Introduction
The human skeletal system comprises highly dynamic
tissue that exhibits continuous bone formation by osteoblasts and
bone resorption by osteoclasts to maintain homeostasis (1). Previous studies have suggested that
dysfunction of remodeling and metabolism of osteoblasts and
osteoclasts are associated with the progression of osteoporosis in
individuals aged >50 years (2,3).
Osteoporosis exhibits complex pathology and etiology, and is
considered as a multifactorial polygenic disease associated with
environmental, hormonal, nutritional and genetic factors. Studies
have indicated that estrogen concentration is correlated with the
pathogenesis of osteoporosis by regulating bone homeostasis and
preventing postmenopausal bone loss (4,5).
Although numerous perspective studies on postmenopausal
osteoporosis have been reported, the molecular mechanisms of the
pathogenesis induced by postmenopausal osteoporosis are not
completely understood.
The clinical consequences of osteoporosis include
fractures in the upper extremities, hip and even spine, which
result in the loss of function and independence, impaired quality
of life, as well as increased morbidity and mortality (6–8). Various
treatments for postmenopausal osteoporosis, including farnesyl
pyrophosphate synthase inhibitor, have been proposed in a large
number of preclinical and clinical studies (9–11). In
addition, the downregulation of levels of osteoassociated hormones
have been detected in the blood plasma of postmenopausal women with
and without osteoporosis in clinical trials (12,13).
Furthermore, comprehensive treatments with anti-resorptive agents,
such as alendronate, have been investigated for the prevention of
new non-vertebral fractures in patients with postmenopausal
osteoporosis (14). In the present
study, the alendronate-mediated benefits and mechanism for the
treatment of rats with ovariectomy-induced osteoporosis were
investigated.
Previous studies have demonstrated the efficacy of
long-term alendronate treatment on postmenopausal osteoporosis and
bone material properties (13,15). In
addition, arterial stiffness was improved following monthly
bisphosphonate treatment in women with postmenopausal osteoporosis
(16). The effects of other drug
treatments on endothelial function, glucose metabolism and
inflammation markers have also been investigated in patients with
postmenopausal osteoporosis (17–19).
Furthermore, a previous study examined the effects of short-term
treatment with alendronate (70 mg; once-a-week) in women with
postmenopausal osteoporosis, determined by bone turnover markers,
and suggested that alendronate treatment was beneficial in these
patients (18).
Signal transducer and activator of transcription 1
(STAT1) signaling pathway is strongly activated by interferon-β
(IFN-β) in the pathogenesis and progression of osteoporosis, and
previous results contributed to further understanding the
pathological signaling pathways involved in osteoporosis (20). The importance of STAT1 gene
expression in monocytes has been reported, and the progression of
osteoporosis was observed to be regulated by the expression and
phosphorylation levels of STAT1 (21). These findings suggest that STAT1 gene
expression may be associated with the initiation and development of
osteoporosis.
In the present study, the therapeutic effects and
molecular mechanism of alendronate were investigated in osteoblast
cells and in rats with osteoporosis induced by ovariectomy. The
potential involvement of the IFN-β/STAT1 signaling pathway mediated
by alendronate in the progression of ovariectomy-induced
osteoporosis was also examined. The findings suggested that
alendronate regulated osteoblast differentiation and bone formation
through upregulation of IFN-β/STAT1 pathway in the rats. These
results may contribute to the clinical treatment of patients with
postmenopausal osteoporosis.
Materials and methods
Ethical statement
The present study was conducted in strict accordance
with the guidelines for the Care and Use of Laboratory Animals of
Jining No. 1 People's Hospital (Jining, China). All experimental
protocols and animals were approved by Ethics Committee of the
Animal Experiments Defense Research of Jining No. 1 People's
Hospital (Jining, China). All surgery and euthanasia procedures
aimed to minimize animal suffering.
Animal study
Female Wistar albino rats (age, 6–8 weeks) were
purchased from Shanghai Slack Experimental Animals Co., LTD
(Shanghai, China). Rats (n=40) were fed under pathogen-free
conditions and maintained in a controlled environment (temperature,
23±1°C; humidity, 50–60%), as well as an artificial simulation of
12h light/dark cycles. All rats underwent oophorectomy in order to
establish an osteoporosis model and to analyze the efficacy of
alendronate treatment. Experimental procedures were conducted
according to a previous study (22).
The induction lasted 14 days and the mandibular cortical width was
used as an indicator of osteoporosis to determine whether the model
was successfully established (23).
Rats with osteoporosis were randomly divided into two groups (n=20
in each group) and received treatment with alendronate (2.5 mg/kg;
Sigma-Aldrich; Merck KGaA; Darmstadt, Germany) or
phosphate-buffered saline (PBS; 2.5 mg/kg). The treatments were
performed once a day in a total of 10 times. On day 60, the
therapeutic efficacies of alendronate were analyzed according to a
previous study (24).
Bone mineral density
Bone tissues were isolated from experimental animals
according to previous study (25).
Bone cells were obtained from bone tissues as previously described
(26). A total of 1×105
bone cells were used to analyze bone mineral density. Bone mineral
density of the tibia of experimental rats was analyzed with an
automated software (N-Vivo 9.0; QSR International Ltd., Melbourne,
Australia) and quantified for analysis of the efficacy of
alendronate. All procedures were performed as previously described
(27). Intermediate bone density was
defined as mg/cm3.
Measurement of alkaline phosphatase
(ALP) activity
Osteoblasts were isolated from experimental mice as
described previously (28) and
cultured in 6-well plates (1×105 per well) with
alendronate at concentrations of 0, 0.50, 4.00, 8.00 and 16 mg/ml
for 48 h. The effect of alendronate treatment on osteoblasts cells
was analyzed by performing ALP activity assays and ALP staining
(Colorimetric Alkaline Phosphatase Assay kit; ab83369; Abcam;
Cambridge, MA, USA) as described in a previous study (29).
Bone resorption assays
Mature osteoclasts and osteoblasts were isolated
from experimental rats with ovariectomy-induced osteoporosis and
cultured in the presence of 10−6 M 1,25-dihydroxyvitamin
D3 and 10−6 M prostaglandin E2
(Sigma-Aldrich, Merck, Darmstadt, Germany) for 7 days. Next, the
cells were treated with alendronate (2 mg/ml) or PBS (2 mg/ml) for
48 h at 37°C. Subsequently, the minimum essential medium (MEM) was
removed from the culture and cells were washed with PBS three
times. Then, the total bone resorption was analyzed using ImagePro
Plus version 4.0 (Media Cybernetics, Inc., Silver Spring, MD,
USA).
Analysis of the effects of alendronate
on osteoclast differentiation
Bone marrow cells (BMCs) were obtained from female
Wistar albino rats as described previously (30) and cultured in MEM supplemented with
10% fetal bovine serum (FBS) for 48 h at 37°C. The medium was
removed, and fresh MEM was added before cells were cultured for 72
h in 10 ng/ml macrophage colony-stimulating factor (M-CSF;
Sigma-Aldrich; Merck KGaA) at 37°C. Tartrateresistant acid
phosphatase (TRAP) activity was measured (Sigma-Aldrich; Merck
KGaA; 387A-1KT) and used to examine the osteoclast differentiation
(31).
Measurement of osteoblast
viability
The viability of osteoblasts was analyzed in order
to examine the effects of alendronate on osteoporosis, as
descripted in a previous study (32). All experiments were performed in
triplicate.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assays
Total RNA from the femurs of rats was extracted
using RNAeasy Mini kit (Qiagen, Gaithersburg, MD, USA). The mRNA
expression levels of osteocalcin, osterix, Runt-related
transcription factor 2 (Runx2), TRAP, osteoblast differentiation
factor (ODF), osteoprotegerin (OPG), collagen 1A1 (COL1A1),
Fos-related antigen 1 (Fra1), TNF-receptor-associated factor 6
(TRAF6), suppressor of cytokine signaling 1 (SOCS1) and ALP in
osteoblasts and osteoclasts were determined by RT-qPCR with β-actin
as an endogenous control (33). All
the forward and reverse primers were synthesized by Thermo Fisher
Scientific, Inc. (Table I). PCR was
performed as follows: Preliminary denaturation at 94°C for 2 min,
followed by 45 cycles of 95°C for 30 sec, the annealing temperature
reduced to 57.5°C for 30 sec, and 72°C for 10 min in a total
reaction volume of 20 µl containing 50 ng of genomic DNA, 200 µM
dNTP, 2.5 units of Taq DNA polymerase, and 200 µM primers. Relative
mRNA expression level changes were calculated by 2−ΔΔCq
method (25). The results are
expressed as the n-fold way compared with the β-actin control.
 | Table I.Sequences of primers were used in the
present study. |
Table I.
Sequences of primers were used in the
present study.
|
| Sequence |
|---|
|
|
|
|---|
| Gene name | Reverse | Forward |
|---|
| ALP |
5′-CTAGCTGCTCCTATGTCTCACATC-3′ |
5′-GGTCAAGACCATTTGTAGGCTC-3′ |
| TRAP |
5′-TGGTCATTTCTTTGGGGCTTATCT-3′ |
5′-GCTACTTGCGGTTTCACTATGGA-3′ |
| ODF |
5′-CACCTGGTTGCTGACTAATTGAGA-3′ |
5′-CTTGCTGTCCGACCAAATA-3′ |
| OPG |
5′-AACGGCAACACAGCTCACAAGAAC-3′ |
5′-TGCTCGAAGGTGAGGTTAGCATGT-3′ |
| COL1A1 |
5′-CACCAATCACCTGCGGTACAGAA-3′ |
5′-CAGATCACGTCATCGCACAAC-3′ |
| Osteocalcin |
5′-ACACTCCTCGCCCTATTG-3′ |
5′-GATGTGGTCAGCCAACTC-3′ |
| Runx2 |
5′-TCTGGAAAAAAAAGGAGGGACTATG-3′ |
5′-GGTGCTCGGATCCCAAAAGAA-3′ |
| Osterix |
5′-GGAGGTTTCACTCCATTCCA-3′ |
5′-TAGAAGGAGCAAGGGGACAGAA-3′ |
| Fra1 |
5′-ATGTACCGAGACTTCGGGGAACC-3′ |
5′-CACTGTGTTGGCGTAGA-3′ |
| TRAF6 |
5′-CAAGTGTCGTGCCAAGTGAT-3′ |
5′-TTCCAGAAGTGCCAGGTTAATAC-3′ |
| SOCS1 |
5′-GCCTGCTGGCCATCCTAA-3′ |
3′-ACCGCCAGCACCACAGT-3′ |
| β-actin |
5′-CATCTCTTGCTCGAAGTCCA-3′ |
5′-ATCATGTTTGAGACCTTCAACA-3′ |
Western blot assay and histological
analysis
Osteoblasts from experimental rats with osteoporosis
treated with alendronate or PBS were homogenized in lysate buffer.
The supernatant of the mixture obtained by centrifugation at 6,000
× g for 10 min at 37°C and used for analysis of the target protein
levels. Transmembrane protein was extracted using a Transmembrane
Protein Extraction kit (71772-3; Qiagen) according to the
manufacturer's instructions. SDS-PAGE assays were performed as
previous described (34). For
western blotting, primary antibodies: IFN-β (1:1,000; ab77246;
Abcam), STAT1 (1:1,000; ab31369; Abcam), pSTAT1 (1:1,000; ab30645;
Abcam), β-actin (1:1,000; ab8226; Abcam) were added for 12 h at 4°C
after blocking in 5% skimmed milk for 1 h at 37°C, followed by
incubation with horseradish peroxidase-conjugated anti-rabbit IgG
(1706515; Bio-Rad Laboratories, Inc., Hercules, CA, USA) were used
at a 1:5,000 dilution for 24 h at 4°C. The results were visualized
using an enhanced chemiluminescence detection system (ChemiDoc™
Imaging Systems, Bio-Rad Laboratories, Inc.).
For histological analysis, experimental rats were
anesthetized using intraperitoneal injection of sodium
pentobarbital anesthesia (40 mg/kg; Invitrogen; Thermo Fisher
Scientific, Inc.) and the femur was isolated from experimental rats
after treatment with alendronate or PBS. Femur tissues were
sectioned (4-µm thick), fixed using 10% formaldehyde for 15 min at
30°C followed by embedding in paraffin wax. Hematoxylin-eosin
staining for 120 min at 30°C was used to analyze the bone quality
and density using a microscope (Olympus Corporation; Tokyo, Japan)
at a magnification, ×400.
Transfection with small interference
RNA (siRNA)
All siRNAs were synthesized by Invitrogen (Thermo
Fisher Scientific, Inc.), including the siRNA-IFN-β for endogenous
inhibition of IFN-β (EI-IFN-β) and siRNA-vector. Osteoblasts
(1×106) were transfected with 100 pmol siRNA-IFN-β using
Lipofectamine 2000 (Sigma-Aldrich; Merck KGaA), according to the
manufacturer's protocol. While siRNA-vector transfection was
performed in the control group.
Statistical analysis
All data are represented as the mean ± standard
error of triplicate experiments. Analysis was performing by SPSS
software (version 14.0; SPSS, Inc., Chicago, IL, USA). Unpaired
data were analyzed by Student's t-test, while comparisons of data
between multiple groups were conducted by analysis of variance.
P<0.05 was considered as an indicator of a statistically
significant difference.
Results
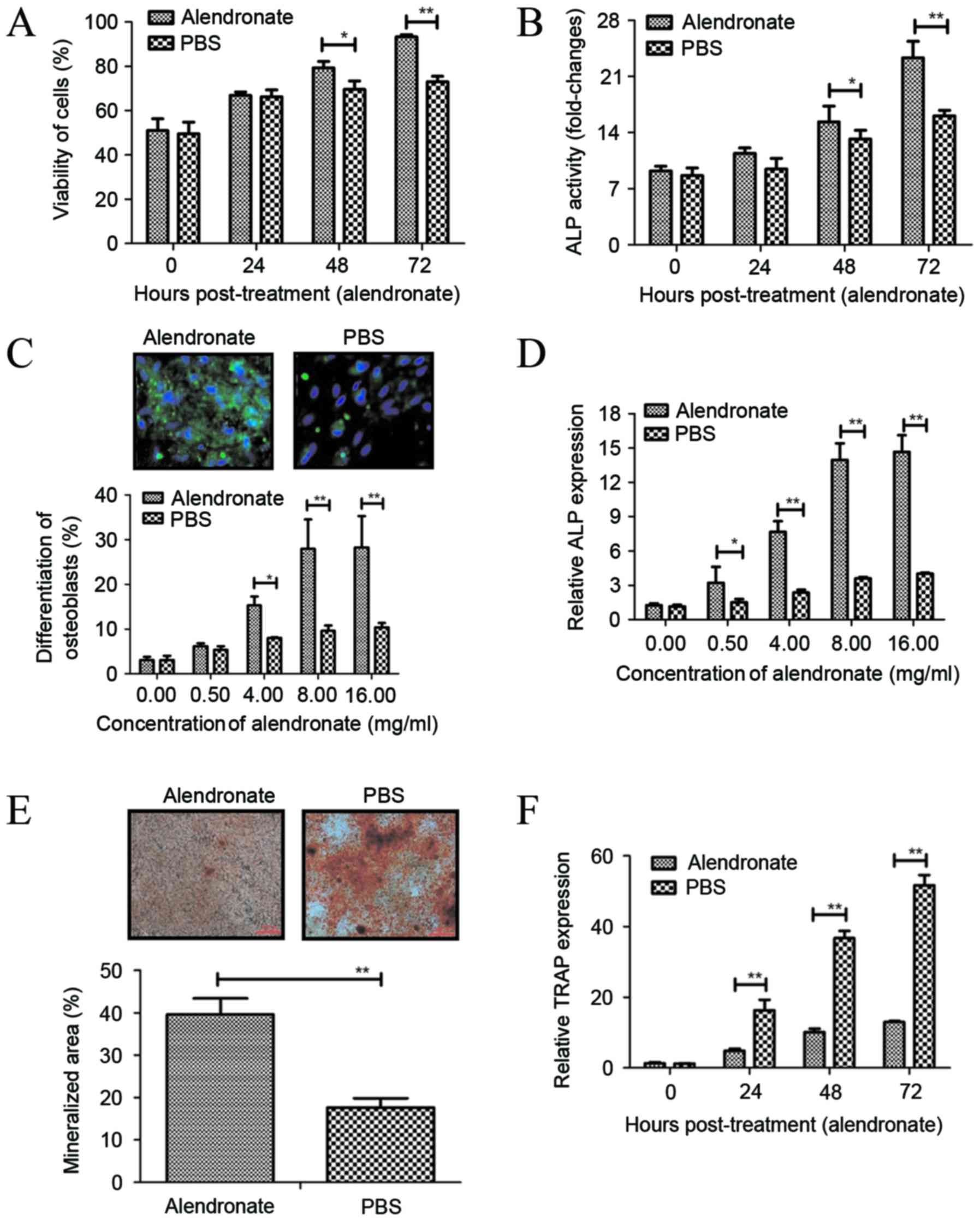
Effects of alendronate treatment on
the viability, ALP activity and differentiation of osteoblasts
In order to analyze the efficacy of alendronate
treatment on osteoporosis, the ALP activity and differentiation of
osteoblasts were analyzed following treatment with alendronate. It
was observed that alendronate significantly increased the viability
of osteoblasts after 48- and 72-h incubation (Fig. 1A). In addition, ALP activity was
upregulated in osteoblasts subsequent to treatment with alendronate
for 48 and 72 h, as compared with the control group (Fig. 1B). It was also revealed that
alendronate treatment at different concentrations (0.5–16 mg/ml)
promoted early differentiation marker of osteoblasts derived from
newborn rat calvaria determined by staining for ALP, with a
significant effect observed at ≥4 mg/ml (Fig. 1C). Furthermore, the results indicated
that alendronate (0.5–16 mg/ml) significantly enhanced ALP mRNA
expression in osteoblasts (Fig. 1D).
Alendronate treatment at 8 mg/ml markedly increased the mineralized
area, as determined by alizarin red S staining for calcium
(Fig. 1E). Notably, treatment with 8
mg/ml alendronate markedly decreased the mRNA expression levels of
TRAP compared with those in the PBS-treated group (Fig. 1F). These results suggest that
alendronate promotes osteoblast differentiation and bone
formation.
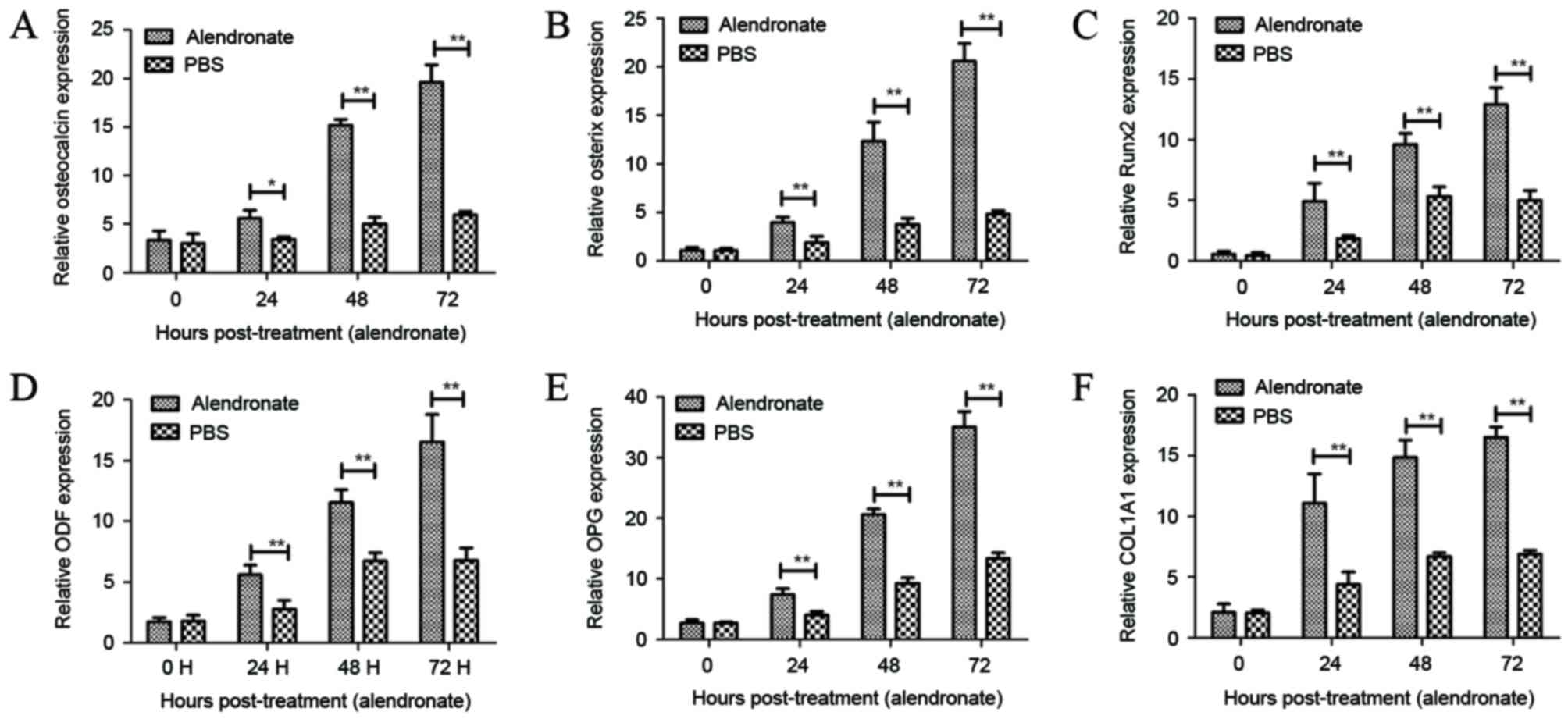
Alendronate stimulates the expression
levels of mRNAs associated with osteoblast differentiation
Next, the present study investigated the effects of
alendronate on the mRNAs expression levels of osteoblast
differentiation-associated factors in osteoblasts. It was observed
that alendronate markedly promoted the mRNA expression levels of
osteocalcin, osterix and Runx2, which are essential transcription
factors for osteoblast differentiation (Fig. 2A-C). ODF, OPG and COL1A1 are
essential for the osteoblast differentiation (35). As shown in Fig. 2D-F, alendronate stimulation also
increased the expression levels of ODF, OPG and COL1A1 in
osteoblasts. These results suggest that alendronate regulates
osteoblast differentiation by stimulating the expression levels of
various mRNAs associated with osteoblast differentiation.
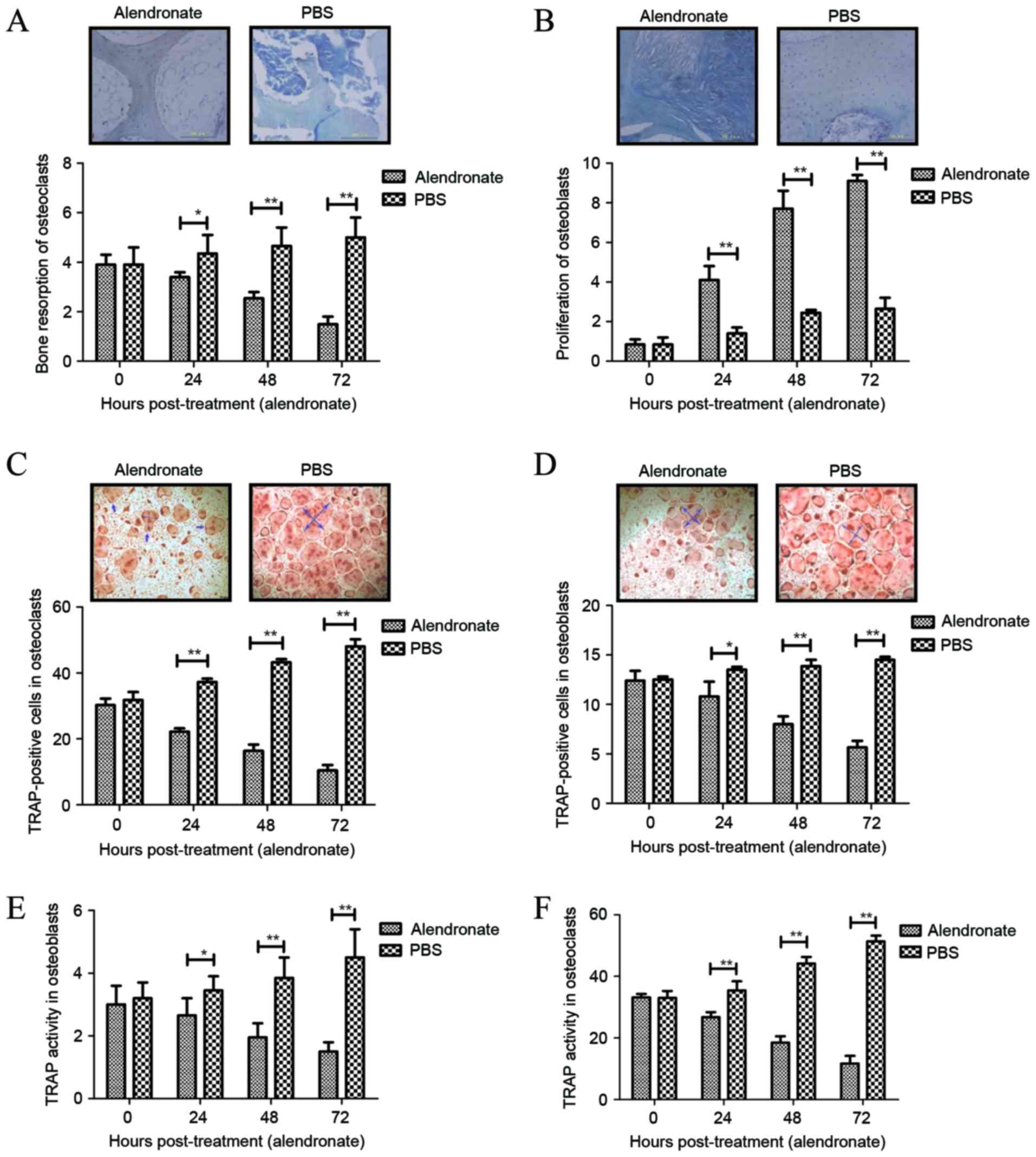
Resorption ability and TRAP activity
of osteoclasts following treatment with alendronate
The resorption ability and TRAP activity of
osteoclasts are essential for the progression of rarefaction of
bone and function of bone tissue (36). The present study results demonstrated
that alendronate treatment significantly inhibited the resorption
ability of osteoclasts (Fig. 3A),
but promoted the proliferation of osteoblasts during the
differentiation period, as determined by toluidine blue staining
(Fig. 3B). In addition, the TRAP
activity of osteoclasts and osteoblasts following treatment with
alendronate during the differentiation period was examined, and the
results indicated that TRAP-positive cells were inhibited in
osteoclasts and osteoblasts following treatment (Fig. 3C and D). Furthermore, TRAP activity
in osteoclasts and osteoblasts was suppressed by the treatment of
alendronate during the differentiation period when compared with
non-treated cells (Fig. 3E and F).
Taken together, these findings suggest that alendronate treatment
markedly inhibits the resorption ability and TRAP activity of
osteoclasts during the differentiation period, which may be
beneficial for the treatment of osteoporosis.
Alendronate regulates osteoblast
differentiation through the IFN-β/STAT1 signaling pathway
In order to analyze the molecular mechanism of
alendronate-mediated osteoblast differentiation, the IFN-β/STAT1
signaling pathway was investigated in osteoblasts. The results
shown in Fig. 4A reveal that
alendronate promoted IFN-β expression in osteoblasts. It was also
identified that STAT1 and pSTAT1 were enhanced in osteoblasts after
treatment with alendronate (Fig.
4B). Alendronate treatment also increased the mRNA expression
levels of Fra1, TRAF6 and SOCS1 in osteoblasts compared with the
control group (Fig. 4C). However,
endogenous inhibition of IFN-β (EI-IFN-β) markedly reduced the
inhibitory effects of alendronate (AEI-IFN-β) on osteoblast
differentiation and differentiation-associated gene expression
levels (Fig. 4D). In addition,
inhibition of IFN-β expression (EI-IFN-β) downregulated of STAT1
and pSTAT1 protein levels and abolished their upregulation induced
by alendronate (AEI-IFN-β) in osteoblasts (Fig. 4E), which also reduced
alendronate-promoted (AEI-IFN-β) expression levels of
differentiation-associated genes, including osteocalcin, osterix
and Runx2 (Fig. 4F). Taken together,
these results revealed that alendronate treatment regulates
osteoblast differentiation through upregulation of the IFN-β/STAT1
signaling pathway.
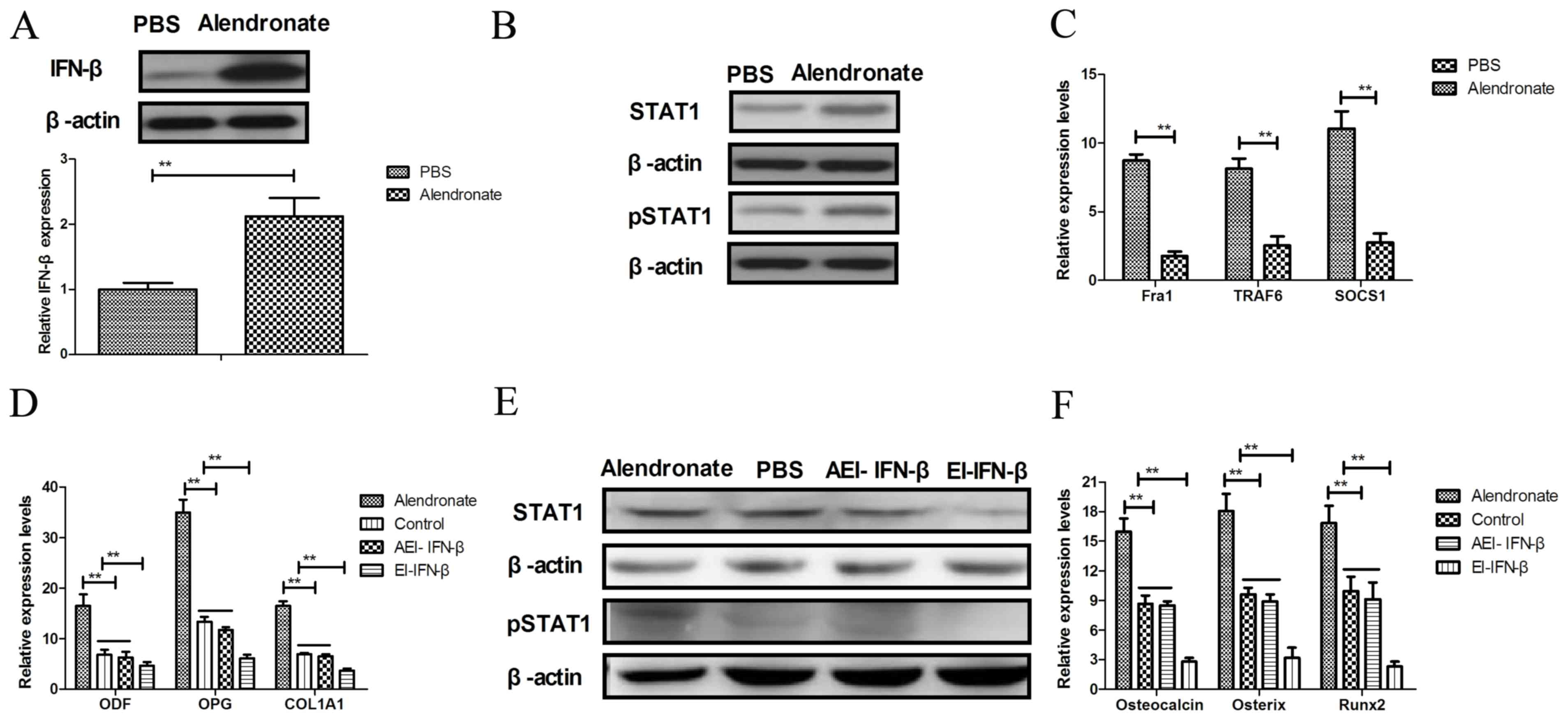 | Figure 4.Alendronate regulates osteoblast
differentiation through the IFN-β/STAT1 signaling pathway. (A)
Alendronate promoted IFN-β protein expression levels in
osteoblasts. (B) Protein expression levels of STAT1 and pSTAT1 in
osteoblasts following treatment with alendronate. (C) Analysis of
mRNA expression levels of Fra1, TRAF6 and SOCS1 in osteoblasts.
Inhibition of IFN-β expression suppresses (D) ODF, OPG and COL1A1
expression levels, (E) STAT1 and pSTAT1 protein expression in
osteoblasts, and (F) the differentiation-associated gene expression
levels of osteocalcin, osterix and Runx2. The data are presented as
the mean ± standard error. **P<0.01 alendronate vs. control
(PBS) group. PBS, phosphate-buffered saline; IFN-β, interferon-β;
STAT1, signal transducer and activator of transcription 1; Fra1,
Fos-related antigen 1; TRAF6, TNF-receptor-associated factor 6;
SOCS1, suppressor of cytokine signaling 1; ODF, osteoblast
differentiation factor; OPG, osteoprotegerin; COL1A1, collagen 1A1;
Runx2, Runt-related transcription factor 2; EI-IFN-β, endogenous
inhibition of IFN-β; AEI, alendronate + EI-IFN-β. |
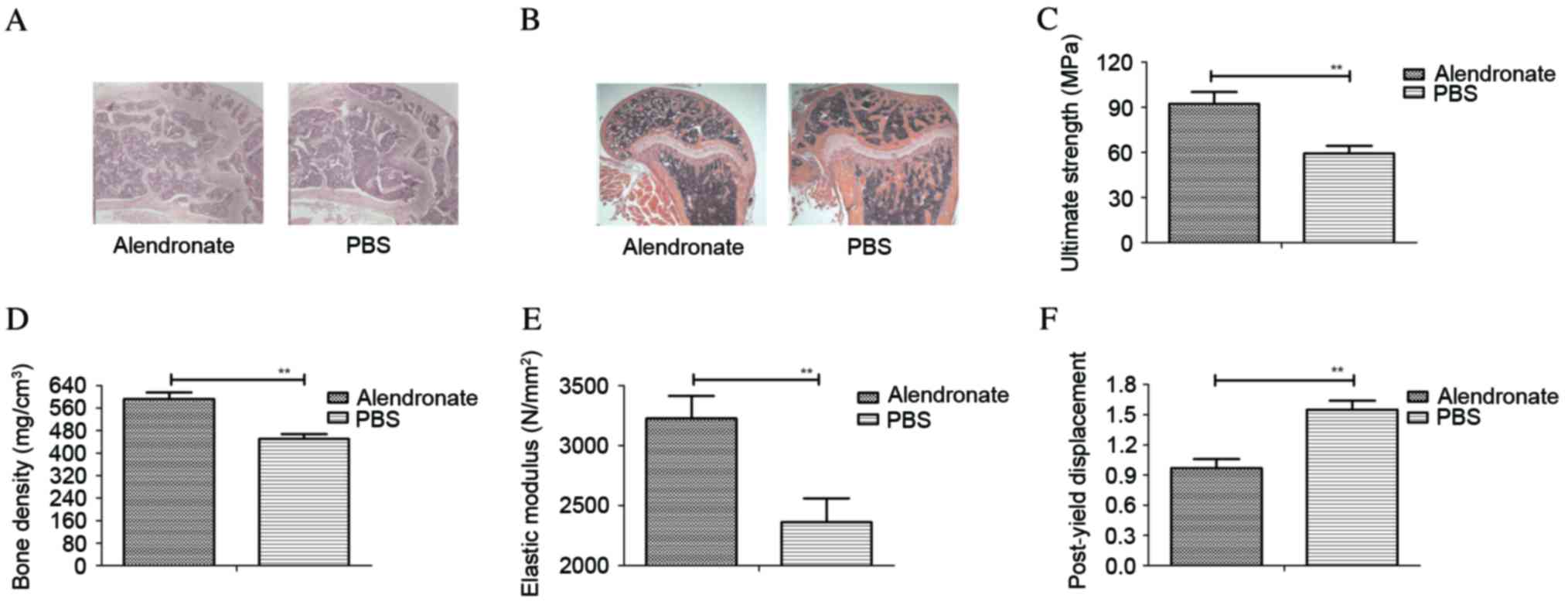
Alendronate treatment exerts a
beneficial effect in rats with ovariectomy-induced osteoporosis,
determined by histological
The current study further analyzed the therapeutic
effects of alendronate on rats with ovariectomy-induced
osteoporosis, which was determined by histological analysis of
experimental rats on day 60. The results in Fig. 5A revealed that bone structure was
significantly improved in rats treated with alendronate that
exhibited ovariectomy-induced osteoporosis compared with the PBS
group. It was also observed that alendronate treatment prevented
loss of trabeculae in rats with osteoporosis induced by ovariectomy
(Fig. 5B). In addition, bone
strength and bone density assays demonstrated that alendronate
significantly improved osteoporosis in rats induced by ovariectomy,
when compared with the PBS control group (Fig. 5C and D). Furthermore, the elastic
modulus and post-yield displacement of bone were also increased in
rats with osteoporosis following treatment with alendronate
(Fig. 5E and F). Therefore, these
data suggest that alendronate treatment was beneficial in rats with
osteoporosis induced by ovariectomy, as determined by histological
and index detection.
Discussion
Osteoporosis is identified as a systemic skeletal
disease that affects mostly postmenopausal women characterized by
reduction of bone strength and mass loss of bone mineral density
(37,38). Evidence suggested that postmenopausal
women have increased risk of osteoporosis, resulting in fragility
fracture due to the lack of estrogen secretion (39,40).
Previous studies have also demonstrated that alendronate (a
bisphosphonate) is an efficient agent for the treatment of
osteoporosis, functioning through inhibition of bone resorption by
accumulating on the bone surface and inhibition of osteoclast
apoptosis (41,42). In the present study, the therapeutic
efficacy and potential underlying mechanism of alendronate
treatment were investigated in a rat model of osteoporosis induced
by ovariectomy. The presented results indicated that alendronate
therapy promoted osteoblast differentiation in the calvarial
osteoblastic cells isolated from newborn rats. In addition, the
resorption ability and TRAP activity of osteoclasts were inhibited
following treatment with alendronate, which may contribute to bone
remodeling in rats with osteoporosis. Alendronate treatment
stimulated ALP expression and activity, as well as the expression
levels of mRNAs associated with osteoblast differentiation,
including Fra1, TRAF6 and SOCS1. The expression levels of ODF, OPG
and COL1A1 in osteoblasts were also upregulated in osteoblasts
subsequent to alendronate administration. Furthermore, the findings
of the current study indicated that alendronate treatment regulated
the osteoblast differentiation through upregulation of IFN-β/STAT1
signaling pathway.
A previous study has demonstrated that stimulation
with IFN-β leads to significant inhibition of osteoporotic
osteoclasts (20). IFN-β production
is mainly secreted by fibroblasts, and subsequently binds to the
IFN-α/β receptor (43,44). The activity of IFN-β depends on the
transcriptional activator of the expression and phosphorylation of
STAT1 and STAT2, which leads to the activation of the JAK signal
pathway (45). Studies also
indicated that IFN-β/STAT1 signaling pathway regulates numerous
metabolism disorder-associated molecular transcriptions (46–48).
Additionally, IFN-β functions as a potential drug for multiple
sclerosis and exhibits various beneficial clinical outcomes
(49,50). In the present study, the involvement
of the IFN-β/STAT1 signaling pathway in osteoblast differentiation
was analyzed. The results indicated that IFN-β/STAT1 signaling
pathway was enhanced in osteoporotic osteoblasts, resulting in the
increase of Fra1, TRAF6 and SOCS1 expression levels following
treatment with alendronate. Furthermore, alendronate increased the
differentiation-associated gene expression levels of osteocalcin,
osterix and Runx2 through upregulation of IFN-β and enhanced the
phosphorylated production of IFN-β. However, inhibition of IFN-β
expression also suppressed STAT1 and pSTAT1 expression in
osteoblasts. These findings indicated that alendronate mediated
improvements in osteoporosis through regulation of IFN-β/STAT1
signaling pathway in osteoblasts.
Bone strength and bone density are the most
important indicators in the progression of patients with
osteoporosis (51,52). Various types of treatments in
ovariectomized rats have been investigated in a large number of
studies (53–55). In addition, histological analysis of
bone qualities is essential for examining the improvement of
osteoporosis, which is regarded as an evaluation criterion for
drugs used in the treatment of osteoporosis (56,57). In
the present study, the therapeutic effects of alendronate-mediated
improvements of osteoporosis were investigated by histological
analysis, including examining the bone density and loss of
trabeculae. The present study findings also indicated that bone
strength, bone density, elastic modulus and post-yield displacement
were significantly improved following treatment with alendronate in
rats with osteoporosis induced by ovariectomy.
In conclusion, although numerous reports have
provided important evidence for identifying the efficacy of
alendronate treatment in osteoblast differentiation, the underlying
molecular mechanism remains poorly understood (58,59). In
the present study, it was demonstrated that alendronate treatment
not only presents stimulatory effects on osteoblast differentiation
and mineralization, but also enhances the bone formation in rats
with osteoporosis induced by ovariectomy. The findings also
revealed that alendronate improves bone loss of osteoporosis
through upregulation of the IFN-β/STAT1 signaling pathway, which
suggests that alendronate may be a potential therapeutic agent for
osteoporosis.
References
|
1
|
Wong SK, Chin KY, Suhaimi FH, Ahmad F and
Ima-Nirwana S: The Relationship between metabolic syndrome and
osteoporosis: A Review. Nutrients. 8:pii: E3472016. View Article : Google Scholar
|
|
2
|
Petty SJ, Wilding H and Wark JD:
Osteoporosis associated with epilepsy and the use of
anti-epileptics-a review. Curr osteoporos Rep. 14:54–65. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Newman M, Lowe C Minns and Barker K:
Spinal orthoses for vertebral osteoporosis and osteoporotic
vertebral fracture: A systematic review. Arch Phys Med Rehabil.
97:1013–1025. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gennari L, Merlotti D, De Paola V, Calabrò
A, Becherini L, Martini G and Nuti R: Estrogen receptor gene
polymorphisms and the genetics of osteoporosis: A HuGE review. Am J
Epidemiol. 161:307–320. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bushardt RL, Turner JL, Ragucci KR and
Askins DG Jr: Non-estrogen treatments for osteoporosis: An
evidence-based review. JAAPA. 19:25–30. 2006.PubMed/NCBI
|
|
6
|
Ziablitsev DS and Larin OS: Influence of
single nucleotide polymorphisms of vitamin D receptor-gene on the
level of osteoassociated hormones linkage with postmenopausal
osteoporosis. Fiziol Zh. 61:21–27. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dempster DW, Zhou H, Recker RR, Brown JP,
Bolognese MA, Recknor CP, Kendler DL, Lewiecki EM, Hanley DA, Rao
SD, et al: A longitudinal study of skeletal histomorphometry at 6
and 24 months across four bone envelopes in postmenopausal women
with osteoporosis receiving teriparatide or zoledronic acid in the
SHOTZ Trial. J Bone Miner Res. 31:1429–1439. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Iba K, Sonoda T, Takada J, Dohke T and
Yamashita T: Further significant effects of eldecalcitol on bone
resorption markers and bone mineral density in postmenopausal
osteoporosis patients having undergone long-term bisphosphonate
treatment. J Bone Miner Metab. 35:171–176. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Darba J, Kaskens L, Vilela F Sorio and
Lothgren M: Cost-utility of denosumab for the treatment of
postmenopausal osteoporosis in Spain. Clinicoecon Outcomes Res.
7:105–117. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Inderjeeth CA, Glendenning P, Ratnagobal
S, Inderjeeth DC and Ondhia C: Long-term efficacy, safety, and
patient acceptability of ibandronate in the treatment of
postmenopausal osteoporosis. Int J Womens Health. 7:7–17. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang ZB, Wan SL, Lu YJ, Ning L, Liu C and
Fan SW: Does vitamin K2 play a role in the prevention and treatment
of osteoporosis for postmenopausal women: A meta-analysis of
randomized controlled trials. Osteoporos Int. 26:1175–1186. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ouyang L, Zhang Q, Ruan X, Feng Y and Wang
X: Treatment effect of Bushen Huayu extract on postmenopausal
osteoporosis in vivo. Exp Ther Med. 7:1687–1690. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhdan VM, Kitura O, Kitura IeM, Babanina M
and Tkachenko MV: Treatment of postmenopausal osteoporosis in the
general medical practice (clinical case). Lik Sprava. 1–89.
2013.
|
|
14
|
Massafra U, Integlia D, Broccoli S and
Migliore A: Mixed treatment comparison to rank antiresorptive
agents in preventing new non vertebral fractures in postmenopausal
osteoporosis. Value Health. 18:A6362015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hassler N, Gamsjaeger S, Hofstetter B,
Brozek W, Klaushofer K and Paschalis EP: Effects of long-term
alendronate treatment on postmenopausal osteoporosis bone material
properties. Osteoporos Int. 26:339–352. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Igase M, Kohara K, Tabara Y, Ohara M,
Takita R, Ochi M, Okada Y and Miki T: Change in arterial stiffness
associated with monthly bisphosphonate treatment in women with
postmenopausal osteoporosis. Menopause. 21:962–966. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Celer O, Akalin A and Oztunali C: Effect
of teriparatide treatment on endothelial function, glucose
metabolism and inflammation markers in patients with postmenopausal
osteoporosis. Clin Endocrinol (Oxf). 85:556–560. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rugpolmuang L and Waikakul S: Effect of a
short-term treatment with once-a-week medication of alendronate 70
Mg on bone turnover markers in postmenopausal women with
osteoporosis. J Med Assoc Thai. 98 Suppl 8:S70–S75. 2015.PubMed/NCBI
|
|
19
|
Schultz TC, Valenzano JP, Verzella JL and
Umland EM: Odanacatib: An emerging novel treatment alternative for
postmenopausal osteoporosis. Womens Health (Lond). 11:805–814.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Seeliger C, Schyschka L, Kronbach Z,
Wottge A, van Griensven M, Wildemann B and Vester H: Signaling
pathway STAT1 is strongly activated by IFN-β in the pathogenesis of
osteoporosis. Eur J Med Res. 20:12015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen XD, Xiao P, Lei SF, Liu YZ, Guo YF,
Deng FY, Tan LJ, Zhu XZ, Chen FR, Recker RR and Deng HW: Gene
expression profiling in monocytes and SNP association suggest the
importance of the STAT1 gene for osteoporosis in both Chinese and
Caucasians. J Bone Miner Res. 25:339–355. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kushwaha P, Khedgikar V, Ahmad N, Karvande
A, Gautam J, Kumar P, Maurya R and Trivedi R: A neoflavonoid
dalsissooal isolated from heartwood of Dalbergia sissoo Roxb. Has
bone forming effects in mice model for osteoporosis. Eur J
Pharmacol. 788:65–74. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Papamanthos M, Varitimidis S, Dailiana ZH,
Kogia E and Malizos K: Computer-assisted evaluation of Mandibular
Cortical Width (MCW) index as an indicator of osteoporosis.
Hippokratia. 18:251–257. 2014.PubMed/NCBI
|
|
24
|
Shum LC, White NS, Nadtochiy SM, Bentley
KL, Brookes PS, Jonason JH and Eliseev RA: Cyclophilin D Knock-out
mice show enhanced resistance to osteoporosis and to metabolic
changes observed in aging bone. PLoS One. 11:e01557092016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cao H, Nazarian A, Ackerman JL, Snyder BD,
Rosenberg AE, Nazarian RM, Hrovat MI, Dai G, Mintzopoulos D and Wu
Y: Quantitative (31)P NMR spectroscopy and (1)H MRI measurements of
bone mineral and matrix density differentiate metabolic bone
diseases in rat models. Bone. 46:1582–1590. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
McCann RM, Colleary G, Geddis C, Clarke
SA, Jordan GR, Dickson GR and Marsh D: Effect of osteoporosis on
bone mineral density and fracture repair in a rat femoral fracture
model. J Orthop Res. 26:384–393. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Leamy LJ, Kelly SA, Hua K, Farber CR and
Pomp D: Quantitative trait loci for bone mineral density and
femoral morphology in an advanced intercross population of mice.
Bone. 55:222–229. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jonason JH and O'Keefe RJ: Isolation and
culture of neonatal mouse calvarial osteoblasts. Methods Mol Biol.
1130:295–305. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saiyed ZM, Sharma S, Godawat R, Telang SD
and Ramchand CN: Activity and stability of alkaline phosphatase
(ALP) immobilized onto magnetic nanoparticles (Fe3O4). J
Biotechnol. 131:240–244. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Subramanian K, Geraerts M, Pauwelyn KA,
Park Y, Owens DJ, Muijtjens M, Ulloa-Montoya F, Jiang Y, Verfaillie
CM and Hu WS: Isolation procedure and characterization of
multipotent adult progenitor cells from rat bone marrow. Methods
Mol Biol. 636:55–78. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Idris AI, Landao-Bassonga E and Ralston
SH: The TRPV1 ion channel antagonist capsazepine inhibits
osteoclast and osteoblast differentiation in vitro and ovariectomy
induced bone loss in vivo. Bone. 46:1089–1099. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hiemer B, Ziebart J, Jonitz-Heincke A,
Grunert PC, Su Y, Hansmann D and Bader R: Magnetically induced
electrostimulation of human osteoblasts results in enhanced cell
viability and osteogenic differentiation. Int J Mol Med. 38:57–64.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xiao S, Wang J and Xiao N: MicroRNAs as
noninvasive biomarkers in bladder cancer detection: A diagnostic
meta-analysis based on qRT-PCR data. Int J Biol Markers.
31:e276–e285. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wai-Hoe L, Wing-Seng L, Ismail Z and
Lay-Harn G: SDS-PAGE-based quantitative assay for screening of
kidney stone disease. Biol Proced Online. 11:145–160. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Johnson RW, White JD, Walker EC, Martin TJ
and Sims NA: Myokines (muscle-derived cytokines and chemokines)
including ciliary neurotrophic factor (CNTF) inhibit osteoblast
differentiation. Bone. 64:47–56. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Takahashi M, Kushida K, Kawana K, Hoshino
H and Inoue T: Discrimination ability of pyridinoline crosslinks
related markers for bone resorption in postmenopause and
osteoporosis. Endocr Res. 23:105–117. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ma J, Ma Y, Liu X, Chen S, Liu C, Qin A
and Fan S: Gambogic acid inhibits osteoclast formation and
ovariectomy-induced osteoporosis by suppressing the JNK, p38 and
Akt signalling pathways. Biochem J. 469:399–408. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Song SH, Wang D, Mo YY, Ding C and Shang
P: Antiosteoporotic effects of naringenin on ovariectomy-induced
osteoporosis in rat. Yao Xue Xue Bao. 50:154–161. 2015.(In
Chinese). PubMed/NCBI
|
|
39
|
Lee SN, Cho JY, Eun YM, Song SW and Moon
KW: Associations between osteoporosis and coronary artery disease
in postmenopausal women. Climacteric. 19:458–462. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Alami S, Hervouet L, Poiraudeau S, Briot K
and Roux C: Barriers to effective postmenopausal osteoporosis
treatment: A qualitative study of patients' and practitioners'
views. PLoS One. 11:e01583652016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Karlsson L, Lundkvist J, Psachoulia E,
Intorcia M and Ström O: Persistence with denosumab and persistence
with oral bisphosphonates for the treatment of postmenopausal
osteoporosis: A retrospective, observational study, and a
meta-analysis. Osteoporos Int. 26:2401–2411. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kim MJ, Kim SN, Lee IS, Chung S, Lee J,
Yang Y, Lee I and Koh SE: Effects of bisphosphonates to treat
osteoporosis in children with cerebral palsy: A meta-analysis. J
Pediatr Endocrinol Metab. 28:1343–1350. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Diringer MN, Dhar R, Scalfani M, Zazulia
AR, Chicoine M, Powers WJ and Derdeyn CP: Effect of high-dose
simvastatin on cerebral blood flow and static autoregulation in
subarachnoid hemorrhage. Neurocrit Care. 25:56–63. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dauletbaev N, Cammisano M, Herscovitch K
and Lands LC: Stimulation of the RIG-I/MAVS Pathway by
polyinosinic: Polycytidylic acid upregulates IFN-β in airway
epithelial cells with minimal costimulation of IL-8. J Immunol.
195:2829–2841. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Grumbach IM, Fish EN, Uddin S, Majchrzak
B, Colamonici OR, Figulla HR, Heim A and Platanias LC: Activation
of the Jak-Stat pathway in cells that exhibit selective sensitivity
to the antiviral effects of IFN-beta compared with IFN-alpha. J
Interferon Cytokine Res. 19:797–801. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Imaizumi T, Yoshida H, Hayakari R, Xing F,
Wang L, Matsumiya T, Tanji K, Kawaguchi S, Murakami M and Tanaka H:
Interferon-stimulated gene (ISG) 60, as well as ISG56 and ISG54,
positively regulates TLR3/IFN-β/STAT1 axis in U373MG human
astrocytoma cells. Neurosci Res. 105:35–41. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ma JS, Kim WJ, Kim JJ, Kim TJ, Ye SK, Song
MD, Kang H, Kim DW, Moon WK and Lee KH: Gold nanoparticles
attenuate LPS-induced NO production through the inhibition of
NF-kappaB and IFN-beta/STAT1 pathways in RAW264.7 cells. Nitric
Oxide. 23:214–219. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yun CH, Yang JS, Kang SS, Yang Y, Cho JH,
Son CG and Han SH: NF-kappaB signaling pathway, not IFN-beta/STAT1,
is responsible for the selenium suppression of LPS-induced nitric
oxide production. Int immunopharmacol. 7:1192–1198. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bustamante MF, Morcillo-Suárez C, Malhotra
S, Rio J, Leyva L, Fernández O, Zettl UK, Killestein J, Brassat D,
García-Merino JA, et al: Pharmacogenomic study in patients with
multiple sclerosis: Responders and nonresponders to IFN-β. Neurol
Neuroimmunol Neuroinflamm. 2:e1542015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Damiano S, Sasso A, De Felice B,
Terrazzano G, Bresciamorra V, Carotenuto A, Orefice NS, Orefice G,
Vacca G, Belfiore A, et al: The IFN-β 1b effect on Cu Zn superoxide
dismutase (SOD1) in peripheral mononuclear blood cells of
relapsing-remitting multiple sclerosis patients and in
neuroblastoma SK-N-BE cells. Brain Res Bull. 118:1–6. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Takahashi T, Watanabe T, Nakada H,
Tanimoto Y, Kimoto S, Mijares DQ, Zhang Y and Kawai Y: Effect of a
dietary supplement on peri-implant bone strength in a rat model of
osteoporosis. J Prosthodont Res. 60:131–137. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li X, Niu QT, Warmington KS, Asuncion FJ,
Dwyer D, Grisanti M, Han CY, Stolina M, Eschenberg MJ, Kostenuik
PJ, et al: Progressive increases in bone mass and bone strength in
an ovariectomized rat model of osteoporosis after 26 weeks of
treatment with a sclerostin antibody. Endocrinology. 155:4785–4797.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Brennan O, Kuliwaba JS, Lee TC, Parkinson
IH, Fazzalari NL, McNamara LM and O'Brien FJ: Temporal changes in
bone composition, architecture, and strength following estrogen
deficiency in osteoporosis. Calcif Tissue Int. 91:440–449. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ying SH, Kao HC, Chang MC, Yu WK, Wang ST
and Liu CL: Fixation strength of PMMA-augmented pedicle screws
after depth adjustment in a synthetic bone model of osteoporosis.
Orthopedics. 35:e1511–e1516. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Simon JA, Recknor C, Moffett AH Jr, Adachi
JD, Franek E, Lewiecki EM, McClung MR, Mautalen CA, Ragi-Eis S,
Nicholson GC, et al: Impact of denosumab on the peripheral skeleton
of postmenopausal women with osteoporosis: Bone density, mass, and
strength of the radius, and wrist fracture. Menopause. 20:130–137.
2013.PubMed/NCBI
|
|
56
|
Mashiba T: Morphological analysis of bone
dynamics and metabolic bone disease. Histological findings in
animal fracture model-effects of osteoporosis treatment drugs on
fracture healing process. Clin Calcium. 21:551–558. 2011.(In
Japanese).
|
|
57
|
Delling G, Ritzel H and Werner M:
Histological characteristics and prevalence of secondary
osteoporosis in systemic mastocytosis. A retrospective analysis of
158 cases. Pathologe. 22:132–140. 2001.(In German).
|
|
58
|
Saag KG, Agnusdei D, Hans D, Kohlmeier LA,
Krohn KD, Leib ES, MacLaughlin EJ, Alam J, Simonelli C, Taylor KA
and Marcus R: Trabecular bone score in patients with chronic
glucocorticoid-induced osteoporosis treated with alendronate or
teriparatide. Arthritis Rheumatol. 68:2122–2128. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ryu TK, Kang RH, Jeong KY, Jun DR, Koh JM,
Kim D, Bae SK and Choi SW: Bone-targeted delivery of
nanodiamond-based drug carriers conjugated with alendronate for
potential osteoporosis treatment. J Control Release. 232:152–160.
2016. View Article : Google Scholar : PubMed/NCBI
|