Introduction
Hearing impairment is one of the most relevant
chronic disorders in humans worldwide with an increasing incidence
in industrial countries. According to the WHO, 360 million people
currently suffer from disabling hearing loss (1). Congenital deafness, aging, acoustic
trauma or ototoxins (e.g., uremic, drug induced) are the most
common causes of partial or total hearing loss. Hearing loss due to
ototoxic drugs such as aminoglycosides or platinum-based
chemotherapeutic agents can be either reversible and temporary or
irreversible and permanent. The severity of the effect depends on
several factors including the level of the dose, duration of
treatment, genetic predisposition of the patient or animal and
route of administration (2–4). The hearing loss is typically
sensorineural, bilateral and progressive (5). Hearing loss is a major limiting factor
in the clinical use of aminoglycosides and represents one of the
main preventable causes of deafness (6–8). An
otoprotective compound is yet to be found.
Early and accurate detection of cochlear damage
during aminoglycoside administration is a major concern for health
care professionals. The treatment of choice to prevent drug-induced
ototoxicity is once-daily dosing and careful monitoring of serum
drug concentrations as well as auditory testing. At present,
finding otoprotective compounds is of vast interest. Cochlear hair
cell damage in vitro by aminoglycosides such as Gentamicin
and Kanamycin has been described in literature by several different
research groups and is an established known phenomenon. In
vivo experiments are performed to investigate drug-induced
cochelotoxicity and tolerance while co-morbidities ought to be
avoided. In order to research otoprotection in
aminoglycoside-induced hearing impairment, a reliable model for
preliminary tests performed in small cohorts needs to be
established. Common experimental animals have included guinea pigs,
rats and chinchillas. The larger rodents' pharmacokinetic profiles
differ significantly from those of small rodents such as mice
(9–11). In the past, large rodents have been
used not only because of their body size and easier breeding or
surgical intervention but also because of the greater drug-induced
toxicity of their auditory system compared to mice (3,12,13).
Nevertheless, mice are widely used and suitable as research models
and can be genetically modified. Genetic standardization and
relative ease of engineering make the mouse a preferred model for
hearing research in vivo, especially when studied at a young
age (up to 20 weeks) (5,12). Furthermore, the inner ear anatomy of
mice is similar to that of humans (14). The mouse has served as a successful
model in inner ear research, including auditory and vestibular
disorders (15), inner ear
development, noise-induced hearing loss and drug-induced
ototoxicity (13,16–18).
Yet, mouse models have shown high resistance to
aminoglycoside-induced cochlear damage (13). Some researchers have combined
aminoglycosides and loop diuretics to take advantage of their
synergistic effect to increase the rate of cochlear injury
(17,19–22).
To identify a mouse model for cochlear injury,
different research groups (5,9,12,13,16–18,22,23) have
investigated aminoglycoside ototoxicity in different strains of
mice, which has yielded variable results for similar drug dose
regimens. Also, similar dose regimens in identical mouse strains
have produced dissimilar results. Kendall et al (24), have suggested the use of C57BL/6J
strains in auditory research instead of C57BL/6N, as the latter
have genetically drifted within their subpopulation and show
irregular auditory thresholds among different populations. A robust
in vivo mouse model for aminoglycoside-induced ototoxicity
remains to be established. The most commonly used method to assess
functional auditory effects is the auditory brainstem response
(ABR). Less frequently, distortion product otoacoustic emission
(DPOAE) have been used. Wu et al (13), presented an adult mouse model for
kanamycin ototoxicity using ABR only. We, however, assessed
different dose regimens for gentamicin- and kanamycin-induced
ototoxocity, that is capable of causing substantial cochlear damage
in young mice while maintaining a low mortality rate and being able
to detect cochlear damage in a small cohort, which can be used as a
model for future preliminary cochelotoxic tests.
Materials and methods
Animals
Four- to 6-week-old C57BL/6J mice supplied by
Charles River (Charles River, Freiburg im Breisgau, Germany) were
used to assess the optimal aminoglycoside dose regiment needed to
induce ototoxic hair cell injury for purposes of research. Mice
were divided and housed in cages (Type 2 IVC; Allentown Inc.,
Allentown, PA, USA) in groups of four (M1-M4), with wood shaving
litter (Lignocel select premium; J. Rettenmaier & Söhne GmbH,
Rosenberg, Germany) under standard conditions: 12:12 h photoperiod,
room temperature between 20 and 22°C, relative humidity of 45–55%
and 20–22 air changes per hour. Standard mouse food (cat. no. 3436,
Kliba Nafag Provimi Kliba AG, Kaiseraugst, Switzerland) and water
were available ad libitum. All mice were allowed to acclimatize to
the animal facility for at least one week before testing was begun.
Health monitoring was performed following FELASA recommendations
(25). All animal procedures were
carried out according to our approved animal research protocols
(permission number 19/2011, Veterinary Department of Zurich,
Kantonales Veterinäramt Zürich, Switzerland). Due to the previously
described high mortality rates with systemic gentamicin
administration (5), the local
veterinary department (Kantonales Veterinäramt Zürich, Switzerland)
permitted us to use a limited cohort of animals (n=16) for the
experiments. Three groups received different aminoglycoside
treatment protocols including gentamicin, kanamycin and kanamycin
plus furosemide. The fourth group served as a control and received
no treatment. Every mouse was checked daily during the experiment
for weight and activity behavior.
Drug administration
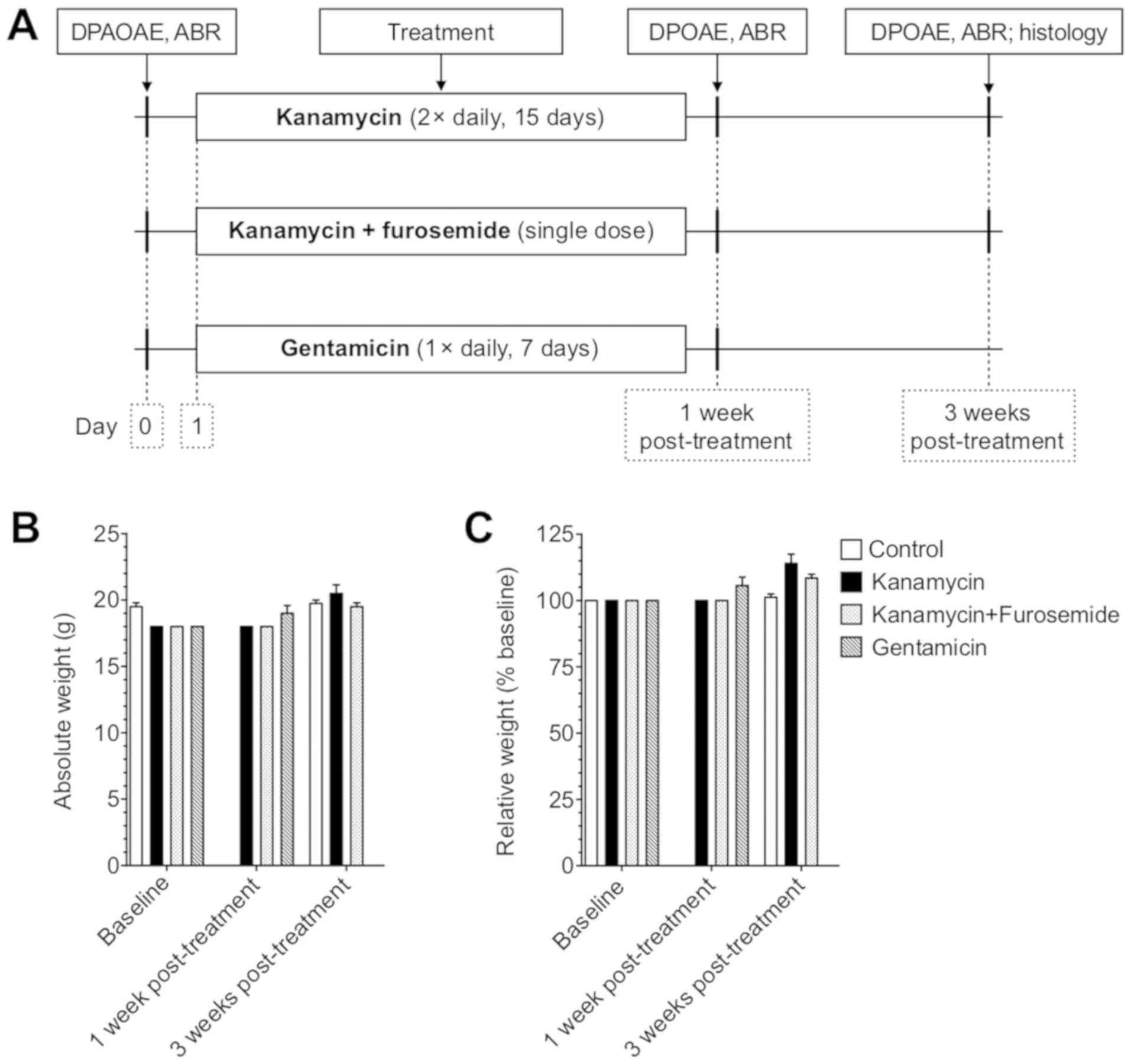
The details of dosing and timing are listed in
Table I and Fig. 1A. Mice from each experimental group
were treated with the corresponding protocol. Kanamycin (Sigma
Aldrich, Buchs, Switzerland) was dissolved in physiological saline
to obtain the desired concentration according to the instructions
provided by the supplier. Each mouse from the gentamicin (Hexal,
Holzkirchen, Germany) group received a once-daily dose by
intraperitoneal (i.p.) drug administration at 8:00 a.m. for 7 days.
Each mouse from the kanamycin group received twice daily
subcutaneous (s.c.) drug administration at 8:00 a.m. and 8:00 p.m.
for 15 days. Kanamycin plus furosemide (Sanofi-Aventis, Vernier,
Switzerland) was administered as a single dose in which furosemide
was infused into the tail vein within 5 min after kanamycin s.c.
injection. The dosage administered was selected on the basis of
previously published and weight-adapted ototoxic protocols using
gentamicin at 200 mg/kg body weight (BW) (12), kanamycin at 800 mg/kg BW (5), kanamycin plus furosemide at 1,000 mg/kg
BW plus 100 mg/kg BW (22). The
injections were adjusted daily according to body weight and were
administered using a 1-ml syringe and 25-gauge needle in a
standardized manner by the same person in the same setting, at the
same time each day.
 | Table I.Treatment regimens and mortality
rate. |
Table I.
Treatment regimens and mortality
rate.
| Drug/Group | Dose, rate,
route | No. of deaths/No.
of injected mice (mortality rate) (%) |
|---|
| Kanamycin | 800 mg/kg/12 h, 15
days, s.c | 0/4 (0) |
| Kanamycin +
furosemide | 1,000 mg/kg, s.c. +
100 mg/kg i.v. | 0/4 (0) |
| Gentamicin | 200 mg/kg/24 h, 7
days, i.p. | 2/4 (50) |
Measures of auditory function
The ABR and DPOAE measurements were performed on all
animals before starting the treatment protocols, at 1-week and the
two groups with kanamycin were also tested at 3 weeks
post-treatment (Fig. 1A). The DPOAE
were performed one day after ABR measurements, therefore in two
separate anesthesias. Mice were anaesthetized by i.p. injection of
a combination of ketamine (65 mg/kg body weight, Graeub,
Switzerland), xylazine (13 mg/kg body weight, Bayer HealthCare,
Leverkusen, Germany) and acepromazine (2 mg/kg body weight, Fatro
S.p.A., Ozzano dell'Emilia, Italy). After loss of the withdrawal
reflex, the animals were placed on a heating pad in a soundproof
chamber. Photos showing the method for hearing assessment are
provided as supplementary material (Fig. S1).
ABR testing
Hearing was evaluated by ABR as previously described
(26). Needle electrodes were
inserted subcutaneously at the vertex (active electrode), in the
ipsilateral mastoid region (reference electrode) and in the lumbar
region (ground electrode). Tone bursts of 4, 8, 12, 16, 24, 32 kHz
were generated with SigGenRP software (Tucker-Davis Technologies
TDT, Gainesville, FL, USA). The stimuli were delivered through a
closed acoustic system and were calibrated using a sound level
meter (Precision Integrating Sound Level Meter Type 2218; Brüel
& Kjaer, Naerum, Denmark) and an ear simulator (Type: 4157,
Brüel & Kjaer, Naerum, Denmark). Sounds were delivered from a
free-field electrostatic speaker (ES1, TDT) placed into the ear
canal. The ABR recordings were obtained with a Tucker-Davis
Technologies (TDT) System III workstation running BioSig RP. The
tone-type sounds of 5 ms duration were presented at a rate of 10
per second and reduced in level from 80 dB SPL to 5 dB SPL in 5-dB
steps. The ABR waveforms were averaged in response to 300 tones.
Hearing threshold was defined as the lowest level that induced the
appearance of a visually detectable peak in the response
waveform.
DPOAE testing
DPOAE at 2f1-f2 were obtained 24 h following ABR
recordings from mice that were anesthesized as described above. We
used the Real-time Signal Processing System III from Tucker-Davis
Technologies and procedures described previously (27). The primary tones produced by two
separate speakers (ES1, TDT) were placed as a combination
microphone/speaker system in the animal's sealed ear canal near the
tympanic membrane. The DPOAE recordings were made with a low-noise
microphone ER 10B (Etymotic Research, Elk Grove Village, IL, USA).
All stimuli were digitally synthesized at 200 kHz using TDT SigGen
software. Primary tone frequencies (f1 and f2) differed by a factor
of 1.25 and were presented initially at 65- and 50-dB SPL
respectively. The test frequencies were at 4, 8, 16, 32 kHz and
levels were reduced in 10-dB steps from 80 to 30 dB. A fast Fourier
Transform (FFT) was performed to obtain the magnitude of the 2f1-f2
distortion product. A peak at 2f1-f2 in the spectrum was accepted
as a DPOAE if it was 6 dB above the noise floor in the same
frequency.
Hair cell count
Mice were transcardially perfused with freshly
prepared 4% paraformaldehyde dissolved in PBS (pH 7.2) during
anesthesia. The cochleae were removed and the round and oval
windows were opened. The cochleae were postfixed for 48 h in the
same fixative, decalcified for 96 h in RDF Mild Decalcifier
(CellPath Ltd., Newtown Powys, Wales, UK) and embedded in paraffin.
Serial sections (2 µm) were cut using a HM 355S Automatic Microtome
(Thermo Fisher Scientific, Inc., Waltham, MA, USA). Sections were
collected on SuperFrost Plus slides (Thermo Fisher Scientific,
Inc.), dried on a heating plate at 37°C overnight and stained with
H&E. Observer-blinded inner hair cell (IHC) and outer hair cell
(OHC) counts at each of the basal, middle and apical cochlear turn
were performed on para-midmodiolar sections obtained from five
non-overlapping 20-µm segments in each cochlea. For each segment,
three consecutive sections were mounted and the best-preserved
section was used for counting. The presence of a hair cell was
assumed if a nucleus was clearly visible in the typical anatomical
location next to the tunnel of Corti. Additionally, hair cells were
distinguished from other cells, e.g., supporting cells, by their
denser and therefore darker stained nuclei. Images were acquired
using an AxioCam ICc5 (Carl Zeiss Microscopy GmbH, Jena, Germany)
on a Leica DM RB light microscope (Leica Camera AG, Wetzlar,
Germany) and processed with Adobe Photoshop CS5 software (v.12.0,
Adobe Systems, San Jose, CA, USA).
Statistical analysis
ABR, DPOAE and hair cell counts were analysed by
two-way ANOVA with Dunnett's multiple comparison testing using
Prism (v.6.0 for Apple Macintosh, GraphPad Software, San Diego, CA,
USA). Data are presented as mean ± SEM. P<0.05 was considered to
indicate a statistically significant difference.
Results
Systemic administration of kanamycin
is a safe method to cause hearing damage in young mice
The administration of kanamycin in two different
regimens (Table I) yielded a
mortality rate of 0%. None of the animals appeared ill and no
weight change >15% was observed during and after treatment
(Fig. 1B and C). In contrast,
systemic administration of previously reported ototoxic doses of
aminoglycosides (5,12–22), has
been associated with adverse effects. The gentamicin-group had the
highest systemic toxicity with a mortality rate of 50%. One mouse
was found dead within 24 h after the first injection and another
mouse died within 24 h after the last injection on day 7.
Hearing assessment
Hearing impairment due to ototoxic effects was
assessed by measuring frequency-specific ABR and DPOAE threshold
before and after antibiotic treatment. After conferring with the
veterinary department, the gentamicin group experiment was
dismissed on Day 7 after occurrence of a 50% mortality rate. The
control group was measured on day 0 and day 21. At baseline,
average hearing threshold of all mice was 13 dB SPL (range +/−
maximum 15 dB) for the 16 kHz tone ABR, where mice are reported to
hear most sensitively (28).
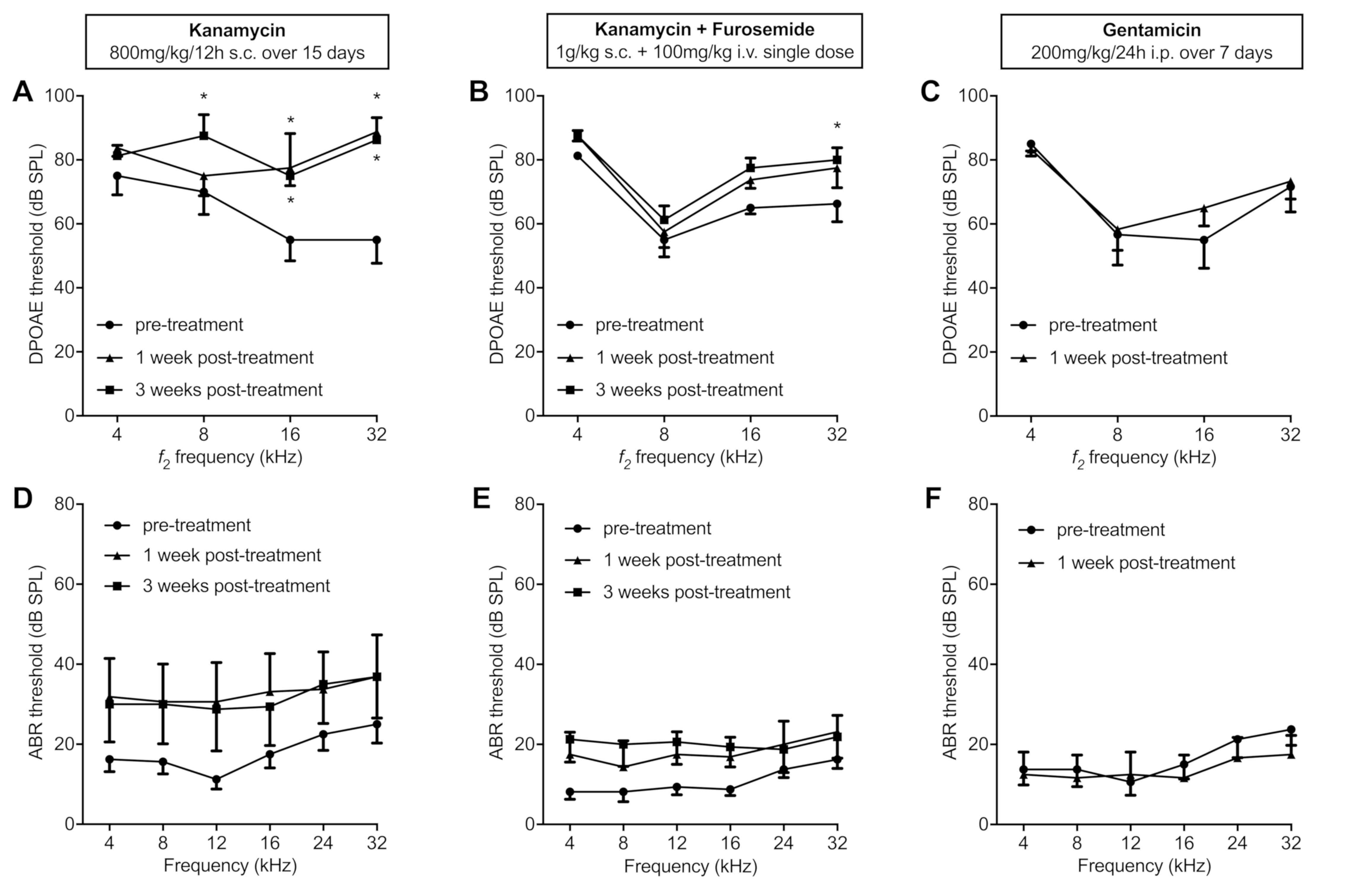
DPOAE recordings showed significant
threshold shifts in the higher frequencies at early stage
The kanamycin regimen caused significant cochlear
damage, which was detectable by DPOAE, showing threshold shifts in
the higher frequencies (8, 16 and 32 kHz). Combined with furosemide
as a single dose, a significant threshold shift was detected at 32
kHz. In the gentamicin group, DPOAE were not significantly altered
post treatment (Fig. 2A-C). The
control group had no significant changes in DPOAE threshold.
ABR recordings showed a tendency
toward an increase of hearing thresholds
The overall threshold shift between pre- and
post-treatment for the kanamycin only group (Fig. 2D) ranged from 12–18 dB (mean 14.3
dB). The kanamycin-furosemide group (Fig. 2E) had overall threshold shifts of
5–13 dB (mean 9.6 dB) between pre- and post-treatment. The
gentamicin group (Fig. 2F) as well
as the control group showed no substantial threshold shifts. When
focused on individual animal threshold measurements, the shifts
showed a heterogenic variance between the mice and between the two
ears of the same mouse (data not shown). No statistical significant
threshold shifts were observed in any group by ABR, however ABR
show a trend for hearing threshold elevation after
kanamycin-induced ototoxicity.
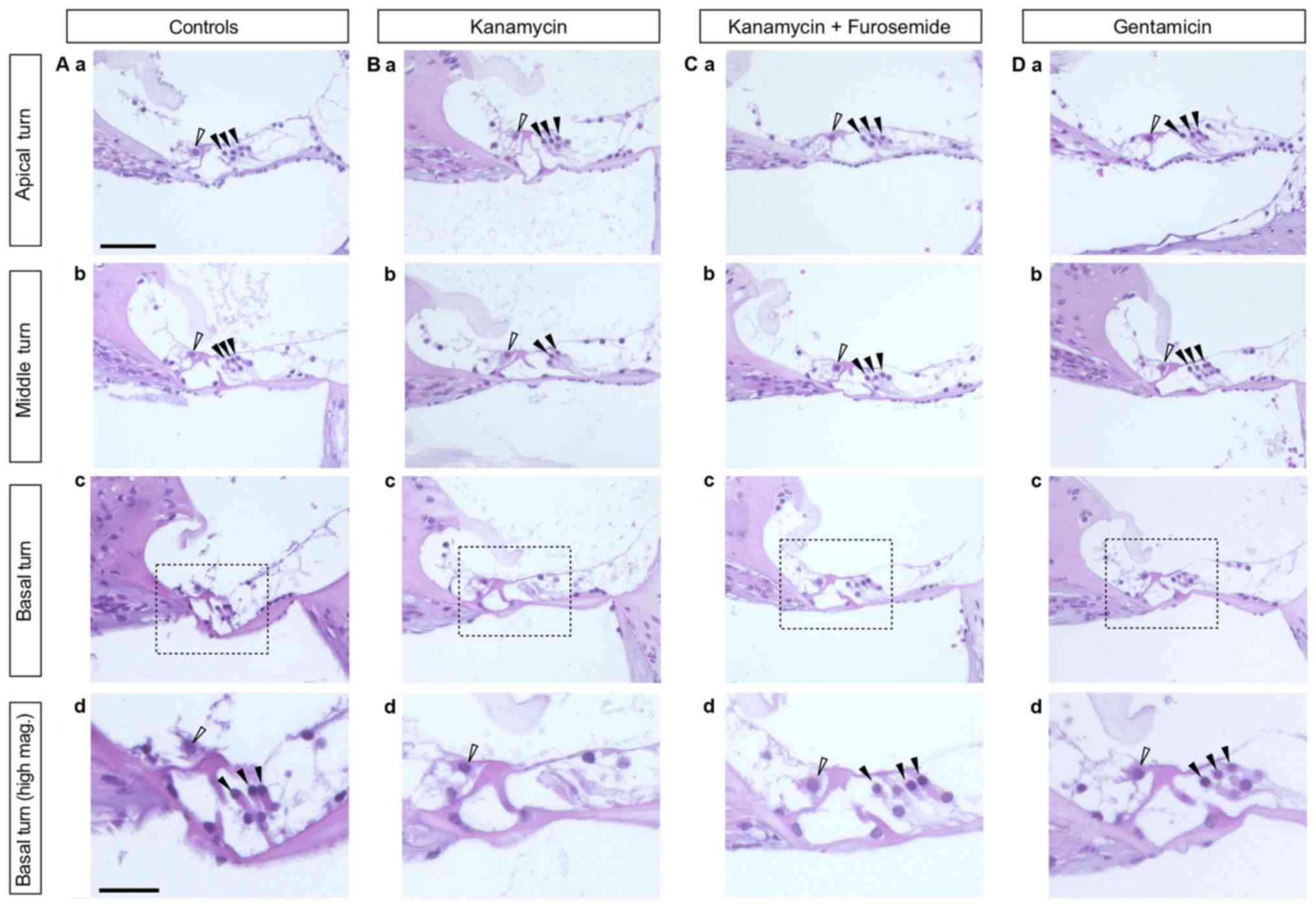
Kanamycin treatment causes a
significant hair cell loss in the organ of Corti
Histological evaluation of the cochleae revealed a
considerably reduced amount of OHC per organ of Corti (OC) in the
kanamycin-only group, especially in the basal turn, compared to the
control group, where no OHC loss was observed (Fig. 3). No IHC loss was found in any group
and no OHC loss was found in the gentamicin group. To quantify
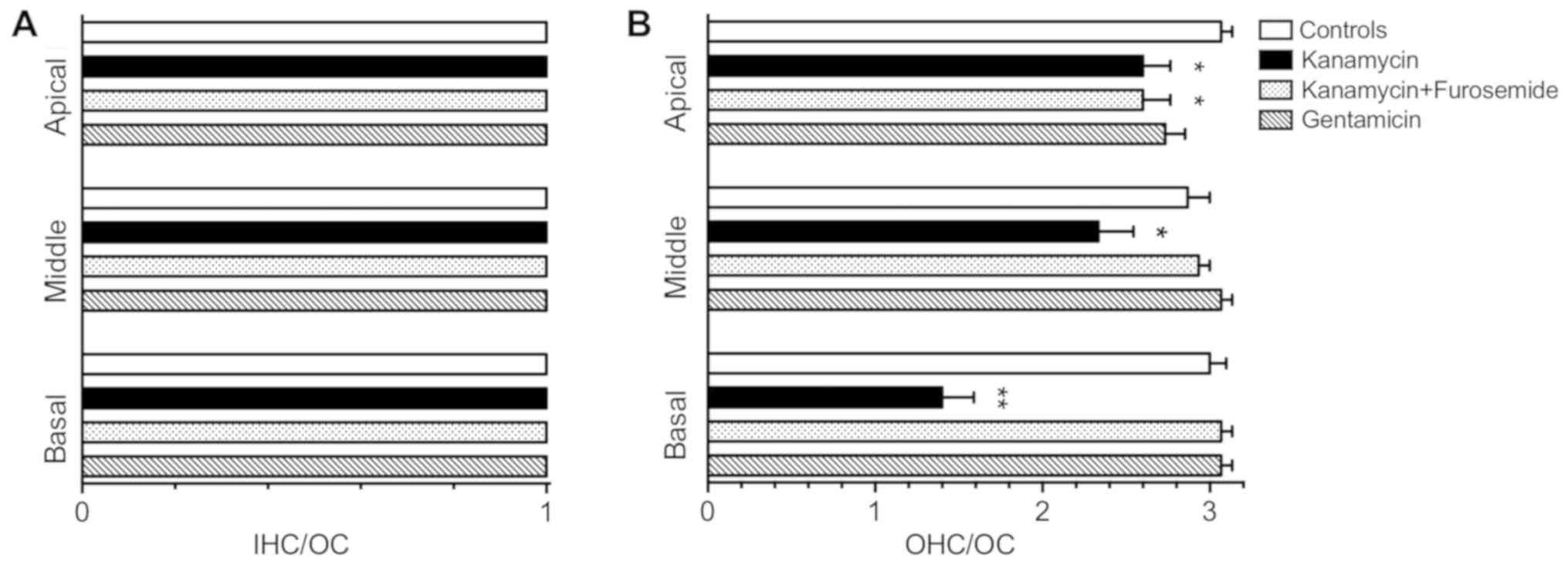
these changes, we performed IHC and OHC counts on sections of 3
animals per group (Fig. 4). OHC loss
in the kanamycin group was predominant in the basal turn of the
cochlea (1.4±0.2 OHC/OC vs. 3.0±0.1 OHC/OC, P<0.0001), less
marked in the middle turn (2.3±0.2 OHC/OC vs. 2.9±0.1 OHC/OC,
P<0.05) and only a slight hair cell loss was observed in the
apical turn (2.6±0.16 OHC/OC vs. 3.1±0.1 OHC/OC, P<0.05). No
significant OHC loss was found in the kanamycin-furosemide and the
gentamicin group, except in the apical turn, where the
kanamycin-furosemide produced a minor but significant OHC loss
compared to the control group (2.6±0.16 OHC/OC vs. 3.1±0.1 OHC/OC,
P<0.05). No significant IHC loss was found in any treatment
group compared to the control group.
Discussion
Hearing impairment caused by cochleotoxic agents is
a prevalent clinical issue. Ongoing studies are trying to find
protective agents against ototoxicity. The mouse has served as a
consistent model to study the inner ear in vivo. So far,
there have been no precise recommendations regarding ototoxic
regimen in a mouse model, fulfilling requirements such as being
simple to perform, safe and causing substantial inner ear damage.
In order to evaluate various mouse models for ototoxic injury
caused by various aminoglycoside regimens, preliminary tests in
small cohorts need to be performed preceding large scaled
experiments. The aim of our study was to investigate different
aminoglycoside regimens that cause a substantial ototoxic damage
in vivo, which are simple, safe and produce a detectable
hearing threshold shift already in a small cohort of mice. We show
that an ototoxic regimen with kanamycin twice daily for 15
consecutive days is simple to employ, well tolerated and produces a
significant early hearing threshold shift in a small cohort of
mice. At the same time, the small cohort itself and the lack of
saline injection of the control group is a limitation to our study.
We demonstrate a higher sensitivity of DPOAE, as thereby a hearing
threshold shift is detectable earlier in high frequencies and
correlates with the outer hair cell damage mainly in the basal turn
of the cochlea. The known target of aminoglycoside ototoxicity are
the hair cells of the organ of Corti, which are responsible for the
mechano-sensory transduction of sound in the auditory system
(29). These vulnerable hair cells
are not capable of regeneration, and their damage leads to lifelong
hearing impairment. ABR have been used to characterize the auditory
system of various mouse strains and mutants. The DPOAE serve as a
noninvasive tool to assess cochlear function, specifically that of
the vulnerable OHCs. While ABR have been used as the main
non-invasive method to assess auditory function in mouse models due
to the simplicity and reliability of the method, DPOAE can provide
a more direct means of assessing peripheral function by assessing
OHC functional changes. Aminoglycosides have a narrow therapeutic
range causing large spreads in hearing thresholds leading to high
standard deviations. Mice have proven to have a high resistance to
aminoglycoside-induced hair cell loss and to cochlear damage in
general (18). Before appreciable
alterations in auditory function after aminoglycoside treatment was
detected, our mouse cohorts required close to lethal doses for
treatment, with the gentamicin group showing obvious systemic toxic
effects. We chose to study the C57BL/6 strain because it is widely
used as a transgenic modified mouse model and is therefore
interesting as an in vivo animal model, especially to study
aminoglycoside-induced ototoxicity. Age is also a factor, as young
mice are more ideal as aminoglycoside ototoxicity models because
they have a higher sensitivity to aminoglycosides than in adult
life (12,13). Poirrier et al (9), hypothesized that genetic and
environmental differences between mouse strains influence responses
to various toxins including ototoxins. Researchers have been
combining aminoglycosides and loop diuretics to study the loss of
hair cells in rodents and describe a significant increase in
hearing loss (16–18,23–30). The
synergistic effects of a loop diuretic and an aminoglycoside occur
because the aminoglycosides pass the blood-brain barrier allowing
to spread effectively through the cochlea (31,32)
combined with a decreased renal clearance (33). Because our kanamycin plus furosemide
group received only a single dose and showed no significant hearing
loss nor hair cell damage, we conclude that this dosage is
insufficient in producing enough cochlear damage. In the inner ear,
damage is first evident as a loss of OHCs at the base of the
cochlea, spreading further towards the apex with continued drug
treatment (7). Clinically, the
ototoxic effect of aminoglycosides is characterized by a hearing
loss initially in the high frequencies corresponding to hair cell
damage in the lower basal turn (7,34). We
also found early detection of hearing threshold shifts in the high
frequencies with kanamycin only and with kanamycin plus furosemide
in a small cohort. DPOAE were able to detect these threshold
changes early. Tan et al (35) examined the effect of sub-damaging
aminoglycoside doses on noise-induced hearing loss in guinea-pigs
using DPOAE and ABR before and after treatment. Aksoy et al
(36,37) assessed hearing in rats before and
after amikacin and trimetazidine or betahistin application.
Moreover, Shi et al (38)
were one of the first to demonstrate that DPOAE are preferable to
ABR testing in the early detection of gentamicin toxicity of the
cochlea in guinea pigs. They suggest that DPOAE provide earlier
detection of cochlear damage caused by gentamicin than do ABRs.
Kakigi et al (39) compared
ABR, DPOAE and transiently evoked otoacoustic emissions (TEOAE) in
chinchillas treated with aminoglycosides. Their results suggest
that DPOAE can be used to monitor hair cell function more
accurately at specific anatomical locations than can the other
methods. Peguero et al (40)
assessed hearing with both methods in different mouse strains
(CBA/CaJ, 129S6/SvEvTac and 101/H). They determined that DPOAE
detect early-onset OHC dysfunction despite normal ABR thresholds.
It was evident in our study that DPOAE revealed hearing impairment
early, while ABR still did not show any significant elevation of
thresholds. Furthermore, we showed that Kanamycin alone for a
prolonged application is able to cause damage in this small cohort
of mice. A number of research groups have attempted to find an
aminoglycoside inner ear damage regimen for rodents using different
animals and strains, various drugs, doses and application methods.
They have used primarily ABR for this purpose.
Three mouse groups living under identical conditions
received different drug regimens to induce cochlear damage by
systemic administration of high aminoglycoside doses. Significant
threshold changes after kanamycin treatment were detected by DPOAE
in the high frequencies (8, 32 and 16 kHz). ABR was not able to
detect significant threshold changes after 3 weeks. The kanamycin
group had an overall shift of 12–19 dB SPL. The combination of
kanamycin and furosemide lead to threshold shifts at 32 kHz. None
of the kanamycin-treated groups had any deaths during or after the
experiments. Histological examination and hair cell counts
particularly showed a marked hair cell loss in the basal turn of
the cochlea in mice treated with kanamycin, which in accordance
with the functional measurements. The DPOAE measures detected
hearing impairment primarily in the high frequencies, corresponding
to the basal cochlear hair cells, defining OHC damage. While ABRs
reflect the summed activity of the peripheral neuronal auditory
system, DPOAEs are generated presynaptically and depend only on the
integrity of the OHC. Gentamicin at 200 mg/kg/24 h caused no
significant threshold shift in DPOAE and ABR and was unsafe causing
a high mortality rate. ABR showed a tendency towards hearing
threshold shifts. While DPOAE are able to detect cochlear damage in
a small cohort already, we postulate that a larger cohort is needed
to show significant changes with ABR. Therefore, DPOAE can be used
in preliminary experiments finding cochlear damage in early stage
and Kanamycin is a preferred method for cochlear damage in
mice.
In summary, the present study compares for the first
time DPOAE and ABR with aforementioned aminoglycoside regimens and
demonstrates that Kanamycin treatment is a simple, reliable and
safe regimen to induce damage to auditory function. Despite a small
cohort and high-dosed administration period, the mortality rate was
0%. Significant hearing threshold changes were produced by the
kanamycin regimen in a small cohort of mice and detected by DPOAE
in early stage. DPOAE can therefore be used for early detection of
outer hair cell damage induced by Kanamycin 800 mg s.c. 2×/day for
15 days.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Mrs. Katherine
Horvath for the correction of the manuscript.
Funding
The present study was supported by the Zürcher
Stiftung für das Hören (to Arianne Monge Naldi).
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LH, DBa and AMN designed the study. LH performed
drug administration, ABR and DPOAE testing. DBa and TH performed
the hair cell count. DBa performed the statistical analysis. DBo
contributed to the designing the study and revising the manuscript
critically for important intellectual content. LH and DBa wrote the
manuscript. The final version of the manuscript has been read and
approved by all authors.
Ethics approval and consent to
participate
The study has been granted ethics approved with
consent to participate by the Veterinary Department of Zurich
(permission number 19/2011, Kantonales Veterinäramt Zürich,
Switzerland).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
World Health Organization, Geneva. WHO
Fact sheet 2018. Deafness and hearing loss, . http://www.who.int/mediacentre/factsheets/fs300/en/March
20–2019
|
|
2
|
Brummett RE and Morrison RB: The incidence
of aminoglycoside antibiotic-induced hearing loss. Arch Otolaryngol
Head Neck Surg. 116:406–410. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Blakley BW, Hochman J, Wellman M, Gooi A
and Hussain AE: Differences in ototoxicity across species. J
otolaryngol Head Neck Surg. 37:700–703. 2008.PubMed/NCBI
|
|
4
|
Guthrie OW: Aminoglycoside induced
ototoxicity. Toxicology. 249:91–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Murillo-Cuesta S, Contreras J, Cediel R
and Varela-Nieto I: Comparison of different aminoglycoside
antibiotic treatments to refine ototoxicity studies in adult mice.
Lab Anim. 44:124–131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Corrado AP, de Morais IP and Prado WA:
Aminoglycoside antibiotics as a tool for the study of the
biological role of calcium ions. Historical overview. Acta Physiol
Pharmacol Latinoam. 39:419–430. 1989.PubMed/NCBI
|
|
7
|
Forge A and Schacht J: Aminoglycoside
antibiotics. Audiol Neurootol. 5:3–22. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pichler M, Wang Z, Grabner-Weiss C, Reimer
D, Hering S, Grabner M, Glossmann H and Striessnig J: Block of
P/Q-type calcium channels by therapeutic concentrations of
aminoglycoside antibiotics. Biochemistry. 35:14659–14664. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Poirrier AL, Van den Ackerveken P, Kim TS,
Vandenbosch R, Nguyen L, Lefebvre PP and Malgrange B: Ototoxic
drugs: Difference in sensitivity between mice and guinea pigs.
Toxicol Lett. 193:41–49. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Walton K, Dorne JL and Renwick AG:
Species-specific uncertainty factors for compounds eliminated
principally by renal excretion in humans. Food Chem Toxicol.
42:261–274. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang B and Bankir L: Urea and urine
concentrating ability: New insights from studies in mice. Am J
Physiol Renal Physiol. 288:F881–F896. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen L, Xiong S, Liu Y and Shang X: Effect
of different gentamicin dose on the plasticity of the ribbon
synapses in cochlear inner hair cells of C57BL/6J mice. Mol
Neurobiol. 46:487–494. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu WJ, Sha SH, McLaren JD, Kawamoto K,
Raphael Y and Schacht J: Aminoglycoside ototoxicity in adult CBA,
C57BL and BALB mice and the Sprague-Dawley rat. Hear Res.
158:165–178. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Steel KP and Bock GR: Hereditary inner-ear
abnormalities in animals. Relationships with human abnormalities.
Arch Otolaryngol. 109:22–29. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Probst FJ and Camper SA: The role of mouse
mutants in the identification of human hereditary hearing loss
genes. Hear Res. 130:1–6. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hartman BH, Basak O, Nelson BR, Taylor V,
Bermingham-McDonogh O and Reh TA: Hes5 expression in the postnatal
and adult mouse inner ear and the drug-damaged cochlea. J Assoc Res
Otolaryngol. 10:321–340. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hirose K and Sato E: Comparative analysis
of combination kanamycin-furosemide versus kanamycin alone in the
mouse cochlea. Hear Res. 272:108–116. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Taylor RR, Nevill G and Forge A: Rapid
hair cell loss: A mouse model for cochlear lesions. J Assoc Res
Otolaryngol. 9:44–64. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
West BA, Brummett RE and Himes DL:
Interaction of kanamycin and ethacrynic acid. Severe cochlear
damage in guinea pigs. Arch Otolaryngol. 98:32–37. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nourski KV, Miller CA, Hu N and Abbas PJ:
Co-administration of kanamycin and ethacrynic acid as a deafening
method for acute animal experiments. Hear Res. 187:131–133. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Russell NJ, Fox KE and Brummett RE:
Ototoxic effects of the interaction between kanamycin and
ethacrynic acid. Cochlear ultrastructure correlated with cochlear
potentials and kanamycin levels. Acta Otolaryngol. 88:369–381.
1979. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jansen TT, Bremer HG, Topsakal V,
Hendriksen FG, Klis SF and Grolman W: Deafness induction in mice.
Otol Neurotol. 34:1496–1502. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Oesterle EC and Campbell S: Supporting
cell characteristics in long-deafened aged mouse ears. J Assoc Res
Otolaryngol. 10:525–544. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kendall A and Schacht J: Disparities in
auditory physiology and pathology between C57BL/6J and C57BL/6N
substrains. Hear Res. 318:18–22. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
FELASA working group on revision of
guidelines for health monitoring of rodents and rabbits, ; Mähler
Convenor M, Berard M, Feinstein R, Gallagher A, Illgen-Wilcke B,
Pritchett-Corning K and Raspa M: FELASA recommendations for the
health monitoring of mouse, rat, hamster, guinea pig and rabbit
colonies in breeding and experimental units. Lab Anim. 48:178–192.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Horvath L, Bodmer D, Radojevic V and Monge
Naldi A: Activin signaling disruption in the cochlea does not
influence hearing in adult mice. Audiol Neurootol. 20:51–61. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mhatre AN, Li Y, Bhatia N, Wang KH, Atkin
G and Lalwani AK: Generation and characterization of mice with Myh9
deficiency. Neuromolecular Med. 9:205–215. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Koay G, Heffner R and Heffner H:
Behavioral audiograms of homozygous med(J) mutant mice with sodium
channel deficiency and unaffected controls. Hear Res. 171:111–118.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sedo-Cabezon L, Boadas-Vaello P,
Soler-Martin C and Llorens J: Vestibular damage in chronic
ototoxicity: A mini-review. Neurotoxicology. 43:21–27. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Versnel H, Agterberg MJ, de Groot JC,
Smoorenburg GF and Klis SF: Time course of cochlear
electrophysiology and morphology after combined administration of
kanamycin and furosemide. Hear Res. 231:1–12. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu H, Ding DL, Jiang HY, Wu XW, Salvi R
and Sun H: Ototoxic destruction by co-administration of kanamycin
and ethacrynic acid in rats. J Zhejiang Univ Sci B. 12:853–861.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ding D, McFadden SL, Browne RW and Salvi
RJ: Late dosing with ethacrynic acid can reduce gentamicin
concentration in perilymph and protect cochlear hair cells. Hear
Res. 185:90–96. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ohtani I, Ohtsuki K, Omata T, Ouchi J and
Saito T: Potentiation and its mechanism of cochlear damage
resulting from furosemide and aminoglycoside antibiotics. ORL J
Otorhinolaryngol Relat Spec. 40:53–63. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fausti SA, Rappaport BZ, Schechter MA,
Frey RH, Ward TT and Brummett RE: Detection of aminoglycoside
ototoxicity by high-frequency auditory evaluation: Selected case
studies. Am J Otolaryngol. 5:177–182. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tan CT, Hsu CJ, Lee SY, Liu SH and
Lin-Shiau SY: Potentiation of noise-induced hearing loss by
amikacin in guinea pigs. Hear Res. 161:72–80. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Aksoy F, Dogan R, Ozturan O, Eren SB,
Veyseller B, Pektas A and Hüseyinbas Ö: Protective effect of
trimetazidine on amikacin-induced ototoxicity in rats. Int J
Pediatr Otorhinolaryngol. 78:663–669. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Aksoy F, Dogan R, Ozturan O, Yildirim YS,
Veyseller B, Yenigun A and Ozturk B: Betahistine exacerbates
amikacin ototoxicity. Ann Otol Rhinol Laryngol. 124:280–287. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shi Y and Martin WH: ABR and DPOAE
detection of cochlear damage by gentamicin. J Basic Clin Physiol
Pharmacol. 8:141–155. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kakigi A, Hirakawa H, Harel N, Mount RJ
and Harrison RV: Comparison of distortion-product and transient
evoked otoacoustic emissions with ABR threshold shift in
chinchillas with ototoxic damage. Auris Nasus Larynx. 25:223–232.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Peguero B and Tempel BL: A chromosome 17
locus engenders frequency-specific non-progressive hearing loss
that contributes to age-related hearing loss in mice. J Assoc Res
Otolaryngol. 16:459–471. 2015. View Article : Google Scholar : PubMed/NCBI
|