Introduction
Atherosclerosis (AS), a chronic inflammatory
disorder, is the most prevalent cardiovascular disease and the
world's leading cause of fatality and morbidity (1–3).
Evidence has indicated that modified lipids, particularly oxidized
low-density lipoprotein (ox-LDL), are predominantly engulfed by
macrophages, resulting in increased pro-inflammatory cytokine
expression and further macrophage recruitment, thus contributing to
increases in the size and complexity of atherosclerotic plaques
(4–7). Therefore, it is of crucial importance
to investigate the interaction between macrophages and modified
lipids as well as the regulatory mechanisms of inflammation in
macrophages.
Autophagy, a highly controlled intracellular process
whereby long-lived proteins and damaged organelles are degraded and
recycled, has an essential role in cell survival and in responses
to environmental or internal pressure (8–12).
Recent studies have demonstrated that autophagy is involved in
complicated interactions with the innate and adaptive immunity, and
thus is associated with the pathogenesis of inflammatory diseases
(13–17). Macrophages lacking autophagy-related
protein 16-1 (Atg16L1) or autophagy-related (Atg)7, key components
of the autophagic process, resulted in upregulated secretion of
interleukin (IL)-1β and IL-18 in response to inflammatory
stimulation (13). Using mice with
knocked out autophagy protein Atg5 in macrophages and neutrophils,
researchers identified that Atg5 serves an important role in
resistance to the intracellular pathogens Listeria
monocytogenes and Toxoplasma gondii (18). Atg5 was required for interferon
(IFN)-γ/lipopolysaccharide (LPS)-induced harm to the T.
gondii parasitophorous vacuole membrane, and thus the
elimination of intracellular pathogens (18). With respect to the pathogenesis of
AS, studies have reported that ox-LDL-induced autophagy impairment
in HUVECs contributes to the development of this disease (19). Furthermore, autophagy dysfunction in
macrophages may accelerate atherosclerotic progression in
vivo (20). Thus, modulation of
autophagy may lead to therapeutic interventions for AS.
Hydrogen, possessing anti-oxidative properties by
selectively neutralizing cytotoxic reactive oxygen species, exerts
organ-protective effects by regulating oxidative stress,
inflammation and apoptosis (21).
Recently, accumulating evidence has demonstrated the protective
role of hydrogen on diverse models of disease, including diseases
of the central nervous system, cardiovascular system, lungs and
renal system (21–26). Hydrogen treatment attenuated
LPS-induced upregulation of pro-inflammatory cytokines, including
tumor necrosis factor (TNF)-α, IL-1β and high mobility group box 1
in macrophages (27). Chronic
hydrogen-rich saline treatment depressed pro-inflammatory cytokines
expression, including IL-6 and IL-1β (28). In addition, hydrogen-rich saline
mediates neuroprotection and attenuates acute kidney injury through
the regulation of autophagy (29,30).
Oral treatment with hydrogen-rich water for 6 months alleviated the
progression of AS in apolipoprotein E-knockout mice (31). However, the therapeutic mechanism
underlying its protective effects has not yet been completely
elucidated. Considering the inhibited effect of autophagy on
inflammation and the promoted effect of hydrogen on autophagy
(32,33), the present study intended to explore
the effect of hydrogen on ox-LDL-induced inflammation and to
investigate the potential role of autophagy in hydrogen-mediated
protection.
Materials and methods
Reagents
Bovine serum albumin (BSA; cat. no. A4161),
rapamycin (cat. no. R117), bafilomycin A1 (BafA1; cat. no. B1793)
and protease inhibitor cocktail (cat. no. P8340) were purchased
from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Ox-LDL was
obtained from Yiyuan Biotech Co., Ltd (Guangzhou, Guangdong,
China). RPMI-1640 medium and fetal bovine serum were obtained from
Gibco (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Radioimmunoprecipitation assay (RIPA) lysis buffer (cat. no.
P0013B) was purchased from Beyotime Institute of Biotechnology
(Haimen, China).
Hydrogen preparation
As previously described (21,27),
purified hydrogen was dissolved into RPMI-1640 medium at 0.4 MPa
pressure for 2 h to reach saturation. O2 was dissolved
into a second medium by bubbling O2 gas at the saturated
level (42.5 mg/l), and CO2 into a third medium by
bubbling CO2 gas. A total of 0.6 mM hydrogen medium was
prepared by combining the three media (H2
medium:O2 medium:CO2 medium) in the
proportion of 90%:5%:5% (v/v/v). The control medium was prepared by
combining two media (95% O2 medium:5% CO2
medium). H2, O2 and CO2
concentrations were confirmed with gas chromatography.
Cell culture
The mouse macrophage-like cell line Raw264.7,
obtained from the Type Culture Collection of the Chinese Academy of
Sciences (Shanghai, China), was cultured in RPMI-1640 medium
supplemented with 10% fetal bovine serum and maintained at 37°C in
an atmosphere containing 5% CO2.
Cell viability assay
Cells were seeded in 96-well plates at a density of
1×106 cells/well and were cultured with or without
various concentrations (0.00, 0.15, 0.30 and 0.60 mM) of hydrogen
for 24 h at 37°C. Subsequently, cell viability was determined using
water-soluble tetrazolium salt-8 staining with a CCK8 (Cell
Counting kit-8) assay, (cat. no. CK04; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) according to the
manufacturer's instructions. Optical density was determined at 450
nm with a plate reader (Multiskan FC; Thermo Fisher Scientific,
Inc.).
Cell transfection experiment
Control small interfering (si)RNA (non-targeting;
siControl) and siRNA specific for mouse sirtuin 1 (SIRT1;
ON-TARGETplus SMARTpool; cat. no. L-303540-00) were obtained from
GE Healthcare Dharmacon, Inc. (Lafayette, CO, USA). The sequences
of the SIRT1 siRNAs utilized are presented in Table I. A total of 20 nM siRNA was
transfected into osteoblasts using Lipofectamine 2000 (cat. no.
11668027; Invitrogen; Thermo Fisher Scientific, Inc.), as reported
previously (34). Cells were
incubated for 24 h before being used for subsequent
experiments.
 | Table I.Sequences of SIRT1 siRNAs. |
Table I.
Sequences of SIRT1 siRNAs.
| SIRT1 siRNAs | Sequence |
|---|
| 1 SIRT1 siRNA
sense |
5′-GAUGAAGUUGACCUCCUCATT-3′ |
| 1 SIRT1 siRNA
antisense |
5′-UGAGGAGGUCAACUUCAUCTT-3′ |
| 2 SIRT1 siRNA
sense |
5′-TCAGTGTCATGGTTCCTTTGC-3′ |
| 2 SIRT1 siRNA
antisense |
5′-AATCTGCTCCTTTGCCACTCT-3′ |
| 3 SIRT1 siRNA
sense |
5′-GCAACAGCAUCUUGCCUGACUUGUA-3′ |
| 3 SIRT1 siRNA
antisense |
5′-UCAUAGAGCCAUGAAGUAUGACAAA-3′ |
| 4 SIRT1 siRNA
sense |
5′-TTTCAGAACCACCAAAGCG-3′ |
| 4 SIRT1 siRNA
antisense |
5′-TCCCACAGGAAACAGAAACC-3′ |
Western blot analysis
Cells were lysed in RIPA lysis buffer with a protein
inhibitor cocktail for 30 min on ice, and the lysates were
centrifuged at 12,000 × g for 10 min at 4°C. The supernatants were
collected, and 12 or 15% SDS-PAGE was used to separate the proteins
before transfer to polyvinylidene fluoride membranes (EMD
Millipore; Billerica, MA, USA). The membranes were blocked with 5%
non-fat milk for 1 h at room temperature, followed by incubation
with the following primary antibodies: Anti-microtubule-associated
protein 1A/1B-light chain 3 (LC3; 1:1,000; cat. no. L7543;
Sigma-Aldrich; Merck KgaA), anti-p62 (1:1,000; cat. no. 39749),
anti-SIRT1 (1:1,000; cat. no. 8469; both Cell Signaling Technology,
Inc., Danvers, MA, USA) and anti-GAPDH (1:1,000; cat no. A01020;
Abbkine Scientific Co., Ltd., Wuhan, China) at 4°C overnight.
Membranes were washed with Tris-buffered saline with Tween-20 (20
mM Tris-HCl buffer; pH=7.4; containing 150 mM NaCl and 0.05%
Tween-20) three times and then incubated with horseradish
peroxidase (HRP)-conjugated anti-rabbit immunoglobulin G (1:2,000;
cat. no. 7074; Cell Signaling Technology, Inc.) and HRP-conjugated
anti-mouse IgG antibodies (1:1,000; cat. no. sc-2005; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) at room temperature for 2 h.
After probing with specific primary antibodies and an
HRP-conjugated secondary antibody, the protein bands were detected
using a super ECL plus detection reagent (Thermo Fisher Scientific,
Inc.), and the band density was analyzed using ImageJ 1.41
(National Institutes of Health; Bethesda, MD, USA).
ELISA
The levels of inflammatory cytokines [IL-6 (cat. no.
CME0006), TNF-α (cat. no. CME0004) and IL-1β (cat. no. CME0015)]
were detected using ELISA kits (4A Biotech Co. Ltd., Beijing
China). All procedures were performed in accordance with the
manufacturer's instructions.
Statistical analysis
Results were expressed as the mean ± standard error
of the mean. The differences between groups were analyzed using the
Brown-Forsythe test and, if appropriate, by one-way analysis of
variance followed by Dunnett's post hoc test or the Bonferroni
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Ox-LDL impairs autophagic flux in
macrophages
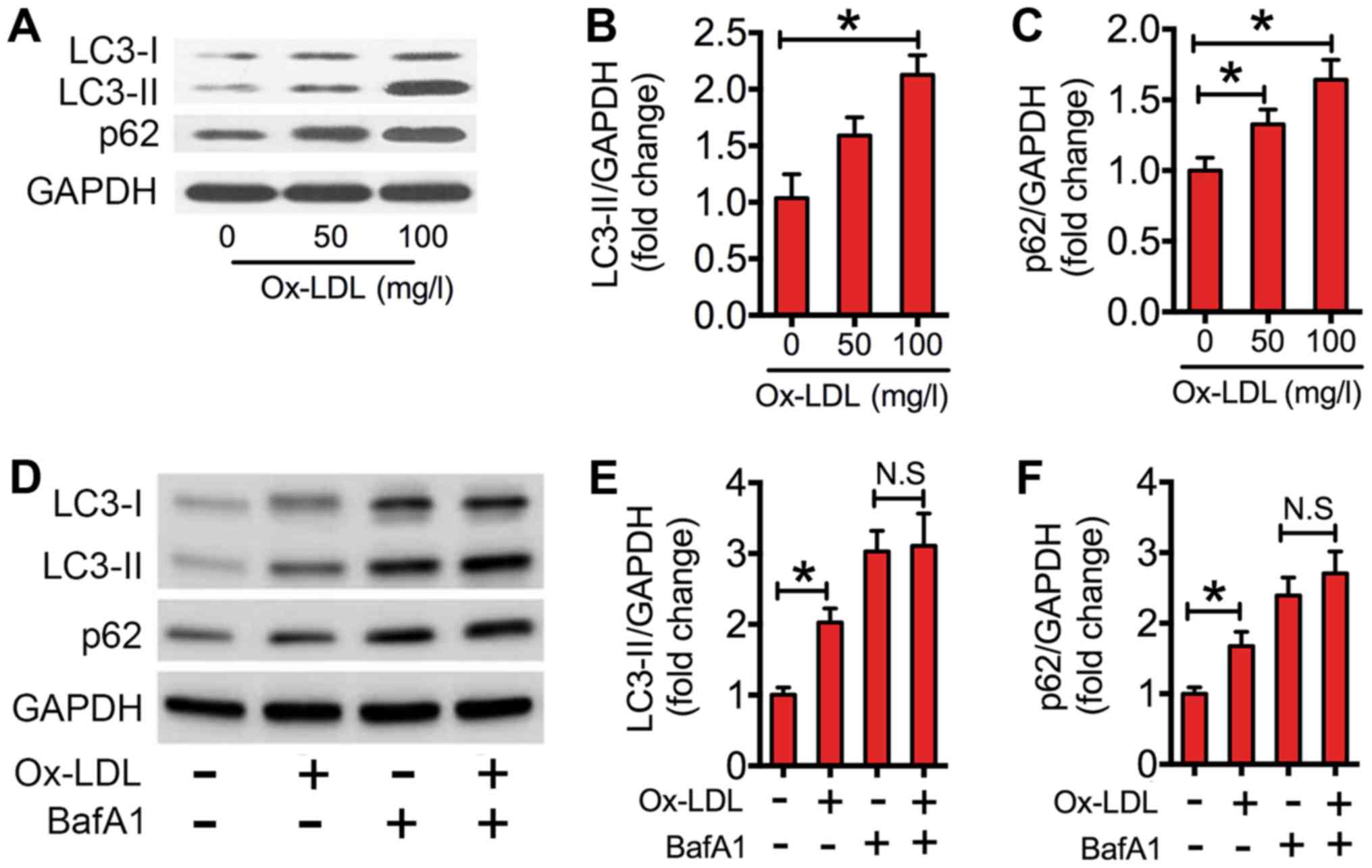
In the present study, western blotting of
LC3-II/GAPDH ratios in ox-LDL-treated RAW264.7 cells was performed.
As indicated in Fig. 1A and B, the
expression level of LC3-II was significantly increased in
macrophages treated with 100 mg/l ox-LDL compared with that of the
control. The increased level of LC3-II could be due to increased
initiation of autophagy or decreased clearance of autophagosomes,
as reflected by accumulation of the autophagic substrate p62.
Therefore, the protein expression levels of p62 in ox-LDL
co-cultured macrophages were measured. When compared with the
control, ox-LDL significantly increased p62 expression levels in
macrophages (Fig. 1A and C). To
further interpret the decreased clearance of autophagosomes, BafA1,
which inhibits the late degradation stage of autophagy. Results
indicated that ox-LDL significantly elevated the expression levels
of LC3-II and p62 relative to the control (Fig. 1D-F). However, the ox-LDL-induced
increase in LC3-II and p62 was nullified in the presence of BafA1
(Fig. 1D-F). These results
demonstrated that ox-LDL impaired autophagic flux in
macrophages.
Ox-LDL-induced inflammation in
macrophages
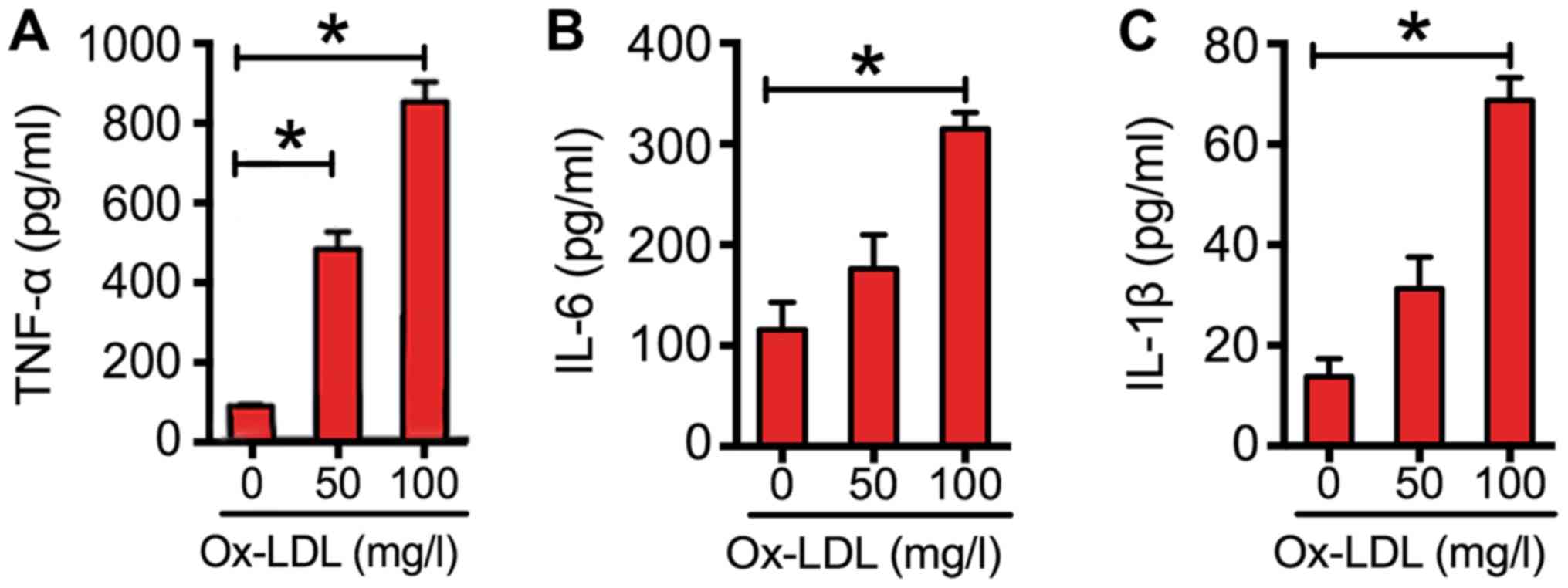
To explore the effects of ox-LDL on inflammatory
cytokine expression in macrophages, RAW264.7 cells were exposed to
various concentrations of ox-LDL. The results indicated that ox-LDL
dose-dependently increased the expression of cytokines (Fig. 2). At a concentration of 100 mg/l, the
levels of TNF-α were increased by almost 7-fold; those of IL-6 were
increased by 2-fold; and those of IL-1β were increased by 4-fold
(Fig. 2).
Hydrogen enhances autophagic flux in
macrophages
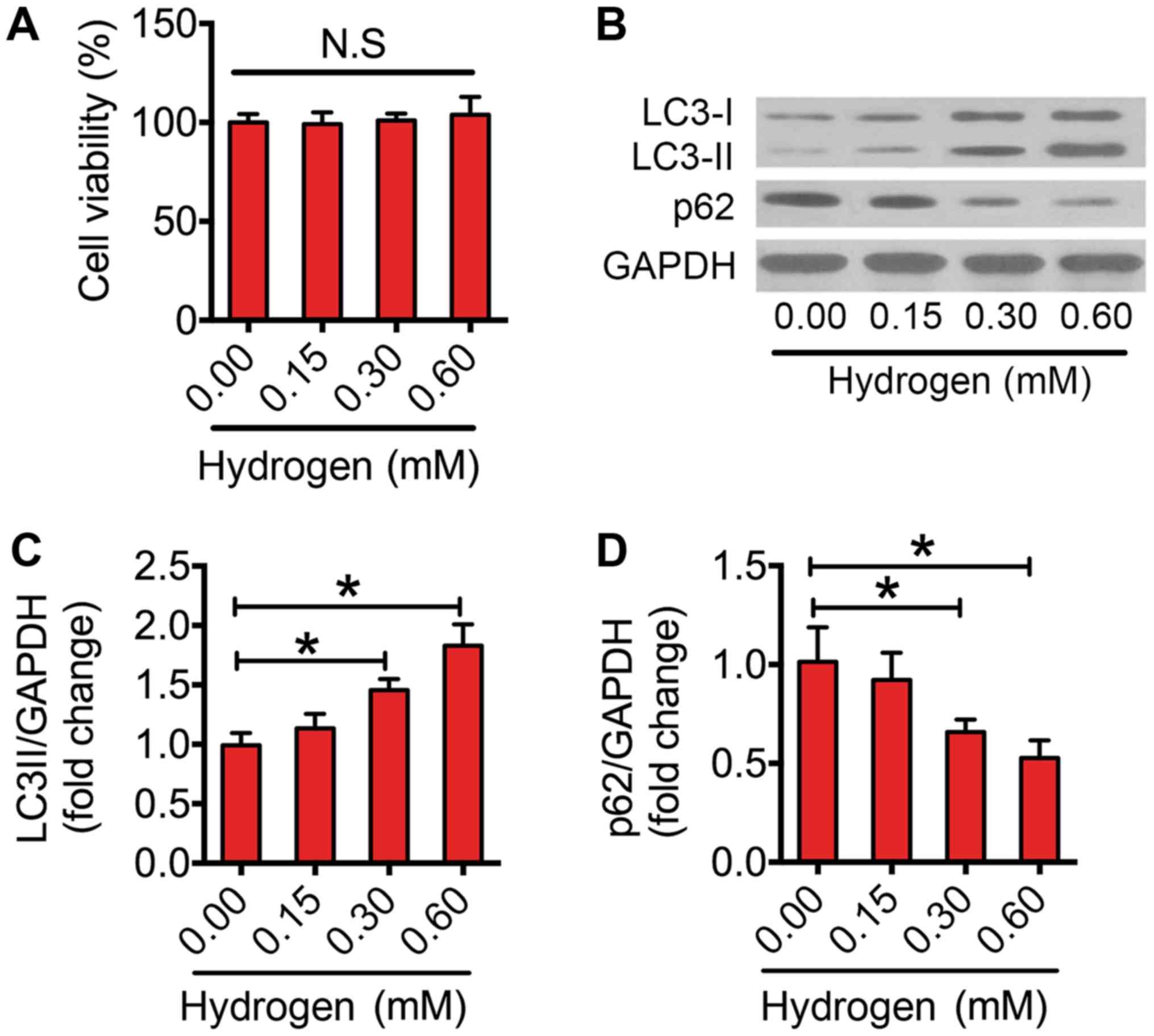
Previous studies have suggested that hydrogen
triggers autophagy in several diseases (29,30). In
the present study, it was investigated whether hydrogen could
activate autophagy in macrophages. Firstly, the results indicated
that hydrogen did not impair the cell viability at the selected
dose (Fig. 3A). Following this, the
effect of hydrogen on autophagic flux was assessed. Notably,
hydrogen increased the protein expression levels of LC3-II in a
dose-dependent manner (Fig. 3B and
C). Furthermore, 0.30 and 0.60 mM hydrogen significantly
reduced the expression level of p62 in macrophages (Fig. 3B and D). These results indicated that
treatment with hydrogen enhanced autophagic flux in
macrophages.
Enhanced autophagic flux mediates the
anti-inflammatory effect of hydrogen in ox-LDL-treated
macrophages
The above results demonstrated that ox-LDL increased
inflammatory cytokine expression levels and impaired autophagic
flux in macrophages. In Fig. 4A-C,
the data suggest that hydrogen downregulated cytokines in a
dose-dependent manner in macrophages. To further investigate
whether the effect of hydrogen on ox-LDL-induced cytokines was
involved in autophagic flux, RAW264.7 cells were pretreated with
BafA1 (an inhibitor of autophagy) or rapamycin (an activator of
autophagy) before being cultured with ox-LDL or ox-LDL and
hydrogen. As indicated in Fig. 4D-F,
rapamycin significantly decreased the expression of cytokines
induced by ox-LDL. Similarly, the expression of cytokines induced
by ox-LDL was lower in hydrogen cotreated cells compared with the
ox-LDL alone group. In addition, inhibition of hydrogen-induced
autophagy by BafA1 partly reversed its effects on cytokine
expression. Collectively, these results indicated that hydrogen
ameliorated ox-LDL-induced inflammation, and the anti-inflammatory
effect was mediated by enhanced autophagic flux.
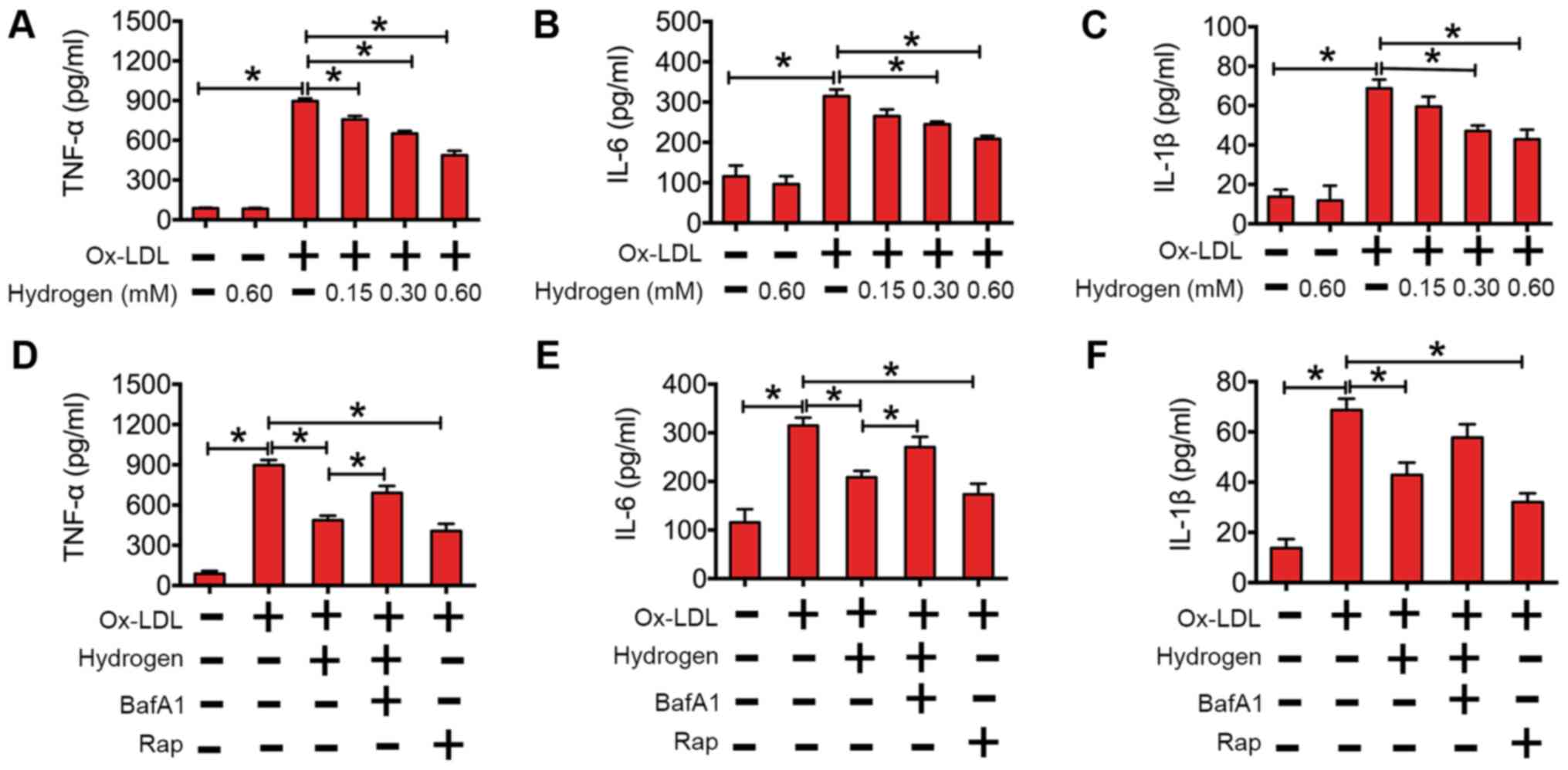 | Figure 4.Hydrogen inhibited ox-LDL-induced
inflammatory cytokine expression by upregulation of autophagic flux
in RAW264.7 cells. The expression of (A) TNF-α, (B) IL-6 and (C)
IL-1β after RAW264.7 cells were incubated with/without hydrogen
(0.00, 0.15, 0.30 and 0.60 mM), prior to being stimulated
with/without ox-LDL (100 mg/l) for 24 h. The expression of (D)
TNF-α, (E) IL-6 and (F) IL-1β after RAW264.7 cells were incubated
with/without hydrogen (0.60 mM), bafilomycin A1 (50 nM) and/or
rapamycin (200 nM), prior to being stimulated with/without ox-LDL
(100 mg/l) for 24 h. The cytokine concentrations were quantified by
ELISA. Data are presented as the mean ± standard error of the mean
from three independent experiments. *P<0.05 as indicated. IL,
interleukin; BafA1, bafilomycin A1; Rap, rapamycin; TNF-α, tumor
necrosis factor-α; Ox-LDL, oxidized low-density lipoprotein. |
Upregulation of SIRT1 is essential for
the rescuing effect of hydrogen on autophagy flux in ox-LDL-treated
macrophages
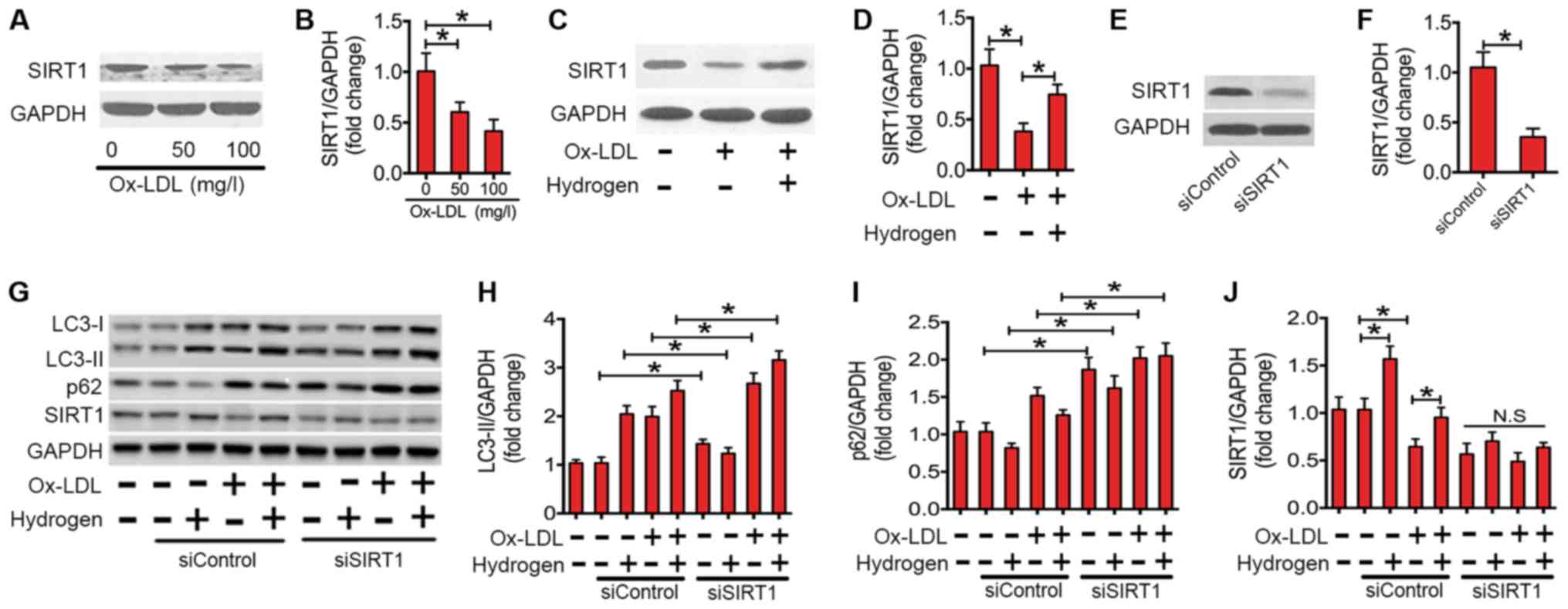
SIRT1, a member of the NAD+-dependent
deacetylases, serves an important role in regulating autophagic
flux (35,36). Recently, studies have demonstrated
that SIRT1 is involved in the inflammatory responses of various
conditions (37). In the present
study, the expression of the SIRT1 protein in ox-LDL-stimulated
macrophages was investigated (Fig.
5). Ox-LDL treatment significantly decreased the expression of
SIRT1 protein in a dose-dependent manner (Fig. 5A and B). However, co-treatment with
hydrogen significantly rescued the expression level of SIRT1
protein in ox-LDL-stimulated macrophages (Fig. 5C and D). To investigate whether the
ability of hydrogen to ameliorate impaired autophagic flux depends
on SIRT1, siRNA was used to knock down SIRT1 (Fig. 5E-G and J). As indicated in Fig. 5G and H, following the transfection
with SIRT1 siRNA, the expression level of LC3-II was significantly
increased. Furthermore, the downregulating effect of hydrogen on
p62 expression was abolished (Fig. 5G
and I). These results indicated that SIRT1 is essential for the
ameliorating the effect of hydrogen on ox-LDL-induced impaired
autophagic flux in macrophages.
Discussion
AS predisposes patients to various cardiovascular
diseases, including hypertension, coronary heart disease and stroke
(38–41). Ox-LDL-induced inflammation serves a
crucial role in the pathogenesis of atherosclerosis. Hydrogen, as a
novel antioxidant, can exert potent cellular protective effects in
different disease models (22).
Notably, oral administration of hydrogen-rich water for 6 months
prevented the progress of atherosclerosis in an animal model
(31). Furthermore, hydrogen
represents a novel therapeutic choice for cardiovascular disease
(22). However, the molecular
mechanisms require elucidation. In the present study, the present
data suggested that hydrogen attenuated ox-LDL-induced inflammatory
cytokine expression through the stimulation of autophagy via
SIRT1.
Hydrogen, as the lightest and most abundant chemical
element, may act as a scavenger to selectively ameliorate reactive
oxygen species (ROS) and exert potent cellular protective effects
in a variety of clinical and experimental models of diseases
(21,30). In rats, hydrogen gas reduced infarct
sizes without affecting hemodynamic parameters during
ischemia-reperfusion (I/R) injury of the left anterior descending
coronary artery (42).
Intraperitoneal injection of hydrogen-rich saline exerted
cardioprotection against I/R injury by reducing plasma and
myocardial malondialdehyde concentrations, cardiac cell apoptosis
and caspase-3 activity (43).
Furthermore, continued consumption of hydrogen water decreased the
oxidative stress level in the aorta and reduced the number of
atherosclerotic lesions in a previous study (31). In the present study, the effects of
hydrogen on ox-LDL-treated macrophages were examined in
vitro, and the results indicated that hydrogen markedly
decreased the secretion of pro-inflammatory cytokines compared with
the control. Considering the pivotal role of ox-LDL-induced
inflammation in the pathogenesis of AS, hydrogen may serve as an
anti-inflammatory agent and have therapeutic potential in the
treatment of atherosclerosis.
Autophagy is a key component of the cellular stress
adaptation responses that maintain cellular homeostasis. It is
important in protection against different types of disease,
including neurodegenerative disease, aging, cancer and inflammatory
illness (44–46). Perturbations in autophagy contribute
to chronic inflammatory diseases and autoimmune diseases, including
Crohn's disease, systemic lupus erythematosus, obesity and diabetes
(47). In mice, loss of autophagy
via knockout of Atg5 in macrophages results in enhanced systemic
and hepatic inflammation, which is due to the disturbance between
pro-inflammatory M1 and anti-inflammatory M2 polarization (48). Atg16L1-deficient activated caspase-1
by Toll/IL-1 receptor domain-containing adaptor inducing IFN-β in
macrophages and resulted in increased secretion of the inflammatory
cytokines IL-1β and IL-18 (49). The
present data revealed that ox-LDL treatment resulted in impaired
autophagic flux. Furthermore, enhanced autophagy by rapamycin (a
classical autophagy activator) or hydrogen decreased the effects on
ox-LDL-induced pro-inflammatory cytokine expression. However,
downregulation of autophagy by bafilomycin A1 alleviated the
inhibitory effect of hydrogen on pro-inflammatory cytokine
secretion.
SIRT1, belonging to the silent information regulator
2 family, has been reported to be involved in age-associated
diseases, including neurodegenerative diseases, metabolic diseases,
cardiovascular diseases and osteoporosis (50). The target proteins of SIRT1 include
those associated with regulating cell proliferation, DNA damage
repair and gene transcription (51).
A recent study suggested that SIRT1 regulates autophagy through
several cellular pathways, either directly or indirectly (35). Previous results have indicated that
activation of SIRT1 can induce autophagy, whereas SIRT1-deficient
mouse embryonic fibroblasts insufficiently activated autophagy
under starvation conditions (35,52). In
hepatocellular carcinoma cells, inhibition of SIRT1 was
demonstrated to impair cell proliferation and rapamycin-induced
autophagy (35). In addition,
overexpression of SIRT1 in neurons prevents the accumulation of the
prion protein and neurotoxicity by inducing autophagy (52). In human colorectal or cervical cancer
cells, SIRT1 is necessary for the induction of autophagy under
nutrient deprivation or caloric restriction conditions, whereas
knockdown of SIRT1 expression hampers the activation of autophagy
(53). The present data suggested
that SIRT1 was involved in alleviating the impairment of autophagy
caused by hydrogen. With siRNAs against SIRT1, the hydrogen-trigged
enhancing effect on autophagy was abolished in ox-LDL-treated
macrophages. In conclusion, the present study demonstrated that
ox-LDL decreased the expression of SIRT1, resulting in impaired
autophagic flux, which consequently resulted in the upregulation of
pro-inflammatory cytokines. Furthermore, the findings suggested
that hydrogen may enhance autophagy via increasing SIRT1 expression
and decreasing ox-LDL-induced inflammation. The present study
provided novel insights into the role of defective autophagy in the
pathogenesis of AS and identified autophagy as a promising
therapeutic target for the treatment of AS. Furthermore, hydrogen
may represent a potential agent for treating AS.
Acknowledgements
Not applicable.
Funding
The present study was supported by Tianjin Municipal
Health Bureau of Science and Technology Fund Projects (grant no.
2014KZ024).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
The current study was designed by JH and SY. SY, XL
and HL performed western blotting, ELISA and CCK8 assays. SY, XL
and JZ helped with hydrogen preparation and cell culture. The
statistical analysis was conducted by ML. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Moore KJ and Tabas I: Macrophages in the
pathogenesis of atherosclerosis. Cell. 145:341–355. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tuttolomondo A, Di Raimondo D, Pecoraro R,
Arnao V, Pinto A and Licata G: Atherosclerosis as an inflammatory
disease. Curr Pharm Des. 18:4266–4288. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Horio E, Kadomatsu T, Miyata K, Arai Y,
Hosokawa K, Doi Y, Ninomiya T, Horiguchi H, Endo M, Tabata M, et
al: Role of endothelial cell-derived angptl2 in vascular
inflammation leading to endothelial dysfunction and atherosclerosis
progression. Arterioscler Thromb Vasc Biol. 34:790–800. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sergin I and Razani B: Self-eating in the
plaque: What macrophage autophagy reveals about atherosclerosis.
Trends Endocrinol Metab. 25:225–234. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xue F, Nie X, Shi J, Liu Q, Wang Z, Li X,
Zhou J, Su J, Xue M, Chen WD and Wang YD: Quercetin inhibits
LPS-induced inflammation and ox-LDL-induced lipid deposition. Front
Pharmacol. 8:402017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang XH, Wang F, You SJ, Cao YJ, Cao LD,
Han Q, Liu CF and Hu LF: Dysregulation of cystathionine γ-lyase
(CSE)/hydrogen sulfide pathway contributes to ox-LDL-induced
inflammation in macrophage. Cell Signal. 25:2255–2262. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Du J, Huang Y, Yan H, Zhang Q, Zhao M, Zhu
M, Liu J, Chen SX, Bu D, Tang C and Jin H: Hydrogen sulfide
suppresses oxidized low-density lipoprotein (ox-LDL)-stimulated
monocyte chemoattractant protein 1 generation from macrophages via
the nuclear factor κB (NF-κB) pathway. J Biol Chem. 289:9741–9753.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang P, Shao BZ, Deng Z, Chen S, Yue Z and
Miao CY: Autophagy in ischemic stroke. Prog Neurobiol.
163–164:98–117. 2018. View Article : Google Scholar
|
|
9
|
Farré JC and Subramani S: Mechanistic
insights into selective autophagy pathways: Lessons from yeast. Nat
Rev Mol Cell Biol. 17:537–552. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fraldi A, Klein AD, Medina DL and
Settembre C: Brain disorders due to lysosomal dysfunction. Annu Rev
Neurosci. 39:277–295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Coutts AS and La Thangue NB: Actin
nucleation by WH2 domains at the autophagosome. Nat Commun.
6:78882015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Orogo AM and Gustafsson AB: Therapeutic
targeting of autophagy: Potential and concerns in treating
cardiovascular disease. Circ Res. 116:489–503. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qian M, Fang X and Wang X: Autophagy and
inflammation. Clin Transl Med. 6:242017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Riffelmacher T, Clarke A, Richter FC,
Stranks A, Pandey S, Danielli S, Hublitz P, Yu Z, Johnson E,
Schwerd T, et al: Autophagy-dependent generation of free fatty
acids is critical for normal neutrophil differentiation. Immunity.
47(466–480): e4652017.
|
|
15
|
Cadwell K: Crosstalk between autophagy and
inflammatory signalling pathways: Balancing defence and
homeostasis. Nat Rev Immunol. 16:661–675. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Munz C: Autophagy beyond intracellular MHC
class II antigen presentation. Trends Immunol. 37:755–763. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhong Z, Sanchez-Lopez E and Karin M:
Autophagy, inflammation, and immunity: A Troika governing cancer
and its treatment. Cell. 166:288–298. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao Z, Fux B, Goodwin M, Dunay IR, Strong
D, Miller BC, Cadwell K, Delgado MA, Ponpuak M, Green KG, et al:
Autophagosome-independent essential function for the autophagy
protein Atg5 in cellular immunity to intracellular pathogens. Cell
Host Microbe. 4:458–469. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Y, Cao X, Zhu W, Liu Z, Liu H, Zhou
Y, Cao Y, Liu C and Xie Y: Resveratrol enhances autophagic flux and
promotes Ox-LDL degradation in HUVECs via upregulation of SIRT1.
Oxid Med Cell Longev. 2016:75898132016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Razani B, Feng C, Coleman T, Emanuel R,
Wen H, Hwang S, Ting JP, Virgin HW, Kastan MB and Semenkovich CF:
Autophagy links inflammasomes to atherosclerotic progression. Cell
Metab. 15:534–544. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ohsawa I, Ishikawa M, Takahashi K,
Watanabe M, Nishimaki K, Yamagata K, Katsura K, Katayama Y, Asoh S
and Ohta S: Hydrogen acts as a therapeutic antioxidant by
selectively reducing cytotoxic oxygen radicals. Nat Med.
13:688–694. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang CS, Kawamura T, Toyoda Y and Nakao
A: Recent advances in hydrogen research as a therapeutic medical
gas. Free Radic Res. 44:971–982. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ge L, Wei LH, Du CQ, Song GH, Xue YZ, Shi
HS, Yang M, Yin XX, Li RT, Wang XE, et al: Hydrogen-rich saline
attenuates spinal cord hemisection-induced testicular injury in
rats. Oncotarget. 8:42314–42331. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang G, Gao S, Li X, Zhang L, Tan H, Xu
L, Chen Y, Geng Y, Lin Y, Aertker B and Sun Y: Pharmacological
Postconditioning with lactic acid and hydrogen rich saline
alleviates myocardial reperfusion injury in rats. Sci Rep.
5:98582015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zou R, Wang MH, Chen Y, Fan X, Yang B, Du
J, Wang XB, Liu KX and Zhou J: Hydrogen-rich saline attenuates
acute lung injury induced by limb ischemia/reperfusion via
down-regulating chemerin and NLRP3 in rats. Shock. 2018. View Article : Google Scholar
|
|
26
|
Chen J, Zhang H, Hu J, Gu Y, Shen Z, Xu L,
Jia X, Zhang X and Ding X: Hydrogen-rich saline alleviates kidney
fibrosis following AKI and retains Klotho expression. Front
Pharmacol. 8:4992017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen HG, Xie KL, Han HZ, Wang WN, Liu DQ,
Wang GL and Yu YH: Heme oxygenase-1 mediates the anti-inflammatory
effect of molecular hydrogen in LPS-stimulated RAW 264.7
macrophages. Int J Surg. 11:1060–1066. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zheng H and Yu YS: Chronic hydrogen-rich
saline treatment attenuates vascular dysfunction in spontaneous
hypertensive rats. Biochem Pharmacol. 83:1269–1277. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Du H, Sheng M, Wu L, Zhang Y, Shi D, Weng
Y, Xu R and Yu W: Hydrogen-rich saline attenuates acute kidney
injury after liver transplantation via activating p53-mediated
autophagy. Transplantation. 100:563–570. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bai X, Liu S, Yuan L, Xie Y, Li T, Wang L,
Wang X, Zhang T, Qin S, Song G, et al: Hydrogen-rich saline
mediates neuroprotection through the regulation of endoplasmic
reticulum stress and autophagy under hypoxia-ischemia neonatal
brain injury in mice. Brain Res. 1646:410–417. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ohsawa I, Nishimaki K, Yamagata K,
Ishikawa M and Ohta S: Consumption of hydrogen water prevents
atherosclerosis in apolipoprotein E knockout mice. Biochem Biophys
Res Commun. 377:1195–1198. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pankratz F, Hohnloser C, Bemtgen X,
Jaenich C, Kreuzaler S, Hoefer I, Pasterkamp G, Mastroianni J,
Zeiser R, Smolka C, et al: MicroRNA-100 suppresses chronic vascular
inflammation by stimulation of endothelial autophagy. Circ Res.
122:417–432. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ilyas G, Zhao E, Liu K, Lin Y, Tesfa L,
Tanaka KE and Czaja MJ: Macrophage autophagy limits acute toxic
liver injury in mice through down regulation of interleukin-1β. J
Hepatol. 64:118–127. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Z, Liu N, Liu K, Zhou G, Gan J, Wang
Z, Shi T, He W, Wang L, Guo T, et al: Autophagy mediated CoCrMo
particle-induced peri-implant osteolysis by promoting osteoblast
apoptosis. Autophagy. 11:2358–2369. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qiu G, Li X, Che X, Wei C, He S, Lu J, Jia
Z, Pang K and Fan L: SIRT1 is a regulator of autophagy:
Implications in gastric cancer progression and treatment. FEBS
Lett. 589:2034–2042. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee IH, Cao L, Mostoslavsky R, Lombard DB,
Liu J, Bruns NE, Tsokos M, Alt FW and Finkel T: A role for the
NAD-dependent deacetylase Sirt1 in the regulation of autophagy.
Proc Natl Acad Sci USA. 105:3374–3379. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Deng Z, Wang Z, Jin J, Wang Y, Bao N, Gao
Q and Zhao J: SIRT1 protects osteoblasts against particle-induced
inflammatory responses and apoptosis in aseptic prosthesis
loosening. Acta. Biomater. 49:541–554. 2017.
|
|
38
|
Banerjee C and Chimowitz MI: Stroke caused
by atherosclerosis of the major intracranial arteries. Circ Res.
120:502–513. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lonardo A, Nascimbeni F, Mantovani A and
Targher G: Hypertension, diabetes, atherosclerosis and NASH: Cause
or consequence? J Hepatol. 68:335–352. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Skochko OV and Kaidashev IP: Effect of
pioglitazone on insulin resistance, progression of atherosclerosis
and clinical course of coronary heart disease. Wiad Lek.
70:881–890. 2017.PubMed/NCBI
|
|
41
|
Cao J, Steffen BT, Guan W, Remaley AT,
McConnell JP, Palamalai V and Tsai MY: A comparison of three
apolipoprotein B methods and their associations with incident
coronary heart disease risk over a 12-year follow-up period: The
multi-ethnic study of atherosclerosis. J Clin Lipidol. 12:300–304.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hayashida K, Sano M, Ohsawa I, Shinmura K,
Tamaki K, Kimura K, Endo J, Katayama T, Kawamura A, Kohsaka S, et
al: Inhalation of hydrogen gas reduces infarct size in the rat
model of myocardial ischemia-reperfusion injury. Biochem Biophys
Res Commun. 373:30–35. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sun Q, Kang Z, Cai J, Liu W, Liu Y, Zhang
JH, Denoble PJ, Tao H and Sun X: Hydrogen-rich saline protects
myocardium against ischemia/reperfusion injury in rats. Exp Biol
Med (Maywood). 234:1212–1219. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lopez A, Lee SE, Wojta K, Ramos EM, Klein
E, Chen J, Boxer AL, Gorno-Tempini ML, Geschwind DH, Schlotawa L,
et al: A152T tau allele causes neurodegeneration that can be
ameliorated in a zebrafish model by autophagy induction. Brain.
140:1128–1146. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pott J, Kabat AM and Maloy KJ: Intestinal
epithelial cell autophagy is required to protect against
TNF-induced apoptosis during chronic colitis in mice. Cell Host
Microbe. 23(191–204): e42018.
|
|
46
|
Nabavi SF, Sureda A, Dehpour AR, Shirooie
S, Silva AS, Devi KP, Ahmed T, Ishaq N, Hashim R, Sobarzo-Sanchez
E, et al: Regulation of autophagy by polyphenols: Paving the road
for treatment of neurodegeneration. Biotechnol Adv. 2017.
|
|
47
|
Levine B, Mizushima N and Virgin HW:
Autophagy in immunity and inflammation. Nature. 469:323–335. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu K, Zhao E, Ilyas G, Lalazar G, Lin Y,
Haseeb M, Tanaka KE and Czaja MJ: Impaired macrophage autophagy
increases the immune response in obese mice by promoting
proinflammatory macrophage polarization. Autophagy. 11:271–284.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Saitoh T, Fujita N, Jang MH, Uematsu S,
Yang BG, Satoh T, Omori H, Noda T, Yamamoto N, Komatsu M, et al:
Loss of the autophagy protein Atg16L1 enhances endotoxin-induced
IL-1beta production. Nature. 456:264–268. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zeng L, Chen R, Liang F, Tsuchiya H, Murai
H, Nakahashi T, Iwai K, Takahashi T, Kanda T and Morimoto S: Silent
information regulator, Sirtuin 1, and age-related diseases. Geriatr
Gerontol Int. 9:7–15. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang RH, Sengupta K, Li C, Kim HS, Cao L,
Xiao C, Kim S, Xu X, Zheng Y, Chilton B, et al: Impaired DNA damage
response, genome instability, and tumorigenesis in SIRT1 mutant
mice. Cancer Cell. 14:312–323. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Jeong JK, Moon MH, Lee YJ, Seol JW and
Park SY: Autophagy induced by the class III histone deacetylase
Sirt1 prevents prion peptide neurotoxicity. Neurobiol Aging.
34:146–156. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Morselli E, Maiuri MC, Markaki M, Megalou
E, Pasparaki A, Palikaras K, Criollo A, Galluzzi L, Malik SA,
Vitale I, et al: Caloric restriction and resveratrol promote
longevity through the Sirtuin-1-dependent induction of autophagy.
Cell Death Dis. 1:e102010. View Article : Google Scholar : PubMed/NCBI
|