Introduction
Polycystic ovarian syndrome (PCOS) is the most
common endocrine and metabolic disease in women, occurring in ~15%
at procreation age (1). Its
characteristics include hyperandrogenism, ovulation failure and
polycystic ovarian morphology (2),
combined with multiple clinical symptoms including menoxenia, acne,
hirsutism, alopecia, central obesity and infertility (3). PCOS, is a life-long disease, which not
only causes short-term problems, including infertility due to
anovulation, but also exhibits long-term metabolic issues including
obesity, insulin resistance, non-alcoholic fatty liver disease,
type 2 diabetes mellitus and cardiac vascular disease (4), as well as hyperinsulinemia and estrogen
dependent cancer (5).
Hyperandrogenism, occasionally combined with
hypoestrogenism, is a classic diagnostic criterion for PCOS, as
excess androgen induces the dysfunction of the
hypothalamic-pituitary-ovarian axis, which is the neurohormonal
system that regulates gonadal hormone balance, follicle maturation
and ovulation (6). Mainstream
therapy including hormonal therapy, antiandrogens and
insulin-sensitizing drugs used to reduce androgen levels is usually
combined with oral contraceptives (7). Among them, diane-35 (a combination of
ethinyl estradiol and cyproterone acetate) is commonly used to
treat PCOS (8).
However, diane-35 produces significant hormonal side
effects and negatively impacts triglyceride metabolism and
pancreatic function (9).
Salvia miltiorrhiza (S. miltiorrhiza), a
Chinese herbal medicine, has been used to treat PCOS. Components
isolated from S. miltiorrhiza, including cryptotanshinone,
attenuate PCOS by reversing dexamethasone-induced ovarian insulin
resistance in mice (10). However,
whether other components of S. miltiorrhiza have the
capability to treat PCOS remains unclear. Tanshinone IIA (TSIIA) is
another primary component of S. miltiorrhiza (11). Serving as an antioxidant,
anti-inflammatory and immunomodulatory element, it is primarily
used to treat cardiovascular diseases (12). TSIIA also inhibits oxidative stress
injury in neuronal cells to treat Alzheimer's disease (13). The results of the present study
confirmed the therapeutic effect of TSIIA on PCOS induced by
neonatal exposure to estradiol in mice by regulating
follicle-stimulating hormone (FSH) receptor (FSHR) expression in
the ovary.
Materials and methods
Animals
Animal care and experiment procedures were performed
in accordance with the guidelines of and approved by the Animal
Care and Ethical Committee of Affiliated Hospital of Nanjing
University of Chinese Medicine (Nanjing, China). A total of 57
female (C57Bl/6J × A/J) F1 (B6A) 5-day-old mice (weight, 4–5 g)
purchased from Beijing Vital River Laboratory Animal Technology
Co., Ltd. (Beijing, China) were used in the current study. All mice
were maintained in standard cages under a 12 h light/dark cycle at
a temperature of 22–24°C and a relative humidity of 55–65%, with
ad libitum access to food and water.
Experimental design and drug
administration
Estradiol (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) dissolved in dimethyl sulfoxide and diluted with sesame
oil (BSZH Co. Ltd., Beijing, China), was subcutaneously injected
into 5-day-old female mice (PCOS mice, n=56) at a dose of 20 µg/day
for 3 days (14). Vehicle of the
same volume was administrated to mice (control group, n=28). On the
56th postnatal day, half PCOS mice (estradiol+TSIIA group, n=28)
received TSIIA (Nanjing Spring and Autumn Biological Engineering
Co. Ltd, Nanjing, China) for 4 weeks at a dose of 100 mg/kg/day by
gavage. The PCOS mice left (estradiol group, n=28) and control mice
received the same weight-based volume of vehicle. Estrous cyclicity
was determined during the last 18 days prior to sacrifice.
Thereafter, mice were weighed (22–25 g) and sacrificed on the 96th
postnatal day by decapitation.
Assessment of estrous cycles
Estrous cycles were examined daily at 0800–0900 h.
The fluid obtained by vaginal lavage with 0.9% saline was spotted
thinly on a microscope slide. Following air-drying, slides were
stained with toluidine blue (0.1%) at room temperature for 2 sec.
According to the types of vaginal epithelial cells present
(leukocyte, nucleated and cornified cells), diestrus, estrus and
proestrus were identified using light microscopy at magnification,
×100 as previously described (15).
Ovarian histology
Ovaries were excised from mice, fixed in 4%
paraformaldehyde at room temperature for 48 h, dehydrated in
ascending grades of ethanol and embedded in paraffin in 5-µm
sections. Samples were then deparaffinized, rehydrated and stained
with hematoxylin and eosin (H&E) for 3 and 0.5 min at room
temperature, respectively. Follicles were counted using a
conventional light microscope (Olympus DP70; Olympus Corporation,
Tokyo, Japan) with ×40 objective. The number of follicles (atretic
cyst-like, large antral and preovulatory follicles) were counted in
every sixth section (30 µm apart) and multiplied by 6 to provide
the total number of follicles in each ovary. Only follicles
containing an oocyte with a visible nucleus were counted to avoid
double counting. The classification of follicular stages was made
following the morphological criteria as described previously
(16,17). The number of corpora lutea was scored
in a blinded fashion using one section per ovary and one ovary per
mouse (18).
Hormonal measurements
Serum was obtained when mice were sacrificed
following the determination of estrous cyclicity. Levels of FSH,
luteinizing hormone (LH), progesterone (P), estradiol and
testosterone (T) were quantified using ELISA kits (cat. nos.
E0830Mu, E0441Mu, E0459Mu, E0461Mu and E0458Mu, respectively; Uscn
Life Sciences, Inc., Wuhan, China) according to the manufacturer's
protocol.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from murine ovary tissue
using a TRIzol reagent kit (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) according to the manufacturer's protocol.
RNA (1 µg) was transcribed into cDNA using a PrimeScript™ RT
reagent kit (Takara Biotechnology Co., Ltd., Dalian, China). qPCR
reactions were performed using a Light Cycler Fast Start DNA Master
SYBR-Green I kit and an ABI Prism 7300 Sequence Detection System
(Applied Biosystems; Thermo Fisher Scientific, Inc.) The
thermocycling conditions were as follows: 95°C for 3 min; 95°C for
15 sec; 60°C for 15 sec; 72°C for 1 min (35 cycles); and 72°C for
10 min. The relative expression of genes was determined using the
2−ΔΔCq method (19) with
normalization to 36B4 expression. The primer sequences were as
follows: LH receptor (LHR) forward, 5′-AATGAGTCCATCACGCTGAAAC-3′
and reverse, 5′-CCTGCAATTTGGTGGAAGAGA-3′; FSHR forward,
5′-CCTTGCTCCTGGTCTCCTTG-3′ and reverse,
5′-CTCGGTCACCTTGCTATCTTG-3′; aromatase forward,
5′-ATGTTCTTGGAAATGCTGAACCC-3′ and reverse,
5′-AGGACCTGGTATTGAAGACGAG-3′; peroxisome proliferator-activated
receptor γ (PPARγ) forward, 5′-TTTTCCGAAGAACCATCCGATT-3′ and
reverse, 5′-ATGGCATTGTGAGACATCCCC-3′; and 34B4 forward,
5′-AAGCGCGTCCTGGCATTGTCT-3′ and reverse,
5′-CCGCAGGGGCAGCAGTGGT-3′.
Isolation of granule cells
As described previously (20), ovaries were isolated from mice and
the large and transparent follicles residing within were punctured
to liberate granule cells into the medium, Dulbecco's modified
Eagle's medium (DMEM)/nutrient mixture F-12 (F12; Gibco; Thermo
Fisher Scientific, Inc.). Granule cells were mixed and collected
via centrifugation at 100 × g at room temperature for 2 min. Cells
were then suspended in DMEM/F12 complete medium containing 10%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and
seeded into 12-well plates at 3×105 cells/well for 48 h.
Medium was changed every 24 h to discard non-adherent granulosa
cells and oocytes. Cells were further cultured at 37°C for 24 h for
further experimentation.
Western blotting
Granule cells were homogenized in ice-cold lysis
buffer [50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 2 mM EDTA, 1% Nonidet
P-40, 10 mM
Na4P2O7·10H2O, and
protein inhibitors including 100 mM NaF, 1 mM
Na3VO4, 1 mM phenylmethylsulfonyl fluoride,
10 mg/ml aprotinin, 10 mg/ml leupeptin; all components purchased
from Aladdin Shanghai Biochemical Technology Co., Ltd., Shanghai,
China) and boiled at 95°C for 5 min in 5X loading buffer [250 mM
Tris-HCl (pH 6.8), 10% SDS, 0.5% bromophenol blue, 50% glycerol, 5%
mercaptoethanol; all components purchased from Aladdin Shanghai
Biochemical Technology Co., Ltd.]. Protein concentration was
assessed using a bicinchoninic acid protein assay kit (Pierce;
Thermo Fisher Scientific, Inc.). Protein (25 µg) was separated by
10% SDS-PAGE and transferred to polyvinylidene difluoride
membranes. The membranes were incubated with rabbit anti-FSHR
antibodies (1:1,000; cat. no. K002799P; Beijing Solarbio Science
and Technology Co., Ltd., Beijing, China), anti-LHR antibodies
(1:1,000; cat. no. sc-25828; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA), anti-aromatase antibodies (1:1,000; cat. no.
PA1-21398; Invitrogen; Thermo Fisher Scientific, Inc.) and mouse
anti-β-actin (1:1,000; cat. no. CBL171-I; EMD Millipore, Billerica,
MA, USA) at 4°C overnight. Membranes were then further incubated
with horseradish peroxidase-conjugated goat anti-rabbit
Immunoglobulin G (IgG; 1:5,000; cat. no. AP183P) or goat anti-mouse
IgG (1:5,000; cat. no. AP192P; both EMD Millipore) for 2 h at room
temperature. Membranes were visualized using the Tanon-5200
Chemiluminescence Imager (Tanon Science and Technology, Co., Ltd.,
Shanghai, China) with an ECL Western blotting substrate (EMD
Millipore). ImageJ version 1.48 (National Institutes of Health,
Bethesda, MA, USA) was used for the densitometry analysis of
bands.
Cyclic adenosine monophosphate (cAMP)
levels in granule cells
Following culture for 24 h, non-adherent granule
cells were removed and adherent cells were treated with FSH (100
IU/l; Ningbo Second Hormone Factory, Ningbo, China) for 10 min at
37°C. Cells were then rinsed with cold PBS once and lysed using 0.1
M hydrochloric acid for the protein concentration assay with total
protein assay kit (cat. no. A045-4; Nanjing Jiancheng
Bioengineering Institute, Nanjing, China). Whole cell lysate was
centrifuged at 10,000 × g at 4°C for 10 min and the supernatant was
used for cAMP measurement using a cAMP ELISA kit (cat. no.
10R-2133; Fitzgerald Industries International, North Acton, MA,
USA). Total cAMP amount was normalized to total protein amount in
each sample to calculate cAMP level in cells.
Statistical analysis
Data were expressed as the mean ± standard error of
the mean. Data between groups were analyzed using one-way analysis
of variance followed by Dunn's multiple comparisons test. P<0.05
was considered to indicate a statistically significant result.
Results
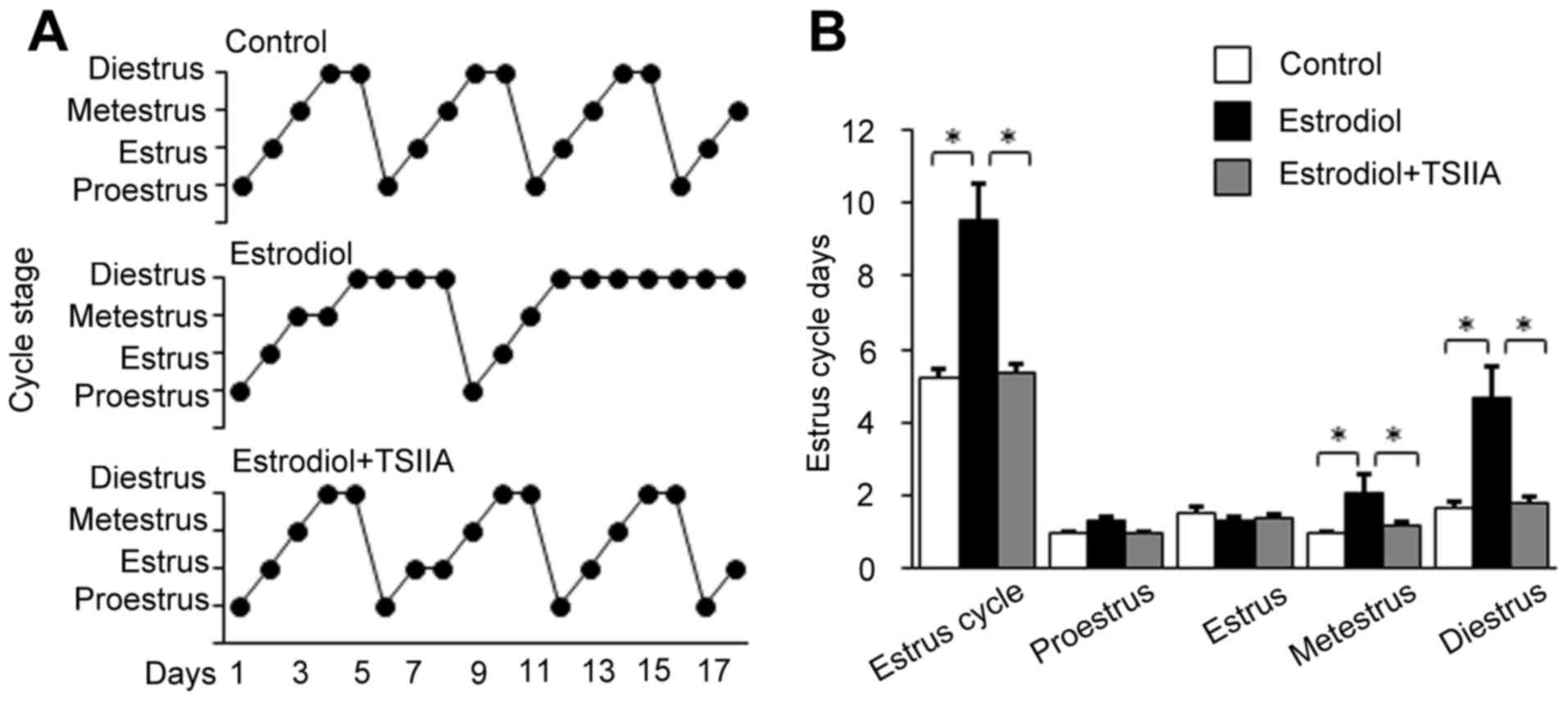
TSIIA attenuates estrous cycle
disorder in mice with PCOS
Normal control mice exhibited a regular estrous
cycle of 5 days on average (Fig.
1A). The entire estrous cycle comprises four phases, which
includes proestrus, estrus, metestrus and diestrus. Neonatal
estradiol-treatment increased metestrus and diestrus compared with
control mice, which contributed to the prolongation of the estrous
cycle to ~9 days (Fig. 1A and B).
TSIIA treatment significantly reduced entire estrous cycle,
metestrus and diestrus and led to a slight increase in the estrous
period (Fig. 1A and B).
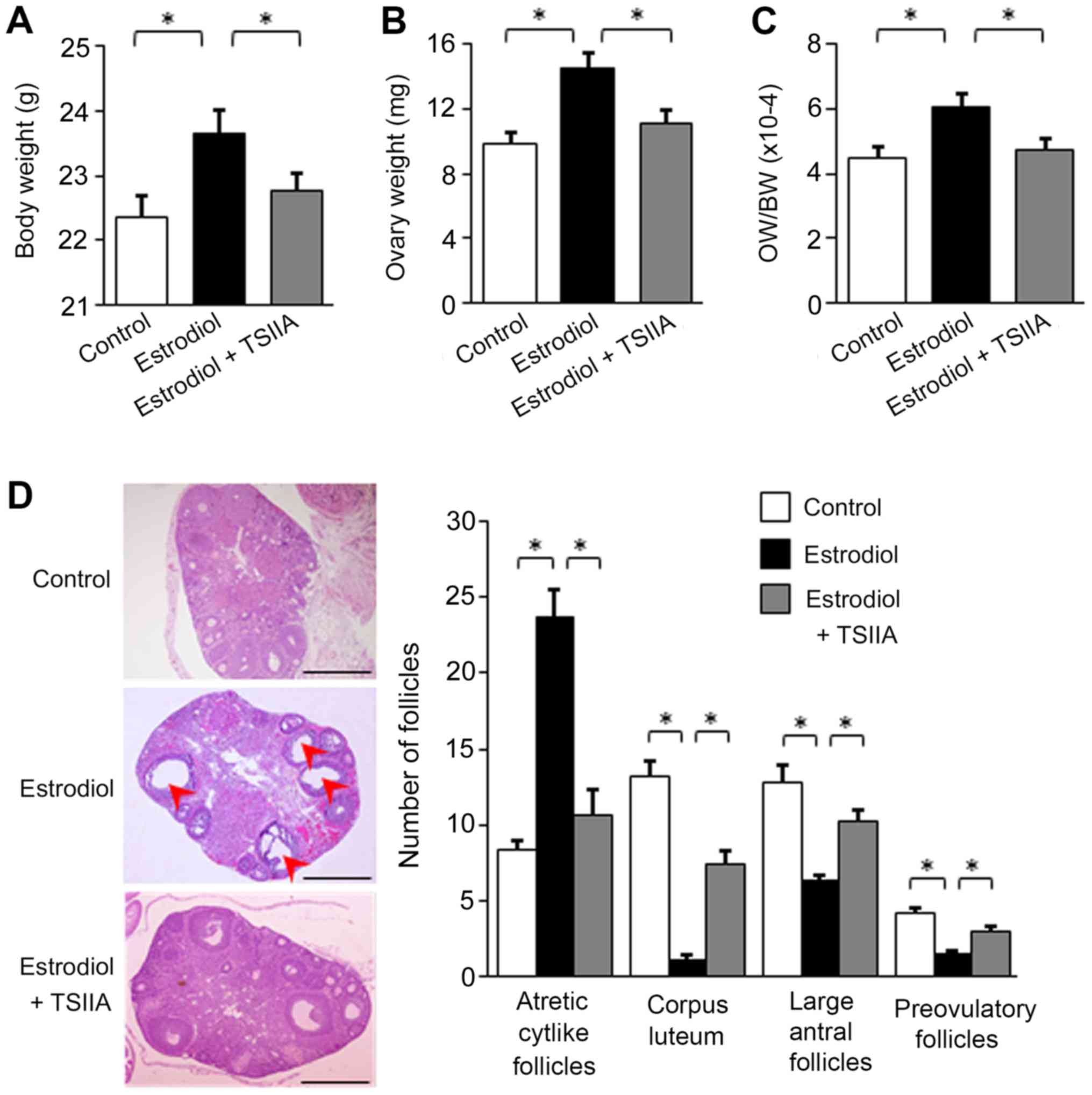
Effects of TSIIA on ovarian morphology
in mice with PCOS
Body weight and ovary weight in each group were
assayed following sacrifice. The results revealed that neonatal
estradiol treatment increased body and ovarian weight in mice
(Fig. 2A and B). The increase in
ovarian weight was more pronounced than that of body weight, and
the ratio of ovary to body weight also significantly increased
following estradiol treatment (Fig.
2C). TSIIA treatment reversed the estradiol-induced increase in
body weight, ovary weight and the ratio of ovary to body weight
(Fig. 2A-C).
Pathological changes in murine ovaries were assessed
via H&E staining. In congruence with a previous study (14), multiple enlarged follicles were
observed in the ovaries of estradiol-treated mice in the present
study (Fig. 2D), which mimicked
PCOS. Neonatal estradiol-treatment reduced the number of large
antral follicles, preovulatory follicles and corpus luteum, but
increased the number of atretic cyst-like follicles (Fig. 2D), which were also similar to changes
typically observed in PCOS (6).
TSIIA treatment reversed the observed pathological change in the
number of follicles (Fig. 2D).
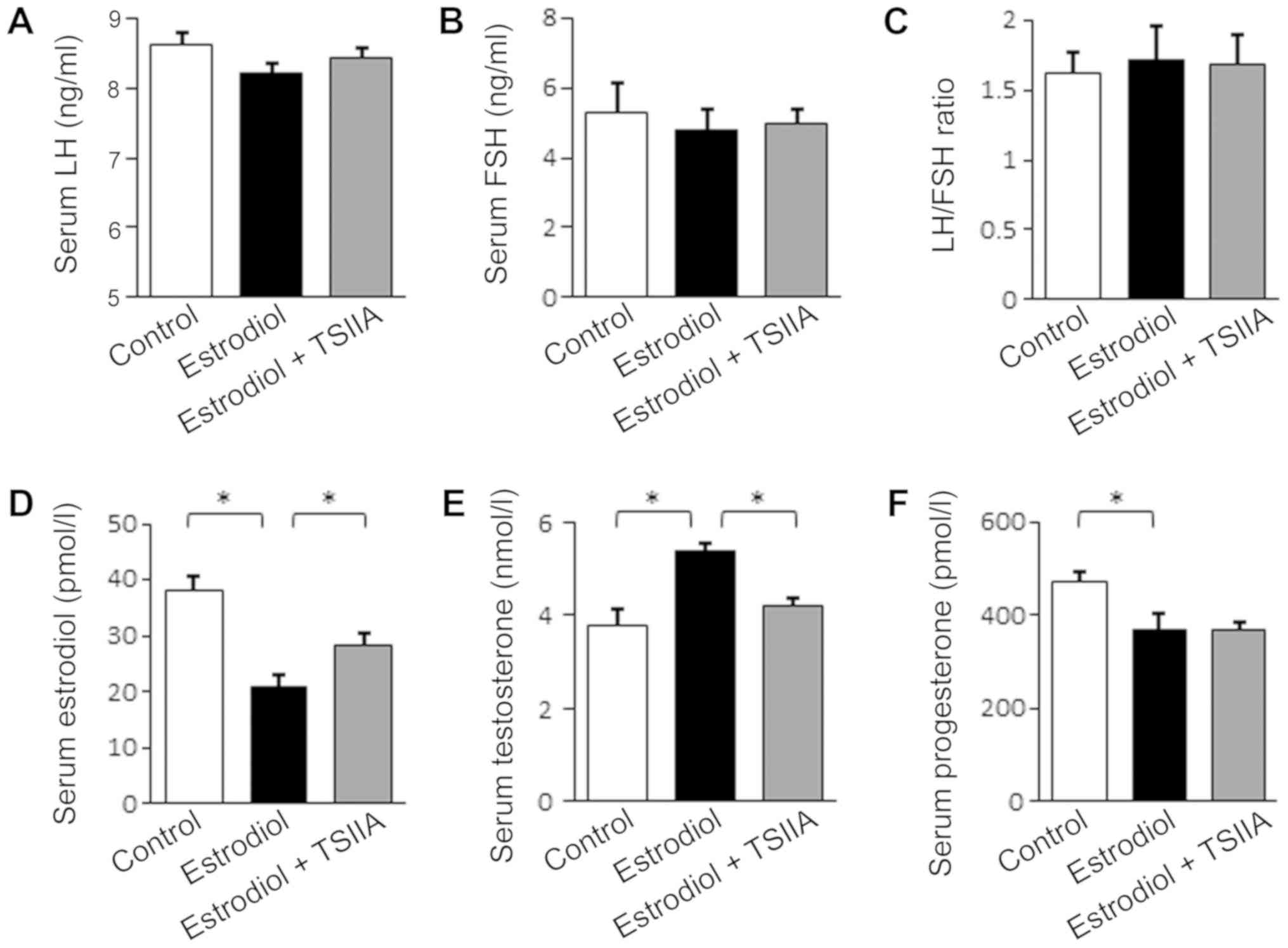
TSIIA regulates serum hormone levels
in mice with PCOS
Ovarian structure and function is primarily
regulated by serum hormones (6).
Thus, levels of FSH, LH, estradiol, T and P in murine serum were
assayed. As presented in Fig. 3A-C,
serum levels of LH, FSH and the ratio of LH to FSH were not
significantly affected by neonatal estradiol-treatment nor by
TSIIA. However, serum estradiol was reduced and T levels were
elevated following neonatal estradiol treatment, which were each
reversed when further treated with TSIIA (Fig. 3D and E). Serum P levels were also
reduced following neonatal estradiol-treatment, but TSIIA
demonstrated no significant effect.
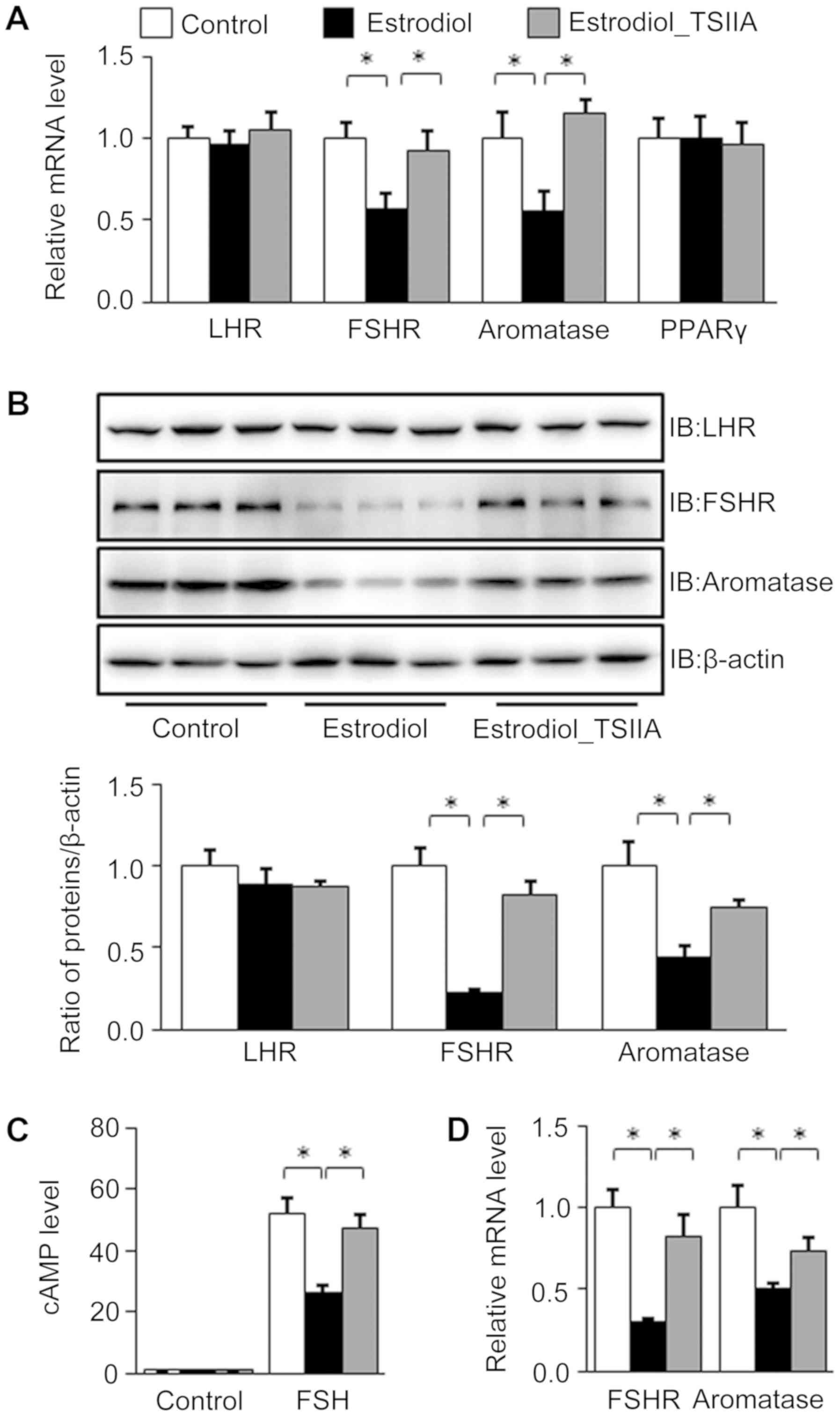
Effect of TSIIA on LHR, FSHR,
aromatase and PPARγ expression in the ovaries of mice with
PCOS
Neonatal mice treated with estradiol exhibited
significantly lower mRNA levels of aromatase, the key enzyme for
the transferal of T to estradiol in ovaries (21). However, mRNA levels of ovarian
aromatase recovered following TSIIA treatment. FSHR, LHR and PPARγ
function to regulate the expression of aromatase in the ovary
(22–24); however, only FSHR exhibited
significantly lower mRNA levels following neonatal estradiol
treatment, which was recovered by TSIIA (Fig. 4A).
In ovarian granule cells (Fig. 4B-D) of neonatal estradiol-treatment
mice, western blot analysis revealed lower FSHR and aromatase
protein levels, which were improved following TSIIA treatment
(Fig. 4B).
The FSH-induced cAMP levels in granule cells
isolated from estradiol-treated mice were significantly lower than
that from control mice, whereas TSIIA treatment elevated the
granular cAMP level in estradiol-treated group similar to control
group (Fig. 4C).
The results also demonstrated that granular FSHR and
aromatase mRNA levels in estradiol treated group were significantly
lower than in the control group, but were reversed following TSIIA
treatment (Fig. 4D).
Discussion
The present study revealed the therapeutic effect of
TSIIA, a component isolated from Salvia miltiorrhiza, on
murine PCOS induced by the neonatal treatment of estradiol.
In a previous study (25), PCOS was successfully replicated in
mice via neonatal treatment with estradiol. This was determined as
mice exhibited a prolonged diestrus phase, enlarged ovaries, a
thickened theca layer, a decreased number of corpus luteums and
preovulatory follicles, an increased number of atretic follicles
and multiple cysts in ovaries, which are characteristics similar to
those in women with PCOS (19). In
the current study, estrus cycles were returned to normal status
following TSIIA treatment and this also reversed the pathological
change of murine ovaries with estradiol-induced PCOS.
The alteration of granulosa cell hormonal
equilibrium is a cause of disturbed follicular development and
maturation that results in the premature arrest of follicular
growth in PCOS (26,27). Estrogen suppresses granulosa and
luteal cell apoptosis (28,29) and triggers the release of FSH and LH,
promoting follicle maturation and ovulation (30). An excess of intraovarian androgen
leads to follicular atresia (28),
as it activates primordial follicles via the
phosphatidylinositol-3-kinase/protein kinase B/Forkhead box O3a
pathway and inhibits growth differentiation factor 9 expression in
oocytes, resulting in the arrest of preantral follicle development
(31).
In the present study, neonatal treatment of
estradiol decreased serum estradiol but increased serum T levels,
which may lead to pathological and functional changes in the ovary.
Thus, the therapeutic effect of TSIIA observed in the current study
may be the result of disordered estradiol and T, as other hormones
affecting follicle maturation, including LH, FSH and P, were either
not changed or not reversed following TSIIA therapy.
The hormonal balance between estrogen and androgen
is primarily sustained by the normal function of granulosa cells,
which contain a series of enzymes converting T to estradiol.
Disorder in the activity or expression of aromatase, the key enzyme
catalyzing the conversion of T to estradiol, induces excessive
androgen accumulation in serum (32). Thus, aromatase mRNA levels of the
ovaries and granule cells were determined in the current study. It
was demonstrated that they were reduced by neonatal
estradiol-treatment, but ameliorated following TSIIA therapy. This
implies that TSIIA targets the expression of aromatase to revise
the T/estradiol balance in mice.
The transcription of aromatase is regulated by the
LH, FSH and PPARγ pathways (22–24). LH
and FSH mediate the down- or upregulation of aromatase via LHR and
FSHR, respectively. Since the serum levels of LH and FSH were not
affected by neonatal estradiol treatment or TSIIA in the current
study, the expression of LHR and FSHR should predict the
transcription of aromatase. FSHR mRNA levels were reduced by
neonatal estradiol treatment, which was then ameliorated by TSIIA.
However, neither LHR nor PPARγ expression was affected. The results
indicate that the downregulation of FSHR induced PCOS in the
present murine model and that TSIIA exerted its therapeutic effect
by targeting FSHR expression. Furthermore, the present study
revealed that estradiol treatment reduced FSHR and aromatase
protein levels, thus inhibiting FSH-induced cAMP elevation in
granule cells isolated from mice. TSIIA treatment also ameliorated
FSHR expression and an FSH induced cAMP increase. As a result,
TSIIA primarily affected the ovarian expression of FSHR, which
improved ovarian function.
In summary, the present study revealed that PCOS
induced by neonatal exposure to estradiol in mice is attenuated by
TSIIA treatment. The therapeutic effect of TSIIA is likely due to
the recovery of FSHR expression in the ovary, thus leading to
elevated serum levels of estradiol and T, an improvement in the
structure of ovaries and the attenuation of the disordered estrous
cycle. The current study revealed a novel monomer (TSIIA), which
may be utilized to further the application of natural drugs in the
treatment of PCOS to reduce the side effects of hormone drugs.
However, the present study was limited by the lack of a positive
control group and intensive studies in granule cells. Further study
is therefore required to assess the mechanism of how TSIIA
upregulates FSHR in granule cells.
Acknowledgements
The authors would like to thank Professor Liang
Sheng from Nanjing Medical University (Nanjing, China) for
assisting with the experimental design and modifications to the
manuscript.
Funding
The present study was supported by the Natural
Science Foundation of China (grant nos. 81473713 and 81774354),
Jiangsu Provincial Hospital of TCM (grant no. Y18070) and Leading
talents of Chinese medicine in Jiangsu province (grant no.
SLJ0202).
Availability of data and materials
All data and materials supported the results of the
present study are available in the published article.
Authors' contributions
HFZ was responsible for the experimental design and
drafting the manuscript. JJ, QYH, WWX, WJZ, BL, JL and WW performed
the experiments and data analysis. HFZ reviewed the manuscript. All
authors gave final approval of the version to be published.
Ethics approval and consent to
participate
Animal care and experimental procedures were
approved by and performed in accordance with the guidelines of the
Animal Care and Ethics Committee of Affiliated Hospital of Nanjing
University of Chinese Medicine.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fauser BC, Tarlatzis BC, Rebar RW, Legro
RS, Balen AH, Lobo R, Carmina E, Chang J, Yildiz BO, Laven JS, et
al: Consensus on women's health aspects of polycystic ovary
syndrome (PCOS): The Amsterdam ESHRE/ASRM-Sponsored 3rd PCOS
Consensus Workshop Group. Fertil Steril. 97:28–38.e25. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Abbott DH, Tarantal AF and Dumesic DA:
Fetal, infant, adolescent and adult phenotypes of polycystic ovary
syndrome in prenatally androgenized female rhesus monkeys. Am J
Primatol. 71:776–784. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jones GL, Hall JM, Balen AH and Ledger WL:
Health-related quality of life measurement in women with polycystic
ovary syndrome: A systematic review. Human Reprod Update. 14:15–25.
2008. View Article : Google Scholar
|
|
4
|
Jeanes YM and Reeves S: Metabolic
consequences of obesity and insulin resistance in polycystic ovary
syndrome: Diagnostic and methodological challenges. Nutr Res Rev.
30:97–105. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lauretta R, Lanzolla G, Vici P, Mariani L,
Moretti C and Appetecchia M: Insulin-sensitizers, polycystic ovary
syndrome and gynaecological cancer risk. Int J Endocrinol.
2016:86717622016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
De Leo V, Musacchio MC, Cappelli V,
Massaro MG, Morgante G and Petraglia F: Genetic, hormonal and
metabolic aspects of PCOS: An update. Reprod Biol Endocrinol.
14:382016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rocca ML, Venturella R, Mocciaro R, Di
Cello A, Sacchinelli A, Russo V, Trapasso S, Zullo F and Morelli M:
Polycystic ovary syndrome: Chemical pharmacotherapy. Expert Opin
Pharmacother. 16:1369–1393. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Saeed R, Akram J, Changezi HU and Saeed M:
Treatment of hirsutism in polycystic ovarian syndrome with Diane,
50 mcg ethinyl estradiol and 2 mg cyproterone acetate. Specialist.
9:109–112. 1993.
|
|
9
|
Jing Z, Liang-Zhi X, Tai-Xiang W, Ying T
and Yu-Jian J: The effects of Diane-35 and metformin in treatment
of polycystic ovary syndrome: An updated systematic review. Gynecol
Endocrinol. 24:590–600. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang Y, Li W, Wang CC, Wu X and Zheng J:
Cryptotanshinone reverses ovarian insulin resistance in mice
through activation of insulin signaling and the regulation of
glucose transporters and hormone synthesizing enzymes. Fertil
Steril. 102:589–596.e4. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li P, Wang GJ, Li J, Hao HP and Zheng CN:
Identification of tanshinone IIA metabolites in rat liver
microsomes by liquid chromatography-tandem mass spectrometry. J
Chromatogr A. 1104:366–369. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Z and Xu H: Anti-inflammatory and
immunomodulatory mechanism of tanshinone IIA for atherosclerosis.
Evid Based Complement Alternat Med. 2014:2679762014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang ZY, Liu JG, Li H and Yang HM:
Pharmacological effects of active components of chinese herbal
medicine in the treatment of Alzheimer's disease: A review. Am J
Chin Med. 44:1525–1541. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chapman JC, Min SH, Freeh SM and Michael
SD: The estrogen-injected female mouse: New insight into the
etiology of PCOS. Reprod Biol Endocrinol. 7:472009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Goldman JM, Murr AS and Cooper RL: The
rodent estrous cycle: Characterization of vaginal cytology and its
utility in toxicological studies. Birth Defects Res B Dev Reprod
Toxicol. 80:84–97. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Caldwell AS, Middleton LJ, Jimenez M,
Desai R, McMahon AC, Allan CM, Handelsman DJ and Walters K:
Characterization of reproductive, metabolic, and endocrine features
of polycystic ovary syndrome in female hyperandrogenic mouse
models. Endocrinology. 155:3146–3159. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Myers M, Britt KL, Wreford NG, Ebling FJ
and Kerr JB: Methods for quantifying follicular numbers within the
mouse ovary. Reproduction. 127:569–580. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Glidewell-Kenney C, Hurley LA, Pfaff L,
Weiss J, Levine JE and Jameson JL: Nonclassical estrogen receptor
alpha signaling mediates negative feedback in the female mouse
reproductive axis. Proc Natl Acad Sci USA. 104:8173–8177. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liang N, Xu Y, Yin Y, Yao G, Tian H, Wang
G, Lian J, Wang Y and Sun F: Steroidogenic Factor-1 is required for
TGF-β3-mediated 17β-estradiol synthesis in mouse ovarian granulosa
cells. Endocrinology. 152:3213–3225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Simpson ER and Davis SR: Minireview:
Aromatase and the regulation of estrogen biosynthesis-some new
perspectives. Endocrinology. 142:4589–4594. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fan W, Yanase T, Morinaga H, Mu YM, Nomura
M, Okabe T, Goto K, Harada N and Nawata H: Activation of peroxisome
proliferator-activated receptor-gamma and retinoid X receptor
inhibits aromatase transcription via nuclear factor-kappaB.
Endocrinology. 146:85–92. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Komar CM: Peroxisome
proliferator-activated receptors (PPARs) and ovarian
function-implications for regulating steroidogenesis,
differentiation, and tissue remodeling. Reprod Biol Endocrinol.
3:412005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Komar CM, Braissant O, Wahli W and Curry
TE Jr: Expression and localization of PPARs in the rat ovary during
follicular development and the periovulatory period. Endocrinology.
142:48312001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
van Houten EL and Visser JA: Mouse models
to study polycystic ovary syndrome: A possible link between
metabolism and ovarian function. Reprod Biol. 14:32–43. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lombardi LA, Simãμes RS, Maganhin CC,
Baracat MC, Silva-Sasso GR, Florencio-Silva R, Soares JM Jr and
Baracat EC: Immunohistochemical evaluation of proliferation,
apoptosis and steroidogenic enzymes in the ovary of rats with
polycystic ovary. Rev Assoc Med Bras (1992). 60:349–356. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Salilew-Wondim D, Wang Q, Tesfaye D,
Schellander K, Hoelker M, Hossain MM and Tsang BK: Polycystic
ovarian syndrome is accompanied by repression of gene signatures
associated with biosynthesis and metabolism of steroids,
cholesterol and lipids. J Ovarian Res. 8:242015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Billig H, Furuta I and Hsueh AJ: Estrogens
inhibit and androgens enhance ovarian granulosa cell apoptosis.
Endocrinology. 133:2204–2212. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Goodman SB, Kugu K, Chen SH, Preutthipan
S, Tilly KI, Tilly JL and Dharmarajan AM: Estradiol-mediated
suppression of apoptosis in the rabbit corpus luteum is associated
with a shift in expression of bcl-2 family members favoring
cellular survival. Biol Reprod. 59:820–827. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Caligioni CS: Assessing reproductive
status/stages in mice. Curr Protoc Neurosci. 4:Appendix 4I. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang JL, Zhang CP, Li L, Huang L, Ji SY,
Lu CL, Fan CH, Cai H, Ren Y, Hu ZY, et al: Testosterone induces
redistribution of forkhead box-3a and down-regulation of growth and
differentiation factor 9 messenger ribonucleic acid expression at
early stage of mouse folliculogenesis. Endocrinology. 151:774–782.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Capellino S, Straub RH and Cutolo M:
Aromatase and regulation of the estrogen-to-androgen ratio in
synovial tissue inflammation: Common pathway in both sexes. Ann N Y
Acad Sci. 1317:24–31. 2014. View Article : Google Scholar : PubMed/NCBI
|