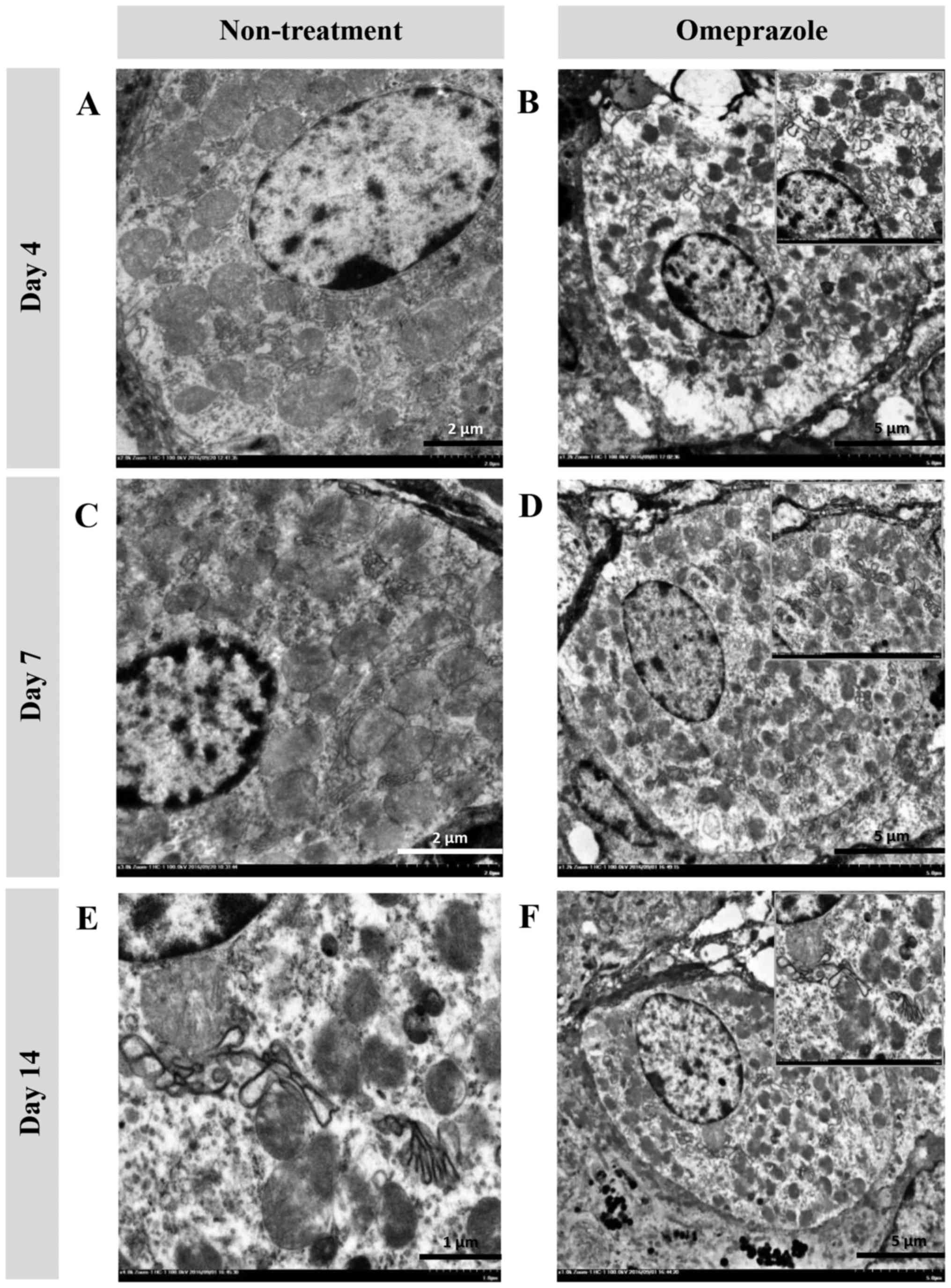
Ultrastructural changes (TEM) in chief and
parietal cells
In the present study, the ultrastructural changes of
cells were examined using TEM (Fig.
4). RER changes in chief cells were mostly found in untreated
rats at 4 and 7 days post induction (Fig. 4A, B, E and F), which were
characterized by a dark electron-dense crinkled-like structure with
dilatation and enlargement. The highest severity of RER changes was
presented at 7 days post induction, which was accompanied by
mitochondrial swelling (Fig. 4E and
F). At 14 days post induction, the RER was completely
regenerated as shown by the sheath-like structure in all groups of
rats (Fig. 4I-L). In addition,
H-score demonstrated that untreated rats had a number of defected
RERs in chief cells at 4 and 7 days post induction with increasing
severity (Table I). Unlike chief
cells, defected RERs in parietal cells were similarly low in both
groups of rats (Table I). It
appeared that parietal cells were more preserved than chief cells
in ethanol-induced gastritis as shown in the Fig. 5.
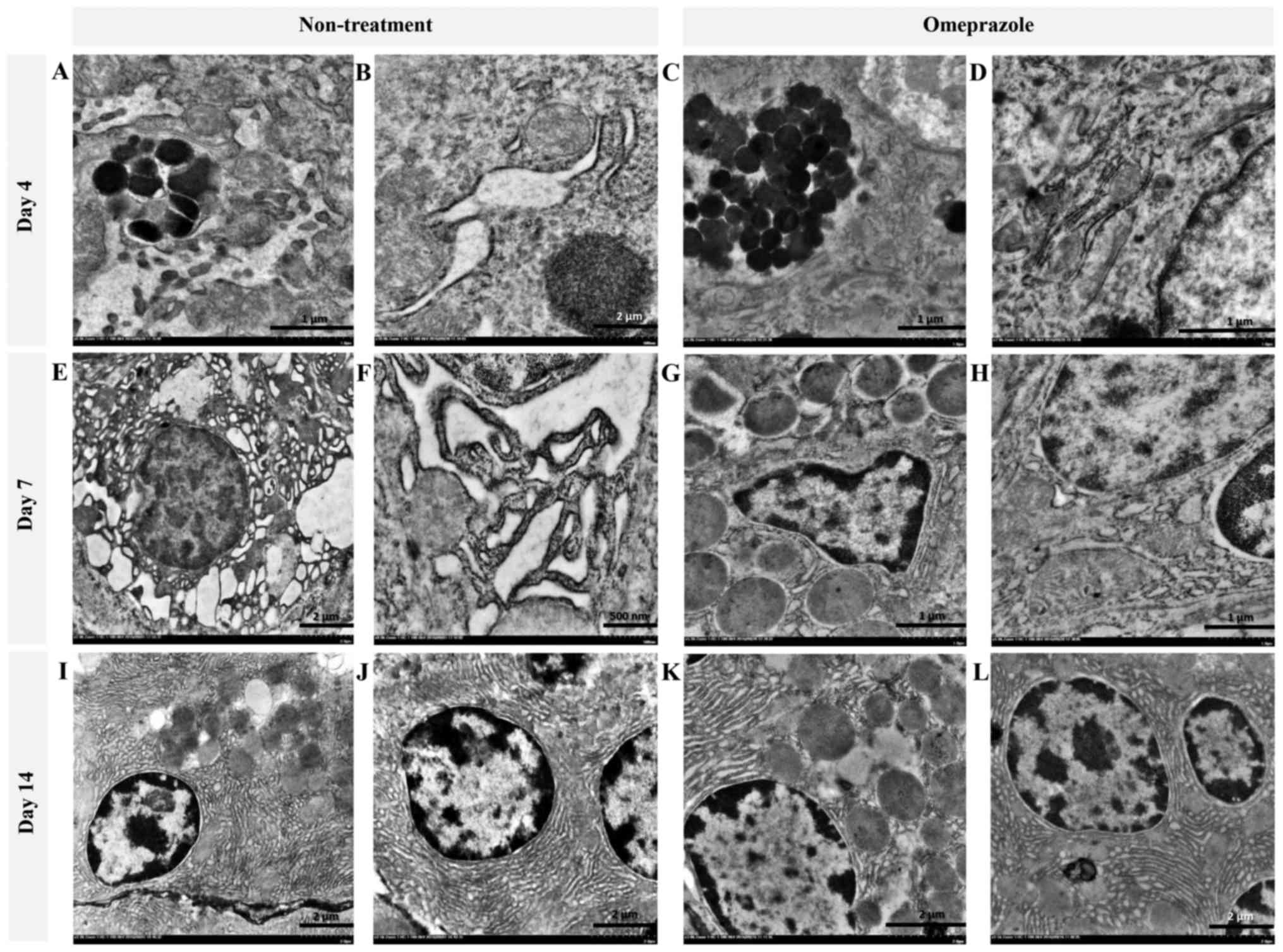 | Figure 4.Ultrastructure changes of chief cells
in omeprazole-treated rats compared with untreated rats. RER
alterations were found in untreated rats as indicated by the
presence of cytoplasmic vacuolation and moderate RER dilatation (A
and B) while the RER architecture in omeprazole-treated rats was
preserved (C, D, G, H, K and L). A severe grade of peculiar dark,
dense and dilated RERs accompanied by degenerative mitochondria
were observed in non-treated rats at 7 days post-induction (E and
F). RER integrity was preserved in rats with or without omeprazole
treatment at 14 days post-induction (I-L). B, E and I-L scale bar,
2 µm; F scale bar, 500 nm; A, C, D, G and H, scale bar, 1 µm. |
PGE-2 activity
PGE-2 activity is described in Table I. The results revealed that
omeprazole treatment induced a significantly higher level of
gastric PGE-2 compared with untreated rats at all time points.
PGE-2 activity was increased over time until 7 days post induction
and then declined at 14 days post induction. Unfortunately, in the
present study, there was no correlation of the level of PGE-2
activity with histopathological or ultrastructural changes.
Immunohistochemistry
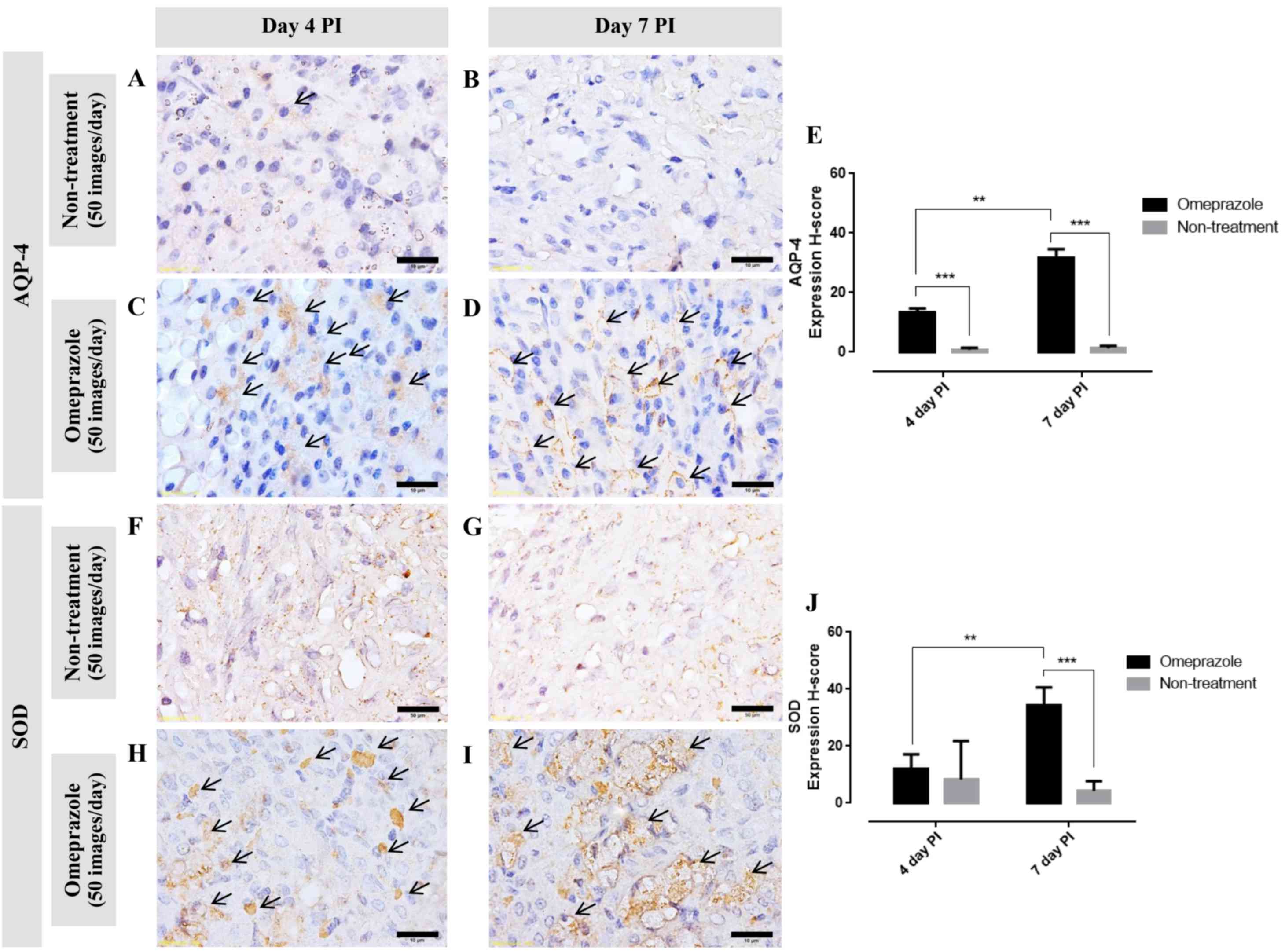
AQP-4 expression in gastric mucosa
Generally, AQP-4 is expressed on the baso-lateral
area of the parietal cells. In this study, at 4 and 7 days post
induction, the AQP-4 expression score in omeprazole treated rats
was significantly higher than that in untreated rats (Fig. 6A-E). Furthermore, the expression
increased over time. Concerning to the localized area, at 4 days
post induction, AQP-4 was labeled in the cytoplasm which located on
either side of the cell (Fig. 6C
arrow). In contrast to 7 days post induction, AQP-4 was labeled on
the baso-lateral cell as presented by ‘fried egg appearance’
(Fig. 6D arrow). These labeling
patterns are associated with the regeneration processes of the
parietal cells.
SOD expression in gastric mucosa
Similar to the AQP-4 expression score, at 7 days
post induction, SOD expression in omeprazole treated rats was
significantly higher than that in untreated rats (Fig. 6F-J). Up-regulation of SOD was
depended on the post induction time. The positive
immuno-localization of SOD was presented on the cytoplasm of
gastric mucosal cells.
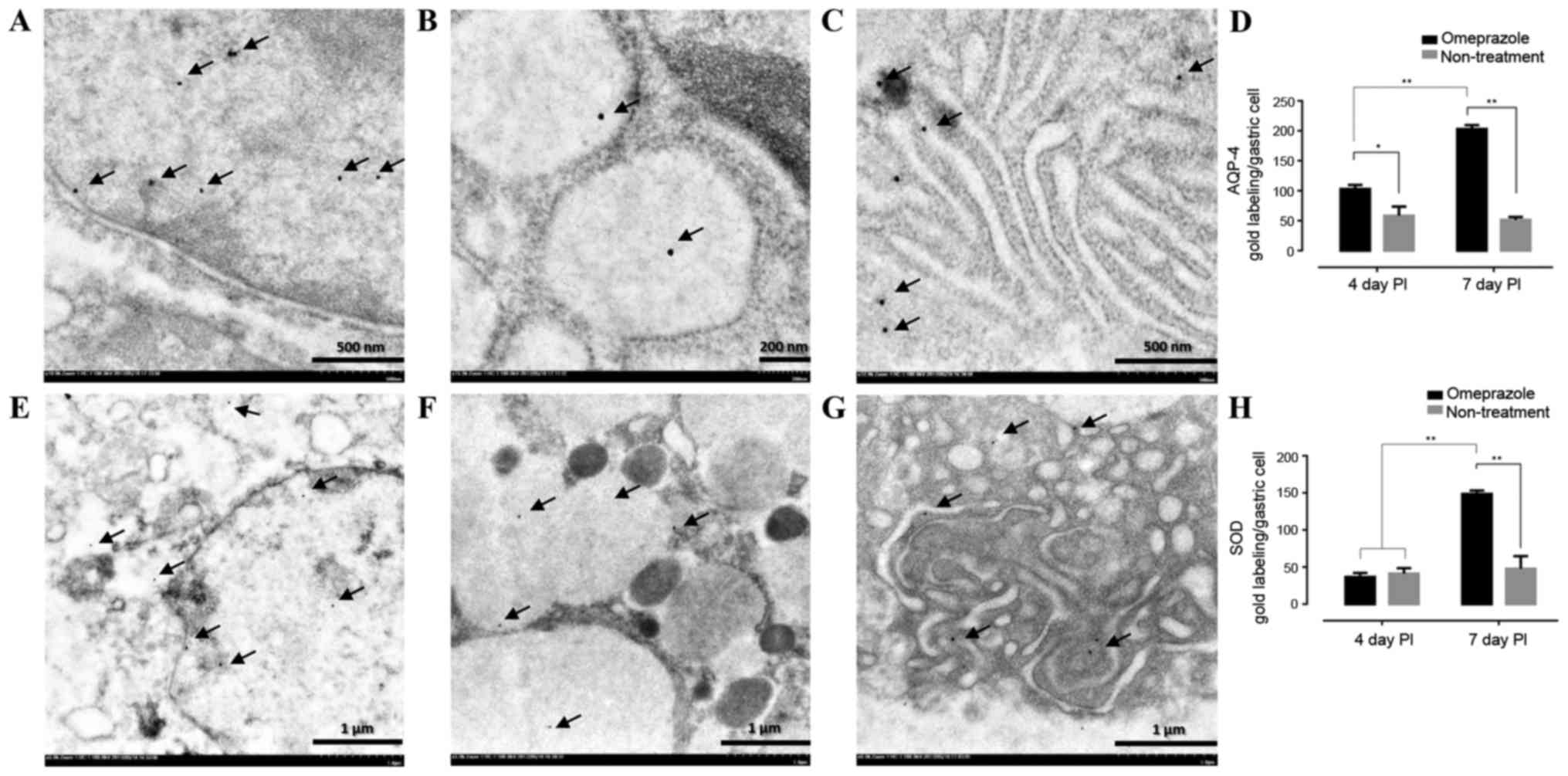
AQP-4 and SOD gold labeling in gastric
mucosa
To confirm the levels of AQP-4 and SOD expression in
the gastric cells, immunoelectron microscopic study was performed.
The results revealed that both AQP-4 and SOD were labeled in the
nucleus, cytoplasm and rough endoplasmic reticulum (Fig. 7A-C and E-G). Like immunohistochemical
staining, the treated rats had significant level of AQP-4 when
compared to untreated rats in all observation periods while a level
of SOD in treated rats was up-regulated on day 7 post induction
(Fig. 7D and H).
Discussion
The balance of protective and aggressive factors in
the gastric mucosal barrier is an important consideration in
gastrointestinal diseases (4).
Gastric acid, pepsin, and external stimuli are known aggravating
factors of gastric mucosal cells, while microcirculatory system,
HCO3−, prostaglandins, epidermal growth
factor synthesis, and epithelial cell reconstitution are
gastro-protective factors that maintain integrity of the gastric
mucosal layer. Obviously, there are several viewpoints in
association with the effect of ethanol on gastric mucosal cells.
However, the main pathogenesis of ethanol-induced gastritis is
vascular damage followed by mucosal cell-hypoxia, -degeneration,
and -necrosis, respectively. The decline of anti-oxidant level and
over production of oxygen free radicals especially super-oxide and
hydrogen peroxide, are also link to ethanol-induced gastric injury
and play a crucial role to further induce gastric inflammatory
response. In addition, ethanol itself destroys gastric mucosal
cells and causes them to slough. Then, the remaining cells and
submucosal layer are directly exposed to luminal gastric acid.
Submucosal edema is a consequence of the increase in vascular
permeability, while vascular congestion is exacerbated by alcohol
penetration. According to the histopathological and
immunohistochemical results, omeprazole attenuated the severity of
hemorrhage and inflammation on gastric mucosa (Table I) with upregulation of anti-oxidative
stress marker, SOD (Figs. 6 and
7), which contributed to the mucosal
protective effect of omeprazole in ethanol-induced gastritis.
Within a past decade, although several products have
been considered as candidates to alleviate gastrointestinal
diseases according to the mucosal lesion index, mucous production,
level of PGE-2, and specific cytokines (15), no studies have described improvement
of RER integrity in ethanol-induced gastritis because RER
alterations are also crucially involved in the pathogenesis of
several gastrointestinal disturbances. SEM usually employed to
evaluate mucosal surface cells affected by ethanol, which reveals a
flattened or swollen mucosal epithelium and irregularly gastric
pits in animal models (16). The
gastric mucosal epithelium in human with chronic gastritis presents
the swelling, vacuolar degeneration, ribosome dissociation,
dilatation of the RER and Golgi's apparatus together with
mitochondrial swelling in chief cells (17,18). In
the present study, we demonstrated additional histopathological
changes in ethanol-induced gastritis other than those in previous
reports, including fatty degeneration in submucosal layer and
vacuolated degeneration in muscular layer (Table I and Fig.
3I and L). The results also presented that omeprazole improved
cellular architecture in the stomach after 4 days post induction.
In treatment group, the gastric cells were almost recovered and
complete healed on 7 and 14 days post induction, respectively.
However, at ultrastructural level, some defects on RER and
mitochondria were still observed. It has been appeared that ethanol
and luminal gastric acid penetrated deeply to damage all of the
epitheliums from mucosal to muscularis layer. We also examined
alterations in chief and parietal cells induced by ethanol.
Interestingly, we firstly confirmed that ethanol-induced gastritis
caused RER alterations in chief cells indicated by a number of
large, dilated, and fragmented RERs as shown in Fig. 4A, B, E and F, and omeprazole
preserves their integrity in relation to its anti-oxidative and
anti-inflammatory effects.
Although, currently it is well described that
omeprazole, which is prescribed to suppression acid secretion in
the stomach, preserves the gastric mucosal layer, there is no clear
evidence that omeprazole reduces ER stress in ethanol-induced
gastritis. A recent study suggested that ethanol exacerbates ER
stress responses and promotes hepatic, cardiac, and pancreatic
diseases through apoptotic pathways (19) however, ethanol-induced ER stress in
the stomach is still controversial because of the lack of reports
(16). The ER is a protein factory
for protein synthesis and folding, after which synthesized proteins
are transported to the Golgi. ER stress is caused by retention of
proteins in the ER because of unfolding (20) or the so called ‘unfolded protein
response’, leading to protein degradation and initiation of
cellular apoptosis (21). Moreover,
ER stress and impaired protein folding cause up-regulation of
reactive oxygen species production (22). A Collapsed RER is closely connected
to misfolded protein as shown by our results. It has been
postulated that omeprazole may reduce ER stress. Another proposed
mechanism for ethanol-induced gastritis is energetic mitochondrial
disturbance by oxidative stress (23). In the present study, ultrastructural
finding demonstrated that mitochondrial swelling was accompanied by
RER alterations (Table I and
Fig. 4E and F). This result supports
that ER stress similarly triggers mitochondrial alterations and
dysfunction, and especially apoptosis (24) with an increase in cytosolic calcium
(25). However this mechanistic
detail needs to be further studies.
There are several markers used to study parietal
cells morphology and functions depending on their roles, AQP-4,
water channel membrane protein in gastric acid physiology, is
relates to the degree of regenerating parietal cells (10). Early immature parietal cells weakly
express AQP-4, which gradually increases upon maturation.
Furthermore, because of its function, AQP-4 is typically used as an
improvement criterion in studies of gastric mucosal injury
(26,27). In the present study, omeprazole
maintained the integrity and induced re-epithelialization of the
parietal cell as indicated by a number of cell positive for AQP-4
at 4 and 7 days post gastritis induction with increasing trends
(Fig. 6A, D and E) as confirmed by
immunogold electron microscopic study (Fig. 7). The results indicated that the
expression was located throughout of cell especially in the nucleus
and rough endoplasmic reticulum. Apart from regenerative aspect,
the present of AQP-4 is associated with acid suppression by proton
pump inhibitors (11,12). Moreover, inflammatory condition
induced by histamine results gastric AQP-4 rearrangement or
down-regulation (28). Therefore,
other than the re-epithelializing effect, omeprazole exerts
anti-acidic and anti-inflammatory effects in the gastric
environment. Unfortunately, the correlation between PGE-2 activity
to histopathological and ultrastructural changes was not observed
in the present study. This limitation may relate to the selection
of sampling date post induction due to the difference recovering
rat e of gastric mucosal between treated and non-treated rats. The
earlier detection, such as on 3, 5 and 7 day, may give better
correlation.
In conclusion, deformation of the RER in chief cells
is caused by ethanol-induced gastritis. Omeprazole modulates
mucosal cells injury affected by ethanol as shown by the
improvement of the RER architecture in chief cells and the
enhancement of parietal cell regeneration together with exerting
gastric anti-inflammatory and anti-oxidative effects.
Acknowledgements
The present study was supported by the National
Laboratory Animal Center, Faculty of Tropical Medicine, Faculty of
Medicine Siriraj Hospital, Mahidol University, and National
Research Council of Thailand (NRCT) under the project ‘The Added
Value of Herbs Used for Cooking in Households’.
References
|
1
|
Schneider AS and Szanto PA: Board Review
SeriesPathology. Fifth Edition. Lippincott William & Wilkins;
Philadelphia, PA: 2013
|
|
2
|
Al-Humayed SM, Mohamed-Elbagir AK,
Al-Wabel AA and Argobi YA: The changing pattern of upper
gastro-intestinal lesions in southern Saudi Arabia: An endoscopic
study. Saudi J Gastroenterol. 16:35–37. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lou LX, Geng B, Yu F, Zhang J, Pan CS,
Chen L, Qi YF, Ke Y, Wang X and Tang CS: Endoplasmic reticulum
stress response is involved in the pathogenesis of stress induced
gastric lesions in rats. Life Sci. 79:1856–1864. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Suzuki H, Nishizawa T, Tsugawa H, Mogami S
and Hibi T: Roles of oxidative stress in stomach disorders. J Clin
Biochem Nutr. 50:35–39. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zarebska A, Sekita-Krzak J, Hernik D,
Matusiewicz J and Czerny K: Histological examinations of chief and
parietal cells of the rat gastric glands after experimental
administration of cephalexin and ethanol. Ann Univ Mariae Curie
Sklodowska Med. 59:99–104. 2004.PubMed/NCBI
|
|
6
|
Karam SM: A focus on parietal cells as a
renewing cell population. World J Gastroenterol. 16:538–546. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Teschke R, Wolff A, Frenzel C, Eickhoff A
and Schulze J: Herbal traditional Chinese medicine and its evidence
base in gastrointestinal disorders. World J Gastroenterol.
21:4466–4490. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zaidi SF, Muhammad JS, Usmanghani K and
Sugiyama T: Review: Pharmacological ins and outs of medicinal
plants against Helicobacter pylori: A review. Pak J Pharm Sci. 28 3
Suppl:S1171–S1176. 2015.
|
|
9
|
Chanchal SK, Mahajan UB, Siddharth S,
Reddy N, Goyal SN, Patil PH, Bommanahalli BP, Kundu CN, Patil CR
and Ojha S: In vivo and in vitro protective effects of omeprazole
against neuropathic pain. Sci Rep. 6:300072016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Matsuoka T, Kobayashi M, Sugimoto T and
Araki K: An immunocytochemical study of regeneration of gastric
epithelia in rat experimental ulcers. Med Mol Morphol. 38:233–242.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fukuhara S, Matsuzaki J, Tsugawa H,
Masaoka T, Miyoshi S, Mori H, Fukushima Y, Yasui M, Kanai T and
Suzuki H: Mucosal expression of aquaporin-4 in the stomach of
histamine type 2 receptor knockout mice and Helicobacter
pylori-infected mice. J Gastroenterol Hepatol. 4 29 Suppl:S53–S59.
2014. View Article : Google Scholar
|
|
12
|
Matsuzaki J, Suzuki H, Minegishi Y, Sugai
E, Tsugawa H, Yasui M and Hibi T: Acid suppression by proton pump
inhibitors enhances aquaporin-4 and KCNQ1 expression in gastric
fundic parietal cells in mouse. Dig Dis Sci. 55:3339–3348. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rahim NA, Hassandarvish P, Golbabapour S,
Ismail S, Tayyab S and Abdulla MA: Gastroprotective effect of
ethanolic extract of Curcuma xanthorrhiza leaf against
ethanol-induced gastric mucosal lesions in Sprague-Dawley rats.
Biomed Res Int. 2014:4164092014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ampawong S and Aramwit P: Tolerogenic
responses of CD206+, CD83+, FOXP3+, and CTLA-4 to sericin/polyvinyl
alcohol/glycerin scaffolds relevant to IL-33 and HSP60 activity.
Histol Histopathol. 31:1011–1027. 2016.PubMed/NCBI
|
|
15
|
Wang QS, Zhu XN, Jiang HL, Wang GF and Cui
YL: Protective effects of alginate-chitosan microspheres loaded
with alkaloids from Coptis chinensis Franch. and Evodia rutaecarpa
(Juss.) Benth. (Zuojin Pill) against ethanol-induced acute gastric
mucosal injury in rats. Drug Des Devel Ther. 9:6151–6165.
2015.PubMed/NCBI
|
|
16
|
Cho KR, Kwon KY and Chang ES:
Ultrastructural study of alcohol-induced gastric mucosal change of
rat. Korean J Pathol. 27:362–370. 1993.
|
|
17
|
Zhang B and Jin R: Ultrastructural
characteristics of gastric mucosal epithelial cells of AIDS
patients complicated with chronic gastritis. Zhonghua Shi Yan He
Lin Chuang Bing Du Xue Za Zhi. 21:17–19. 2007.(In Chinese).
PubMed/NCBI
|
|
18
|
Zhang ZL, Bu JK and Zhao JX:
Ultrastructural observation of the gastric mucosa in chronic
gastritis patients treated by traditional Chinese medicine. World J
Gastroenterol. 3:185–188. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ji C: New insights into the pathogenesis
of alcohol-induced ER stress and liver diseases. Int J Hepatol.
2014:5137872014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang K and Kaufman RJ: From
endoplasmic-reticulum stress to the inflammatory response. Nature.
454:455–462. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim I, Xu W and Reed JC: Cell death and
endoplasmic reticulum stress: Disease relevance and therapeutic
opportunities. Nat Rev Drug Discov. 7:1013–1030. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Malhotra JD, Miao H, Zhang K, Wolfson A,
Pennathur S, Pipe SW and Kaufman RJ: Antioxidants reduce
endoplasmic reticulum stress and improve protein secretion. Proc
Natl Acad Sci USA. 105:pp. 18525–18530. 2008, View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pan JS, He SZ, Xu HZ, Zhan XJ, Yang XN,
Xiao HM, Shi HX and Ren JL: Oxidative stress disturbs energy
metabolism of mitochondria in ethanol-induced gastric mucosa
injury. World J Gastroenterol. 14:5857–5867. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sano R and Reed JC: ER stress-induced cell
death mechanisms. Biochim Biophys Acta. 1833:3460–3470. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Apostolova N, Gomez-Sucerquia LJ, Alegre
F, Funes HA, Victor VM, Barrachina MD, Blas-Garcia A and Esplugues
JV: ER stress in human hepatic cells treated with Efavirenz:
Mitochondria again. J Hepatol. 59:780–789. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen GX, Lao SX and Huang ZX: Effect of
Chinese herbs on expression of aquaporin 3,4 gene in gastric mucosa
of patients with Pi-Wei Damp-Heat syndrome. Zhongguo Zhong Xi Yi
Jie He Za Zhi. 25:199–202. 2005.(In Chinese). PubMed/NCBI
|
|
27
|
Feng G, Xu X, Wang Q, Liu Z, Li Z and Liu
G: The protective effects of calcitonin gene-related peptide on
gastric mucosa injury after cerebral ischemia reperfusion in rats.
Regul Pept. 160:121–128. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Carmosino M, Procino G, Nicchia GP,
Mannucci R, Verbavatz JM, Gobin R, Svelto M and Valenti G:
Histamine treatment induces rearrangements of orthogonal arrays of
particles (OAPs) in human AQP4-expressing gastric cells. J Cell
Biol. 154:1235–1243. 2001. View Article : Google Scholar : PubMed/NCBI
|