Introduction
In China, the number of patients with avascular
necrosis of the femoral head (ANFH) is estimated to be between 5
and 7 million and there are 100,000–150,000 new cases each year
(1). At present, gene therapy is a
large area of ANFH research (2).
Basic fibroblast growth factor (bFGF), transforming growth factor-β
(TGF-β), bone morphogenetic proteins (BMPs), and vascular
endothelial growth factor (VEGF) all serve important roles in the
development of applicable gene therapies, in particular BMPs and
VEGF (3–5). Bone morphogenetic proteins (BMPs) are
signal molecules that promote bone regeneration (6–8). As
important members of the TGF-β superfamily, BMPs have effects on
the stimulation of osteoblast differentiation (9,10) and
this process involves the differentiation of progenitor cells into
chondrocytes and osteoblasts, endochondral ossification and new
bone formation (11). Studies have
revealed that BMP-2, BMP-4, BMP-6, BMP-7 and BMP-9 are crucial for
bone formation (12–15). Furthermore, a number of studies have
suggested that BMP-6 is the most potent osteoinductive BMP and has
greater potential for bone regeneration than BMP-2 and BMP-4
(16–18). Angiogenesis is the most basic
physiological process in bone repair (19); furthermore, VEGF, which is expressed
by endothelial cells, is one of the most important cytokines in
angiogenesis (20) and is associated
with all steps of bone formation, including mesenchymal
condensation, cartilage formation, cartilage resorption and blood
vessel invasion (21,22). It has been indicated that BMPs
stimulate angiogenesis via the osteoblast production of VEGF-A,
which plays an important role in bone formation and angiogenesis by
acting as a chemoattractant (23). A
number of studies have attempted to determine the effects of
combining VEGF with BMP in order to encourage bone formation. One
such study reported that the combination of VEGF and BMP-4 enhances
bone formation (24,25). Another revealed that, when combined,
VEGF and BMP-2 are effective in stimulating bone regeneration
(26). In addition, the
co-administration of VEGF and BMP-4 has been reported to induce
osteogenesis more effectively than VEGF and BMP-2 (27). These studies have focused on the
delivery of BMP-2 and BMP-4 in conjunction with VEGF. However, to
the best of our knowledge, few studies have focused on the effects
of the combination of VEGF and BMP-6 on bone repair.
In the present study, adeno-associated viruses (AAV)
co-expressing VEGF and BMP-6 were constructed to investigate the
potential of bone mesenchymal stem cells (BMSCs) treated with a
combination of VEGF and BMP-6 genes for the treatment of ANFH.
Materials and methods
Ethics statement
The animal study was approved by the First
Affiliated Hospital of Sun Yat-sen University (Guangzhou,
China).
Isolation, culture, and verification
of rat BMSCs
A total of 20 healthy 3–4 weeks specific pathogen
free grade Sprague Dawley rats (weighing 100–120 g, 10 males and 10
females) were purchased from the Animal Center of Sun Yat-Sen
University (Guangzhou, China). The rats were housed in an
environment of 25±5°C with 35±5% humidity and a standard 12 h
light/dark cycle and ad libitum access to food and water.
After three days, the rats were sacrificed by intraperitoneal
injection of 300 mg/kg pentobarbital, soaked in 75% ethanol for 10
min and then the tibiae and femora were isolated under sterile
condition. BMSCs were obtained from the tibiae and femora of the
rats following sacrifice. Density gradient centrifugation and
adherent screening were used to isolate BMSCs as previously
described (28). In short, tibiae
and femora were flushed with low glucose Dulbecco's modified
Eagle's medium (L-DMEM, GE Healthcare Life Sciences, Logan, UT,
USA) to harvest the BMSCs, which were cultured in L-DMEM
supplemented with 10% fetal bovine serum (FBS, Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and 20 mg/ml
penicillin-streptomycin (Gibco; Thermo Fisher Scientific, Inc.).
Cells were subsequently incubated with 5% CO2 at 37°C
for 7 days and split, following which cells at the fourth passage
were used for the following experiments. The obtained cells were
added into Percoll separation solution (Sigma Aldrich: Merck KGaA,
Darmstadt, Germany) with a density of 1.073 g/ml and centrifuged at
4,000 × g for 25 min at 20°C. The cells were further resuspended in
complete medium containing 90% L-DMEM, 10% FBS, 100 µl/ml
penicillin and 100 µl/ml streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) and incubated at 37°C in a 5% CO2
supplemented incubator (Thermo Fisher Scientific. Company, USA).
Cell surface antigens were used to identify BMSC characteristics
via flow cytometry. Briefly, cells were collected and washed twice
with PBS (Gibco; Thermo Fisher Scientific, Inc.) and then incubated
with 0.5% BSA blocking buffer in 1× PBS (Thermo Fisher Scientific,
Inc.) for 30 min at 4°C. Following washing with PBS, cells were
immunofluorescently stained with fluorochrome-conjugated antibodies
specific to the cell surface antigens cluster of differentiation
CD90 (cat no. ab225; 1:200, Abcam, Cambridge, UK), CD29 (cat no.
bs-20631R; 1:200; Bioss, Beijing, China), CD44 (cat no. bs-0521R;
1:200, Bioss), CD11 (cat no. bs-2508R; 1:200; Bioss), CD45 (cat no.
bs-0522R; 1:200; Bioss) and CD34 (cat no. ab81289; 1:200, Abcam) at
4°C for 30 min. Flow cytometry was performed on a FACSCanto II flow
cytometer (BD Biosciences, Franklin Lakes, NJ, USA) using FACSDiva
software (BD Biosciences) and the data was analyzed by FlowJo
software version 10.06 (Tree Star, Inc., Ashland, OR, USA). Each
surface antigen assay was performed in triplicate.
Adeno-associated virus type 2 (AAV)
vector production and infection in vitro
The recombinant vector was packaged by the
adeno-associated virus (AAV) Helper-Free System (Hanbio
Biotechnology Co., Ltd, Shanghai, China). The internal ribosome
entry site (IRES) fragment was incorporated into the plasmid AAV
multiple cloning site (pAAV MCS) (Hanbio Biotechnology Co., Ltd)
and sub-cloned into two multiple cloning sites (Hanbio
Biotechnology Co., Ltd). Next, VEGF and BMP-6 primers were designed
by Primer Premier 5 software (Premier Biosoft Inc., Palo Alto, CA,
USA) and synthesized by Sangon Biotech Company (Shanghai, China),
then VEGF and BMP-6 were inserted into the upstream and downstream
MCS, respectively. The bicistronic frame was 2.5 kb in length,
which is within the vector capacity. The recombinant
(r)AAV-VEGF-IRES-BMP-6 (AAV-VEGF-BMP-6), rAAV-VEGF-green
fluorescent protein (GFP) (AAV-VEGF), rAAV-BMP-6-GFP (AAV-BMP-6),
and rAAV-IRES-GFP (AAV-GFP) vectors were co-transfected into
AAV-293 cells (Forevergen, Guangzhou, China) with the plasmid
(p)AAV-helper and pAAV-RC (carrying AAV-2 replication and capsid
genes) using the calcium phosphate method by adding
CaCl2, according to the manufacturer's instructions
(Invitrogen; Thermo Fisher Scientific, Inc.). A primary virus stock
was collected 72 h following transfection and chloroform/PEG8000
protocols as previously described (29) were used to concentrate and purify the
primary virus stock. In order to achieve the greatest
cytopathogenic effect, infection efficiency and cost of the r
virus, it was determined that 5×104 viral particle/cell
was the best multiplication of infection (MOI) for infecting rat
BMSCs (30). The BMSCs
(2×105/ml) were seeded onto 96-well plates and incubated
for 30 min at 37°C in an atmosphere containing 5% CO2.
BMSCs were subsequently transfected with AAV-GFP, AAV-VEGF,
AAV-BMP-6, or AAV-VEGF-BMP-6 vectors at the optimum MOI using
lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
After 48 h, samples were collected for subsequent
experimentation.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from BMSCs infected with the AAV-GFP,
AAV-VEGF, AAV-BMP-6 or AAV-VEGF-BMP-6 viruses was extracted using
TRIzol Reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol and reverse transcribed to
cDNA with Super Script TM III Reverse Transcriptase (Invitrogen;
Thermo Fisher Scientific, Inc.). The target genes were analyzed
using a Takara SYBR® Green I qPCR kit (Takara Bio, Inc.
Otsu, Japan) performed on the ABI 7500 system (Applied Biosystems;
Thermo Fisher Scientific, Inc.) using the following thermocycling
conditions: 94°C for 3 min, 35 cycles at 94°C for 30 sec, 55°C for
30 sec, and 72°C for 45 sec. The primers used were as follows:
GAPDH, forward 5′-CCTCGTCTCATAGACAAGATGGT-3′ and reverse
5′-GGGTAGAGTCATACTGGAACATG-3′; BMP6, forward
5′-ACAGCATAACATGGGGCTTC-3′ and reverse 5′-CTCGGGGTTCATAAGGTGAA-3′;
VEGF, forward 5′-TTGCTGCTCTACCTCCAC-3′ and reverse
5′-AATGCTTTCTCCGCTCTG-3′; osteocalcin (OCN), forward
5′-CTCTGTCTCTCTGACCTCACAG-3′ and reverse
5′-GGAGCTGCTGTGACATCCATAC-3′; runt-related transcription factor 2
(RUNX2), forward 5′-GAAACTCTTGCCTCGTCCGCT-3′ and reverse
5′-GATGATGACACTGCCACCTCTG-3′; alkaline phosphatase (AKP), forward
5′-CTGGTGGAAGGAGGCAGAATT-3′ and reverse
5′-ATGTGAAGACGTGGGAATGGT-3′. The relative mRNA expression was
calculated using the 2−∆∆Cq method (31).
ELISA
A total of 500 µl from each culture supernatant was
harvested from the AAV-GFP, AAV-VEGF, AAV-BMP-6 and AAV-VEGF-BMP-6
transfection groups. VEGF and BMP-6 levels in the culture medium
were examined using ELISA kits (Rat VEGF ELISA kit, cat no. RRV00,
R&D Systems, Minneapolis, MN, USA; and Rat BMP-6 ELISA kit, cat
no. MBS704699, MyBiosource, San Diego, CA, USA) following
manufacturer's protocol. The absorbance was determined at 450 nm
using a microplate reader (Multiskan Go; Thermo Fisher Scientific,
Inc.). The concentrations of VEGF and BMP-6 were determined by
comparing the absorbance with those of the standards.
Western blotting
For protein extraction, cells were lysed in cell
lysis buffer containing 140 mM NaCl, 10 mM Tris-HCl, 1% Triton
X-100, 1 mM ECTA and 1X protease inhibitor cocktail (Gibco; Thermo
Fisher Scientific, Inc.). The protein concentration was determined
using the BCA assay. A total of 20 µg protein was loaded per lane
and separated on 12% SDS-PAGE gels. Proteins were subsequently
transferred to polyvinylidene difluoride membranes, which were
blocked with 5% non-fat dry milk for 1 h at room temperature and
incubated overnight at 4°C with the following primary antibodies:
Anti-GAPDH (cat no. sc-25778; 1:2,000) anti-BMP6 (cat no. sc-7406;
1:100; both Santa Cruz Biotechnology, Inc.) and anti-VEGF (cat no.
ab105219; 1:1,000; Abcam). They were subsequently incubated with an
anti-goat horseradish peroxidase (HRP)-conjugated secondary
antibody (cat no. sc-2004, 1:1,000, Santa Cruz Biotechnology, Inc.)
at 4°C for overnight, and detected with an enhanced
chemiluminescence detection kit (EMD Millipore, Billerica, MA,
USA).
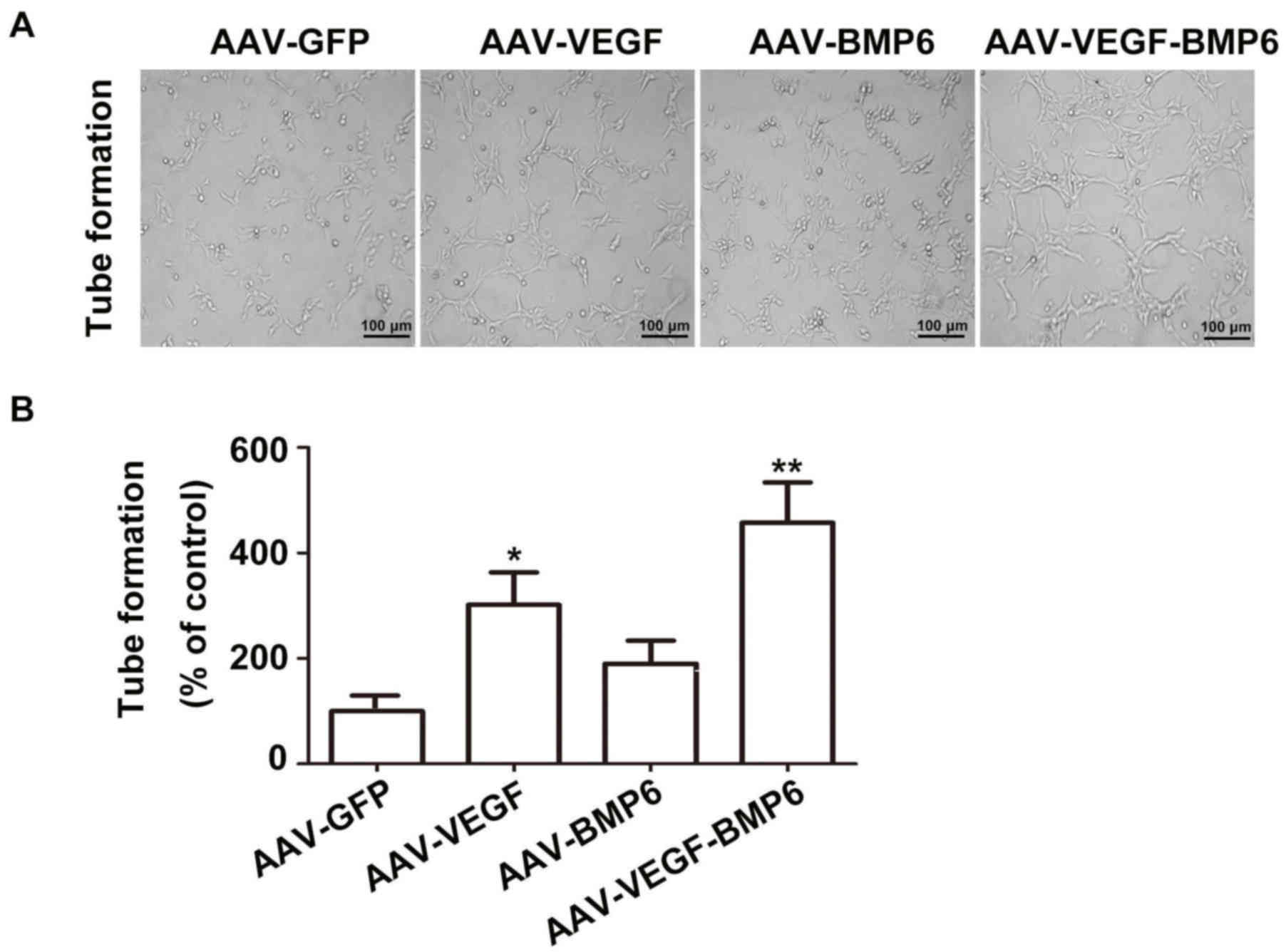
Angiogenic assay in vitro
The basement membrane Matrigel matrix (BD
Biosciences,) was incubated at 37°C for 30 min in a 24-well plate
following dilution with serum-free medium. When solidification
occurred, human umbilical vein epithelial cells (HUVECs;
5×104 cells/well; (Forevergen) were seeded with fresh
L-DMEM medium supplemented with 10% FBS. Tube formation of HUVECs
was observed by co-culture with BMSCs. At 14 days following after
infection, 1 ml of each culture supernatant obtained from AAV-GFP,
AAV-VEGF, AAV-BMP-6 and AAV-VEGF-BMP-6 cells was added to the
24-well plate and incubated at 37°C for 12 h in an atmosphere
containing 5% CO2. Images of tube formation were
acquired from three random fields under a light microscope
(magnification, ×200) and analyzed with Image Pro Plus 6.0 (Media
Cybernetics, Inc., Rockville, MD, USA)
Osteogenic assay in vitro
Following transfection with AAV-GFP, AAV-VEGF,
AAV-BMP-6 or AAV-VEGF-BMP-6, BMSCs were cultured in L-DMEM
supplemented with 10% FBS containing 20 mg/ml
penicillin-streptomycin in an atmosphere containing 5%
CO2 at 37°C for 30 min. During the second week, the
mineralization effects were detected by Von Kossa staining,
alizarin red staining (ARS), and AKP staining. For AKP staining,
cells were washed with PBS, fixed with 4% paraformaldehyde at 4°C
overnight and stained with AKP solution at room temperature for 10
min (Beyotime Institute of Biotechnology). Images of mineral
nodules were captured from three random fields under a light
microscope (magnification, ×200) and subsequently analyzed with
Image Pro Plus 6.0 (Media Cybernetics, Inc.). For ARS staining, ARS
was prepared in double distilled H2O and the pH was
adjusted to 4.1 using 10% (v/v) ammonium hydroxide. Sections were
fixed in 70% ethanol at room temperature for 10 min and then
stained with 40 mM ARS for 5 min at room temperature. Calcium
deposition was assessed by eluting ARS staining with distilled
water including 10% acetic acid and 20% methanol. The absorbance of
supernatants was measured at 405 nm.
In vitro cell seeding on poly
lactide-co-glycolide (PLAGA) scaffolds
The biomimetic synthetic PLAGA scaffolds were
sterilized using 70% ethanol for 1 h and rinsed three times in PBS.
PLAGA scaffolds were then exposed to ultraviolet light for 24 h and
treated with PC-2000 plasma cleaner (South Bay Technology, Inc.,
San Clemente, CA, USA) to enhance cell attachment. Finally, the
cells (1.67×105/ml) were seeded on each PLAGA scaffold.
A total of 12 6 weeks old nude mice (weighing 22–25 g; 6 males and
6 females) were purchased from the Animal Center of Sun Yat-sen
University (Guangzhou, China). The rats were housed in an
environment of 25±5°C with 35±5% humidity and a standard 12 h
light/dark cycle and ad libitum access to food and water.
After one day, the PLAGA scaffolds were implanted subcutaneously
into the nude mice (n=3 per group). Implants were retrieved
following 2 or 3 weeks, embedded in paraffin, sectioned into 5 mm
slices and mounted onto glass slides.
Histological analysis of in vivo bone
formation
Slides were washed in xylene twice for 10 min each
in order to remove the paraffin. A graded series of ethanol
solutions, including 100, 95, and 70% ethanol were used for
rehydration. The sections were washed twice in deionized water for
5 min each. Tissues were fixed in 4% paraformaldehyde at 4°C for
overnight, embedded in paraffin, sliced into sections 4 mm thick
and stained with hematoxylin and eosin (H&E) at room
temperature for 15 min. For Von Kossa staining, sections were
stained with von Kossa stain at room temperature for 20 min to
examine calcium deposition. In addition, immunohistochemistry was
used to examine the expression of VEGF and BMP-6 in implants. Fixed
tissues were deparaffinized, incubated in citrate buffer at room
temperature for 10 min and subsequently incubated with the primary
antibodies, including anti-BMP6 antibody (cat no. ab155963; 1:200;
Abcam) and anti-VEGF antibody (cat no. ab81289; 1:200; Abcam)
overnight at 4°C, followed by biotinylated goat anti-mouse
immunoglobulin G secondary antibodies (cat no. BA1300; 1:500, BD
Biosciences) and treated with streptavidin-horseradish peroxidase
(BD Biosciences). Tissues were then treated with
3,3′-diaminobenzidine substrate and hematoxylin for 15 min at room
temperature. The data was detected using ImageJ software 4.8
(National Institutes of Health, Bethesda, MA, USA).
Analysis of blood vessel
formation
Formalin-fixed paraffin-embedded tissue sections
were immunostained for smooth muscle α-actin in order to detect
blood vessel formation. Following washing in PBS containing 0.1%
(wt/vol) saponin and 2% (wt/vol) bovine serum albumin (Gibco;
Thermo Fisher Scientific, Inc.), CD34 was quantified by
immunohistochemistry in order to detect the number of capillaries.
The sections were labeled for CD34 with CY3-conjugated monoclonal
anti-CD34 antibody (cat no ab81289; 1:200; Abcam) and incubated
overnight at 4°C. Images of tissue sections were subsequently
acquired under a light microscope (magnification, ×200) and
captured using an Olympus MicroFire color digital camera and
PictureFrame image acquisition software 2.0 (Optronics, Goleta, CA,
USA). Histological sections were then examined to quantify the
blood vessel density on the scaffolds (four sections per sample).
Blood vessels were manually counted on the total scaffold and a
circular luminal structure was taken to indicate a blood
vessel.
Statistical analysis
Data were analyzed with the Student's t-test and one
way analysis of variance followed by a Tukey's post hoc test using
SPSS 19.0 software (IBM Corp., Armonk, NY, USA). Each experiment
was repeated at least three times. All results were summarized and
are presented as means ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
Cell verification
The expression of cell surface antigens CD29, CD90,
CD44, CD45, CD11 and CD34 were 97.7, 99.2, 99.0, 2.26, 8.53 and
2.3% positive, respectively (Fig.
1). The cells were demonstrated to high-express CD29, CD90 and
CD44, whereas CD45, CD11 and CD34 were low-expressed, indicating
that the cells were BMSCs rather than hematopoietic cells.
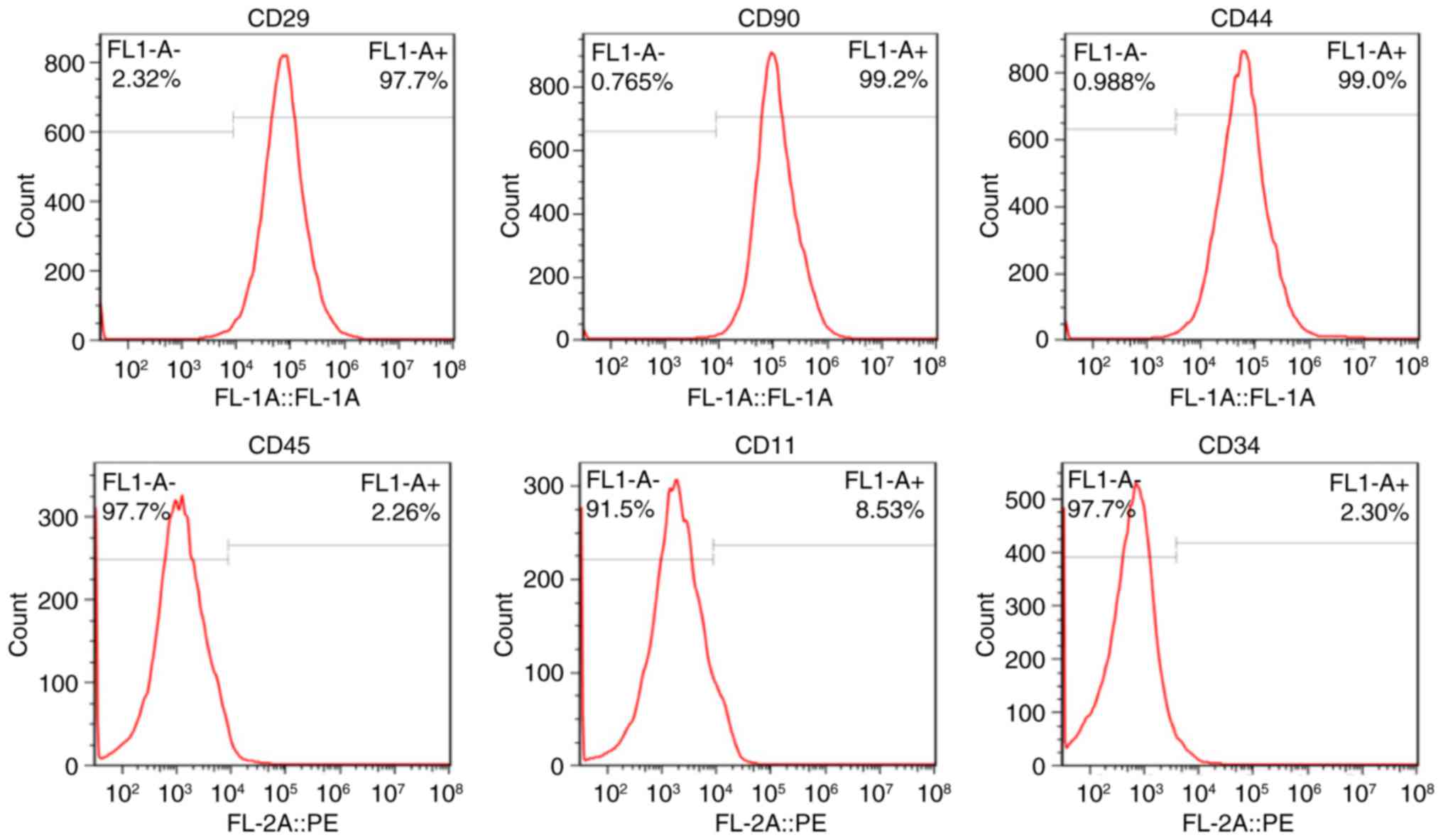 | Figure 1.Representative expression of BMSC
surface markers CD29, CD90, CD44, CD45, CD11 and CD34 as analyzed
by flow cytometry. BMSCs were observed to hyperexpress CD29
(97.7%), CD90 (99.2%) and CD44 (99.0%), whereas CD45 (2.26%), CD11
(8.53%) and CD34 (2.30%) were hypoexpressed. BMSC, bone mesenchymal
stem cell; CD, cluster of differentiation; FL-1,
fluorescence-1. |
Expression of VEGF and BMP-6
The relative mRNA expression of BMP-6, VEGF, OCN,
RUNX2 and AKP was quantified in the BMSCs 6 weeks
post-transfection. VEGF mRNA expression in the AAV-VEGF-BMP-6 and
AAV-VEGF groups was significantly higher compared with the AAV-GFP
group (P<0.05, Fig. 2A). In
addition, BMP-6 mRNA expression in the AAV-VEGF-BMP-6 (P<0.01)
and AAV-BMP-6 (P<0.05) groups were significantly higher compared
with the AAV-GFP group (Fig. 2A).
The expression levels of OCN, RUNX2 and AKP mRNA were all
upregulated in the AAV-BMP-6 and AAV-VEGF-BMP-6 groups compared
with the AAV-GFP group (*P<0.05, **P<0.01, Fig. 2A); Western blotting revealed a
similar pattern for VEGF and BMP-6 protein expression levels, The
expression of BMP6 protein was significantly increased in the
AAV-VEGF-BMP-6 group (P<0.01) and AAV-BMP-6 group (P<0.05)
when compared with the AAV-GFP group Fig. 2B and C). In addition, VEGF expression
was significantly higher in the AAV-VEGF-BMP-6 and AAV-VEGF groups
compared with the AAV-GFP group (both P<0.05, Fig. 2B and C). ELISA was performed to
confirm VEGF and BMP-6 expression. The results revealed that VEGF
and BMP-6 were expressed in all groups. And BMP6 production
increased in the AAV-BMP6 and AAV-VEGF-BMP-6 groups compared with
the AAV-GFP group (P<0.05, P<0.01, Fig. 2D); however, BMP6 production in the
AAV-VEGF-BMP6 group was marked greater than that in AAV-BMP-6 group
(P<0.05, Fig. 2D), which suggests
that VEGF-BMP6 co-expression promotes the secretion of BMP6.
Similarly, VEGF production in the AAV-VEGF and AAV-VEGF-BMP6 groups
was significantly increased compared with the AAV-GFP group
(Fig. 2E).
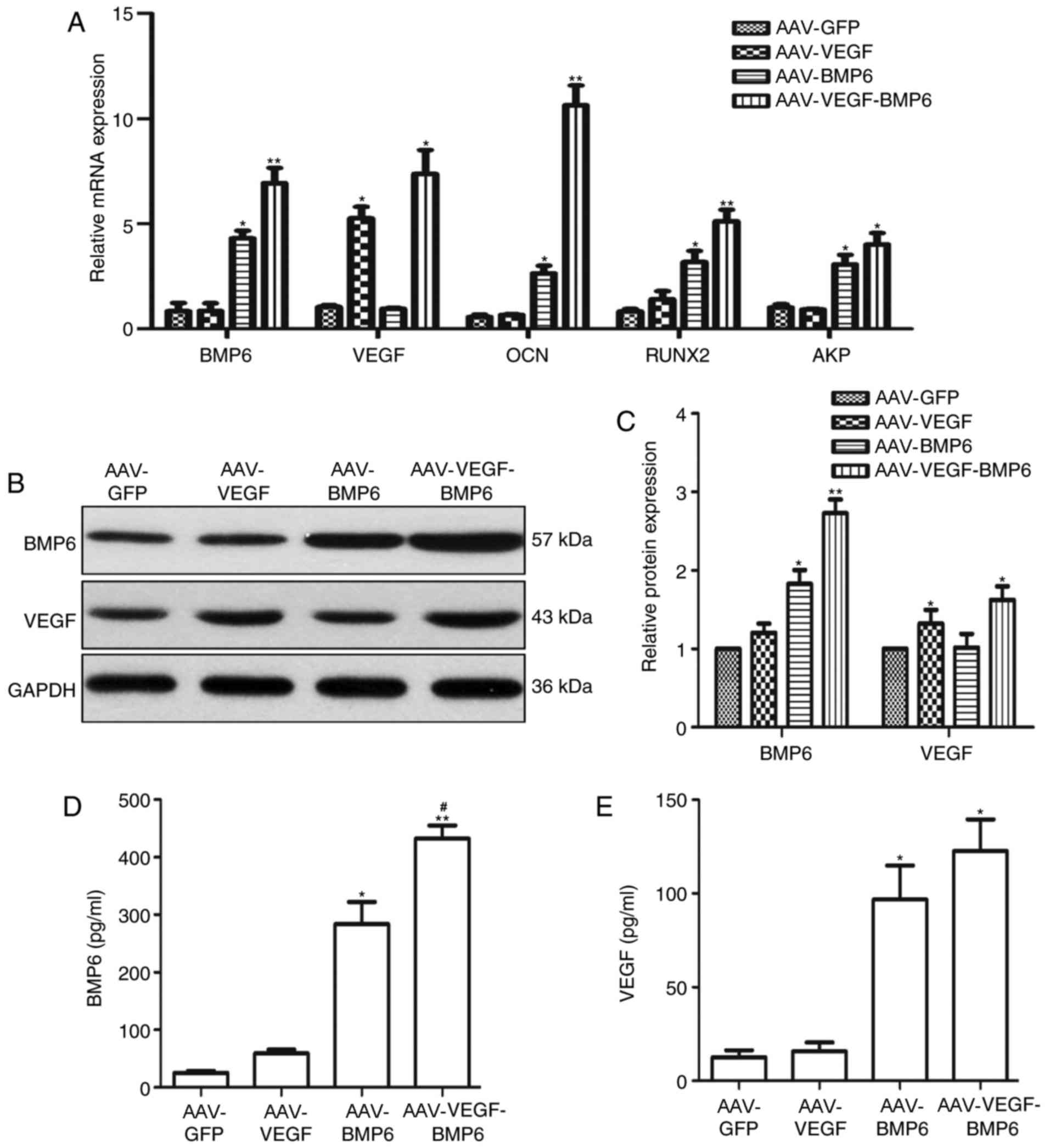 | Figure 2.Levels of VEGF and BMP-6 in the
AAV-GFP, AAV-BMP-6, AAV-VEGF and AAV-VEGF-BMP groups. (A) Relative
mRNA expressions of BMP-6, VEGF, OCN, RUNX2 and AKP analyzed by
reverse transcription-quantitative polymerase chain reaction.
Levels of VEGF and BMP-6 protein in transfected BMSCs were (B)
assessed using western blotting and (C) quantified using
densitometry analysis. GAPDH was used as an internal control. ELISA
kits were used to measure (D) BMP-6 and (E) VEGF protein
expression. *P<0.05 vs. AAV-GFP group, **P<0.01 vs. AAV-GFP
group, #P<0.05 vs. AAV-BMP-6 group. VEGF, vascular
endothelial growth factor; BMP, bone morphogenetic protein; AAV,
adeno-associated virus; GFP, green fluorescent protein; OCN,
osteocalcin; RUNX2, runt-related transcription factor 2; AKP,
alkaline phosphatase; BMSCs, bone mesenchymal stem cells. |
Biological activity of VEGF and BMP6
in vitro
VEGF secreted from BMSCs in the AAV-VEGF-BMP-6 group
enhanced HUVEC proliferation and tube formation in comparison with
the other groups (Fig. 3A). Tube
formation was significantly increased in the AAV-VEGF-BMP-6 and
AAV-VEGF groups compared with the AAV-GFP group (P<0.05,
P<0.01, Fig. 3B). The
mineralization effects of BMP-6 were detected using alizarin red,
AKP and Von Kossa staining (Fig. 4).
The intensity of staining was markedly increased in the
AAV-VEGF-BMP-6 group compared with all other groups, these results
indicated that co-expressing the VEGF and BMP-6 in BMSCs could
enhanced HUVEC tube formation and increased osteogenic ability.
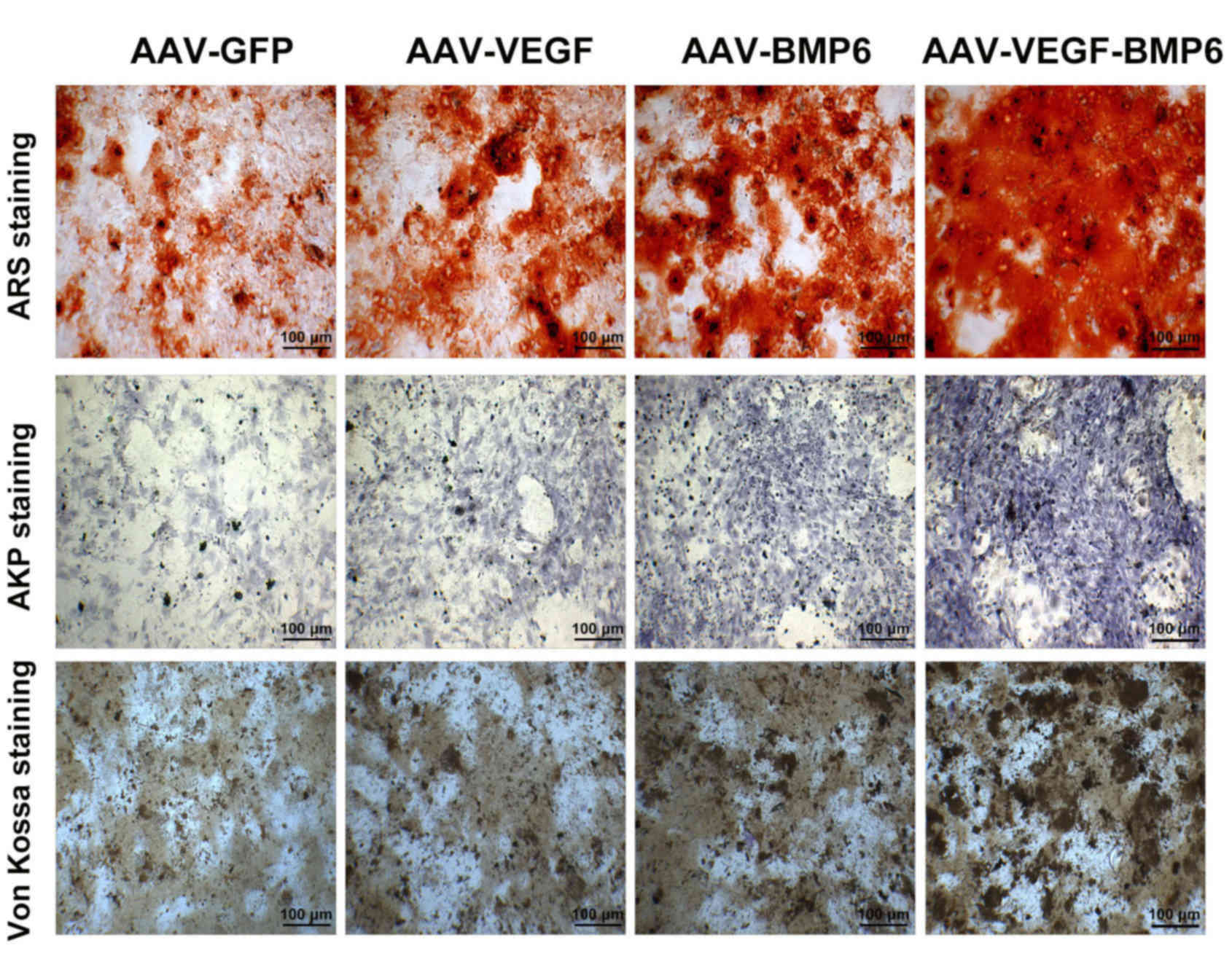 | Figure 4.The mineralization effects of VEGF
and BMP-6 on BMSCs. Representative images of ARS staining, AKP
staining, and Von Kossa staining in the four groups. The
mineralization of ARS staining is seen as mineralized nodules. The
mineralization of BMSCs stained using the AKP and Von Kossa methods
is represented by black dots. Scale bar, 100 µm. ARS, alizarin red
staining; AKP, alkaline phosphatase; AAV, adeno-associated virus;
GFP, green fluorescent protein; VEGF, vascular endothelial growth
factor; BMP, bone morphogenetic protein; BMSCs, bone mesenchymal
stem cells. |
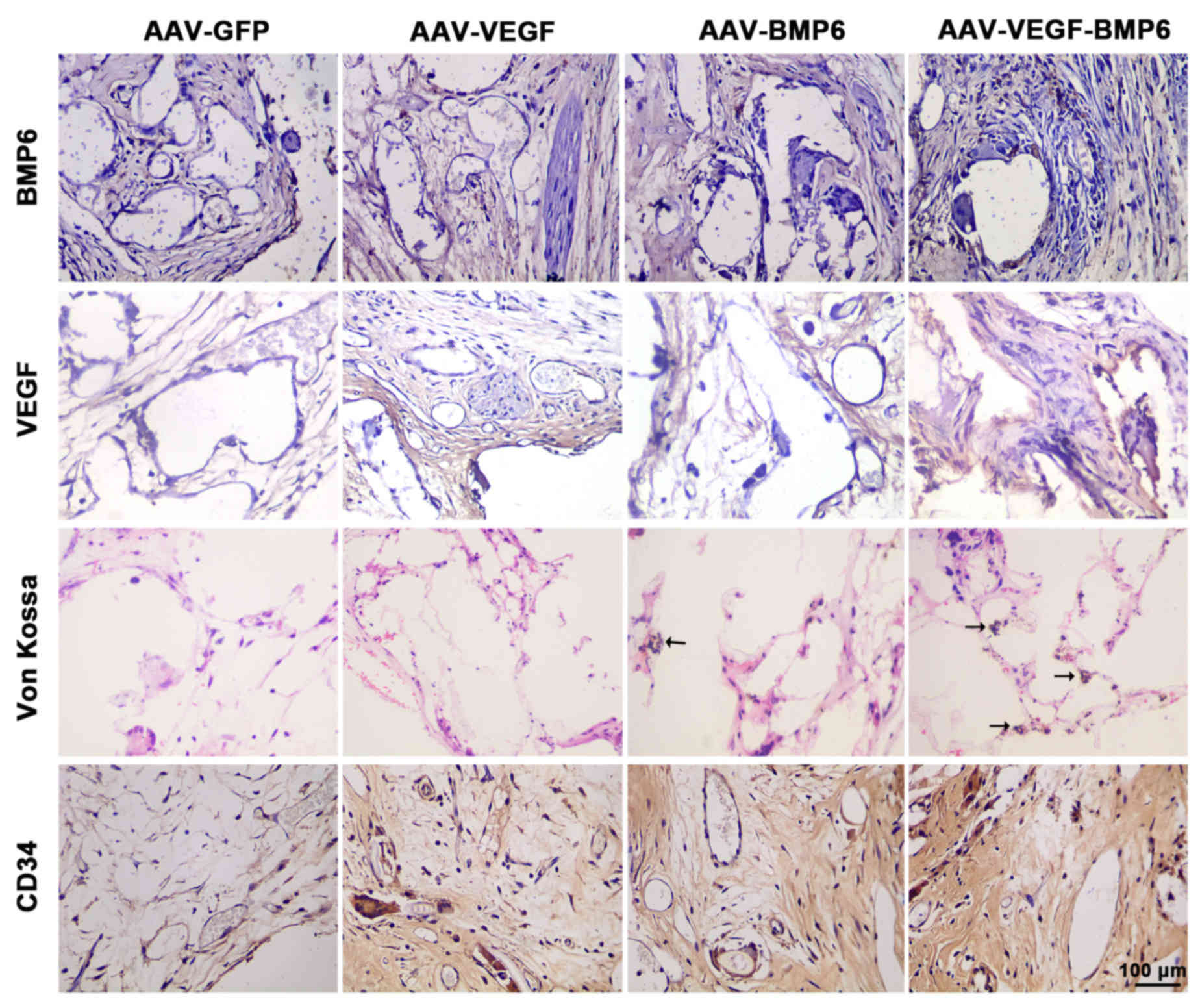
Histological assessment
Histological analysis with H&E and Von Kossa
staining was used to detect bone vessel and bone formation in
vivo. H&E staining analysis revealed that maximum bone
formation was achieved in PLAGA implants with BMSCs transfected
with VEGF and BMP-6. The greatest number of tubes and strongest
capillary integrity were observed in BMSCs transfected with VEGF
and BMP-6, with a significant increase in capillaries compared with
the AAV-GFP group (P<0.05; Fig.
5). The expression of VEGF and BMP-6 in PLAGA implants was
examined and it was observed that VEGF and BMP-6 expression was
markedly increased in the implants that carried BMSC cells
expressing VEGF and BMP-6 compared with the other groups (Fig. 6). Histological analysis with Von
Kossa staining was consistent with the H&E staining results.
Furthermore, cellularity and mineral deposition in the
AAV-VEGF-BMP-6 implants was markedly increased compared with all
other groups (Fig. 6). In order to
further assess the effect of VEGF delivery on angiogenesis,
immunostaining for CD34 was used to detect the number of
capillaries. The results were consistent in indicating that
scaffolds containing BMSCs transfected with VEGF and BMP-6
generated the largest number of blood vessels.
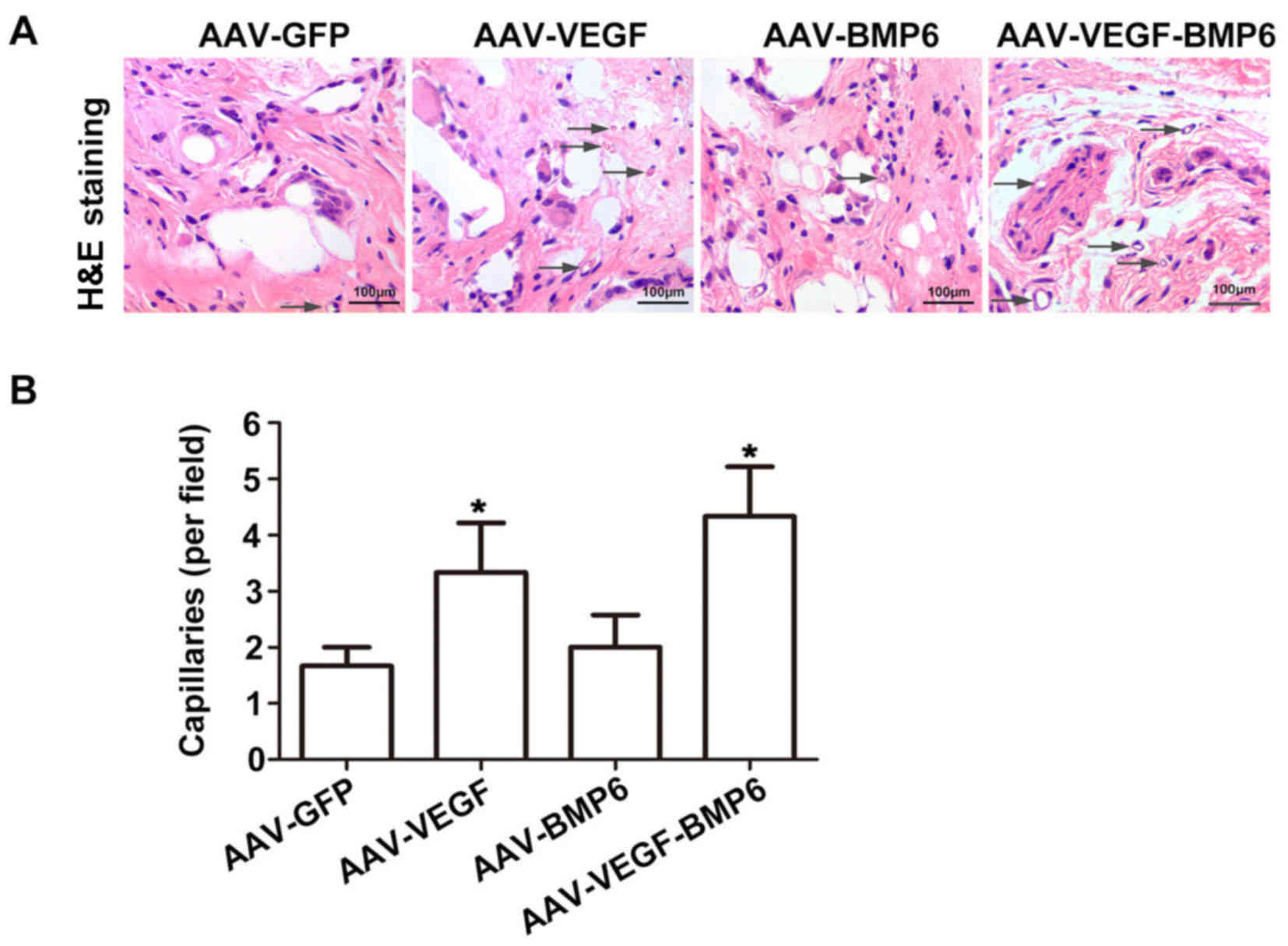 | Figure 5.H&E staining of BMSCs implanted
into PLAGA. (A) H&E staining of AAV-GFP, AAV-BMP-6, AAV-VEGF
and AAV-VEGF-BMP-6 groups was performed to detect in vivo
blood vessel formation. Blood vessels were identified by the
presence of a luminal structure, and the arrows point to the blood
vessels. Scale bar, 100 µm. (B) The number of capillaries in the
four groups was quantified. *P<0.05 vs. AAV-GFP group. PLAGA,
poly lactide-co-glycolide; H&E, hematoxylin and eosin; AAV,
adeno-associated virus; GFP, green fluorescent protein; VEGF,
vascular endothelial growth factor; BMP, bone morphogenetic
protein. |
Discussion
With the rapid development of gene therapy, it is
being increasingly recognized as a potential novel therapeutic
option for the treatment of ANFH (2). Neovascularization and bone formation
through the functions of VEGF and BMP have been extensively studied
(32). The AAV vector is an
attractive non-pathogenic human viral vector with low
immunogenicity (33). It is
efficient for gene transduction in vitro and for local
injection in vivo in the presence of a helper virus, for
example adenovirus or herpes virus (34,35). In
the present study, therefore, AAV was used to transfer and
correctly express the target gene.
Angiogenesis and osteogenesis are critical for bone
renovation and are associated with a number of cytokines, including
bFGF, TGF-β, BMPs, and VEGF (4,36). The
effects of VEGF and BMPs on the process of bone formation have
already been extensively studied (37,38).
VEGF is important in angiogenesis and is able to induce
angiopoiesis by promoting the division and growth of vascular
endothelial cells (7,39). In addition to its effects in
angiogenesis, several studies have reported that VEGF also serves
an important role in osteogenesis by interacting with BMPs
(40,41). It was therefore hypothesized that
VEGF may exhibit distinct effects when combined with different
BMPs. Several studies have shown that the VEGF and BMP-7 proteins
expressed by the rAAV-human (h)VEGF-IRES-hBMP-7 vector enhanced
angiogenesis and bone regeneration both in vitro and in
vivo (30,42). In addition, recent studies have
reported that BMP-6 and BMP-9 exhibited the most potent osteogenic
activity (43,44), and that BMP-6 encourages the
differentiation of human stromal cells to osteoblasts (45). Consequently, the combined effects of
VEGF and BMP-6 were assessed in the present study. Angiogenic and
osteogenic assays were used to identify the biological effects of
VEGF and BMP-6 in vitro and in vivo.
The present study demonstrated that the expression
of VEGF and BMP-6 was increased in the AAV-VEGF-BMP-6 group in
vitro compared with all other groups. VEGF secreted by BMSCs in
the AAV-VEGF-BMP-6 group enhanced HUVEC tube formation. Similarly,
the osteogenic ability of BMP-6 in the AAV-VEGF-BMP-6 group was
significantly higher compared with the other three groups, as
demonstrated by alizarin red, AKP and von Kossa staining. These
results indicate that the vector co-expressing VEGF and BMP-6
enhanced bone repair and regeneration in vitro. The results
in this study are in accord with those of a previous study
(46).
BMSCs are a type of multipotent mesenchymal stem
cell found in bone marrow (47).
They have a high capacity for self-renewal and the potential to
differentiate into several cell types, and so have long been
considered a source of cells for bone tissue engineering (47). Surface antigen detection using flow
cytometry is the gold standard for cell verification; in the
present study, CD29, CD90, CD44, CD45, CD11 and CD34 were selected
as target cell surface antigens based on previous literature
(48,49). The results revealed the
hyperexpression of CD29, CD90 and CD44, as well as hypoexpression
of CD45, CD11 and CD34, suggesting a high BMSC purity. PLAGA is a
biomedical polymer that has been safely used as an implant material
for several decades, including in bone tissue development (50). Prior to transplantation, PLAGA was
fabricated into a highly porous 3D scaffold upon which BMSCs could
be seeded with high efficiency (51). According to Roedersheimer et
al (36) and Cui et al
(46), VEGF and BMP-6 interact
directly at the molecular level or act via their signaling pathways
in order to promote osteogenesis.
In the present study, it was hypothesized that VEGF
and BMP-6 would act in an additive manner to promote the
osteoblastic differentiation of BMSCs in vitro and bone
formation in vivo. A composite bone graft substitute
containing BMSCs transfected with VEGF, BMP-6 or VEGF/BMP-6 loaded
on a PLAGA scaffold was used. The results revealed that there was a
significant increase in the bone volume of implants that carried
BMSCs expressing both VEGF and BMP-6 compared to those expressing
either of the growth factors alone at 2 and 3 weeks following
implantation. A greater number of blood vessels were also observed
in the AAV-VEGF-BMP-6 group compared with other BMSC groups
expressing either growth factor alone. Studies have revealed that
VEGF serves as a chemoattractant for mesenchymal stem cells to
increase the efficiency of bone formation in the presence of BMP-6
(52,53). As demonstrated in the present study,
VEGF and BMP-6 interact directly at the molecular level or via
their signaling pathways to indirectly enhance bone formation by
increasing vascular formation. The increased bone growth and
angiogenesis in subcutaneously implanted PLAGA scaffolds was in
accordance with the in vitro results.
In conclusion, co-expression of VEGF and BMP-6 in
BMSCs enhanced angiogenesis and bone regeneration in vitro
and in vivo. This demonstrates that BMSCs expressing both
VEGF and BMP-6 are responsible for increased numbers of blood
vessels and increased bone formation, which provides theoretical
support for their use in ANFH gene therapy.
Acknowledgements
The present study was supported by the Natural
Science Foundation of Guangdong Province, China (grant no.
2014A030307006), the National Natural Science Foundation of China
(grant no. 31430030) and the Medical Scientific Research Foundation
of Guangdong Province, China (grant no. A2017306).
References
|
1
|
Chamberlain JR, Schwarze U, Wang PR,
Hirata RK, Hankenson KD, Pace JM, Underwood RA, Song KM, Sussman M,
Byers PH and Russell DW: Gene targeting in stem cells from
individuals with osteogenesis imperfecta. Science. 303:1198–1201.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang C, Yang S, Du J, Li J, Xu W and Xiong
Y: Experimental study of vascular endothelial growth factor gene
therapy for avascular necrosis of the femoral head. J Huazhong Univ
Sci Technolog Med Sci. 23:297–299, 316. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wilson CG, Martin-Saavedra FM, Vilaboa N
and Franceschi RT: Advanced BMP gene therapies for temporal and
spatial control of bone regeneration. J Dent Res. 92:409–417. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kuh SU, Zhu Y, Li J, Tsai KJ, Fei Q,
Hutton WC and Yoon ST: Can TGF-beta1 and rhBMP-2 act in synergy to
transform bone marrow stem cells to discogenic-type cells? Acta
Neurochir (Wien). 150:1073–1079. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Evans CH and Huard J: Gene therapy
approaches to regenerating the musculoskeletal system. Nat Rev
Rheumatol. 11:234–242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Herberg S, Susin C, Pelaez M, Howie RN,
Moreno de Freitas R, Lee J, Cray JJ Jr, Johnson MH, Elsalanty ME,
Hamrick MW, et al: Low-dose bone morphogenetic protein-2/stromal
cell-derived factor-1β cotherapy induces bone regeneration in
critical-size rat calvarial defects. Tissue Eng Part A.
20:1444–1453. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
White AP, Vaccaro AR, Hall JA, Whang PG,
Friel BC and McKee MD: Clinical applications of BMP-7/OP-1 in
fractures, nonunions and spinal fusion. Int Orthop. 31:735–741.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
La WG, Jin M, Park S, Yoon HH, Jeong GJ,
Bhang SH, Park H, Char K and Kim BS: Delivery of bone morphogenetic
protein-2 and substance P using graphene oxide for bone
regeneration. Int J Nanomed. 9 Suppl 1:S107–S116. 2014.
|
|
9
|
Wang YJ, Chen S, Deng C, Li F, Wang Y, Hu
X, Shi F and Dong N: MicroRNA-204 Targets Runx2 to Attenuate
BMP-2-induced osteoblast differentiation of human aortic valve
interstitial cells. J Cardiovasc Pharmacol. 66:63–71. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mundy C, Gannon M and Popoff SN:
Connective tissue growth factor (CTGF/CCN2) negatively regulates
BMP-2 induced osteoblast differentiation and signaling. J Cell
Physiol. 229:672–681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hanada K, Solchaga LA, Caplan AI, Hering
TM, Goldberg VM, Yoo JU and Johnstone B: BMP-2 induction and
TGF-beta 1 modulation of rat periosteal cell chondrogenesis. J Cell
Biochem. 81:284–294. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang L, Park P, La Marca F, Than KD and
Lin CY: BMP-2 inhibits tumor-initiating ability in human renal
cancer stem cells and induces bone formation. J Cancer Res Clin
Oncol. 141:1013–1024. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dohin B, Dahan-Oliel N, Fassier F and
Hamdy R: Enhancement of difficult nonunion in children with
osteogenic protein-1 (OP-1): Early experience. Clin Orthop Relat
Res. 467:3230–3238. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takahashi T, Hatakeyama S and Machida T:
Ductal adenocarcinoma of the pancreas with psammomatous
calcification: Report of a case with immunohistochemical study for
bone morphogenetic protein. Pathol Int. 61:603–607. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rahman MS, Akhtar N, Jamil HM, Banik RS
and Asaduzzaman SM: TGF-β/BMP signaling and other molecular events:
Regulation of osteoblastogenesis and bone formation. Bone Res.
3:150052015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Scharpfenecker M, van Dinther M, Liu Z,
van Bezooijen RL, Zhao Q, Pukac L, Löwik CW and Ten Dijke P: BMP-9
signals via ALK1 and inhibits bFGF-induced endothelial cell
proliferation and VEGF-stimulated angiogenesis. J Cell Sci.
120:964–972. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Simic P, Culej JB, Orlic I, Grgurevic L,
Draca N, Spaventi R and Vukicevic S: Systemically administered bone
morphogenetic protein-6 restores bone in aged ovariectomized rats
by increasing bone formation and suppressing bone resorption. J
Biol Chem. 281:25509–25521. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mizrahi O, Sheyn D, Tawackoli W, Kallai I,
Oh A, Su S, Da X, Zarrini P, Cook-Wiens G, Gazit D and Gazit Z:
BMP-6 is more efficient in bone formation than BMP-2 when
overexpressed in mesenchymal stem cells. Gene Ther. 20:370–377.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Street J, Bao M, deGuzman L, Bunting S,
Peale FV Jr, Ferrara N, Steinmetz H, Hoeffel J, Cleland JL,
Daugherty A, et al: Vascular endothelial growth factor stimulates
bone repair by promoting angiogenesis and bone turnover. Proc Natl
Acad Sci USA. 99:pp. 9656–9661. 2002; View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Moens S, Goveia J, Stapor PC, Cantelmo AR
and Carmeliet P: The multifaceted activity of VEGF in
angiogenesis-Implications for therapy responses. Cytokine Growth
Factor Rev. 25:473–482. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gerber HP, Vu TH, Ryan AM, Kowalski J,
Werb Z and Ferrara N: VEGF couples hypertrophic cartilage
remodeling, ossification and angiogenesis during endochondral bone
formation. Nat Med. 5:623–628. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ferrara N: VEGF-A: A critical regulator of
blood vessel growth. Eur Cytokine Netw. 20:158–163. 2009.PubMed/NCBI
|
|
23
|
Joensuu K, Uusitalo L, Alm JJ, Aro HT,
Hentunen TA and Heino TJ: Enhanced osteoblastic differentiation and
bone formation in co-culture of human bone marrow mesenchymal
stromal cells and peripheral blood mononuclear cells with exogenous
VEGF. Orthop Traumatol Surg Res. 101:381–386. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ramazanoglu M, Lutz R, Rusche P, Trabzon
L, Kose GT, Prechtl C and Schlegel KA: Bone response to biomimetic
implants delivering BMP-2 and VEGF: An immunohistochemical study. J
Craniomaxillofac Surg. 41:826–835. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang W, Zhu C, Wu Y, Ye D, Wang S, Zou D,
Zhang X, Kaplan DL and Jiang X: VEGF and BMP-2 promote bone
regeneration by facilitating bone marrow stem cell homing and
differentiation. Eur Cells Mater. 27:1–12. 2014. View Article : Google Scholar
|
|
26
|
Kempen DH, Lu L, Heijink A, Hefferan TE,
Creemers LB, Maran A, Yaszemski MJ and Dhert WJ: Effect of local
sequential VEGF and BMP-2 delivery on ectopic and orthotopic bone
regeneration. Biomaterials. 30:2816–2825. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Peng H, Usas A, Olshanski A, Ho AM,
Gearhart B, Cooper GM and Huard J: VEGF improves, whereas sFlt1
inhibits, BMP2-induced bone formation and bone healing through
modulation of angiogenesis. J Bone Miner Res. 20:2017–2027. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pittenger MF, Mackay AM, Beck SC, Jaiswal
RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S and
Marshak DR: Multilineage potential of adult human mesenchymal stem
cells. Science. 284:143–147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Büning H, Perabo L, Coutelle O,
Quadt-Humme S and Hallek M: Recent developments in adeno-associated
virus vector technology. J Gene Med. 10:717–733. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shi ZB and Wang KZ: Effects of recombinant
adeno-associated viral vectors on angiopoiesis and osteogenesis in
cultured rabbit bone marrow stem cells via co-expressing hVEGF and
hBMP genes: A preliminary study in vitro. Tissue Cell. 42:314–321.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2012.
View Article : Google Scholar
|
|
32
|
Xin CW, Zheng LW, Lei LC, Ma L, Ehrbar M,
Weber FE and Zwahlen RA: Effect of different rhBMP-2 and TG-VEGF
ratios on the formation of heterotopic bone and neovessels. Biomed
Res Int. 2014:5715102014.PubMed/NCBI
|
|
33
|
Ali RR, Reichel MB, Thrasher AJ, Levinsky
RJ, Kinnon C, Kanuga N, Hunt DM and Bhattacharya SS: Gene transfer
into the mouse retina mediated by an adeno-associated viral vector.
Hum Mol Genet. 5:591–594. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Merten OW, Gény-Fiamma C and Douar AM:
Current issues in adeno-associated viral vector production. Gene
Ther. 12 Suppl 1:S51–S61. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Grieger JC, Choi VW and Samulski RJ:
Production and characterization of adeno-associated viral vectors.
Nat Protoc. 1:1412–1428. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Roedersheimer M, West J, Huffer W, Harral
J and Benedict J: A bone-derived mixture of TGF beta-superfamily
members forms a more mature vascular network than bFGF or TGF-beta
2 in vivo. Angiogenesis. 8:327–338. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Samee M, Kasugai S, Kondo H, Ohya K,
Shimokawa H and Kuroda S: Bone morphogenetic protein-2 (BMP-2) and
vascular endothelial growth factor (VEGF) transfection to human
periosteal cells enhances osteoblast differentiation and bone
formation. J Pharmacol Sci. 108:18–31. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sukul M, Nguyen TB, Min YK, Lee SY and Lee
BT: Effect of local sustainable release of BMP2-VEGF from
Nano-cellulose loaded in sponge biphasic calcium phosphate on bone
regeneration. Tissue Eng Part A. 21:1822–1836. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cao QS, Zhang T and Zhang J: Correlation
analysis of STAT3 and VEGF expression and eosinophil infiltration
in nasal polyps. Eur Arch Otorhinolaryngol. 272:1955–1960. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kanczler JM, Ginty PJ, White L, Clarke NM,
Howdle SM, Shakesheff KM and Oreffo RO: The effect of the delivery
of vascular endothelial growth factor and bone morphogenic
protein-2 to osteoprogenitor cell populations on bone formation.
Biomaterials. 31:1242–1250. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jiang J, Wang SH, Chai F, Ai CC and Chen
SY: The study on vascularisation and osteogenesis of BMP/VEGF
co-modified tissue engineering bone in vivo. RSC Adv.
6:41800–41808. 2016. View Article : Google Scholar
|
|
42
|
Zhang C, Wang KZ, Qiang H, Tang YL, Li Q,
Li M and Dang XQ: Angiopoiesis and bone regeneration via
co-expression of the hVEGF and hBMP genes from an adeno-associated
viral vector in vitro and in vivo. Acta Pharmacol Sin. 31:821–830.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yung LM, Sánchez-Duffhues G, Ten Dijke P
and Yu PB: Bone morphogenetic protein 6 and oxidized Low-density
lipoprotein synergistically recruit osteogenic differentiation in
endothelial cells. Cardiovasc Res. 108:278–287. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wei Z, Salmon RM, Upton PD, Morrell NW and
Li W: Regulation of bone morphogenetic protein 9 (BMP9) by
redox-dependent proteolysis. J Biol Chem. 289:31150–31159. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Friedman MS, Long MW and Hankenson KD:
Osteogenic differentiation of human mesenchymal stem cells is
regulated by bone morphogenetic protein-6. J Cell Biochem.
98:538–554. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cui F, Wang X, Liu X, Dighe AS, Balian G
and Cui Q: VEGF and BMP-6 enhance bone formation mediated by cloned
mouse osteoprogenitor cells. Growth Factors. 28:306–317. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Derubeis AR and Cancedda R: Bone marrow
stromal cells (BMSCs) in bone engineering: Limitations and recent
advances. Ann Biomed Eng. 32:160–165. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xiong J, Menicanin D, Zilm PS, Marino V,
Bartold PM and Gronthos S: Investigation of the cell surface
proteome of human periodontal ligament stem cells. Stem Cells Int.
2016:19471572016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hung SC, Chen NJ, Hsieh SL, Li H, Ma HL
and Lo WH: Isolation and characterization of size-sieved stem cells
from human bone marrow. Stem Cells. 20:249–258. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Bongio M, van den Beucken JJJP,
Leeuwenburgh SCG and Jansen JA: Development of bone substitute
materials: From ‘biocompatible’ to ‘instructive’. J Mater Chem.
20:8747–8759. 2010. View Article : Google Scholar
|
|
51
|
Nugroho RWN, Odelius K, Hoglund A and
Albertsson AC: Highlighting the importance of surface grafting in
combination with a Layer-by-Layer approach for fabricating advanced
3D Poly(L-lactide) microsphere scaffolds. Chem Mater. 28:3298–3307.
2016. View Article : Google Scholar
|
|
52
|
Madhu V, Li CJ, Dighe AS, Balian G and Cui
Q: BMP-non-responsive
Sca1+CD73+CD44+ mouse bone marrow
derived osteoprogenitor cells respond to combination of VEGF and
BMP-6 to display enhanced osteoblastic differentiation and ectopic
bone formation. PLoS One. 9:e1030602014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Fiedler J, Leucht F, Waltenberger J, Dehio
C and Brenner RE: VEGF-A and PlGF-1 stimulate chemotactic migration
of human mesenchymal progenitor cells. Biochem Biophys Res Commun.
334:561–568. 2005. View Article : Google Scholar : PubMed/NCBI
|