Introduction
Colon cancer is one of the most common malignancies
severely threatening human health (1). It ranks third highest worldwide in
terms of cancer-associated mortality rates (1). According to statistics, there were
~1,000,000 new cases of colon cancer in 2008 worldwide, and
~500,000 individuals succumbed to mortality (2). The morbidity rate of colon cancer in
China has exhibited an increasing trend in recent years. This can
be ascribed to improvements in nationwide living standards, changes
in dietary patterns, aging of the population, and rapid
developments in endoscopy (3,4).
Therefore, colon cancer has become a serious threat to human health
asone of the most common malignancies (3,4).
Colon cancer has a high mortality rate regardless of
the advances in surgical technology and other treatments and
despite novel molecular preparations emerging in previous years
(5). This may be attributed mainly
to the frequent recurrence and metastasis of colon cancer (5). Therefore, early diagnosis and treatment
is of significance for improving prognosis of patients with colon
cancer (5). At present, the
mechanism underlying its tumorigenesis remains to be fully
elucidated. Understanding its corresponding mechanism is important
to develop novel strategies and identify novel target genes for
effectively treating colon cancer (6).
MicroRNAs are types of small non-coding RNA molecule
of ~19–24 nucleotides in length (7).
They are relatively conserved during biological evolution and do
not encode protein. However, they can regulate target genes through
directly binding with the 3′untranslated region (3′-UTR) of target
mRNA (8).
It has been demonstrated in previous years that
miRNAs are closely associated with tumor genesis and development.
Compared with normal tissue, the miRNA expression profile in tumor
tissue is significantly altered. Furthermore, miRNAs interact with
traditional tumor suppressor genes and oncogenes (9). Therefore, they have dual-identity
during tumorigenesis and development. It is estimated that >50%
of miRNA upstream regulatory genes locate in tumor-associated
regions in chromosomes. There are a variety of miRNAs, including
those functioning as oncogenes and others as tumor suppressor
genes. miRNAs can exert oncogenic functions, inhibit the expression
of tumor suppressor genes, and provide favorable conditions for
tumor proliferation, migration, metastasis and infiltration
(10). By contrast, they can
function as a tumor suppressor gene, inhibit expression of
oncogene, and contribute to tumor cell apoptosis, differentiation
and treatment (11). The earliest
report regarding the correlation between miRNA and tumors was of
the 13q14 gene deletion in chronic lymphocytic leukemia (11). The oncogene miRNA17-92′ was the first
miRNA identified in mammals (11);
it shows abnormally increased expression in lung cancer and
lymphoma. Mutation in relevant miRNAs can activate the expression
of its oncogene or induce loss or changes in a tumor suppressor
gene, which results in tumor genesis and development (11).
MDM4 is a type of proto-oncogene locating in
chromosome 1q23, which encodes 11 exons and 490 amino acids
(8). The MDM4 protein structure
includes a p53-binding domain (p53 BD) locating in the N-terminal,
and an amino acid region locating in the RING figure domain and
center (8). The RING finger domain
is the major determinant of MDM4 and MDM2 acting on p53 (8), and a heterodimer can be formed through
the domain (8). MDM4 binds with p53
through p53BD in its N-terminal, and inhibits the transcriptional
activity of p53. It has been verified in a previous study that MDM4
is partly correlated with tumors (12). For example, the overexpression of
MDM4 can be detected in certain human tumor cell lines, and MDM4
proliferation has be found in solid tumors, including breast cancer
and colon cancer (7,12). Therefore, it is hypothesized that
MDM4 in human tumors can promote tumor genesis or progression. The
present study investigated whether the role of the expression of
miRNA-766 affects human colon cancer survival rate and examined the
underlying molecular mechanisms.
Materials and methods
Study population
A total of 102 patients with human colon cancer and
57 normal volunteers were recruited from the Second Affiliated
Hospital, Shantou University Medical College (Shantou, China) from
March 2012 to July 2012. Colon cancer tissue samples were acquired
in patients undergoing surgery, and para-carcinoma tissues were
collected at a distance of 5 cm from the cancer tissue samples. The
present study was approved by the Ethical Agent Will of the Second
Affiliated Hospital, Shantou University Medical College.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was isolated using a TRIzol-based
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) RNA
isolation protocol and 500 ng RNA was reverse transcribed using the
TaqMan microRNA Reverse Transcription kit (Applied Biosystems;
Thermo Fisher Scientific, Inc.). RT-qPCR analysis was performed
using SYBR-Green PCR Master Mix (PE Applied Biosystems; Thermo
Fisher Scientific, Inc.) with the Applied Biosystems 7900HT
real-time PCR machine (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The reaction conditions were as follows: Hot
start at 94°C for 10 min; 40 cycles of 30 sec at 95°C, 30 sec at
60°C and 30 sec at 72°C, and 10 min at 72°C. miR-766 forward,
5′-TCGAGTACTTGAGATGGAGTTTT-3′ and reverse,
5′-GGCCGCGTTGCAGTGAGCCGAG-3′; U6 forward,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′. miRNA expression was measured using
2−∆∆cq (13).
Microarray analysis
RNA cleanup was performed using the RNeasy Mini kit
(Qiagen, Inc., Valencia, CA, USA). Total RNA was reverse
transcribed into cDNA using an oligo d(T) (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). cRNA was hybridized onto the Affymetrix
Human Genome U133 Plus 2.0 Array, staining was performed with a
Fluidic Station-450 and GeneChips were scanned with the Affymetrix
GeneChip Scanner 7G. Data were quantified and feature-extracted
using Agilent Feature Extraction software (version A.10.7.3.1;
Agilent Technologies, Inc., Santa Clara, CA, USA).
Cell culture and cell transfection
experiments
Human Caco2 cells were purchased from Type Culture
Collection of the Chinese Academy of Sciences (Shanghai, China) and
cultured in Dulbecco's minimal essential medium (DMEM; GE
Healthcare Life Sciences, Logan, UT, USA), 10% fetal bovine serum
(FBS; GE Healthcare Life Sciences) at 37°C in 5% CO2 and 95%
relative humidity. miRNA (miR)-766 mimics and negative control
mimics were purchased from Sangon Biotech Co., Ltd. (Shanghai,
China). Subsequently, 20 ng/ml miR-766, anti-miR-766 mimics and
negative control mimics were transfected into Caco2 cells using
Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol.
Cell viability assay and analysis of
apoptosis
The cells (1×106 cell/ml) were incubated
with medium containing MTT (5 mg/ml) for 4 h and dissolved with 150
µl DMSO. The medium was removed and dissolved with 150 µl DMSO for
15 min at room temperature. The absorbance was measured at 490 nm
using a microplate reader.
To analyze apoptosis, the cells were washed twice
with ice-cold PBS and resuspended in 500 µl binding buffer (Nanjing
KeyGen Biotech Co., Ltd., Nanjing, China). Subsequently, 5 µl
annexin V-fluorescein isothiocyanate and 5 µl propidium iodide
(both Nanjing KeyGen Biotech Co., Ltd.) were added and the cells
were incubated for 15 min at room temperature in the dark.
Apoptosis was analyzed using a FACSort flow cytometer and
quantified using BD CellQuest™ Pro software (BD Biosciences,
Franklin Lakes, NJ, USA).
Cell migration assay
The Caco2 cells (1×105 cell/ml) were
seeded on 24-well plates and were added to the upper chamber of
each migration well (Corning Corporation, Corning, NY, USA). DMEM
(500 µl) with 20% FBS was added to the lower chamber and incubated
for 48 h at 37°C. The lower side were fixed with 75% ice-alcohol
for 30 min and stained with 1% crystal violet solution for 1 h at
room temperature. The cells were counted under a fluorescence
microscope (Axio version II, Carl Zeiss AG, Oberkochen,
Germany).
Western blot analysis
Total protein was extracted in RIPA buffer (Beyotime
Institute of Biotechnology, Haimen, China) on ice for 30 min and
then centrifuged at 12,000 × g for 10 min at 4°C. Total protein was
quantified using a BCA assay (Beyotime Institute of Biotechnology)
and total protein samples (50 µg) were subjected to 8–12%
SDS-polyacrylamide gel electrophoresis and then transferred onto
PVDF membranes. The membranes were blocked with 5% skimmed milk in
TBST, and incubated at 4°C overnight with the following primary
antibodies: MDM4 (cat. no. sc-14738; 1:1,000; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), B-cell lymphoma 2-associated
X protein (Bax; cat. no. sc-6236, 1:1,000), p53 (cat. no. sc-47698;
1:1,000) and GAPDH (cat. no. sc-51631; 1:5,000; all Santa Cruz
Biotechnology). The membranes were washed with TBST for 15 min and
then incubated with the anti-rabbit horseradish peroxidase
secondary antibody (cat. no. sc-2004; 1:5,000; Santa Cruz
Biotechnology, Inc.) for 1 h at 37°C. The protein bands were
detected with enhanced chemiluminescence (Beyotime Institute of
Biotechnology).
Analysis of capase-3/9 activity
levels
Total protein was extracted using RIPA buffer
(Beyotime Institute of Biotechnology) on ice for 30 min and then
centrifuged at 12,000 × g for 10 min at 4°C. Total protein was
quantified using a BCA assay (Beyotime Institute of Biotechnology)
and total protein samples (10 µg) were used to measure the
capase-3/9 activity levels using capase-3/9 activity kits. The
absorbance was measured at 405 nm using a microplate reader.
Statistical analysis
The data are expressed as the mean ± standard
deviation. All statistical analyses were performed using SPSS
version 18.0 statistical software (SPSS, Inc., Chicago, IL, USA).
Statistical comparisons of >2 or three groups were performed
using Student's t-test or a one-way analysis of variance, followed
by a Bonferroni post-hoc test. P<0.05 was considered to indicate
a statistically significant difference.
Results
Expression of miRNA-766 in patients
with colon cancer
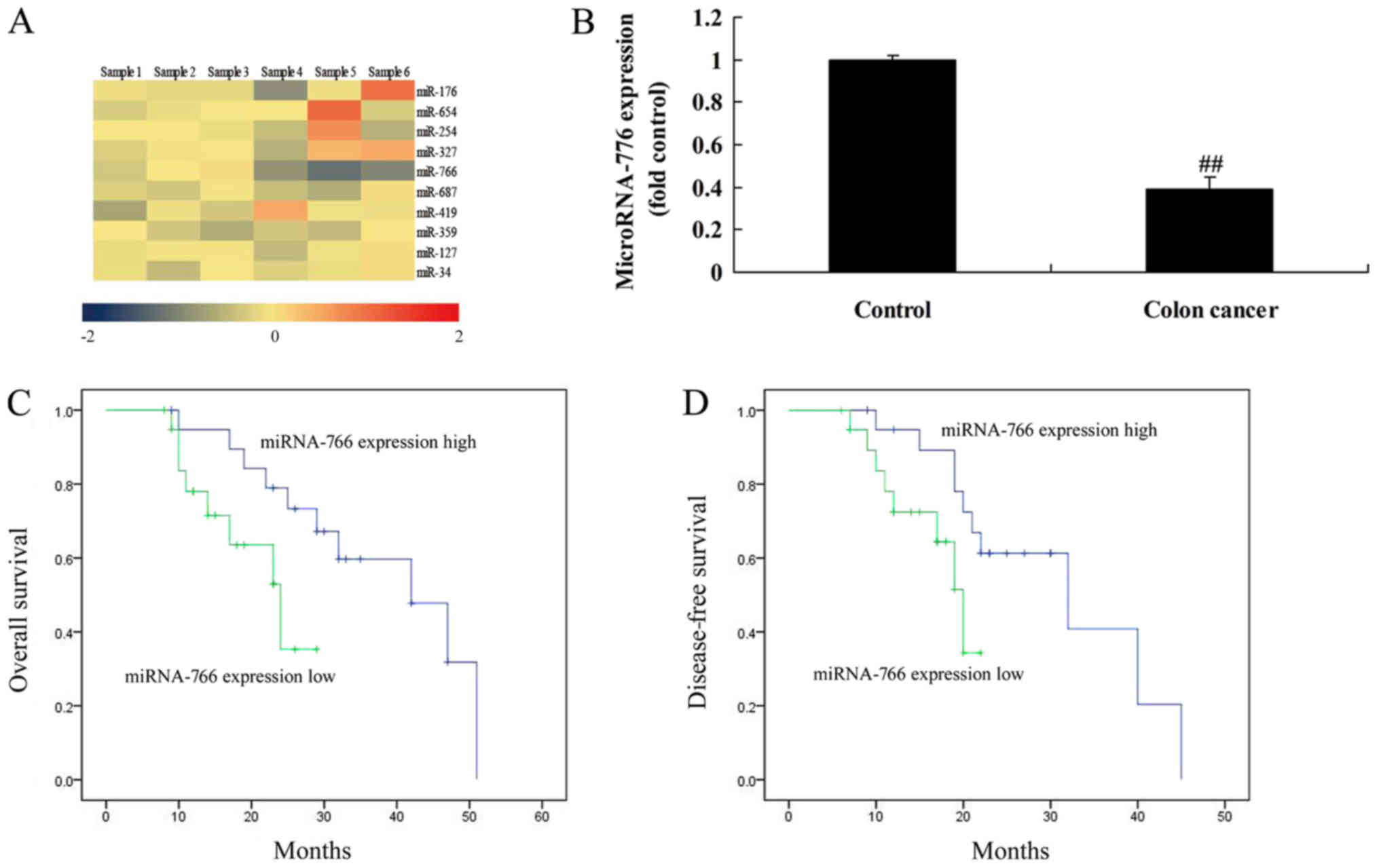
Firstly, the present study analyzed changes in the
expression of miRNAs in patients with colon cancer. As shown in
Fig. 1A and B, the expression of
miRNA-766 in patients with colon cancer was elevated, compared with
that in the control group. The overall survival (OS) and
disease-free survival (DFS) rates of patients with colon cancer
with a high expression of miRNA-766 were prolonged, compared with
those with a low expression of miRNA-766 (Fig. 1C and D). The baseline characteristics
of the 102 patients with colon cancer and 57 normal volunteers are
shown in Table I.
 | Table I.Distribution of selected
characteristics among patients with colon cancer and controls. |
Table I.
Distribution of selected
characteristics among patients with colon cancer and controls.
| Characteristic | Colon cancer, n
(%) | Control, n (%) | P-value |
|---|
| Number of
individuals | 102 | 57 |
|
| Age (years) |
|
|
|
|
<60 | 57 (55.88) | 31 (54.39) |
|
|
<60 | 45 (44.12) | 26 (45.61) |
|
| Sex |
|
|
|
| Male | 78 (76.47) | 46 (80.70) |
|
|
Female | 24 (23.53) | 11 (19.30) |
|
| Smoking status |
|
| 0.009 |
| No | 41 (40.20) | 31 (54.39) |
|
| Yes | 61 (59.80) | 26 (45.61) |
|
| Drinking status |
|
| <0.001 |
| No | 37 (36.27) | 33 (57.89) |
|
| Yes | 65 (63.73) | 24 (42.11) |
|
miRNA-766 regulates cell growth of
colon cancer cells
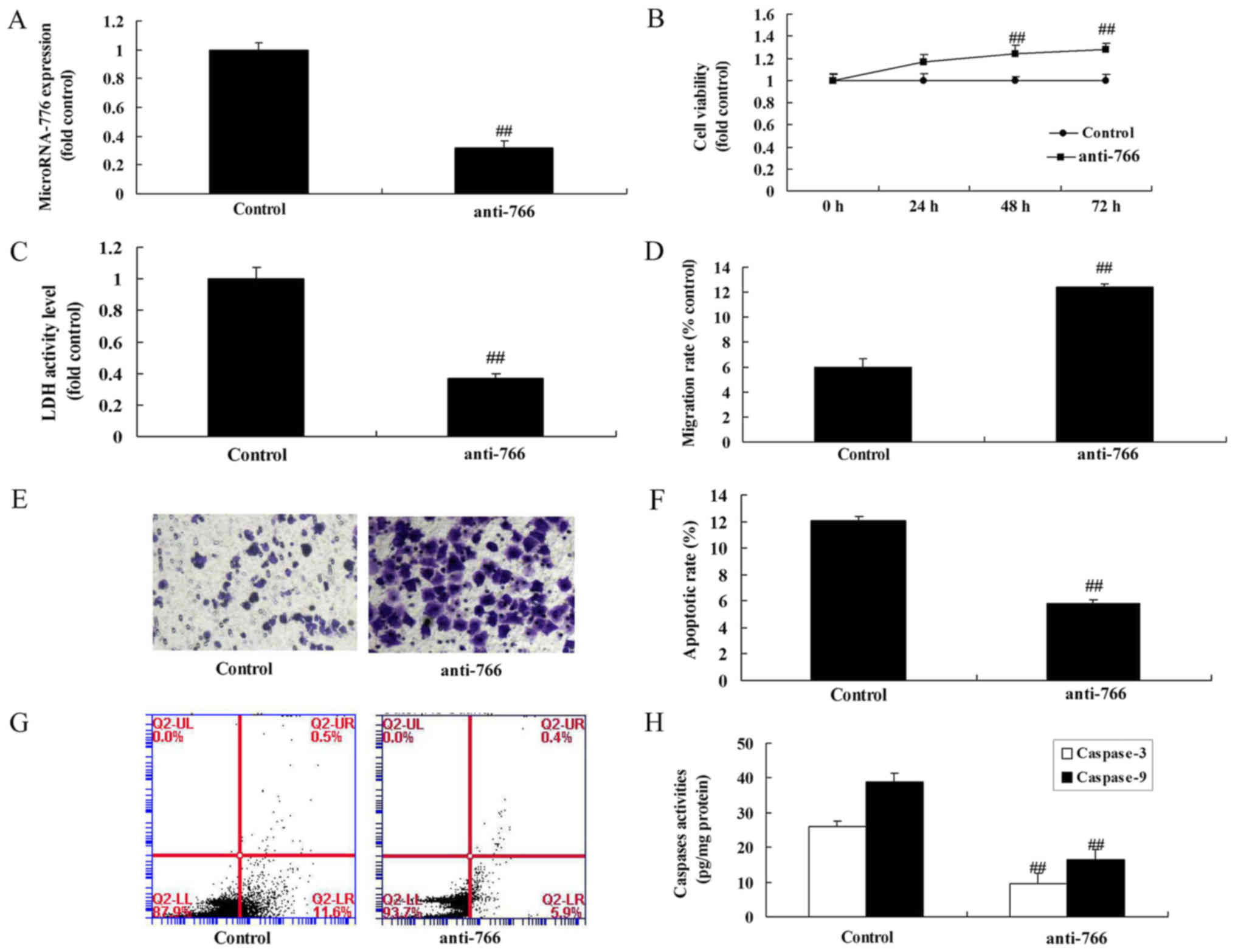
To examine the function of miRNA-766 in the cell
growth of colon cancer cells, the present study analyzed the
effects of miRNA-766 or anti-miRNA-766 on the growth of colon
cancer cells. As shown in Fig. 2A,
the expression of miRNA-766 was increased using miRNA-766 mimics in
Caco2 cells, compared with that in the negative control group. The
overexpression of miRNA-766 reduced cell growth and migration, and
promoted lactate dehydrogenase (LDH) activity, apoptotic rate and
caspase-3/9 activity levels in the Caco2 cells, compared with cells
in the negative control group (Fig.
2B-H). Subsequently, it was confirmed that the expression of
miRNA-766 was inhibited in Caco2 cells by using anti-miRNA-766
mimics, compared with expression in the negative control group
(Fig. 3A). The downregulation of
miRNA-766 promoted cell growth and migration, and reduced the LDH
activity, apoptotic rate and caspase-3/9 activity levels in Caco2
cells, compared with cells in the negative control group (Fig. 3B-H).
miRNA-766 regulates the MDM4/p53
pathway in colon cancer cells
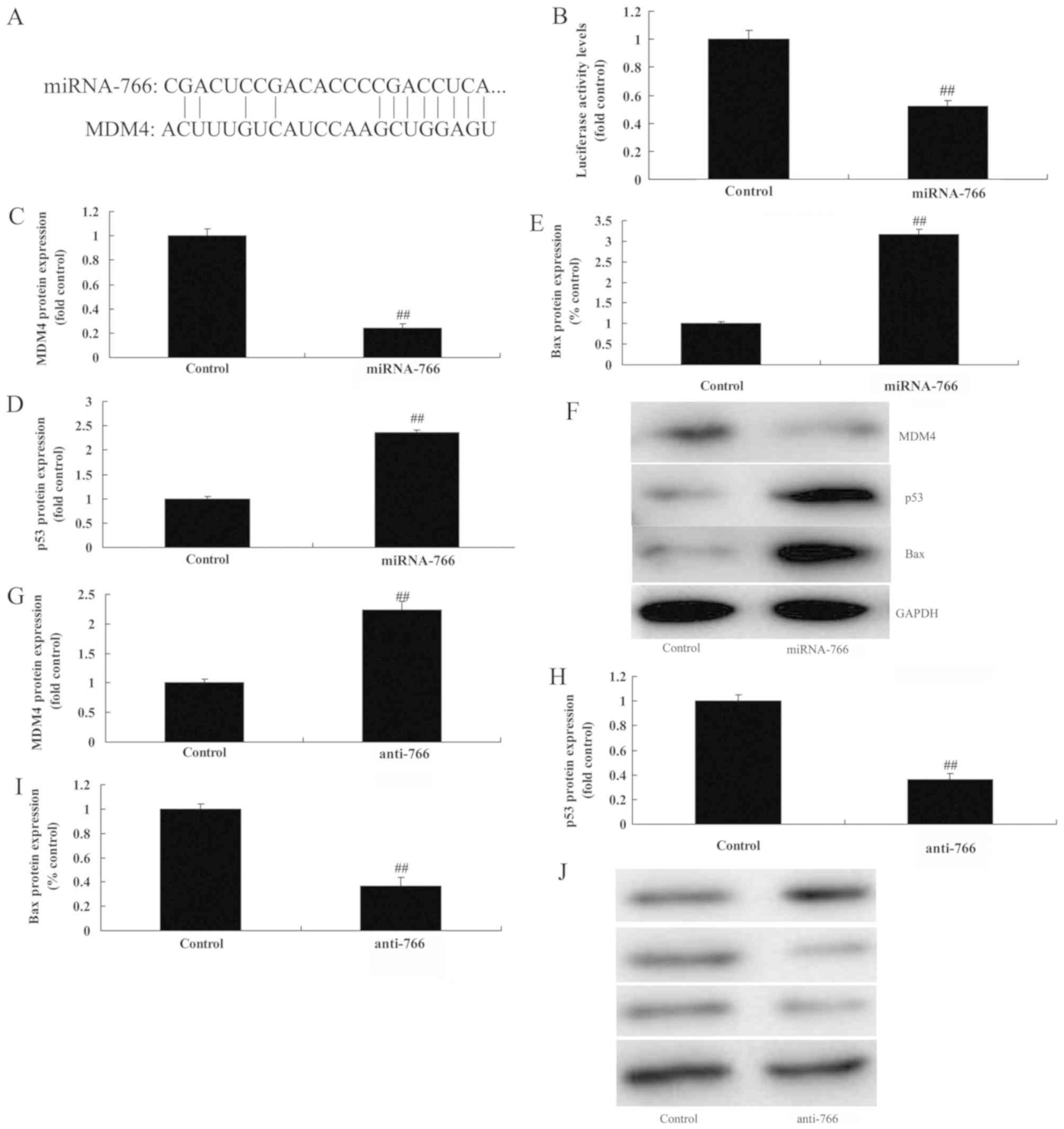
Subsequently, the present study examined the
mechanism underlying the effect of miRNA-766 on colon cancer cell
growth. As shown in Fig. 4A and B,
the putative miR-766-binding sequence in the 3′UTR of MDM4 mRNA and
luciferase activity was attenuated following the overexpression of
miRNA-766, compared with the negative control group. However, the
overexpression of miRNA-766 suppressed the protein expression of
MDM4, and induced that of p53 and Bax in the Caco2 cells, compared
with the negative control group (Fig.
4C-F). The downregulation of miRNA-766 induced the protein
expression of MDM4, and suppressed that of p53 and Bax in Caco2
cells, compared with the negative control group (Fig. 4G-J).
Promotion of MDM4 attenuates the
anticancer effect of miRNA-766 in colon cancer cells
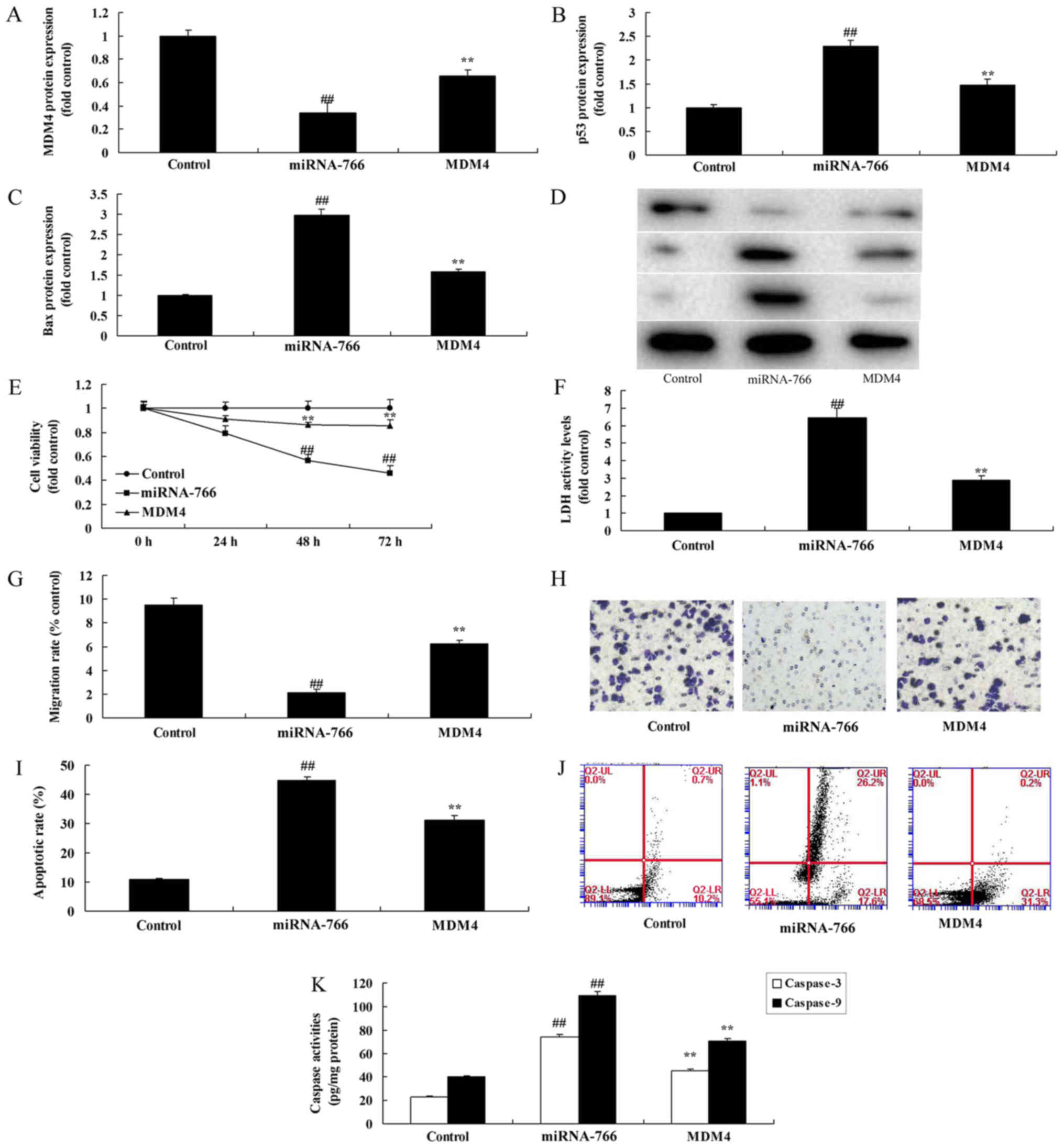
To further assess the relevance of the miR-766/MDM4
interaction in p53 signaling, an MDM4 plasmid was utilized to
induce the expression of MDM4 in Caco2 cells overexpressing
miR-766. As shown in Fig. 5A-D, the
MDM4 plasmid induced the protein expression of MDM4, and suppressed
that of p53 and Bax in Caco2 cells overexpressing miR-766, compared
with the cells overexpressing miR-766 without the plasmid. The
overexpression of MDM4 promoted cell growth and migration, and
reduced LDH activity, apoptotic rate and caspase-3/9 activity
levels in the Caco2 cells overexpressing miR-766, compared with the
negative control group (Fig.
5E-K).
Discussion
Colon cancer has a complicated pathogenic process
(14). It is controlled by multiple
genes, has different stages, and is formed over a long period
(14). Early diagnosis, together
with tumor recurrence monitoring and effective development of novel
treatments, is important for patients with colon cancer (13). It has been found in previous years
that several miRNAs exist in colon cancer tissues and patient
blood, and are important in the pathogenesis of colon cancer
(15). In the in vitro
experiments performed in the present study, the expression of
miRNA-766 in patients with colon cancer was increased, compared
with that in the control group. The OS and DFS rates of patients
with colon cancer and a high expression of miRNA-766 were higher
than those of patients with colon cancer and a low expression of
miRNA-766. Oh et al showed that miRNA-766 affected the
distant metastatic process to a greater extent than cancer cell
proliferation and primary tumor growth of human triple-negative
breast cancer cells (16). Jia et
al showed that miR-766-5p suppressed the tumor growth of
colorectal cancer (17). However,
the present study used only one cell line, Caco2 cells, which is
insufficient for this investigation.
miR-191, which locates in human 3p21.3, has been
found to be overexpressed in numerous types of human tumor
(18). For example, a high
expression of miR-191 has been found in liver, stomach, large
intestine, prostate, and breast cancer (19). It was found in liver cancer that
miRNA-191 promotes epithelial-mesenchymal transition and exerts its
tumor-promoting effect through suppressing the expression of TIM3;
it may serve as a novel target in the treatment of liver cancer
(15). It was confirmed in a study
on gastric cancer that miRNA-191 promotes gastric cancer cell
growth and inhibits cell apoptosis through its target gene NDST1
(15). In the present study, the
overexpression of miRNA-766 reduced cell growth and cell migration,
and promoted LDH activity, apoptotic rate and caspase-3/9 activity
levels in Caco2 cells.
Colon cancer is one of the most common malignancies
clinically. As indicated in numerous studies, tumorigenesis and
development is linked with disruption to the dynamic balance
between cell proliferation and apoptosis (19). Bcl-2 family proteins are important
regulatory factors of cell apoptosis, which can inhibit cell
apoptosis, for example Bcl-2 and Bcl-extra large, or promote
apoptosis, for example, Bax and BCL2-antagonist/killer. Changes in
expression not only affect DNA injury or normal apoptosis of cells
with abnormal cell cycle, but also affect the apoptosis of tumor
cells. The majority of antitumor drugs exert cytotoxic effects
through inducing tumor cell apoptosis (20). In the present study, the
overexpression of miRNA-766 suppressed the protein expression of
MDM4, and induced the protein expression of p53 and Bax in Caco2
cells.
Tumor suppressor p53 is important in regulating cell
cycle, apoptosis, DNA injury and aging (21). It is the gene that is most
susceptible to mutation in human tumors. It is reported that ~50%
of human tumors are associated with abnormalities in the p53 gene,
leading to p53 gene inactivation and abnormal p53 protein function.
Inhibition or inactivation of the p53 gene frequently promotes
tumorigenesis (22). Numerous
factors are involved in the activation of p53, including the MDM4
gene and MDM2 gene (23). MDM4 and
MDM2 are considered to be p53 inhibitors, which can regulate p53
activity (23). FL-MDM4 inhibits
p53-mediated transcriptional activity, and gives rise to cell cycle
arrest and apoptosis (24). MDM2
mainly mediates p53 degradation through E3 ubiquitin ligase. The
overexpression or proliferation of FL-MDM4 has been observed in
human solid tumors and tumor cell lines. The overexpression of
FL-MDM4 mRNA can be detected in colon cancer cells through RT-qPCR
analysis, as reported previously (24). In the present study, the promotion of
MDM4 reduced the anticancer effect of miRNA-766 in colon cancer
cells. Wang et al also demonstrated that miRNA-766 induced
p53 accumulation and G2/M arrest by directly targeting MDM4 in
breast cancer (25).
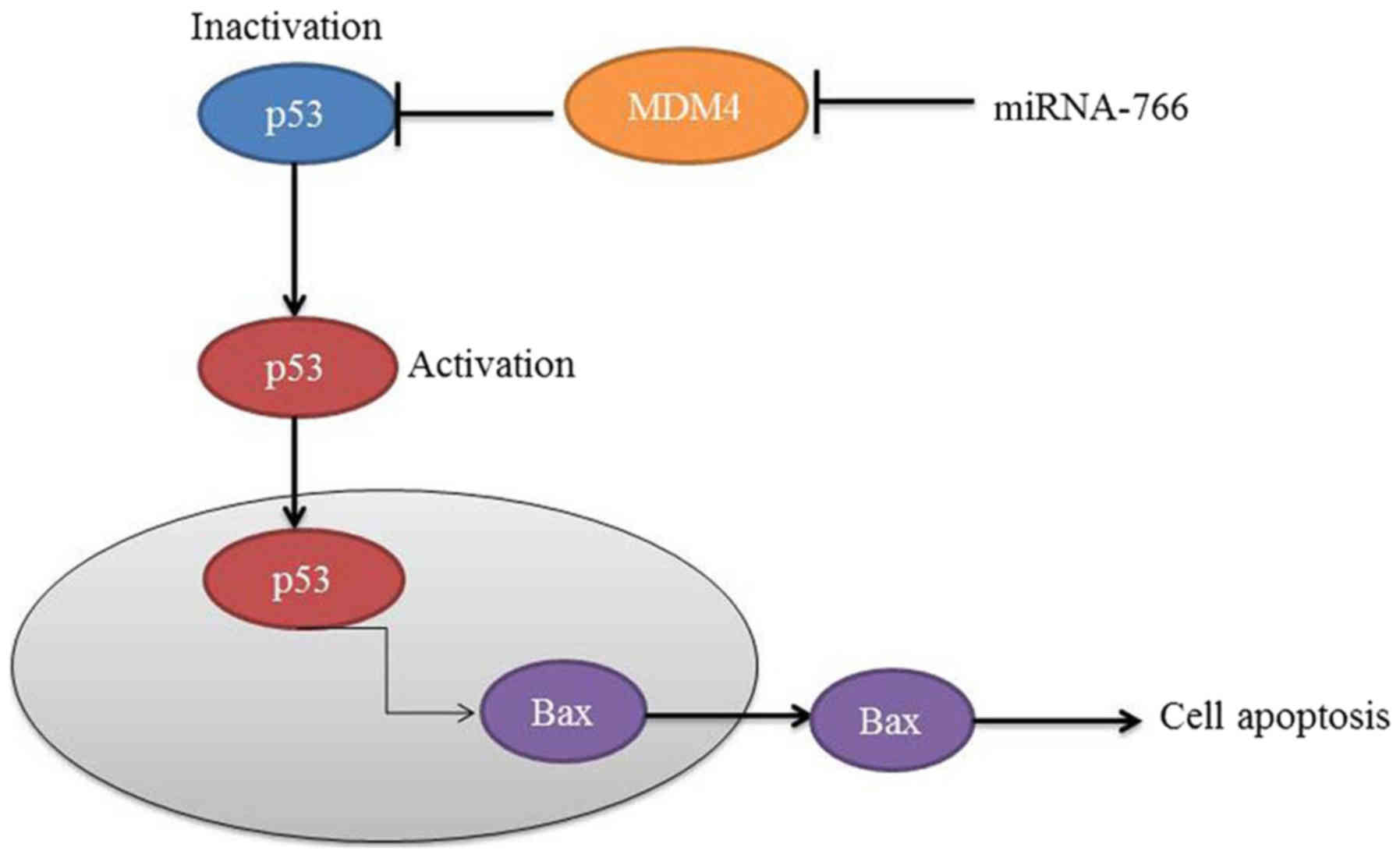
In conclusion, the data obtained in the present
study demonstrated that miRNA-766 reduced cell growth and cell
migration, and promoted LDH activity, apoptotic rate and
caspase-3/9 activity levels through MDM4/p53 in Caco2 cells
(Fig. 6). These findings provide a
direct link between miR-766/MDM4 and human colon cancer survival
rate and cell growth, which provides insight into the p53/Bax
pathway. Additionally, these results support the hypothesis that
genetic variants can interrupt miR-766-mediated gene regulation,
and this type of regulatory gene may be important modifiers of
human colon cancer risk.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
WC designed the experiments; GC, ZL, KL, GL and YL
performed the experiments; WC and GC analysed the data; and WC
wrote the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethical Agent
Will of the Second Affiliated Hospital, Shantou University Medical
College.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests
References
|
1
|
Liu X, Zhao W, Wang W, Lin S and Yang L:
Puerarin suppresses LPS-induced breast cancer cell migration,
invasion and adhesion by blockage NF-κB and Erk pathway. Biomed
Pharmacother. 92:429–436. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gan M and Yin X: Puerarin induced in
mantle cell lymphoma apoptosis and its possible mechanisms
involving multi-signaling pathway. Cell Biochem Biophys.
71:367–373. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Adlakha YK and Saini N: miR-128 exerts
pro-apoptotic effect in a p53 transcription-dependent and
-independent manner via PUMA-Bak axis. Cell Death Dis. 4:e5422013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ye Y, Zhuang J, Wang G, He S, Ni J and Xia
W: MicroRNA-139 targets fibronectin 1 to inhibit papillary thyroid
carcinoma progression. Oncol Lett. 14:7799–7806. 2017.PubMed/NCBI
|
|
5
|
Wang H, Li XT, Wu C, Wu ZW, Li YY, Yang
TQ, Chen GL, Xie XS, Huang YL, Du ZW and Zhou YX: miR-132 can
inhibit glioma cells invasion and migration by target MMP16 in
vitro. Onco Targets Ther. 8:3211–3218. 2015.PubMed/NCBI
|
|
6
|
Li JZ, Wang ZL, Xu WH, Li Q, Gao L and
Wang ZM: MicroRNA-495 regulates migration and invasion in prostate
cancer cells via targeting Akt and mTOR signaling. Cancer Invest.
34:181–188. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yao C, Shi X, Zhang Z, Zhou S, Qian T,
Wang Y, Ding F, Gu X and Yu B: Hypoxia-induced upregulation of
miR-132 promotes Schwann cell migration after sciatic nerve injury
by targeting PRKAG3. Mol Neurobiol. 53:5129–5139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Leinders M, Üçeyler N, Pritchard RA,
Sommer C and Sorkin LS: Increased miR-132-3p expression is
associated with chronic neuropathic pain. Exp Neurol. 283:276–286.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu Q, Zhang M, Tu J, Pang L, Cai W and Liu
X: MicroRNA-122 affects cell aggressiveness and apoptosis by
targeting PKM2 in human hepatocellular carcinoma. Oncol Rep.
34:2054–2064. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Prasad NB, Kowalski J, Tsai HL, Talbot K,
Somervell H, Kouniavsky G, Wang Y, Dackiw AP, Westra WH, Clark DP,
et al: Three-gene molecular diagnostic model for thyroid cancer.
Thyroid. 22:275–284. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Imamura T, Komatsu S, Ichikawa D, Miyamae
M, Okajima W, Ohashi T, Kiuchi J, Nishibeppu K, Konishi H, Shiozaki
A, et al: Depleted tumor suppressor miR-107 in plasma relates to
tumor progression and is a novel therapeutic target in pancreatic
cancer. Sci Rep. 7:57082017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu T, Pang Q, Wang Y and Yan X: Betulinic
acid induces apoptosis by regulating PI3K/Akt signaling and
mitochondrial pathways in human cervical cancer cells. Int J Mol
Med. 40:1669–1678. 2017.PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lages E, Guttin A, El Atifi M, Ramus C,
Ipas H, Dupré I, Rolland D, Salon C, Godfraind C, de Fraipont F, et
al: MicroRNA and target protein patterns reveal physiopathological
features of glioma subtypes. PLoS One. 6:e206002011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eun JW, Kim HS, Shen Q, Yang HD, Kim SY,
Yoon JH, Park WS, Lee JY and Nam SW: MicroRNA-495-3p functions as a
tumour suppressor by regulating multiple epigenetic modifiers in
gastric carcinogenesis. J Pathol. 244:107–119. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Oh K and Lee DS: In vivo validation of
metastasis-regulating microRNA-766 in human triple-negative breast
cancer cells. Lab Anim Res. 33:256–263. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jia B, Xia L and Cao F: The role of
miR-766-5p in cell migration and invasion in colorectal cancer. Exp
Ther Med. 15:2569–2574. 2018.PubMed/NCBI
|
|
18
|
Zaharieva ET, Kamenov ZA and Savov AS:
TLR4 polymorphisms seem not to be associated with prediabetes and
type 2 diabetes but predispose to diabetic retinopathy; TLR4
polymorphisms in glucose continuum. Endocr Regul. 51:137–144. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bahadori M, Baharara J and Amini E:
Anticancer properties of Chrysin on colon cancer cells, in vitro
and in vivo with modulation of caspase-3, −9, Bax and Sall4. Iran J
Biotechnol. 14:177–184. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
LeBlanc H, Lawrence D, Varfolomeev E,
Totpal K, Morlan J, Schow P, Fong S, Schwall R, Sinicropi D and
Ashkenazi A: Tumor-cell resistance to death receptor-induced
apoptosis through mutational inactivation of the proapoptotic Bcl-2
homolog Bax. Nat Med. 8:274–281. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang J, Chu ES, Chen HY, Man K, Go MY,
Huang XR, Lan HY, Sung JJ and Yu J: microRNA-29b prevents liver
fibrosis by attenuating hepatic stellate cell activation and
inducing apoptosis through targeting PI3K/AKT pathway. Oncotarget.
6:7325–7338. 2015.PubMed/NCBI
|
|
22
|
Yang SD, Ma L, Yang DL and Ding WY:
Combined effect of 17β-estradiol and resveratrol against apoptosis
induced by interleukin-1β in rat nucleus pulposus cells via
PI3K/Akt/caspase-3 pathway. PeerJ. 4:e16402016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Z, Shen J, Wu WK, Yu X, Liang J, Qiu G
and Liu J: Leptin induces cyclin D1 expression and proliferation of
human nucleus pulposus cells via JAK/STAT, PI3K/Akt and MEK/ERK
pathways. PLoS One. 7:e531762012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen HX, Xu XX, Tan BZ, Zhang Z and Zhou
XD: MicroRNA-29b inhibits angiogenesis by targeting VEGFA through
the MAPK/ERK and PI3K/Akt signaling pathways in endometrial
carcinoma. Cell Physiol Biochem. 41:933–946. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Q, Selth LA and Callen DF: MiR-766
induces p53 accumulation and G2/M arrest by directly targeting
MDM4. Oncotarget. 8:29914–29924. 2017.PubMed/NCBI
|