Introduction
Liver fibrosis is a common consequence of chronic
liver diseases, including chronic hepatitis B virus infection,
chronic hepatitis C virus infection, alcoholic liver disease and
non-alcoholic fatty liver disease (1–4). During
liver fibrogenesis, hepatic stellate cells (HSCs) are activated,
there is an excessive accumulation of extracellular matrix in the
liver and there is an increased inflammatory response (5). If left untreated or managed
inappropriately, liver fibrosis may rapidly progress to advanced
liver fibrosis or cirrhosis, which may result in patients requiring
an orthotopic liver transplantation (1–3). Indeed,
liver transplantation is currently the only available treatment
option for patients diagnosed with the end-stage liver fibrosis or
cirrhosis. However, a number of disadvantages, including a shortage
of liver transplant donors, a high cost and transplant rejection
following liver transplantation, have largely limited its clinical
application. Therefore, novel therapeutic approaches are required
for the treatment of liver fibrosis and to improve care for such
patients.
Mesenchymal stem cell (MSC) transplantation
(6–10) has emerged as an alternative and novel
method of treating liver fibrosis regardless of the underlying
cause (11). MSCs can be isolated
from a wide range of tissue sources, including bone marrow, adipose
tissue, umbilical cord/cord blood and dental pulp (12,13). Of
these MSCs, adipose tissue-derived MSCs (AD-MSCs) and bone
marrow-derived MSCs (BM-MSCs) are the most easily accessible and
have been extensively studied. Previous studies have demonstrated
that MSCs possess immunomodulatory properties, the ability to
differentiate into hepatocytes to replace damaged hepatocytes, and
the capacity to suppress HSC activation and promote HSC apoptosis
(14,15). Several studies, including one by the
current authors (16), revealed that
BM-MSCs and AD-MSCs are capable of attenuating liver fibrosis using
multiple molecular mechanisms by which HSC activation is inhibited,
activated HSC proliferation is reduced, the inflammatory response
is weakened and hepatocyte regeneration is enhanced (15–19).
Indeed, AD-MSCs and BM-MSCs exhibit similar surface molecular
markers and differentiation abilities; however, AD-MSCs are easier
to obtain and have higher proliferation rates than BM-MSCs
(16).
To the best of our knowledge, AD-MSCs and BM-MSCs
have not been compared with regards to their efficacy in the
treatment of liver fibrosis. It remains unclear whether AD-MSCs and
BM-MSCs exhibit equal anti-liver fibrosis and anti-inflammatory
capabilities. The present study expanded upon the results of
previous studies (16,20) and performed comparative and
comprehensive studies into the effects of BM-MSCs and AD-MSCs on
the activation, proliferation and apoptosis of HSCs in a cell
co-culture system, as well as their anti-inflammatory and
anti-fibrotic properties in a CCl4-induced rat liver
fibrosis model. Furthermore, the secretion of a select group of
cytokines by BM-MSCs and AD-MSCs into the culture medium of a
co-culture system was analyzed.
Materials and methods
Animals
A total of 30 Sprague-Dawley (SD) rats, aged 6 weeks
old and weighing 200-250 g were obtained from the Animal
Experimental Center of Wenzhou Medical College (Wenzhou, China). Of
these, 5 rats were kept for one week until isolation of BM-MSCs and
AD-MSCs, whereas 24 were also acclimatized for one week before they
were used for induction of the CCl4-induced liver
fibrotic rat model. Rats were maintained under conditions included
in the animal care and research protocol, which was approved by the
Animal Care and Use Committee of Wenzhou Medical College. The
current study complied with the guidelines for the Care and Use of
Laboratory Animals by the National Institutes of Health as
described previously (16).
Culture of buffalo rat liver (BRL)
cells and rat HSCs
The BRL-3A cell line was provided by the
Experimental Surgical Center of Wenzhou Medical College (Wenzhou,
China) and originally purchased from Procell Life Science Co. Ltd
(Wuhan, China). BRL cells were incubated in Dulbecco's modified
Eagle medium (DMEM; HyClone; GE Healthcare, Chicago, IL, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) in a humidified atmosphere with
5% CO2 at 37°C. Rat primary HSCs were isolated from a
male SD rat following a previously described protocol (16), and cultured in DMEM supplemented with
10% FBS at 37°C in 5% CO2 for ~5–7 days.
Preparation and culture of BM-MSCs and
AD-MSCs
Fresh bone marrow and subcutaneous adipose tissues
were obtained from male SD rats and used to isolate BM-MSCs and
AD-MSCs. To prepare BM-MSCs, each end of the femur and tibia was
cut to expose the marrow cavity, which was then washed with PBS at
least three times. Fresh bone marrow was collected and centrifuged
at 200 × g at room temperature (RT) for 10 min. Pelleted cells were
resuspended in DMEM supplemented with 10% FBS and centrifuged at
200 × g at RT for 10 min. AD-MSCs were prepared by enzymatic
digestion, as reported previously (16). Briefly, 1–2 mg fresh adipose tissue
was harvested from a male SD rat and washed with PBS. Tissues were
digested with 1 mg/ml collagenase type IV (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) at 37°C for 1 h. The suspension was then
centrifuged at 200 × g at RT for 10 min. Pellets were subsequently
resuspended in DMEM supplemented with 10% FBS and cultured in a
humidified atmosphere with 5% CO2 at 37°C. The culture
medium was changed every 2–3 days and the cells were passaged at
80% confluence. At passages 3–5, BM-MSCs and AD-MSCs were subjected
to analysis of specific surface markers including cluster of
differentiation (CD) 90, CD29 and CD45 by flow cytometry, as
previously described (16). The
proliferation and purity of the two types of MSCs were examined
according to the instructions in the Trypan blue staining kit (cat.
no. C0011-1; Beyotime Institute of Biotechnology, Haimen,
China).
Co-culture of MSCs and HSCs
For co-culture of MSCs and HSCs, third-generation
bone marrow- or AD-MSCs at a density of 3×104 cells/well
were seeded onto the upper layer of Transwells (EMD Millipore,
Billerica, MA, USA), while 3×104 HSCs/well were cultured
in the basal compartment of a 6-well cell culture plate.
3×104 of BRLs/well were seeded instead of MSCs onto the
upper layer of Transwells as a negative control and no cells were
seeded onto the upper layer to serve as a blank control. All cells
were maintained for 72 h in DMEM supplemented with 10% FBS in a
humidified atmosphere with 5% CO2 at 37°C.
Cell proliferation assays
The cell counting kit-8 (CCK-8) (Dojindo Molecular
Technologies, Inc. Kumamoto, Japan) was used to determine cell
proliferation. In brief, 3×104 cells/well of BM-MSCs or
AD-MSCs were co-cultured with HSCs at 3×104 cells/well
for 72 h, while the same amount of BRLs/well were seeded instead of
MSCs as negative control and no cells were seeded as a blank
control. Similarly as described above, under co-culture of MSCs and
HSCs, all cells were maintained for 72 h in DMEM supplemented with
10% FBS in a humidified atmosphere with 5% of CO2 at
37°C. A total of 80 µl CCK-8 reagent was added to each well
containing HSCs in the basal compartment of a 6-well cell culture
plate. Cells were incubated for an additional 2 h and absorbance
was measured using a microplate reader at a wavelength of 450
nm.
Flow cytometric analysis of cells
undergoing apoptosis
Following co-culture of HSCs with BM-MSCs or AD-MSCs
for 72 h, cells were harvested and collected. Cells were washed
twice with PBS and resuspended in 450 µl binding buffer (BD
Biosciences, Franklin Lakes, NJ, USA). Samples were then incubated
with 5 µl Annexin V fluorescein isothiocyanate in the Annexin
V-FITC kit (BD Biosciences) according to the manufacturer's
instructions for 15 min at 4°C and flow cytometric analysis was
performed to examine the proportion of HSCs undergoing apoptosis
using Cell Quest Pro version 5.1 from BD Biosciences.
Characterization of BM-MSCs and AD-MSCs was
performed by using flow cytometry, during which, the expression of
specific surface markers CD90, CD29 and CD45 was measured in
fluorescence intensity over respective isotype and unstained
negative control Ig.
Western blot analysis
Western blot analysis was performed to examine the
expression of α-smooth muscle actin (α-SMA), following a previously
described protocol (20). In brief,
total protein samples were prepared from HSCs following co-culture
with either BM-MSCs or AD-MSCs for 72 h, separated using SDS-PAGE
and transferred onto immunoblot polyvinylidene fluoride membranes.
Membranes were blocked and then incubated with goat anti-mouse
α-SMA monoclonal primary antibody (cat. no. ab5694; Abcam,
Cambridge, UK) at 1:200 dilution or anti-GAPDH (cat. no. ab9485;
Abcam) as internal control at 4°C overnight, followed by an
incubation with the secondary antibody rabbit anti-goat HRP (IgG
H&R) (cat. no. ab6741; Abcam) at a dilution of 1:30,000 at room
temperature for 1 h. Specific bands were visualized after using a
developing solution from Beyotime Institute of Biotechnology (cat.
no. P0019) and analyzed with a Gel-Pro analyzer imaging system from
Media Cybernetics, Inc. (Rockville, MD, USA). The intensity of the
α-SMA bands was normalized to that of GAPDH.
Animal model of liver fibrosis and
transplantation of MSCs
A total of 24 male SD rats weighing 250–300 g were
selected to establish the animal model of CCl4-induced
liver fibrosis. Rats were subcutaneously injected with 1.5 ml/kg
CCl4 diluted 1:1 (v/v) in olive oil twice a week to
induce liver fibrosis. The rats were randomly divided into four
groups: BM-MSC treatment (n=6) and AD-MSC treatment (n=6) groups, a
BRL treatment group acting as a negative control (n=6) and a group
treated with cell culture medium containing no cells acting as a
blank control (n=6). For transplantation of BM-MSCs or AD-MSCs into
the CCl4-treated rat liver, rats were anesthetized and
the abdomen was incised to identify the portal vein to the liver.
Subsequently, 5×106 BM-MSCs or AD-MSCs suspended in 1.5
ml PBS were injected through the portal vein once every 2 weeks.
Rats in the control groups were injected with 1.5 ml PBS alone.
Following treatment of the rats for 4 weeks, they were sacrificed
and used for subsequent experiments.
Immunohistochemical staining
Paraffin-embedded sections (4-mm in thickness) of
the liver were prepared following a previously published protocol
(16), dewaxed in xylene, hydrated
with distilled water and incubated with 3%
H2O2 at room temperature for 30 min.
Following washing with 1X PBS, slides were blocked with 2% goat
serum in 0.01 M PBS containing 0.3% Triton X-100 (PBS-X) for 1 hat
room temperature, and subsequently incubated with α-SMA primary
antibody (cat. no. 5694; Abcam) at a dilution of 1:200 at room
temperature for 1 h and rinsed with 1X PBS at room temperature. A
working solution with secondary antibody rabbit anti-goat HRP (IgG
H&R; cat. no. ab6741; Abcam) was added at a dilution of
1:30,000 and incubated at room temperature for 30 min Subsequently,
the slides were visualized using an Olympus microscope (Model IX71)
with magnification, ×200 following development with diaminobenzine,
counterstaining with hematoxylin, dehydration and mounting on a
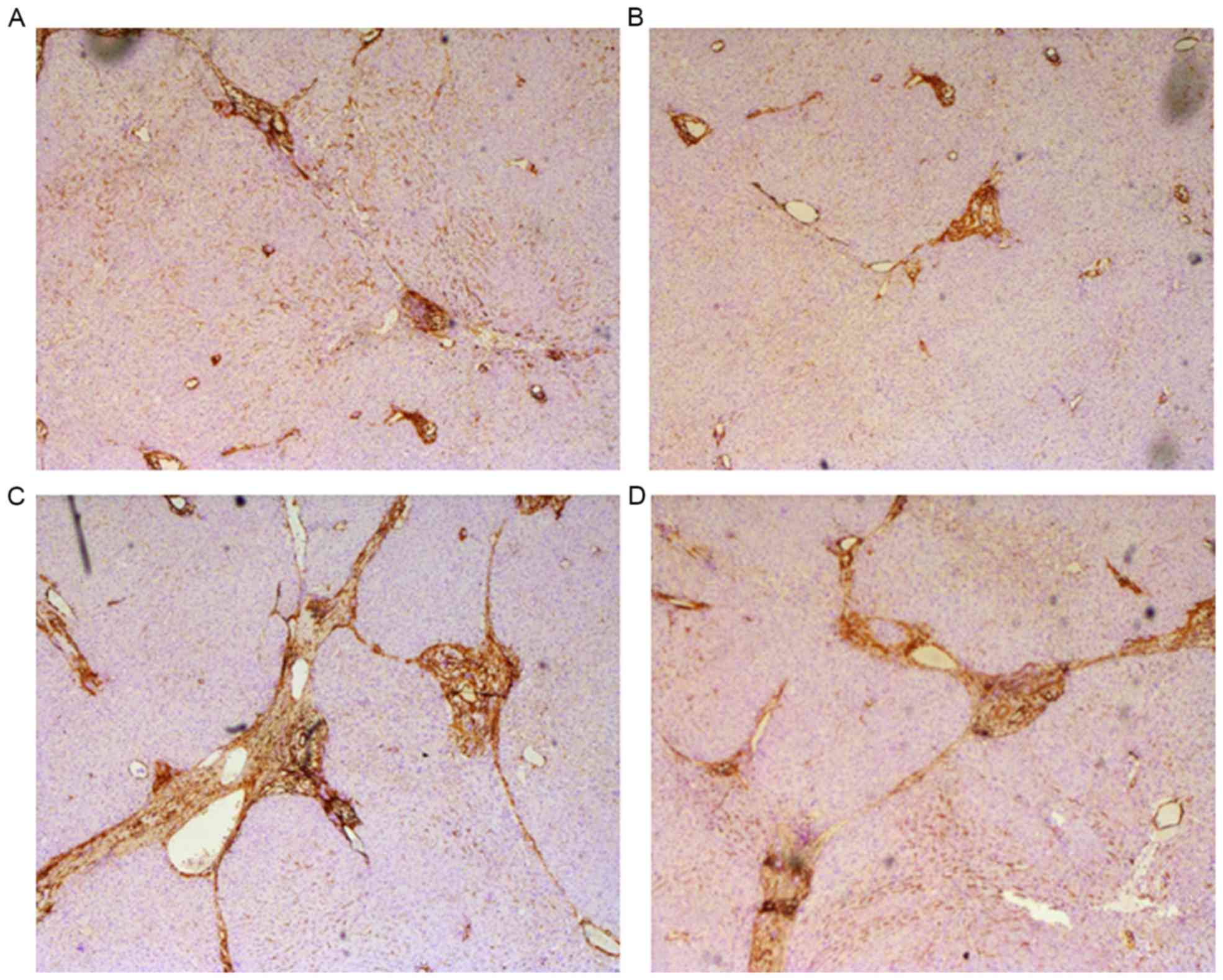
neutral resin. Brown staining indicated α-SMA expression.
Inflammation and fibrosis scores were evaluated according to the
Ishak modified histological activity index (21). Masson staining was performed using a
Masson staining kit (cat. no. HT15; Sigma-Aldrich; Merck KGaA)
according to the manufacturer's instructions.
Measurement of cytokine secretion by
ELISA
BM-MSCs or AD-MSCs at a density of 2×105
cells/well were seeded onto 6-well plates. BRLs were seeded instead
of MSCs to act as a negative control. After 72 h culture, the cell
culture medium was collected to measure cytokine secretion using
ELISA kits (R&D Systems, Inc., Minneapolis, MN, USA) for
vascular endothelial growth factor (VEGF) (cat. no. RRV00),
interleukin-10 (IL-10) (cat. no. R1000), nerve growth factor (NGF)
(cat. no. DY556) and transforming growth factor-β1 (TGF-β1) (cat.
no. MB100B), following the manufacturer's protocol.
Determination of collagen content in
rat liver tissue and serum
Collagen content in rat liver tissue was examined
using a hydroxyproline determination kit (cat. no. KA4552; Abnova,
Taipei, Taiwan) in which hydroxyproline served as a measure of
collagen concentration. Briefly, 80 mg rat liver tissue was
homogenized and centrifuged at 100 × g at 4°C for 60 sec, after
which supernatants were collected for subsequent hydroxyproline
measurement. Absorbance at 550 nm was determined with a
spectrophotometer. collagen content within the liver tissue was
calculated according to the Hydroxyproline Assay kit protocol
provided by Abnova. Serum type III collagen were determined by
ELISA using an ELISA kit from Kamiya Biomedical Company (Shanghai,
China; cat. no. KT-11210), hyaluronic acid levels were measured
with an ELISA kit from Wuhan Fine Biotech Ltd. (Wuhan, China; cat.
no. EU2556).
Statistical analysis
The experiments conducted in the present study
included triplicate samples for each treatment group. All
statistical analyses were performed using SPSS version 13.0 (SPSS
Inc., Chicago, IL, USA). All data are presented as the mean ±
standard deviation, and a comparison among multiple groups was
performed using analysis of variance and Tukey's post-hoc tests.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Preparation and characterization of
BM-MSCs and AD-MSCs
BM-MSCs and AD-MSCs were prepared from the freshly
harvested bone marrow and adipose tissue of a male SD rat. Trypan
blue staining revealed a high degree of proliferation (92–98%) and
purity (80–95%) of the two types of MSCs (data not shown). After
the cells were cultured for 48 h, cell morphology was examined
using an inverted microscope. The vast majority of the cells were
adherent and had a spindle or trigonal shape and only a small
number of suspended cells were visualized. After 3–5 days, the
cells reached ~80% confluence, at which time the detached cells
were washed with PBS and passaged with a small number of the cells
transferred into a new vessel. There were no marked differences
between the morphology of BM-MSCs and AD-MSCs (Fig. 1). Subsequently, the two types of stem
cells were characterized based on the expression of their specific
surface markers by fluorescence-activated cell sorting flow
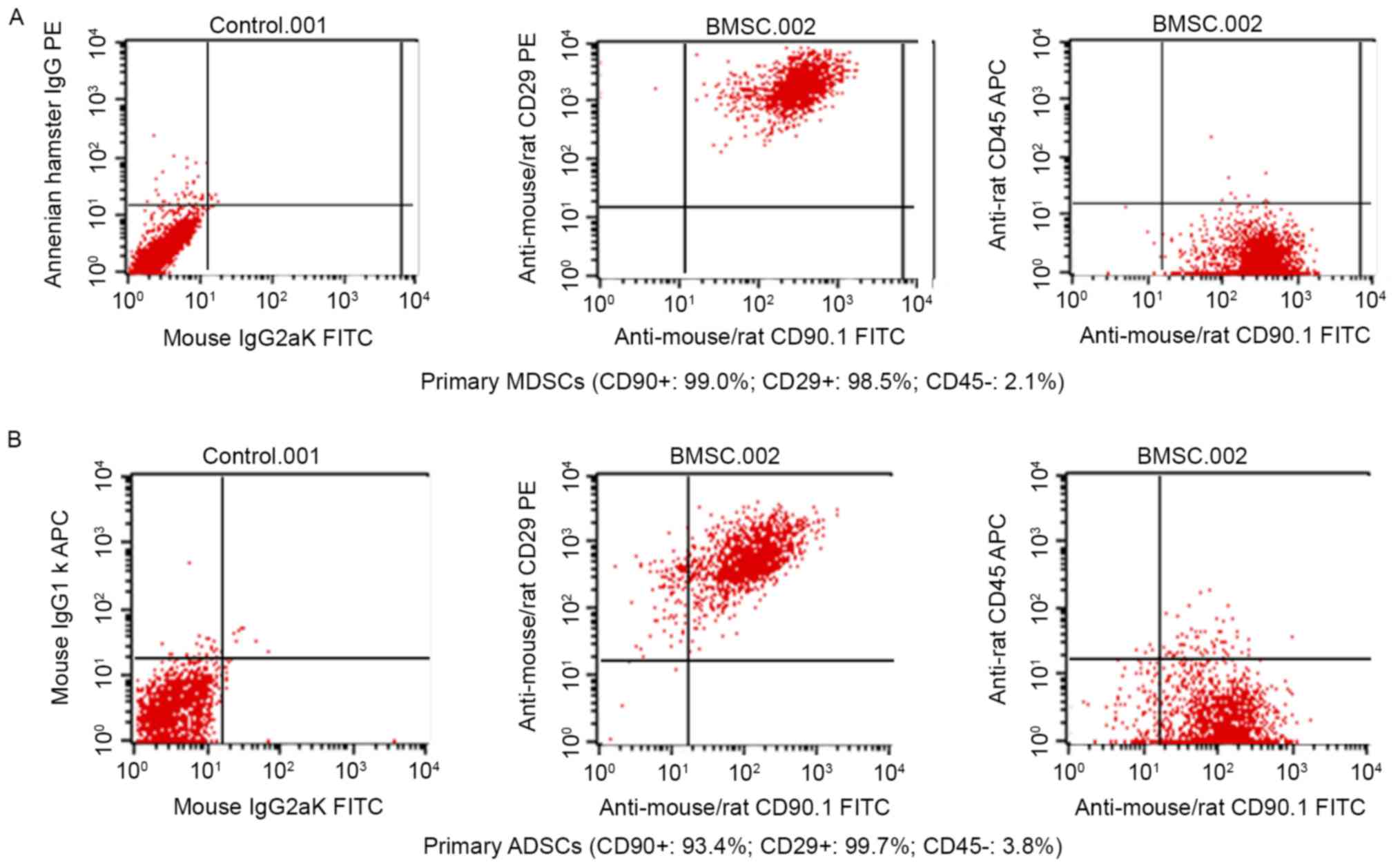
cytometry. As depicted in Fig. 2A and
B, 99.0 and 98.5% of BM-MSCs expressed CD90 and CD29,
respectively, and 93.4 and 99.7% of AD-MSCs expressed CD90 and
CD29, respectively, indicating that MSC surface markers were highly
expressed by BM-MSCs and AD-MSCs. By contrast, the positive rates
of the hematopoietic cell origin marker CD45 expression were low;
only 2.1% of BM-MSCs and 3.8% of AD-MSCs expressed CD45 (Fig. 2). The morphological examination and
the identification of MSC surface markers confirmed the successful
preparation of BM-MSCs and AD-MSCs.
Effects of BM-MSCs and AD-MSCs on the
activation, proliferation and apoptosis of HSCs in a co-culture
system
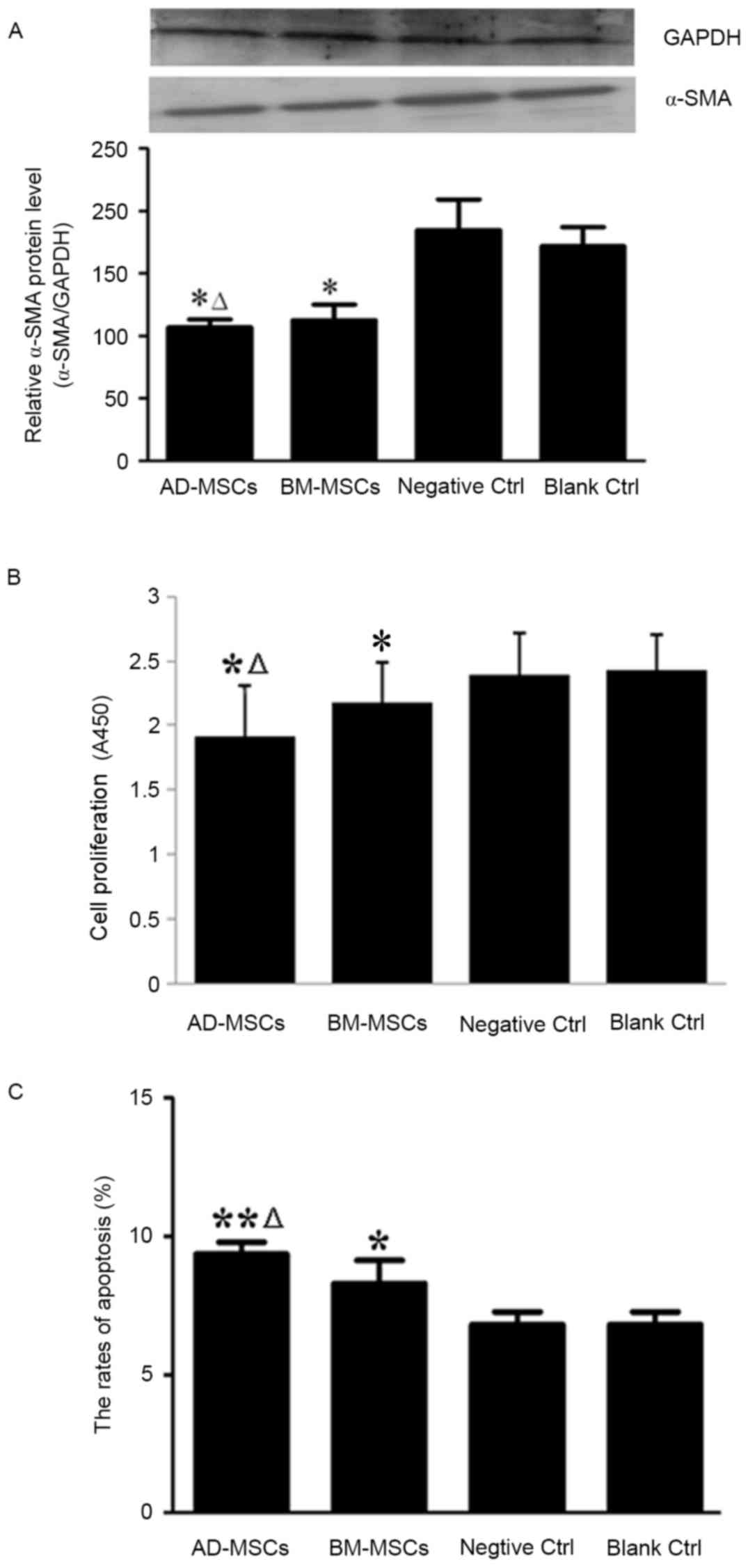
HSC activation serves a pivotal role in the
pathogenesis of liver fibrosis and α-SMA is a classical molecular
marker for activated HSCs (22);
therefore, the effects of the two types of MSCs on the levels of
α-SMA were determined in the current study. Western blot analysis
revealed that expression of α-SMA was significantly reduced in HSCs
co-cultured with either type of MSCs, compared with the expression
in the control groups (P<0.05; Fig.
3A). The effects of BM-MSCs and AD-MSCs on the expression of
α-SMA were compared and it was demonstrated that the AD-MSC
co-culture group exhibited significantly lower α-SMA expression
(P<0.05; Fig. 3A).
Subsequently, the effects of BM-MSCs and AD-MSCs on
the proliferation and apoptosis of HSCs were compared after 72 h
co-culture. Compared with that of the controls, the proliferation
of HSCs was significantly inhibited by co-culture with either
BM-MSCs or AD-MSCs (P<0.05), with a significantly greater
reduction in HSC proliferation achieved by AD-MSCs than BM-MSCs
(P<0.05; Fig. 3B). Data
demonstrated that the two types of MSCs promoted the apoptosis of
HSCs, with a higher rate of apoptosis induced by AD-MSCs compared
with BM-MSCs (P<0.05; Fig.
3C).
Comparison of cytokine secretion by
BM-MSCs and AD-MSCs in co-culture with HSCs
The aforementioned observations indicate that BM-MSC
and AD-MSCs were able to reduce liver fibrosis in the cell
co-culture system. Since there was no direct cell-cell contact
between HSCs and MSCs, the secretion of a select group of cytokines
into the cell culture medium by MSCs was evaluated. It has been
documented that VEGF, IL-10, NGF and TGF-β1 are associated with the
control of cellular functions (18,19,23). As
presented in Table I, compared with
the control cells, each type of MSC secreted significantly more
VEGF (BM-MSCs vs. BRLs: P<0.05; AD-MSCs vs. BRLs, P<0.05) and
significantly less NGF (BM-MSCs vs. BRLs, P<0.05; AD-MSCs vs.
BRLs, P<0.05) and TGF-β1 (BM-MSCs vs. BRLs, P<0.05; AD-MSCs
vs. BRLs, P<0.05) compared with the control. IL-10 levels did
not differ between MSCs and the control. Furthermore, NGF and
TGF-β1 levels in the AD-MSC culture medium were significantly
higher than those in the BM-MSC culture medium (AD-MSCs vs.
BM-MSCs, P<0.05), whereas levels of VEGF and IL-10 in the media
of the two MSC types did not differ significantly (AD-MSCs vs.
BM-MSCs: P>0.05).
 | Table I.Levels of VEGF, IL-10, NGF and TGF-β1
secreted into the cell culture medium of HSCs co-cultured with
BRLs, BM-MSCs or AD-MSCs. |
Table I.
Levels of VEGF, IL-10, NGF and TGF-β1
secreted into the cell culture medium of HSCs co-cultured with
BRLs, BM-MSCs or AD-MSCs.
| Co-culture
group | VEGF (pg/ml) | IL-10 (pg/ml) | NGF (pg/ml) | TGF-β1 (pg/ml) |
|---|
| BRLs |
21.08±5.15 |
17.91±3.16 |
10.13±1.52 |
53.27±12.68 |
| BM-MSCs |
80.33±14.48a |
18.12±1.53 |
3.95±0.71a |
6.36±0.85a |
| AD-MSCs |
78.52±15.79a |
17.37±1.92 |
7.46±0.54a,b |
8.79±0.93a,b |
Anti-inflammatory and anti-fibrotic
effects of BM-MSC and AD-MSC transplantation in a rat model of
CCl4-induced liver fibrosis
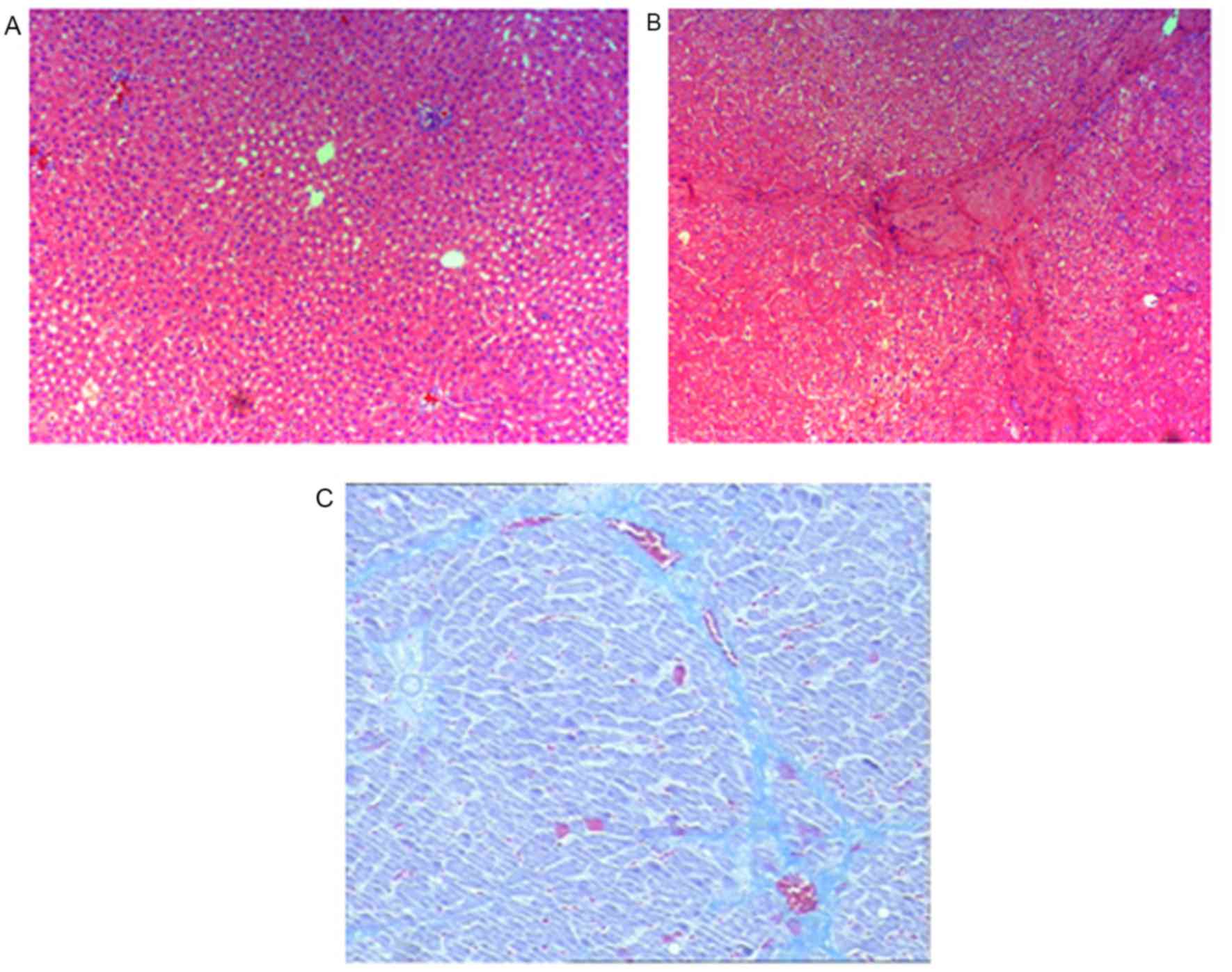
Prior to assessment of the anti-inflammatory and
anti-fibrotic effects of BM-MSC and AD-MSC transplantation in rats,
we validated the rat model of CCl4-induced liver
fibrosis. Compared with the control group treated with vehicle
olive oil only (Fig. 4A), rats
exposed to CCl4 exhibited typical features of liver
inflammation and fibrogenesis as revealed by hematoxylin and eosin
(Fig. 4B), and Masson staining
(Fig. 4C). These features included
an increase in the density of collagen fibers, markedly
proliferated fibrous tissue around the portal areas, inflammatory
cell filtration and the formation of lobules. Pathological
examination and measurement of liver fibrotic marker expression in
the liver and serum samples from the control groups and the groups
treated with the two types of MSCs were performed.
Immunocytochemical staining revealed decreased hepatic steatosis, a
smaller amount of fake leaflets and thinner fibre cords in the
MSC-treated groups. In addition, the formation of fibrosis in the
periportal areas and hepatic parenchyma were reduced in the
MSC-treated groups (Fig. 5A and B).
By contrast, the following pathological changes occurred in the
control groups: Marked hepatic steatosis, fibrogenesis in the
periportal areas and hepatic parenchyma, wide and thick cords of
collagen fibers and a large amount of fake leaflets (Fig. 5C and D).
Furthermore, liver inflammation and fibrosis staging
scores were compared between the control and MSC-treated groups. In
the MSC-treated groups, the inflammation activity (BM-MSCs:
F=51.26, P<0.05; AD-MSCs: F=46.73, P<0.05) and fibrosis
staging scores (BM-MSCs: F=32.29, P<0.05; AD-MSCs: F=40.94,
P<0.05) were significantly lower in the MSC-treated groups
compared with the control groups. Although implantation of AD-MSCs
resulted in a slight improvement in anti-inflammatory and
anti-fibrotic effects compared with implantation of BM-MSCs, the
differences between the two groups were not significant (P>0.05;
Table II).
| Table II.Liver inflammation and fibrosis
staging scores in the control, BM-MSC and AD-MSC transplanted
groups. |
Table II.
Liver inflammation and fibrosis
staging scores in the control, BM-MSC and AD-MSC transplanted
groups.
| Group | Inflammatory
activity | Fibrosis staging
score |
|---|
| Blank control |
13.78±2.53 |
5.09±1.15 |
| Negative
control |
13.34±1.89 |
4.95±1.22 |
| BM-MSCs |
9.87±2.07a |
4.17±0.94a |
| AD-MSCs |
10.13±1.81a |
3.98±0.82a |
Serum hyaluronic acid, collagen type III and
intrahepatic hydroxyproline are well-documented markers of fibrosis
and may indicate the degree of liver fibrosis (24). To further compare the protective
effects of implantation of the two types of MSCs, inflammation and
fibrosis were induced in rats by CCl4 treatment and
ELISA was performed to measure the serum levels of hyaluronic acid,
collagen type III and intrahepatic hydroxyproline. As depicted in
Table III, the serum
concentrations of hyaluronic acid, collagen type III and
intrahepatic hydroxyproline were significantly lower in the two
MSC-treated groups than in the control groups (F=73.51, P<0.05
for hyaluronic acid; F=76.19, P<0.05 for collagen type III; and
F=60.37, P<0.05 for intrahepatic hydroxyproline). Although
transplantation of AD-MSCs resulted in slightly decreased serum
levels of the fibrotic markers, the differences in serum levels in
the two types of MSCs were not significant (P>0.05; Table III).
| Table III.Serum levels of hyaluronic acid,
collagen III and hydroxyproline in control, BM-MSC and AD-MSC
transplanted groups. |
Table III.
Serum levels of hyaluronic acid,
collagen III and hydroxyproline in control, BM-MSC and AD-MSC
transplanted groups.
| Group | Hyaluronic acid
(µg/l) | Collagen III
(µg/l) | Hydroxyproline
(µg/l) |
|---|
| Blank control |
287.5±26.7 |
32.5±4.3 |
473.9±63.7 |
| Negative
control |
282.3±18.7 |
35.3±3.3 |
458.4±38.1 |
| BM-MSCs |
191.5±33.2a |
19.9±5.1a |
312.6±38.8a |
| AD-MSCs |
178.8±28.2a |
21.7±3.3a |
325.8±28.2a |
Discussion
The present study compared the ability of two types
of MSCs; BM-MSCs and AD-MSCs, to attenuate liver fibrosis using an
in vitro co-culture system and an in vivo rat model
of liver fibrosis. The major observations were as follows: i)
BM-MSCs and AD-MSCs significantly inhibited the proliferation and
activation of HSCs, with a statistically greater effect achieved by
AD-MSCs compared with BM-MSCs in a co-culture system; ii)
comparison of secretion of four cytokines by BM-MSCs and AD-MSCs
into the cell culture medium identified similar levels of VEGF and
IL-10 secretion between the two groups but significantly higher
levels of NGF and TGF-β1 secretion by AD-MSCs compared with
BM-MSCs; and iii) the inflammatory activity and fibrosis staging
scores in a rat model of CCl4-induced liver fibrosis
were significantly lower in the BM-MSC- and AD-MSC-treated groups
than in the control groups. AD-MSC implantation induced slightly
improved anti-inflammatory and anti-fibrotic effects compared with
BM-SC implantation; however, this difference was not
significant.
Over the past few decades, studies have demonstrated
that MSCs are able to differentiate into hepatocytes, secrete a
variety of cytokines to modulate the immune responses and indicated
that they possess potent anti-oxidant properties. These
capabilities allow MSCs to combat liver fibrosis regardless of
etiological factors (25–28). Indeed, MSC therapy has been widely
studied and suggested as an alternative novel method of treating
liver fibrosis (29–30). Furthermore, the development of MSC
therapy may bring hope to patients with end-stage liver fibrosis on
the waiting list for a liver transplant (30). MSCs can be derived from a wide range
of tissue sources, including the bone marrow, adipose tissue,
dental pulp and umbilical cord blood (12,13).
Furthermore, the differences and similarities of MSCs isolated from
different sources and maintained in different culture conditions
have been investigated (12,13,16). The
results of the present study indicate that BM-MSCs and AD-MSCs
significantly inhibit the proliferation and activation of HSCs as
well as promote apoptosis in HSCs compared with controls. These
observations are consistent with those described in previous
reports (30). The current study
compared the effects of BM-MSCs and AD-MSCs on the cellular
functions of HSCs and it was revealed that AD-MSCs were
significantly more effective at inhibiting HSC proliferation and
promoting HSC apoptosis than BM-MSCs. There was a similar effect on
HSC activation. Further analysis of cytokine secretion by BM-MSCs
and AD-MSCs in the co-culture system demonstrated that levels of
NGF and TGF-β1 were significantly greater in the AD-MSC culture
medium than in the BM-MSC culture medium, whereas no significant
differences were detected in VEGF and IL-10 levels between the two
groups. It has been widely reported that these cytokines are
involved in regulating the proliferation, activation and apoptosis
of HSCs; therefore, we postulated that the distinct effects of
BM-MSCs and AD-MSCs on HSCs may be attributed to the differential
levels of these cytokines released by BM-MSCs and AD-MSCs. IL-10 is
a well-known anti-inflammatory cytokine capable of inhibiting the
synthesis of pro-inflammatory cytokines, blocking the activity of
nuclear factor-κB, enhancing B-cell proliferation and affecting the
cytokine-activated Janus kinase-signal transducer and activator of
transcription signaling pathway (31). It was demonstrated in the present
study that the secretion of IL-10 into the media of each MSC type
did not differ significantly, which may explain, at least in part,
why the effects of BM-MSCs and AD-MSCs on HSC activation are very
similar.
The present study also evaluated the effectiveness
of BM-MSCs and AD-MSCs in the treatment of liver fibrosis using a
rat model of liver fibrosis, which was induced by injection of
CCl4. The CCl4-induced animal model has a
number of benefits as a suitable animal model for studying liver
fibrosis with patterns similar to those of liver fibrotic disease
in humans (32). However, the
CCl4-induced animal model has a number of limitations
(32), as the model cannot replicate
the exact condition of liver fibrosis in humans and cannot
represent distinctions in immune responses, gene expression and
regulation. This limits the conclusions that can be drawn from the
present study. The current study also investigated an animal model
of liver fibrosis. It was demonstrated that inflammatory activity
and fibrosis staging scores were significantly lower in MSC-treated
groups than in the control groups. Furthermore, implantation of
AD-MSCs slightly improved the anti-inflammatory and anti-fibrotic
effects compared with BM-MSCs. However, these differences were not
significant, which is inconsistent with the in vitro
observations of the present study. In vitro studies
investigating the effectiveness of AD-MSCs and BM-MSCs may lead to
results that do not correspond with the results from animal models.
Furthermore, the possibility that a range of immune responses in
the animal model are involved could not be excluded, since animal
models are far more complex than the co-culture system. Further
investigations are currently under way in our laboratory to improve
understanding of the underlying cellular and molecular mechanisms
responsible for the effects of BM-MSCs and AD-MSCs, and to advance
knowledge regarding the ability of BM-MSCs and AD-MSCs to combat
liver fibrosis.
In conclusion, the results of the present study
indicate that BM-MSCs and AD-MSCs are similarly effective at
attenuating liver fibrosis by inhibiting the activation and
proliferation of HSCs, as well as promoting the apoptosis of HSCs.
Considering that AD-MSCs are easier to prepare and more effective
at inhibiting HSC proliferation and apoptosis in the co-culture
system used in the present study, and that the implantation of
AD-MSCs exhibited slightly improved anti-inflammatory and
anti-liver fibrotic activities compared with BM-MSCs in the rat
model of CCl4-induced liver fibrosis, AD-MSCs may be a
better candidate than other MSCs for cell-based therapy to treat
liver fibrosis. Future clinical studies and an understanding of the
therapeutic mechanisms are required to advance understanding of the
effects of AD-MSCs and BM-MSCs in the treatment of liver
fibrosis.
References
|
1
|
Friedman SL: Liver fibrosis-from bench to
bedside. J Hepatol. 38 Suppl 1:S38–S53. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bosch J and García-Pagán JC: Complications
of cirrhosis. I. Portal hypertension. J Hepatol. 32 1
Suppl:S141–S156. 2000. View Article : Google Scholar
|
|
3
|
Cárdenas A: Hepatorenal syndrome: A
dreaded complication of end-stage liver disease. Am J
Gastroenterol. 100:460–467. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bataller R and Brenner DA: Liver fibrosis.
J Clin Invest. 115:209–218. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Friedman SL: Hepatic stellate cells:
Protean, multifunctional, and enigmatic cells of the liver. Physiol
Rev. 88:125–172. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Salem HK and Thiemermann C: Mesenchymal
stromal cells: Current understanding and clinical status. Stem
Cells. 28:585–596. 2010.PubMed/NCBI
|
|
7
|
Dezawa M, Ishikawa H, Itokazu Y, Yoshihara
T, Hoshino M, Takeda S, Ide C and Nabeshima Y: Bone marrow stromal
cells generate muscle cells and repair muscle degeneration.
Science. 309:314–317. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Das M, Sundell IB and Koka PS: Adult
mesenchymal stem cells and their potency in the cell-based therapy.
J Stem Cells. 8:1–16. 2013.PubMed/NCBI
|
|
9
|
Lu T, Yang C, Sun H, Lv J, Zhang F and
Dong XJ: FGF4 and HGF promote differentiation of mouse bone marrow
mesenchymal stem cells into hepatocytes via the MAPK pathway. Genet
Mol Res. 13:415–424. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ren G, Chen X, Dong F, Li W, Ren X, Zhang
Y and Shi Y: Concise review: Mesenchymal stem cells and
translational medicine: Emerging issues. Stem Cells Transl Med.
1:51–58. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sakaida I, Terai S, Yamamoto N, Aoyama K,
Ishikawa T, Nishina H and Okita K: Transplantation of bone marrow
cells reduces CCl4-induced liver fibrosis in mice. Hepatology.
40:1304–1311. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chamberlain G, Fox J, Ashton B and
Middleton J: Concise review: Mesenchymal stem cells: Their
phenotype, differentiation capacity, immunological features, and
potential for homing. Stem Cells. 25:2739–2749. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qi Z, Zhang Y, Liu L, Guo X, Qin J and Cui
G: Mesenchymal stem cells derived from different origins have
unique sensitivities to different chemotherapeutic agents. Cell
Biol Int. 36:857–862. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Berardis S, Lombard C, Evraerts J, El
Taghdouini A, Rosseels V, Sancho-Bru P, Lozano JJ, van Grunsven L,
Sokal E and Najimi M: Gene expression profiling and secretome
analysis differentiate adult-derived human liver stem/progenitor
cells and human hepatic stellate cells. PLoS One. 9:e861372014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Takeda M, Yamamoto M, Isoda K, Higashiyama
S, Hirose M, Ohgushi H, Kawase M and Yagi K: Availability of bone
marrow stromal cells in three-dimensional coculture with
hepatocytes and transplantation into liver-damaged mice. J Biosci
Bioeng. 100:77–81. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu F, Ji S, Su L, Wan L, Zhang S, Dai C,
Wang Y, Fu J and Zhang Q: Adipose-derived mesenchymal stem cells
inhibit activation of hepatic stellate cells in vitro and
ameliorate rat liver fibrosis in vivo. J Formos Med Assoc.
114:130–138. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu Y, Wang Z, Wang J, Lam W, Kwong S, Li
F, Friedman SL, Zhou S, Ren Q, Xu Z, et al: A histone deacetylase
inhibitor, largazole, decreases liver fibrosis and angiogenesis by
inhibiting transforming growth factor-β and vascular endothelial
growth factor signalling. Liver Int. 33:504–515. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Majumder S, Piguet AC, Dufour JF and
Chatterjee S: Study of the cellular mechanism of Sunitinib mediated
inactivation of activated hepatic stellate cells and its
implications in angiogenesis. Eur J Pharmacol. 705:86–95. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen YX, Huang YH, Zheng WD, Chen ZX,
Zhang LJ and Wang XZ: Interleukin-10 gene modification attenuates
hepatocyte activation of rat hepatic stellate cells in
vitro. Mol Med Rep. 7:371–378. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu F, Su L, Ji S, Zhang S, Yu P, Zheng Y
and Zhang Q: Inhibition of hepatic stellate cell activation and
liver fibrosis by fat-specific protein 27. Mol Cell Biochem.
369:35–43. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sumida Y, Nakajima A and Itoh Y:
Limitations of liver biopsy and non-invasive diagnostic tests for
the diagnosis of nonalcoholic fatty liver disease/nonalcoholic
steatohepatitis. World J Gastroenterol. 20:475–485. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu J, Zhang S, Chu ES, Go MY, Lau RH, Zhao
J, Wu CW, Tong L, Zhao J, vPoon TC and Sung JJ: Peroxisome
proliferator-activated receptors gamma reverses hepatic nutritional
fibrosis in mice and suppresses activation of hepatic stellate
cells in vitro. Int J Biochem Cell Biol. 42:948–957. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu Y, Wang Z, Wang J, Lam W, Kwong S, Li
F, Friedman SL, Zhou S, Ren Q, Xu Z, et al: A histone deacetylase
inhibitor, largazole, decreases liver fibrosis and angiogenesis by
inhibiting transforming growth factor-β and vascular endothelial
growth factor signaling. Liver Int. 33:504–515. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang X, Wang X, Lv Y, Xu L, Lin J and
Diao Y: Protection effect of kallistatin on carbon
tetrachloride-induced liver fibrosis in rats via antioxidative
stress. PLoS One. 9:e884982014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sato Y, Araki H, Kato J, Nakamura K,
Kawano Y, Kobune M, Sato T, Miyanishi K, Takayama T, Takahashi M,
et al: Human mesenchymal stem cells xenografted directly to rat
liver are differentiated into human hepatocytes without fusion.
Blood. 106:756–763. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sharma RR, Pollock K, Hubel A and McKenna
D: Mesenchymal stem or stromal cells: A review of clinical
applications and manufacturing practices. Transfusion.
54:1418–1437. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang J, Bian C, Liao L, Zhu Y, Li J, Zeng
L and Zhao RC: Inhibition of hepatic stellate cells proliferation
by mesenchymal stem cells and the possible mechanisms. Hepatol Res.
39:1219–1228. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Haddad R and Saldanha-Araujo F: Mechanisms
of T-cell immunosuppression by mesenchymal stromal cells: What do
we know so far? Biomed Res Int. 2014:2168062014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sakaida I, Terai S, Yamamoto N, Aoyama K,
Ishikawa T, Nishina H and Okita K: Transplantation of bone marrow
cells reduces CCl4-induced liver fibrosis in mice. Hepatology.
40:1304–1311. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ma ZG, Lv XD, Zhan LL, Chen L, Zou QY,
Xiang JQ, Qin JL, Zhang WW, Zeng ZJ, Jin H, et al: Human
urokinase-type plasminogen activator gene-modified bone
marrow-derived mesenchymalstem cells attenuate liver fibrosis in
rats by down-regulating the Wnt signaling pathway. World J
Gastroenterol. 22:2092–2103. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Iyer SS and Cheng G: Role of interleukin
10 transcriptional regulation in inflammation and autoimmune
disease. Crit Rev Immunol. 32:23–63. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liedtke C, Luedde T, Sauerbruch T,
Scholten D, Streetz K, Tacke F, Tolba R, Trautwein C, Trebicka J
and Weiskirchen R: Experimental liver fibrosis research: Update on
animal models, legal issues and translational aspects. Fibrogenesis
Tissue Repair. 6:192013. View Article : Google Scholar : PubMed/NCBI
|