Introduction
Distal arthrogryposis 3 (DA3)/Gordon syndrome (GS),
DA5 and Marden-Walker syndrome (MWS) share a broad spectrum of
similar phenotypes to describe congenital contractures of multiple
joints that mainly involve congenital contractures of hands and
feet, cleft palate, ptosis, cerebellar malformations,
ophthalmoplegia, as well as pulmonary hypertension, which may be
caused by decreased intrauterine movement, or due to neurological,
muscle or connective tissue development disease (1–8).
Previous studies also indicated that mechanotransduction is
important for these biological and pathological processes,
including sensory perception and embryonic development of organs,
which is mediated by mechanosensation in proprioceptors, including
muscle spindles in or the Golgi tendon organs in tendons that are
able to sense mechanical forces upon cell membranes through
mechanically activated ion channels, and propagate proprioceptive
information by different nerve fibers (9).
Piezo type mechanosensitive ion channels (PIEZOs),
including PIEZO component 1 (PIEZO1) and PIEZO2, are very large
proteins with numerous predicted transmembrane domains per subunit,
and are evolutionarily conserved in plants and animals (10). PIEZOs are expressed in a broad range
of different tissue and cell types, including urinary bladder,
lungs, kidneys, cartilage and dorsal root ganglion (DRGs) (11). In 2010, PIEZO1 and −2 were identified
as the mechanically activated ion channels, and to have crucial
roles in numerous mechanotransduction processes, including touch
perception, proprioception and vascular development (12). A previous study indicated that
constitutive deletion of PIEZO1 and −2 in mice leads to
developmental lethality (13).
PIEZO1 has a crucial role in the development of the mouse
vasculature, and is required for mechanical force-induced cation
influx in red blood cells (14,15).
PIEZO2 is expressed in a subset of DRG neurons that innervate the
skin, hair follicles and Merkel cells to form low-threshold
mechanoreceptors for the detection of light touch in mammals
(13,16–18).
PIEZO2 is also a principal mechanically activated mechanotransducer
in low-threshold skeletal muscle-innervating proprioceptors in mice
(17).
Mutations in PIEZO2 have been reported to cause DA3,
DA5 and MWS. Numerous dominant gain-of-function mutations, as well
as recessive loss-of-function mutations, have been reported,
including gain-of-function mutations that destabilize inactivation
structures and lead to an overall increase of calcium influx, and
frameshift mutations and out-of-frame exon skipping that lead to
termination of protein synthesis through premature termination
codons or nonsense-mediated decay of PIEZO2 transcripts (2–7). In
spite of the marked improvement achieved by previous studies, a
systematic study analyzing and comparing the influence of different
mutations on PIEZO2 transcription, translation and protein function
is currently lacking, to the best of our knowledge. Therefore, the
present study aimed to systematically evaluate the effect of
different pathological mutations of PIEZO2 on its transcription, as
well as on translation and protein structure/function that
contribute to DA3, DA5, MWS and associated diseases based on a
bioinformatics analysis.
Materials and methods
Acquisition of pathological mutation
information of PIEZO2
The pathological mutation information for PIEZO2 was
obtained from the ClinVar database (https://www.ncbi.nlm.nih.gov/clinvar/), and further
information was yielded from published studies found on PubMed. A
number of studies that reported exact mutations in PIEZO2 (1–8) were
included in the present study. However, certain studies that
reported pathological deletions or repetitions in the chromosome
region that included PIEZO2, but did not exactly confirm whether
PIEZO2 was the pathological gene were excluded from the present
study (19–21).
Conservation analysis of mutation
sites in PIEZO2
The amino acid sequences of human PIEZO2 protein, as
well as Monopterus albus, Mus musculus, Odocoileus virginianus
texanus, Pogona vitticeps and Seriola dumerili PIEZO2
protein, were obtained from the National Center for Biotechnology
Information (NCBI) website (https://www.ncbi.nlm.nih.gov/protein/). The amino acid
sequence was saved in FASTA format, and the conservation of
mutation sites in PIEZO2 was compared between different species
using BioEdit software (version 7.0.5; downloaded from http://www.mbio.ncsu.edu/bioedit/bioedit.html).
Effect of gain-of-function mutations
of PIEZO2 on protein structure
In the RaptorX database (http://raptorx.uchicago.edu/StructurePropertyPred/predict/)
(22,23) and the Phyre2 database (http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)
(24), the amino acid sequences of
normal and mutant PIEZO proteins were entered and the protein
structures were predicted and analyzed.
Results
Reported mutation information of
PIEZO2
A total of 27 pathological mutations were obtained
from the ClinVar database and references, which included 6 de
novo mutations, 10 dominant mutations and 11 recessive
mutations. Among these mutations, gain-of-function mutations
(dominant/de novo mutations) were located in the middle and
C-terminal region, but mainly in the C-terminal region,
particularly in the 52nd exon region. Loss-of-function mutations
(recessive mutations) were located in the middle and N-terminal
region, but mainly in the N-terminal region (Table I).
 | Table I.Pathological mutation information of
PIEZO2. |
Table I.
Pathological mutation information of
PIEZO2.
| Author/year | Gene variation | Exon | Protein
variation |
Disease/phenotypes | Inheritance | (Refs.) |
|---|
| McMillin MJ, et
al 2014 |
NM_022068.3:c.8238_8245 delGACTAGAG | 52 |
p.Trp2746Terfsa | GS | De novo | (5) |
| McMillin MJ, et
al 2014 |
NM_022068.3:c.8215T>C | 52 | p.Ser2739Pro | DA5 | Dominant | (5) |
| McMillin MJ, et
al 2014 |
NM_022068.3:c.8208delA | 52 | p.Tyr2737Ilefs | DA5 | De novo | (5) |
| McMillin MJ, et
al 2014 | NM_022068.3:c.
8181_8183delAGA | 52 | p.Glu2727del | DA5 | Dominant/de
novo | (5) |
| Coste B, et al
2013 |
NM_022068.2:c.8179_8181del | 52 | p.Glu2727del | DA5 | Dominant | (6) |
| McMillin MJ, et
al 2014 |
NM_022068.3:c.8153G>C | 52 | p.Arg2718Pro | DA5 | De novo | (5) |
| McMillin MJ, et
al 2014 |
NM_022068.3:c.8153G>T | 52 | p.Arg2718Leu | DA5 | Dominant | (5) |
| McMillin MJ, et
al 2014; Alisch F, et al |
NM_022068.3:c.8057G>A | 52 | p.Arg2686His | DA5, GS | Dominant/de
novo | (2,5) |
| McMillin MJ, et
al 2014 |
NM_022068.3:c.8056C>T | 52 | p.Arg2686Cys | MWS | De novo | (5) |
| McMillin MJ, et
al 2014 |
NM_022068.3:c.7067C>T | 45 | p.Thr2356Met | DA5 | Dominant | (5) |
| McMillin MJ, et
al 2014 |
NM_022068.3:c.6668C>T | 45 | p.Ser2223Leu | DA5 | De novo | (5) |
| McMillin MJ, et
al 2014 |
NM_022068.3:c.6662C>T | 43 | p.Thr2221Ile | DA5 | Dominant/de
novo | (5) |
| a |
NM_022068.3:c.5895G>A | 38 | p.Trp1965Ter | b | Recessive |
|
| Delle Vedove A,
et al 2016 |
NM_022068.3:c.5621delT | 37 | p.Leu1874Argfs | c | Recessive | (3) |
| Chesler AT, et
al 2016 |
NM_022068.3:c.5054G>C | 35 | p.Arg1685Pro | d | Recessive | (7) |
| Chesler AT, et
al 2016 |
NM_022068.3:c.5053C>T | 35 | p.R1685* | d | Recessive | (7) |
| Chesler AT, et
al 2016 |
NM_022068.3:c.4723C>T | 32 | p.R1575* | d | Recessive | (7) |
| Okubo M, et
al. 2015 |
NM_022068.c.4456G>C | 30 | p.Ala1486Pro | DA5 | Dominant | (4) |
| Delle Vedove A,
et al 2016 |
NM_022068.3:c.3020_3030del
CTGAGAACTTC | 20 | p.Pro1007Leufs | c | Recessive | (3) |
| Delle Vedove A,
et al 2016 | NM_022068.
c.3019_3029del | 20 |
p.Pro1007Leufs*3 | c | Recessive | (3) |
| McMillin MJ, et
al 2014 |
NM_022068.3:c.2993T>C | 20 | p.Met998Thr | DA5 | De novo | (5) |
| Mahmud AA, et
al 2017 |
NM_022068.3:c.2708C>G | 18 | p.Ser903Ter | e | Recessive | (8) |
| Coste B, et
al 2013 |
NM_022068.3:c.2404A>T | 17 | p.Ile802Phe | DA5 | Dominant | (6) |
| McMillin MJ, et
al 2014 |
NM_022068.3:c.2134A>G | 15 | p.Met712Val | DA5 | Dominant/de
novo | (5) |
| Delle Vedove A,
et al 2016 |
NM_022068.3:c.1550_1552 delGCTinsCGAA | 13 | p.Ser517Thrfs | c | Recessive | (3) |
| Haliloglu G, et
al 2017 | NM_022068,
c.1384C>T | 9 | p.R462* | f | Recessive | (1) |
| Delle Vedove A,
et al 2016 | NM_022068,
c.493-?_917+del | 6,7 | NMD | c |
| (3) |
Conservation of mutation sites in
PIEZO2
The sequence blast results of the PIEZO2 protein
obtained from BioEdit software (version 7.0.5; downloaded from
http://www.mbio.ncsu.edu/bioedit/bioedit.html)
indicated that most mutation sites exhibited high conservation
among different species, particularly in the C-terminal region. For
mammals, all mutation sites exhibited high conservation in Homo
sapiens, Mus musculus and Odocoileus virginianus
texanus. However, the p.R462* mutation was distinctive between
humans and other species (Table
II).
| Table II.Conservation of mutant sites of
PIEZO2 in different species. |
Table II.
Conservation of mutant sites of
PIEZO2 in different species.
|
| Amino acid in
different species |
|---|
|
|
|
|---|
| Mutant site | Homo
sapiens | Monopterus
albus | Mus
musculus | Odocoileus
virginianus texanus | Pogona
vitticeps | Seriola
dumerili |
|---|
| p.Trp2746Terfs | W | W | W | W | W | W |
| p.Ser2739Pro | S | S | S | S | S | S |
| p.Tyr2737Ilefs | Y | Y | Y | Y | Y | Y |
| p.Glu2727del | E | E | E | E | E | E |
| p.Arg2718Pro | R | R | R | R | R | R |
| p.Arg2718Leu | R | R | R | R | R | R |
| p.Arg2686His | R | R | R | R | R | R |
| p.Arg2686Cys | R | R | R | R | R | R |
| p.Thr2356Met | T | T | T | T | T | T |
| p.Ser2223Leu | S | S | S | S | S | S |
| p.Thr2221Ile | T | T | T | T | Aa | T |
| p.Trp1965Ter | W | W | W | W | W | W |
| p.Leu1874Argfs | L | Ma | L | L | L | Ma |
| p.Arg1685Pro | R | R | R | R | R | R |
| p.Arg1685Ter | R | R | R | R | R | R |
| p.Arg1575Ter | R | R | R | R | R | Ka |
| p.Ala1486Pro | A | Va | A | A | A | Va |
| p.Pro1007Leufs | P | P | P | P | P | P |
| p.Met998Thr | M | M | M | M | M | M |
| p.Ser903Ter | S | S | S | S | S | S |
| p.Ile802Phe | I | Va | I | I | I | I |
| p.Met712Val | M | M | M | M | M | M |
| p.Ser517Thrfs | S | S | S | S | S | S |
| p.R462* | Ra | K | K | K | K | K |
Effect of gain-of-function mutations
of PIEZO2 on protein structure and function
Regarding recessive mutations of PIEZO2, a previous
study has indicated that they lead to loss-of-function of PIEZO2
due to nonsense-mediated decay of PIEZO2 transcripts or termination
of protein synthesis through premature termination codons (3).
Regarding gain-of-function mutations, including
E2727del and I802F, a previous study indicated that these two
abovementioned mutations of PIEZO2 facilitate faster recovery of
mechanically activated currents from inactivation, with E2727del
leading to a slowing of inactivation, resulting in increased
channel activity in response to mechanical stimulus (6). However, how these mutations influence
the structure of PIEZO2 protein to affect the protein function
remains to be elucidated. In the present study, the bioinformatical
analysis results indicated that these mutations, including
p.Ala1486Pro, p.Thr2221Ile and p.Glu2727del, modify the α-helix
structure, whereas p.Thr2356Met would modify the α-helix and
transmembrane helix structure (Fig.
1). In addition, the p.Thr2221Ile, p.Arg2718Leu and
p.Arg2718Pro mutations lead to a reduction of the solvent
accessibility of the PIEZO2 protein, whereas p.Arg2718Pro may
change the α-helix to a loop structure for the 8-state secondary
structure (SS8), and modifies the α-helix to a coil for SS3
(Fig. 2).
 | Figure 2.Prediction of the structural
properties of human PIEZO2 protein through the RaptorX online
database. In each pair of columns, the native site of the PIEZO2
protein is stated on the left and the corresponding mutant site on
the right. The capital letters in the code correspond to the amino
acids. Code in the different rows: SS3 (H, α-helix; E, β-sheet; C,
coil); SS8 [G, 3-helix; I, 5-helix (π-helix); E, extended strand in
β-ladder; B, isolated β-bridge; T, hydrogen-bonded turn; S,
hydrogen-bonded bend; L, hydrogen-bonded loop]; ACC [B, buried (ACC
<10%); M, Medium, 10%<ACC<40%; E, Exposed, ACC >40%].
SEQ, sequence; SS3, 3-state secondary structure; ACC, solvent
accessibility; DISO, disorder (prediction is based on the cutoff
value at 0.25); PIEZO2, Piezo type mechanosensitive ion channel
component 2. |
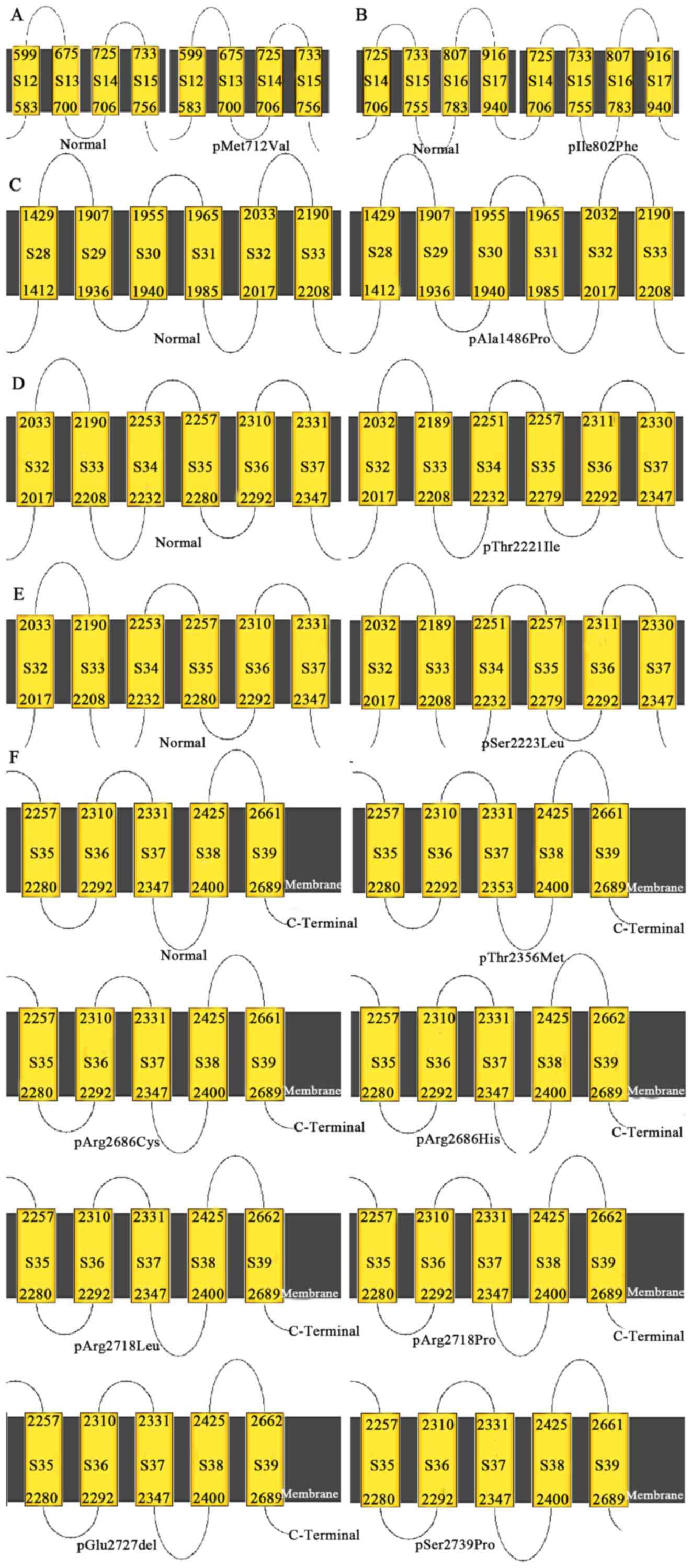
The p.Ala1486Pro mutation changes the 32nd
transmembrane region (S32) of PIEZO2 protein, while the
p.Thr2221Ile and p.Ser2223Leu mutations influence S32-S37 of the
PIEZO2 protein. p.Thr2356Met changes S37. p.Arg2686His,
p.Arg2718Leu, p.Arg2718Pro and p.Glu2727del modify S39 (Fig. 3).
Discussion
Proprioception is the perception of body and limb
position mediated by proprioceptors, namely innervate muscle
spindles and Golgi tendon organs, two types of mechanoreceptor in
skeletal muscles. Mechanotransduction is important for sensory
perception and embryonic development of organs and tissues
(9,10). PIEZO2 is a highly conserved
non-selective and mechanosensitive cation channel (25). In line with this, the results of the
present study demonstrated that these reported mutation sites are
highly conserved. PIEZO2 mutations have been reported to be linked
with arthrogryposis, including GS/DA3, DA5, MWS and other
associated diseases (2,3,5,6). However, the clinical manifestations of
the PIEZO2-associated diseases display a great variation.
Heterozygous gain-of-function mutations in PIEZO2 are of
autosomal-dominant inheritance and contribute to GS/DA3, DA5 and
MWS (6). In addition, numerous
recessively inherited PIEZO2-associated diseases included
arthrogryposis and other symptoms that overlap with GS/DA3, DA5 and
MWS, but which may not be diagnosed as GS/DA3, DA5 or MWS due to
certain distinct clinical manifestations (1,3).
Dominant gain-of-function mutations are mainly localized at the
C-terminal end of PIEZO2. Recessive loss-of-function mutations are
distributed across the PIEZO2 protein but mainly in the N-terminal
region, and do not map to any hotspots as is the case for dominant
gain-of-function mutations. The results of the present study also
confirm this.
A previous study indicated that gain-of-function
mutations in PIEZO2 lead to a deceleration of PIZO2 channel
inactivation and/or faster recovery from inactivation, resulting in
increased channel activity (6). This
may be due to these mutations being mainly located in the last
several exons of PIEZO2 and thus, transcripts carrying these
mutations are expected to escape nonsense-mediated transcript decay
or premature termination codons, while mainly influencing the
protein structure and function. By contrast, loss-of-function
mutations may result in nonsense-mediated transcript decay or
premature termination codons, consequently leading to a lack of
PIEZO2 protein (1,3). The presence of distinct clinical
phenotypes between or even among the gain- and loss-of-function
mutations of PIEZO2 linked with PIEZO2-associated diseases implies
that different mutations in PIEZO2 have different effects on
disease pathophysiology, which may depend on the type of mutation
and the mode of inheritance.
Although the structure of PIEZO2 protein remains to
be determined, the high-resolution cryo-electron microscopy
structure of murine PIEZO1 has been published (26), which offers useful information for
the prediction of the structure and function of PIEZO2 protein. A
previous study speculated that the C-terminal region of PIEZO2 and
PIEZO1 has regulatory roles for this ion channel, which may offer
certain clues regarding gain-of-function mutations (27). In the present study, the
bioinformatical analysis revealed that the p.Ala1486Pro,
p.Thr2221Ile and p.Glu2727del mutations modify the secondary
structure of PIEZO2 protein. Furthermore, the p.Thr2221Ile,
p.Arg2718Leu and p.Arg2718Pro mutations reduce the solvent
accessibility of PIEZO2 protein. The p.Ala1486Pro, p.Thr2221Ile,
p.Ser2223Leu, p.Thr2356Met, p.Arg2686His, p.Arg2718Leu,
p.Arg2718Pro and p.Glu2727del mutations affect the transmembrane
region of PIEZO2. These mutations may change the activity of PIEZO2
to enhance its function. In a previous study, similar
characteristics were reported for other ion channels (28).
In summary, the results of the present study further
confirm that dominant and recessive mutations were present in
PIEZO2, with dominant mutations being mainly located in the
C-terminal region, whereas recessive mutations were mainly located
in the N-terminal region. No overlap was present between these
hotspots, and most reported mutation sites exhibited high
conservation in different species, particularly in the C-terminal
region. Loss-of-function mutations may result in nonsense-mediated
transcript decay or premature termination codons, consequently
leading to a lack of PIEZO2 protein, whereas gain-of-function
mutations of PIEZO2 lead to a slowing of PIZO2 channel inactivation
and/or faster recovery from inactivation, resulting in increased
channel activity. The bioinformatical analysis also suggested that
the p.Ala1486Pro, p.Thr2221Ile and p.Glu2727del mutations modify
the secondary structure of PIEZO2 protein, while the p.Thr2221Ile,
p.Arg2718Leu and p.Arg2718Pro mutations reduce the solvent
accessibility of PIEZO2 protein. In addition, the p.Ala1486Pro,
p.Thr2221Ile, p.Ser2223Leu, p.Thr2356Met, p.Arg2686His,
p.Arg2718Leu, p.Arg2718Pro and p.Glu2727del mutations affect the
transmembrane region of PIEZO2. These mutations may change the
activity of PIEZO2 that may contribute to an enhanced function of
PIEZO2. Variable clinical phenotypes were present between and among
the gain- and loss-of-function mutations linked with
PIEZO2-associated disease, which implied that different mutations
in PIEZO2 have different pathophysiological effects. Of course,
further functional, electrophysiological and high-resolution
cryo-electron microscopy studies are required to explore the
precise structure and function of PIEZO2, which may offer useful
clues for the prevention and treatment of PIEZO2-associated
disease.
Acknowledgements
Not applicable.
Funding
No funding received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YM analyzed the data and wrote the paper. YZ and ZC
analyzed the data. XH designed the study and revised the paper. The
final version of the manuscript has been read and approved by all
authors, and each author believes that the manuscript represents
honest work.
Ethical approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Haliloglu G, Becker K, Temucin C, Talim B,
Küçükşahin N, Pergande M, Motameny S, Nürnberg P, Aydingoz U,
Topaloglu H and Cirak S: Recessive PIEZO2 stop mutation causes
distal arthrogryposis with distal muscle weakness, scoliosis and
proprioception defects. J Hum Genet. 62:497–501. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Alisch F, Weichert A, Kalache K, Paradiso
V, Longardt AC, Dame C, Hoffmann K and Horn D: Familial Gordon
syndrome associated with a PIEZO2 mutation. Am J Med Genet A.
173:254–259. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Delle Vedove A, Storbeck M, Heller R,
Hölker I, Hebbar M, Shukla A, Magnusson O, Cirak S, Girisha KM,
O'Driscoll M, et al: Biallelic loss of proprioception-related
PIEZO2 causes muscular atrophy with perinatal respiratory distress,
arthrogryposis, and scoliosis. Am J Hum Genet. 99:1206–1216. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Okubo M, Fujita A, Saito Y, Komaki H,
Ishiyama A, Takeshita E, Kojima E, Koichihara R, Saito T, Nakagawa
E, et al: A family of distal arthrogryposis type 5 due to a novel
PIEZO2 mutation. Am J Med Genet A 167A. 1100–1106. 2015. View Article : Google Scholar
|
|
5
|
McMillin MJ, Beck AE, Chong JX, Shively
KM, Buckingham KJ, Gildersleeve HI, Aracena MI, Aylsworth AS,
Bitoun P, Carey JC, et al: Mutations in PIEZO2 cause Gordon
syndrome, Marden-Walker syndrome, and distal arthrogryposis type 5.
Am J Hum Genet. 94:734–744. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Coste B, Houge G, Murray MF, Stitziel N,
Bandell M, Giovanni MA, Philippakis A, Hoischen A, Riemer G, Steen
U, et al: Gain-of-function mutations in the mechanically activated
ion channel PIEZO2 cause a subtype of Distal Arthrogryposis. Proc
Natl Acad Sci USA. 110:4667–4672. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chesler AT, Szczot M, Bharucha-Goebel D,
Čeko M, Donkervoort S, Laubacher C, Hayes LH, Alter K, Zampieri C,
Stanley C, et al: The role of PIEZO2 in human mechanosensation. N
Engl J Med. 375:1355–1364. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mahmud AA, Nahid NA, Nassif C, Sayeed MS,
Ahmed MU, Parveen M, Khalil MI, Islam MM, Nahar Z, Rypens F, et al:
Loss of the proprioception and touch sensation channel PIEZO2 in
siblings with a progressive form of contractures. Clin Genet.
91:470–475. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Felsenthal N and Zelzer E: Mechanical
regulation of musculoskeletal system development. Development.
144:4271–4283. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu J, Lewis AH and Grandl J: Touch,
tension, and transduction - the function and regulation of piezo
ion channels. Trends Biochem Sci. 42:57–71. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee W, Leddy HA, Chen Y, Lee SH, Zelenski
NA, McNulty AL, Wu J, Beicker KN, Coles J, Zauscher S, et al:
Synergy between Piezo1 and Piezo2 channels confers high-strain
mechanosensitivity to articular cartilage. Proc Natl Acad Sci USA.
111:E5114–E5122. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Coste B, Mathur J, Schmidt M, Earley TJ,
Ranade S, Petrus MJ, Dubin AE and Patapoutian A: Piezo1 and Piezo2
are essential components of distinct mechanically activated cation
channels. Science. 330:55–60. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nonomura K, Woo SH, Chang RB, Gillich A,
Qiu Z, Francisco AG, Ranade SS, Liberles SD and Patapoutian A:
Piezo2 senses airway stretch and mediates lung inflation-induced
apnoea. Nature. 541:176–181. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li J, Hou B, Tumova S, Muraki K, Bruns A,
Ludlow MJ, Sedo A, Hyman AJ, McKeown L, Young RS, et al: Piezo1
integration of vascular architecture with physiological force.
Nature. 515:279–282. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cahalan SM, Lukacs V, Ranade SS, Chien S,
Bandell M and Patapoutian A: Piezo1 links mechanical forces to red
blood cell volume. eLife. 4:2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Anderson EO, Schneider ER and Bagriantsev
SN: Piezo2 in cutaneous and proprioceptive mechanotransduction in
vertebrates. Curr Top Membr. 79:197–217. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ranade SS, Woo SH, Dubin AE, Moshourab RA,
Wetzel C, Petrus M, Mathur J, Bégay V, Coste B, Mainquist J, et al:
Piezo2 is the major transducer of mechanical forces for touch
sensation in mice. Nature. 516:121–125. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Woo SH, Ranade S, Weyer AD, Dubin AE, Baba
Y, Qiu Z, Petrus M, Miyamoto T, Reddy K, Lumpkin EA, et al: Piezo2
is required for Merkel-cell mechanotransduction. Nature.
509:622–626. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lemmers RJ, van den Boogaard ML, van der
Vliet PJ, Donlin-Smith CM, Nations SP, Ruivenkamp CA, Heard P,
Bakker B, Tapscott S, Cody JD, et al: Hemizygosity for SMCHD1 in
facioscapulohumeral muscular dystrophy type 2: Consequences for 18p
deletion syndrome. Hum Mutat. 36:679–683. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Carter E, Heard P, Hasi M, Soileau B,
Sebold C, Hale DE and Cody JD: Ring 18 molecular assessment and
clinical consequences. Am J Med Genet A 167A. 54–63. 2015.
View Article : Google Scholar
|
|
21
|
Miller DT, Adam MP, Aradhya S, Biesecker
LG, Brothman AR, Carter NP, Church DM, Crolla JA, Eichler EE,
Epstein CJ, et al: Consensus statement: Chromosomal microarray is a
first-tier clinical diagnostic test for individuals with
developmental disabilities or congenital anomalies. Am J Hum Genet.
86:749–764. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang S, Li W, Liu S and Xu J:
RaptorX-Property: A web server for protein structure property
prediction. Nucleic Acids Res. 44:W430–W435. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Källberg M, Wang H, Wang S, Peng J, Wang
Z, Lu H and Xu J: Template-based protein structure modeling using
the RaptorX web server. Nat Protoc. 7:1511–1522. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kelley LA, Mezulis S, Yates CM, Wass MN
and Sternberg MJ: The Phyre2 web portal for protein modeling,
prediction and analysis. Nat Protoc. 10:845–858. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vasquez V, Scherrer G and Goodman MB:
Sensory biology: It takes Piezo2 to tango. Curr Biol. 24:566–R569.
2014. View Article : Google Scholar
|
|
26
|
Saotome K, Murthy SE, Kefauver JM, Whitwam
T, Patapoutian A and Ward AB: Structure of the mechanically
activated ion channel Piezo1. Nature. 554:481–486. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu J, Young M, Lewis AH, Martfeld AN,
Kalmeta B and Grandl J: Inactivation of mechanically activated
piezo1 ion channels is determined by the C-terminal extracellular
domain and the inner pore helix. Cell Rep. 21:2357–2366. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Held K, Gruss F, Aloi VD, Janssens A,
Ulens C, Voets T and Vriens J: Mutations in the voltage-sensing
domain affect the alternative ion permeation pathway in the TRPM3
channel. J Physiol. 2018. View
Article : Google Scholar : PubMed/NCBI
|