Introduction
Lung cancer is one of the most common malignant
tumors that is a major cause of various cancer-related deaths for
men and women, worldwide. In 2012, there were ~1.8 million newly
registered cases of lung cancer, with 1.6 million deaths occurring
globally (1). Non-small cell lung
cancer (NSCLC) accounts for ~85% of all lung-cancer-related deaths
and can be classified as adenocarcinoma (40%), squamous cell
carcinoma (<40%) and large cell carcinoma (5%). It is well
established that tobacco smoke is the main cause of lung cancer;
however, a series of studies have identified that occupational and
environmental exposure to cadmium (Cd) is another important risk
factor (2–5).
Cd is a widely used toxic industrial heavy metal
that causes serious environmental health hazards to humans. The
International Agency for Research on Cancer has classified Cd as a
known human carcinogen (6). The
primary means of Cd intake by humans are via food (particularly
plant products), habitual tobacco smoking, drinking contaminated
sources and Cd-related industry. Increasing evidence has revealed a
correlation between Cd exposure and the formation of lung cancer
(2,7,8). It has
also been reported that oxidative stress, apoptotic resistance,
induction of autophagy, decreased DNA repair capacity and genomic
instability are involved in the development of lung cancer induced
by Cd (9). Recently, exposure to Cd
has been correlated with the migration and invasion of lung cancer
cells (10,11). However, the underlying mechanisms of
Cd-induced cancer are yet to be fully elucidated.
ERK, a member of the mitogen-activated protein
kinase (MAPK) family, is a key molecule that transmits signals from
surface receptors to the nucleus. It has the capacity to regulate
cell growth, survival, mitosis and differentiation. It has also
been demonstrated that ERK is involved in cell migration and
invasion in several types of cancers (12–16),
including lung cancer (17–24). Quintero Barceinas et al
(17) reported that all-trans
retinoic acid promoted growth, survival and migration in A549 lung
cancer cells by activating the ERK signaling pathway. Furthermore,
Li et al (18) reported that
angelicin inhibited human lung carcinoma A549 cell growth and
migration by regulating the ERK pathway. Zhang et al
(20) demonstrated that
transmembrane protein 17 decreased the invasion and metastasis of
lung cancer cells via the ERK signaling pathway.
Based on previous literature, the present study
hypothesized that p-ERK may serve an important role in the
migration and invasion of A549 cells induced by Cd. Therefore, the
aim of the current study was to explore the potential roles of
p-ERK in Cd-induced A549 cell migration and invasion to increase
understanding of the underlying molecular mechanisms and provide a
potential therapeutic target for lung cancer treatment.
Materials and methods
Cell culture and treatment
The human lung cancer A549 cell line was obtained
from the American Type Culture Collection (25). A549 cells were cultured in RPMI-1640
medium (Beijing Solarbio Science & Technology Co., Ltd.)
containing 10% fetal bovine serum (FBS) (GE Healthcare
Bio-Sciences), 100 U/ml penicillin and 100 µg/ml streptomycin in a
humidified incubator at 5% CO2 and 37°C. A549 cells were
cultured in serum-free RPMI-1640 medium for 24 h, followed by
treatment with different concentrations of Cd [cadmium chloride
hemi (pentahydrate); Shanghai Aladdin Bio-Chem Technology Co.,
Ltd.; 0, 0.1, 0.25, 0.5, 1, 2, 4 or 8 µM] for various durations
(0, 8, 24 or 48 h), as indicated. For certain experiments, 10
µM U0126 (Sigma-Aldrich; Merck KGaA), an inhibitor of mitogen
activated protein kinase (MEK)1/2 that is widely used to inhibit
ERK1/2 activation (26–29), was added to the cell culture 1 h
before Cd treatment.
MTT assay
A549 cells were harvested and seeded into 96-well
plates at a density of 1×104 cells per well. Following
incubation at 37°C for 24 h, the cells were treated with 0, 0.1,
0.25, 0.5, 1, 2, 4 or 8 µM Cd for 48 h at 37°C. MTT (10 µl; 5
mg/ml) was then added to each well and cells were incubated at 37°C
for 4 h. Purple formazan was dissolved in 150 µl of dimethyl
sulfoxide, and absorbance was measured on a microplate reader at
492 nm. The group treated with 0 µM Cd was regarded as the control.
Four replicate wells were used for each analysis.
Western blot analysis
Cells were collected and washed three times with
cold PBS. RIPA assay (Pierce; Thermo Fisher Scientific, Inc.)
solution containing protease and phosphatase inhibitors was
subsequently used to lyse cells, after which the protein content
was determined using a bicinchoninic acid assay. Proteins (30 µg)
were separated via SDS-PAGE on a 12% gel then transferred onto a
nitrocellulose membrane. The membrane was then blocked with 5%
bovine serum albumin in Tris-buffered saline and 0.1% Tween-20
(TBST) at room temperature for 1 h. Subsequently, the blots were
incubated at 4°C overnight with primary antibodies against
phosphorylated (p)-ERK1/2 (1:1,000; Thr202/Tyr204; cat. no. 4730S;
Cell Signaling Technology, Inc.), ERK1/2 (1:1,000; cat. no. 4695S;
Cell Signaling Technology, Inc.) and tubulin (1:1,000; cat. no.
sc-365791; Santa Cruz Biotechnology, Inc.). The blots were then
washed with TBST three times, followed by incubation with
horseradish peroxidase-conjugated secondary antibodies (Goat
anti-rabbit IgG; cat. no. ZB-2301; Goat anti-Mouse IgG; cat. no.
ZB-2305; each, OriGene Technologies, Inc.; each, 1:5,000) for 1 h
at room temperature. Finally, the protein bands were visualized
using an enhanced chemiluminescence reagent (cat. no. #1705060;
Bio-Rad Laboratories, Inc.) on a ChemiDoc™ XRS+ image analysis
system (Bio-Rad Laboratories, Inc.). Densitometry was performed
using Image J software v1.31 (National Institutes of Health).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from A549 cells using RNAiso
Plus (Takara Bio, Inc.) according to the manufacturer's protocol.
Following the quantification of total RNA concentrations, the
PrimeScript™ II 1st Strand cDNA Synthesis kit (Takara, Bio, Inc.)
was used to generate cDNA. A SYBR Green PCR master mix kit
(SYBR® Premix Ex Taq™-Tli RNaseH Plus; Takara,
Bio, Inc.) was used for qPCR. The thermocycling conditions were as
follows: 95°C for 30 sec; 40 cycles at 95°C for 5 sec, 60°C for 30
sec and 72°C for 30 sec. mRNA relative expression levels were
quantified using the 2−ΔΔCq method (30) and normalized to GAPDH. The following
primers were used: MMP2 forward, 5′-TGACATACATCTTTGCTGGAGAC-3′ and
reverse, 5′-GGCTTGCGAGGGAAGAAGTT-3′; GAPDH forward,
5′-TTCAGGTAATAGGCACCCTT-3′ and reverse,
5′-CTTCTCCATGGTGGTGAAGA-3′.
Cell migration assay
A migration assay was performed using Transwell
inserts. Cells were maintained at a concentration of
4×105 cells/ml in serum-free RPMI-1640. A total of 300
µl cell suspension was added into the upper chamber, whilst the
lower chamber was treated with 600 µl of RPMI-1640 with 10% FBS.
Following incubation at 37°C for 24 h, the medium was discarded and
non-migrating cells on the top surface of the upper chamber were
removed gently using cotton swabs. Migrated cells were fixed with
pre-chilled methanol for 30 min then stained with 0.5% crystal
violet at room temperature for 20 min. Representative images were
taken under an inverted microscope (magnification, ×10 and ×20)
equipped with a camera (Leica Microsystems GmbH). Subsequently, 33%
glacial acetic acid (200 µl) was added for 10 min to de-stain.
Absorbance was measured at 570 nm using a microplate reader.
Cell invasion assay
For the invasion assay, the upper chamber was
pre-coated with Matrigel. Ice-cold Matrigel was mixed with ice-cold
RPMI-1640 medium at a ratio of 1:1 and spread onto the upper
chamber (50 µl/chamber), which was subsequently incubated at 37°C
for 2 h. The following steps including cell plating, incubation,
fixing and de-staining were conducted as aforementioned for the
migration assay. Representative images were taken under an inverted
microscope (magnification, ×10 and ×20) equipped with a camera
(Leica Microsystems GmbH).
Statistical analysis
Statistical analysis was performed using SPSS v13.0
(SPSS, Inc.). Data were presented as the mean ± standard deviation
based on at least three replicates. The differences among groups
were analyzed using one-way analysis of variance followed by Least
Significant Difference post-hoc test. P<0.05 was considered to
indicate statistical significance.
Results
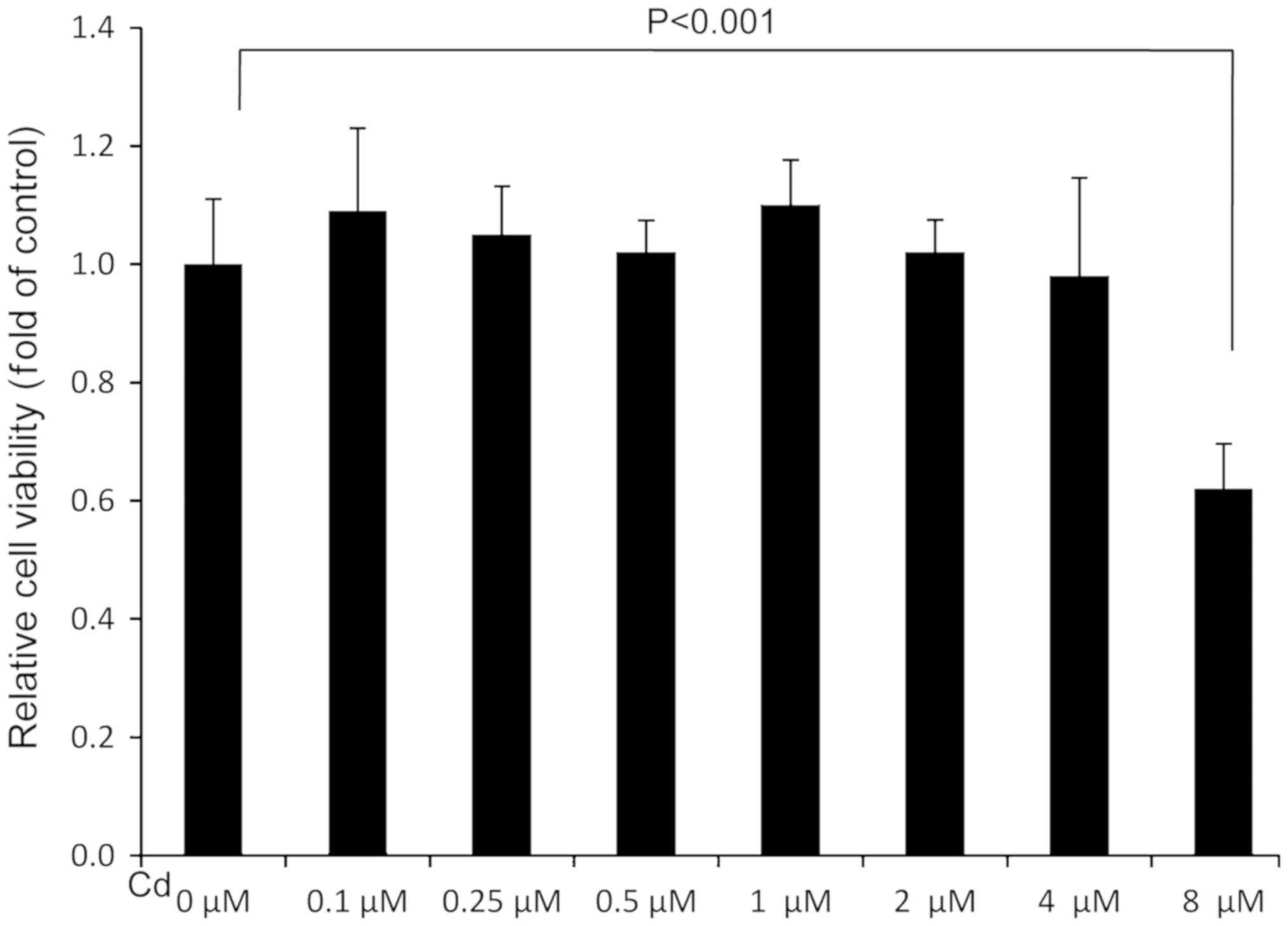
Cd treatment at the highest
concentration decreases A549 cell viability
To assess the effect of Cd treatment on cell
viability, an MTT assay was performed. The results revealed that
the relative cell viability of the 8 µM Cd treatment group was
significantly lower than the solvent control (0 µM), indicating
that 8 µM Cd had a marked cytotoxic effect (Fig. 1). Cd treatment demonstrated no marked
effect on cell viability when the concentration was <4 µM
(Fig. 1).
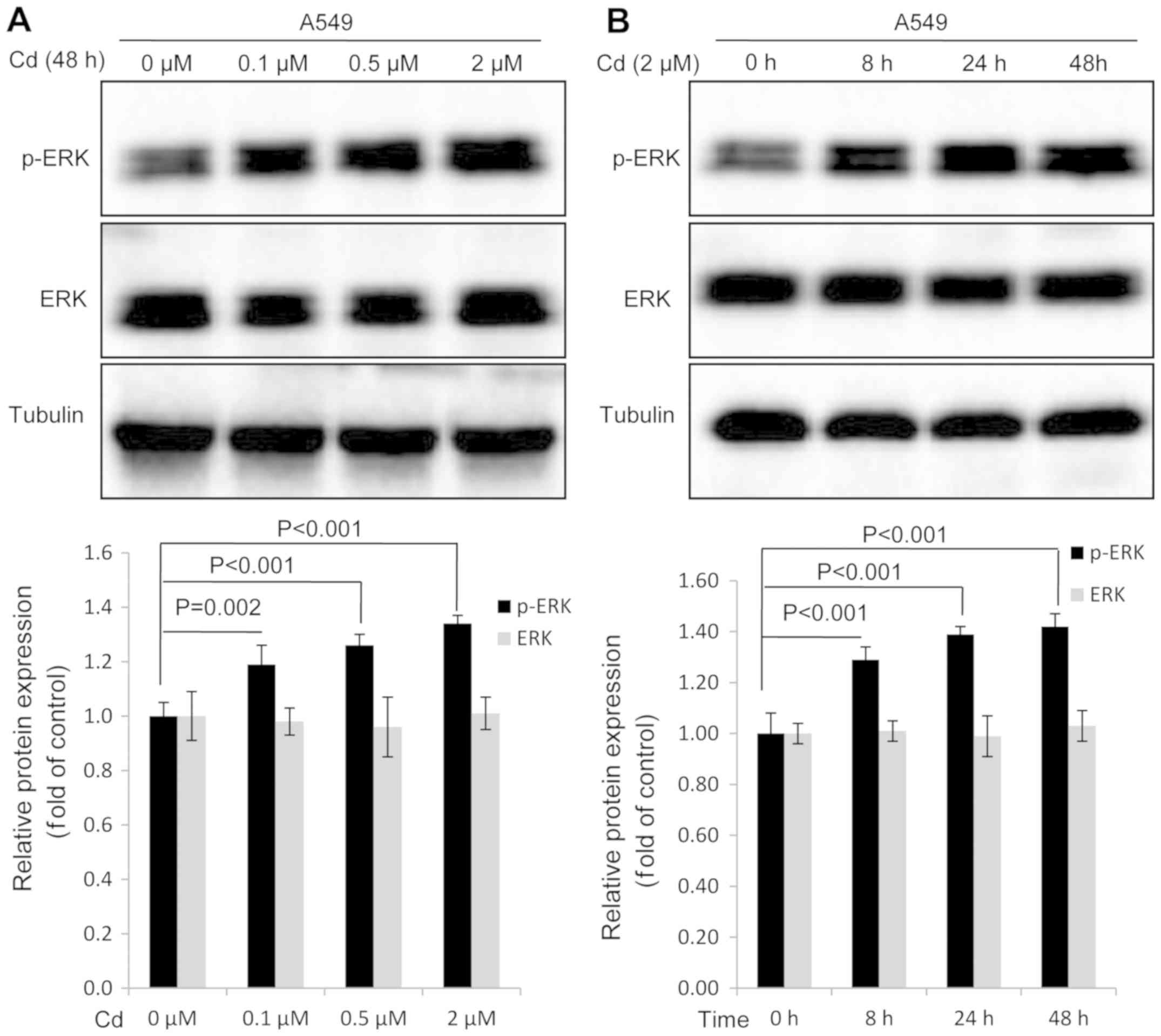
Cd treatment increases the expression
of p-ERK in A549 cells
Western blot analysis was conducted to investigate
whether Cd affected the expression of p-ERK in A549 cells. The
results revealed that the expression of p-ERK was markedly
increased in a dose-dependent manner when A549 cells were treated
with Cd for 48 h (Fig. 2A). When
compared with the 0 h group, a significant increase in p-ERK
expression was observed when A549 cells were treated with 2 µM Cd
for increasing time durations (Fig.
2B). Cd treatment exhibited no effect on the expression of
total ERK protein (Fig. 2). The
results indicated that ERK activation was induced by Cd
treatment.
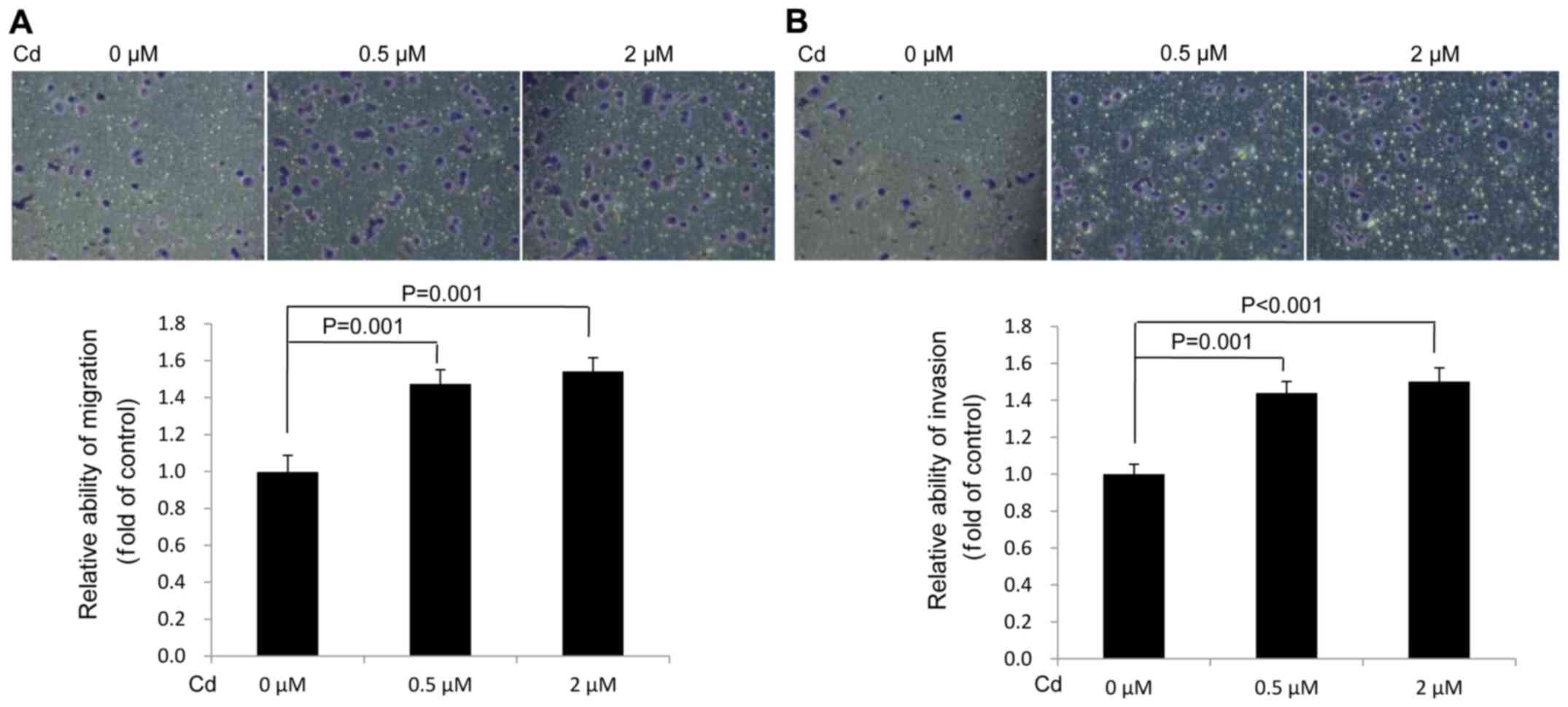
Cd accelerates the migration and
invasion of A549 cells
The data demonstrated that Cd treatment increased
A549 cell migration (Fig. 3A) and
invasion (Fig. 3B) compared with the
solvent control (0 µM). The results verified that Cd treatment
facilitated the migration and invasion of A549 cells.
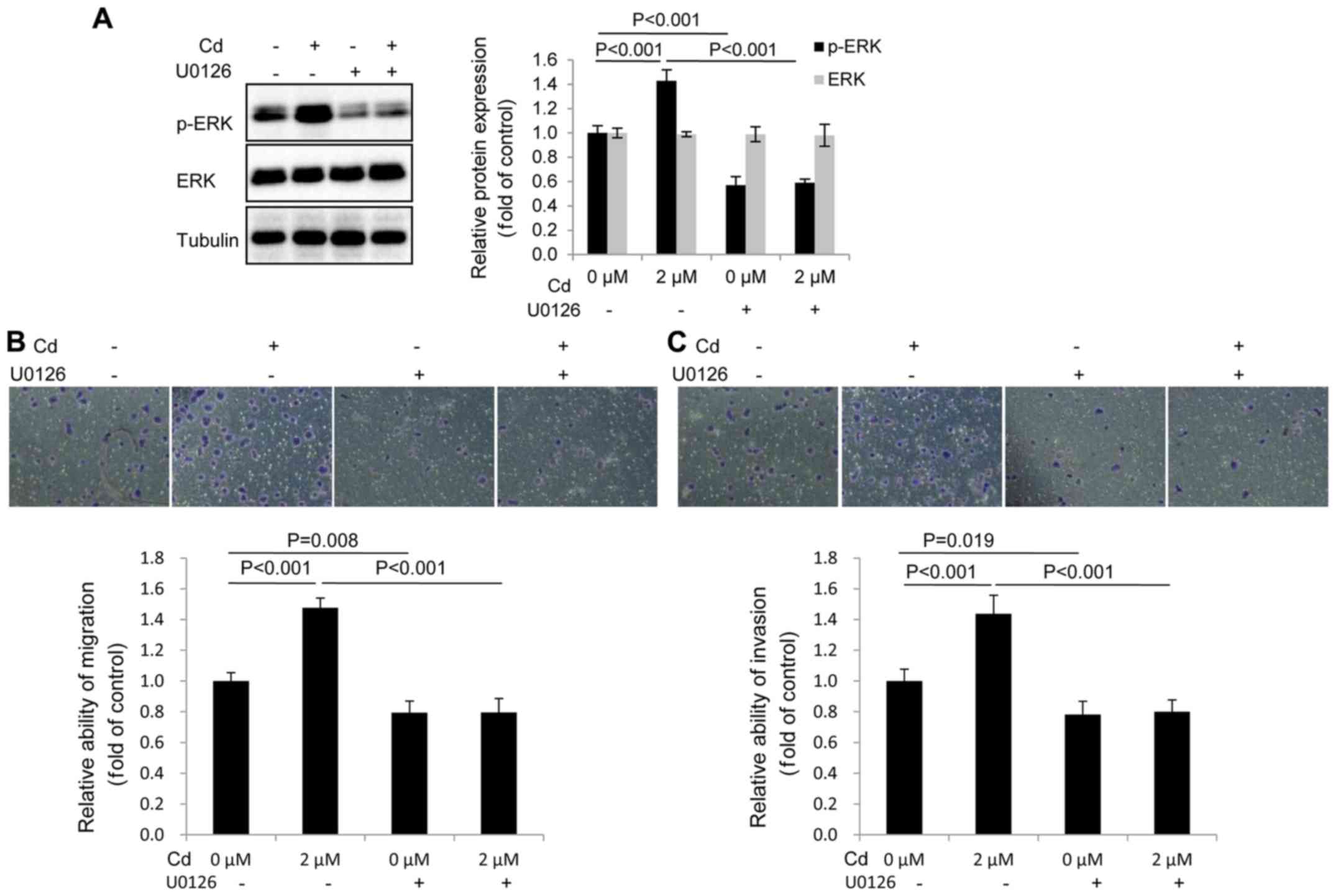
p-ERK has an important role in the
migration and invasion of A549 cells following Cd treatment
The migration and invasion of A549 cells were
assessed following pretreatment with 10 µM U0126 (an inhibitor of
MEK1/2), followed by 2 µM Cd treatment. The results revealed that
the expression of p-ERK was markedly decreased in the U0126-treated
group compared with the control group (Fig. 4A). In addition, U0126 blocked the
expression of p-ERK following Cd treatment. The data therefore
demonstrated that U0126 effectively inhibited the expression of
p-ERK (Fig. 4A). It was also
determined that U0126 inhibited the migration and invasion of A549
cells following Cd treatment (Fig. 4B
and C), indicating that elevated p-ERK activity was required
for Cd-induced migration and invasion.
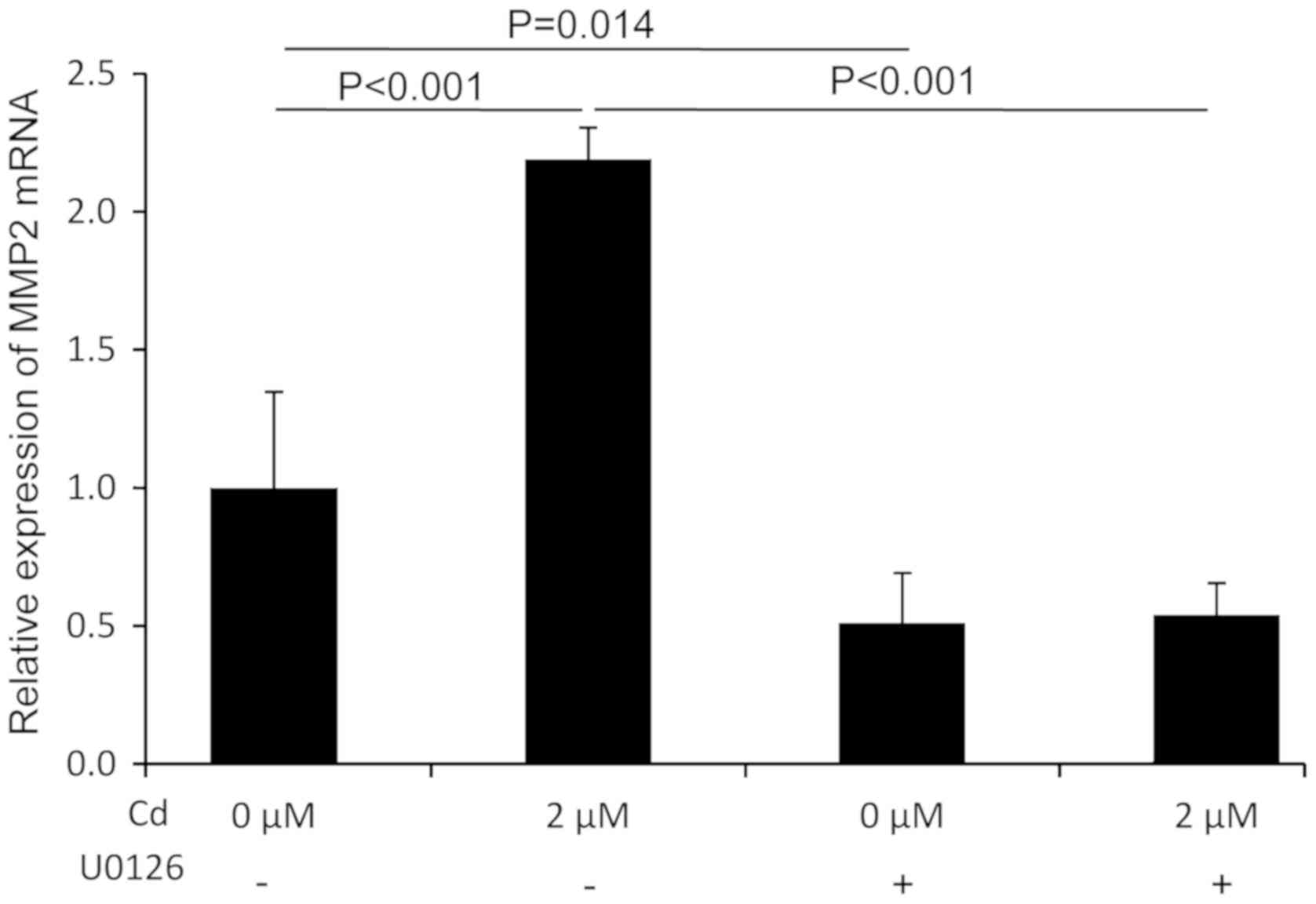
MMP2 mRNA expression following Cd
treatment is mediated by p-ERK
An investigation into whether MMP2 was a downstream
signaling molecule in the ERK pathway was performed using RT-qPCR.
The results revealed that Cd treatment significantly increased the
expression of MMP2 mRNA compared with the control group.
Furthermore, the Cd-induced expression of MMP2 mRNA was
significantly suppressed in the presence of U0126 (Fig. 5). These results indicated that MMP2
mRNA expression was modulated by p-ERK following Cd treatment and
also that MMP2 might be a downstream molecule in the ERK signaling
pathway.
Discussion
Cd is a widely used heavy metal environmental
pollutant. Previous studies have identified that exposure to Cd is
closely associated with the progression of various types of cancer.
He et al (31) reported that
blood Cd levels are positively associated with distant metastasis
and clinical stage of human breast cancer. Demir et al
(32) demonstrated that Cd levels in
tumor tissue are significantly correlated with lung cancer TNM
staging. Furthermore, Son et al (33) reported that chronic Cd treatment
enhanced cell migration and invasion in BEAS-2B cells and increased
tumor growth in a mouse xenograft model. Person et al
(7) also determined that chronic
Cd-treated lung cells exhibit a marked increase in invasion
compared with control cells. Wei and Shaikh (34) reported that increased cell migration
and invasion is demonstrated in prolonged Cd-treated
triple-negative breast cancer cells. Finally, Zhu et al
(12) demonstrated that migration
and invasion is induced in human follicular WRO and anaplastic FRO
thyroid cancer cells when treated with Cd at 250–1,000 nM.
Currently, three published articles have
demonstrated that exposure to Cd induces migration and invasion of
lung cancer cells. Luo et al (10) demonstrated that treatment with Cd
increased the expression of certain invasion-associated proteins in
the lung tissue of mice and in A549 cells, including MMP9, MMP2 and
p-protein tyrosine kinase 2. The study also revealed that
high-mobility group AT-hook (HMGA2) serves a significant role in
Cd-induced A549 cell migration and invasion (10). Lv et al (11) reported that Cd markedly enhances cell
proliferation, migration and invasion in lung cancer A549 cells. It
was also determined that reactive oxygen species-dependent
autophagy and related 4A cysteine peptidase upregulation-mediated
autophagy may affect Cd-induced cell growth, migration and
invasion. Fujiki et al (35)
demonstrated that prolonged Cd exposure induced
epithelial-mesenchymal transition (EMT), stress-fiber formation and
high cell motility in A549 cells, which was partially suppressed by
small interfering RNA-mediated Notch1 silencing. These studies
indicated that Cd promoted lung cancer cell migration and invasion
via different mechanisms.
ERK is activated on both the Thr202 and Tyr204
residues via sequential phosphorylation cascades. It also regulates
several downstream events, such as cell proliferation,
differentiation, motility and cell death (36–38). It
has been reported that the ERK signaling pathway is involved in
cell migration and invasion in several types of cancer cell. Peng
et al (39) reported that
anti-migration and anti-invasion activities of oxyfadichalcone C
were associated with the downregulation of the MAPK/ERK signaling
pathways in melanoma A375 cells. Wang et al (40) reported that the chrysin-induced
inhibition of proliferation, migration and invasion in glioblastoma
cells was mediated by the ERK/Nrf2 signaling pathway. Additionally,
Hong et al (41) reported
that microRNA-508 suppresses the EMT, migration and invasion of
ovarian cancer cells via the MAPK1/ERK signaling pathway. Li et
al (42) also reported that
microRNA-23a promoted cell growth and metastasis in gastric cancer
by targeting sprouty RTK signaling antagonist 2-mediated ERK
signaling. A previous study demonstrated that p-ERK serves an
important role in benzo(a)pyrene-induced Hep-G2 cell migration and
invasion (14).
MMP2, a member of the extracellular matrix degrading
proteinase family, is vital for the promotion of tumor metastasis.
Previous studies have determined that MMP2 is implicated in the
migration and invasion of lung cancer cells. Dong et al
(43) reported that the inhibition
of cell invasion occurs as a result of decreased MMP2 levels in
NSCLC cell lines. In addition, it has been reported that MMP2 is a
downstream molecule of the ERK pathway. For example, Wu et
al (44) determined that MMP2
was mediated by the ERK signaling pathway potentially via
microfibrillar-associated protein 5, which was associated with
tissue development and cancer progression. Furthermore, Li et
al (45) reported that the
activation of the ERK/NF-κB signaling pathway promoted the
expression of MMP2 and induced cell migration and invasion. Yu
et al (46) also reported
that andrographolide enhanced the anti-metastatic effect of
radiation in Ras-transformed cells by suppressing ERK-mediated MMP2
activity. Yang et al (47)
demonstrated that andrographolide suppressed the migratory ability
of human glioblastoma multiforme cells by targeting ERK1/2-mediated
MMP2 expression.
U0126 is a highly selective inhibitor of MEK1
(IC50, 72 nM) and MEK2 (IC50, 58 nM; a type
of MAPK/ERK kinase) (48), and is
also a weak inhibitor of proline rich transmembrane protein 2, Raf,
JNK, mitogen-activated protein kinase kinase kinase 1,
mitogen-activated protein kinase kinase (MKK) 3, MKK-4/EPH receptor
A4, MKK-6, cyclin-dependent kinase (Cdk)-2 and Cdk4. Micromolar
concentrations of U0126 were used to inhibit ERK1/2 activation in
previous studies, including 5 (29),
20 (49) and 40 µM (26). As the most commonly used U0126
concentration (27,29,50,51), 10
µM U0126 was selected in the present study to inhibit the
activation of ERK1/2.
The present study investigated the effects of Cd on
the migration and invasion of A549 cells as well as the potential
role of p-ERK in Cd-induced cell migration and invasion. The
results demonstrated that Cd-treated lung cancer A549 cell
migration and invasion increased compared with the control.
Migration and invasion were also decreased when cells were
pre-treated with U0126, an inhibitor of MEK1/2. Data also indicated
that p-ERK served an important role in Cd-induced A549 cell
migration and invasion. In addition, RT-qPCR determined that the
Cd-induced expression of MMP2 mRNA was significantly reduced in the
presence of U0126, which indicated that MMP2 might be a downstream
molecule in the ERK signaling pathway. Further investigation should
be performed to confirm the pattern of A549 cell MMP2 protein
expression and activity in the presence of Cd and U0126.
HMGA2 is a driver of tumor cell migration, invasion
and metastasis (52–54) and there is a proven link between
HMGA2 and p-ERK. Hawsawi et al (55) demonstrated that HMGA2 promoted EMT
via the MAPK/ERK signaling pathway in prostate cancer. Ping et
al (56) also reported that
angiotensin II type 2 receptor-interacting protein 3a presented
potential in suppressing the proliferation and aggressiveness of
ovarian carcinoma cells via the HMGA2-mediated ERK/EMT signaling
pathway. Additionally, Kao et al (57) indicated that Hsp90 indirectly
regulated HMGA2 by activating the ERK signaling pathway. HMGA2
upregulation is mediated by Cd-induced A549 cell migration and
invasion (10). The present study
determined that ERK activation was involved in Cd-induced A549 cell
migration and invasion. However, the link between HMGA2 and p-ERK
in Cd-induced A549 cell migration and invasion was not investigated
and will be the focus of future studies.
To the best of our knowledge, the present study
demonstrated for the first time that Cd induced A549 cell migration
and invasion by activating the ERK-MMP2 pathway. The results may
contribute to the further understanding of the molecular mechanisms
of Cd-induced lung-cancer cell migration and invasion, and provide
insight to potentially improve lung cancer treatment caused by
environmental Cd.
Acknowledgements
Not applicable.
Funding
The current study was funded by the National Natural
Science Foundation of China (grant no. U1404815) and Henan Province
University Science and Technology Innovation Talent Projects (grant
no. 17HASTIT045). The funder had no role in the study design, data
collection and analysis, decision to publish, or preparation of the
manuscript.
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
YW and HY designed and implemented the current
study, performed the experiments and analyzed the data. TP and HZ
performed western blotting and RT-qPCR analysis. YW, HZ and HW
performed the MTT, migration and invasion assays. HZ and YW wrote
the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nawrot TS, Martens DS, Hara A, Plusquin M,
Vangronsveld J, Roels HA and Staessen JA: Association of total
cancer and lung cancer with environmental exposure to cadmium: The
meta-analytical evidence. Cancer Causes Control. 26:1281–1288.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Verma M: Environmental and occupational
risk factors for lung cancer, methods of molecular biology. Cancer
Epidemiol. 472:3–23. 2009. View Article : Google Scholar
|
|
4
|
Garcia-Esquinas E, Pollan M, Tellez-Plaza
M, Francesconi KA, Goessler W, Guallar E, Umans JG, Yeh J, Best LG
and Navas-Acien A: Cadmium exposure and cancer mortality in a
prospective cohort: The strong heart study. Environ Health
Perspect. 122:363–370. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stayner L, Smith R, Thun M, Schnorr T and
Lemen R: A dose-response analysis and quantitative assessment of
lung cancer risk and occupational cadmium exposure. Ann Epidemiol.
2:177–194. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Meeting of the IARC working group on
beryllium, cadmium, mercury and exposures in the glass
manufacturing industry. Scand J Work Environ Health. 19:360–363.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Person RJ, Tokar EJ, Xu Y, Orihuela R,
Ngalame NN and Waalkes MP: Chronic cadmium exposure in vitro
induces cancer cell characteristics in human lung cells. Toxicol
Appl Pharmacol. 273:281–288. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huff J, Lunn RM, Waalkes MP, Tomatis L and
Infante PF: Cadmium-induced cancers in animals and in humans. Int J
Occup Environ Health. 13:202–212. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luevano J and Damodaran C: A review of
molecular events of cadmium-induced carcinogenesis. J Environ
Pathol Toxicol Oncol. 33:183–194. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Luo H, Li Z, Ge H, Mei D, Zhao L, Jiang L,
Geng C, Li Q, Yao X and Cao J: HMGA2 upregulation mediates
Cd-induced migration and invasion in A549 cells and in lung tissues
of mice. Chem Biol Interact. 277:1–7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lv W, Sui L, Yan X, Xie H, Jiang L, Geng
C, Li Q, Yao X, Kong Y and Cao J: ROS-dependent Atg4 upregulation
mediated autophagy plays an important role in Cd-induced
proliferation and invasion in A549 cells. Chem Biol Interact.
279:136–144. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhu P, Liao LY, Zhao TT, Mo XM, Chen GG
and Liu ZM: GPER/ERK&AKT/NF-κB pathway is involved in
cadmium-induced proliferation, invasion and migration of
GPER-positive thyroid cancer cells. Mol Cell Endocrinol. 442:68–80.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wei Y, Zhao L, He W, Yang J, Geng C, Chen
Y, Liu T, Chen H and Li Y: Benzo[a]pyrene promotes gastric cancer
cell proliferation and metastasis likely through the Aryl
hydrocarbon receptor and ERK-dependent induction of MMP9 and c-myc.
Int J Oncol. 49:2055–2063. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Y, Pan T, Li L, Wang H, Zhang D and
Yang H: Benzo(a)pyrene promotes Hep-G2 cell migration and invasion
by upregulating phosphorylated extracellular signal-regulated
kinase expression. Oncol Lett. 15:8325–8332. 2018.PubMed/NCBI
|
|
15
|
Sharma K, Singh J, Frost EE and Pillai PP:
MeCP2 overexpression inhibits proliferation, migration and invasion
of C6 glioma by modulating ERK signaling and gene expression.
Neurosci Lett. 674:42–48. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun Y, Lan M, Chen X, Dai Y, Zhao X, Wang
L, Zhao T, Li Y, Zhu J, Zhang X, et al: Anti-invasion and
anti-metastasis effects of Valjatrate E via reduction of matrix
metalloproteinases expression and suppression of MAPK/ERK signaling
pathway. Biomed Pharmacother. 104:817–824. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Quintero Barceinas RS, Garcia-Regalado A,
Arechaga- Ocampo E, Villegas-Sepulveda N and Gonzalez-De la Rosa
CH: All-Trans retinoic acid induces proliferation, survival, and
migration in A549 lung cancer cells by activating the ERK signaling
pathway through a transcription-independent mechanism. Biomed Res
Int. 2015:4043682015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li G, He Y, Yao J, Huang C, Song X, Deng
Y, Xie S, Ren J, Jin M and Liu H: Angelicin inhibits human lung
carcinoma A549 cell growth and migration through regulating JNK and
ERK pathways. Oncol Rep. 36:3504–3512. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tang F, Tang S, Guo X, Yang C and Jia K:
CT45A1 siRNA silencing suppresses the proliferation, metastasis and
invasion of lung cancer cells by downregulating the ERK/CREB
signaling pathway. Mol Med Rep. 16:6708–6714. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang X, Zhang Y, Miao Y, Zhou H, Jiang G
and Wang E: TMEM17 depresses invasion and metastasis in lung cancer
cells via ERK signaling pathway. Oncotarget. 8:70685–70694.
2017.PubMed/NCBI
|
|
21
|
Yang J, Kuang XR, Lv PT and Yan XX:
Thymoquinone inhibits proliferation and invasion of human
nonsmall-cell lung cancer cells via ERK pathway. Tumour Biol.
36:259–269. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liao YC, Shih YW, Chao CH, Lee XY and
Chiang TA: Involvement of the ERK signaling pathway in fisetin
reduces invasion and migration in the human lung cancer cell line
A549. J Agric Food Chem. 57:8933–8941. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shih YW, Wu PF, Lee YC, Shi MD and Chiang
TA: Myricetin suppresses invasion and migration of human lung
adenocarcinoma A549 cells: Possible mediation by blocking the ERK
signaling pathway. J Agric Food Chem. 57:3490–3499. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim JH, Cho EB, Lee J, Jung O, Ryu BJ, Kim
SH, Cho JY, Ryou C and Lee SY: Emetine inhibits migration and
invasion of human non-small-cell lung cancer cells via regulation
of ERK and p38 signaling pathways. Chem Biol Interact. 242:25–33.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Y, Wang H, Pan T, Li L, Li J and Yang
H: STIM1 silencing inhibits the migration and invasion of A549
cells. Mol Med Rep. 16:3283–3289. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu X, Price-Schiavi SA and Carraway KL:
Extracellular regulated kinase (ERK)-dependent regulation of
sialomucin complex/rat Muc4 in mammary epithelial cells. Oncogene.
19:4354–4361. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang X, Martindale JL and Holbrook NJ:
Requirement for ERK activation in cisplatin-induced apoptosis. J
Biol Chem. 275:39435–39443. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Silvany RE, Eliazer S, Wolff NC and Ilaria
RL Jr: Interference with the constitutive activation of ERK1 and
ERK2 impairs EWS/FLI-1-dependent transformation. Oncogene.
19:4523–4530. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Raja R, Lata S, Trivedi S and Banerjea AC:
Serum deprivation/starvation leads to reactivation of HIV-1 in
latently infected monocytes via activating ERK/JNK pathway. Sci
Rep. 8:144962018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
He Y, Peng L, Huang Y, Liu C, Zheng S and
Wu K: Blood cadmium levels associated with short distant
metastasis-free survival time in invasive breast cancer. Environ
Sci Pollut Res Int. 24:28055–28064. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Demir N, Enon S, Turksoy VA, Kayaalti Z,
Kaya S, Cangir AK, Soylemezoglu T and Savas I: Association of
cadmium but not arsenic levels in lung cancer tumor tissue with
smoking, histopathological type and stage. Asian Pac J Cancer Prev.
15:2965–2970. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Son YO, Wang L, Poyil P, Budhraja A,
Hitron JA, Zhang Z, Lee JC and Shi X: Cadmium induces
carcinogenesis in BEAS-2B cells through ROS-dependent activation of
PI3K/AKT/GSK-3β/β- catenin signaling. Toxicol Appl Pharmacol.
264:153–160. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wei Z and Shaikh ZA: Cadmium stimulates
metastasis-associated phenotype in triple-negative breast cancer
cells through integrin and β-catenin signaling. Toxicol Appl
Pharmacol. 328:70–80. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fujiki K, Inamura H, Miyayama T and
Matsuoka M: Involvement of Notch1 signaling in malignant
progression of A549 cells subjected to prolonged cadmium exposure.
J Biol Chem. 292:7942–7953. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Eblen ST: Extracellular-regulated kinases:
Signaling from ras to ERK substrates to control biological
outcomes. Adv Cancer Res. 138:99–142. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tanimura S and Takeda K: ERK signalling as
a regulator of cell motility. J Biochem. 162:145–154. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sun Y, Liu WZ, Liu T, Feng X, Yang N and
Zhou HF: Signaling pathway of MAPK/ERK in cell proliferation,
differentiation, migration, senescence and apoptosis. J Recept
Signal Transduct Res. 35:600–604. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Peng X, Wang Z, Liu Y, Peng X, Liu Y, Zhu
S, Zhang Z, Qiu Y, Jin M, Wang R, et al: Oxyfadichalcone C inhibits
melanoma A375 cell proliferation and metastasis via suppressing
PI3K/Akt and MAPK/ERK pathways. Life Sci. 206:35–44. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang J, Wang H, Sun K, Wang X, Pan H, Zhu
J, Ji X and Li X: Chrysin suppresses proliferation, migration, and
invasion in glioblastoma cell lines via mediating the ERK/Nrf2
signaling pathway. Drug Des Devel Ther. 12:721–733. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hong L, Wang Y, Chen W and Yang S:
MicroRNA-508 suppresses epithelial-mesenchymal transition,
migration, and invasion of ovarian cancer cells through the
MAPK1/ERK signaling pathway. J Cell Biochem. 119:7431–7440. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li Y, Chen H, She P, Chen T, Chen L, Yuan
J and Jiang B: microRNA-23a promotes cell growth and metastasis in
gastric cancer via targeting SPRY2-mediated ERK signaling. Oncol
Lett. 15:8433–8441. 2018.PubMed/NCBI
|
|
43
|
Dong QZ, Wang Y, Tang ZP, Fu L, Li QC,
Wang ED and Wang EH: Derlin-1 is overexpressed in non-small cell
lung cancer and promotes cancer cell invasion via EGFR-ERK-mediated
up-regulation of MMP-2 and MMP-9. Am J Pathol. 182:954–964. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wu Z, Wang T, Fang M, Huang W, Sun Z, Xiao
J and Yan W: MFAP5 promotes tumor progression and bone metastasis
by regulating ERK/MMP signaling pathways in breast cancer. Biochem
Biophys Res Commun. 498:495–501. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li X, Bao C, Ma Z, Xu B, Liu X, Ying X and
Zhang X: Perfluorooctanoic acid stimulates ovarian cancer cell
migration, invasion via ERK/NF-κB/MMP-2/-9 pathway. Toxicol Lett.
294:44–50. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yu CC, Chen CA, Fu SL, Lin HY, Lee MS,
Chiou WY, Su YC and Hung SK: Andrographolide enhances the
anti-metastatic effect of radiation in Ras-transformed cells via
suppression of ERK-mediated MMP-2 activity. PLoS One.
13:e02056662018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yang SL, Kuo FH, Chen PN, Hsieh YH, Yu NY,
Yang WE, Hsieh MJ and Yang SF: Andrographolide suppresses the
migratory ability of human glioblastoma multiforme cells by
targeting ERK1/2-mediated matrix metalloproteinase-2 expression.
Oncotarget. 8:105860–105872. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Favata MF, Horiuchi KY, Manos EJ, Daulerio
AJ, Stradley DA, Feeser WS, Van Dyk DE, Pitts WJ, Earl RA, Hobbs F,
et al: Identification of a novel inhibitor of mitogen-activated
protein kinase kinase. J Biol Chem. 273:18623–18632. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gao H, Zhang Y, Dong L, Qu XY, Tao LN,
Zhang YM, Zhai JH and Song YQ: Triptolide induces autophagy and
apoptosis through ERK activation in human breast cancer MCF-7
cells. Exp Ther Med. 15:3413–3419. 2018.PubMed/NCBI
|
|
50
|
Jiang X and Li H: Overexpression of LRIG1
regulates PTEN via MAPK/MEK signaling pathway in esophageal
squamous cell carcinoma. Exp Ther Med. 12:2045–2052. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang Y and Gong L: Palmitoleate inhibits
insulin transcription by activating the ERK1/2 pathway in rat
pancreatic beta-cells. Exp Ther Med. 13:2805–2811. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Morishita A, Zaidi MR, Mitoro A,
Sankarasharma D, Szabolcs M, Okada Y, D'Armiento J and Chada K:
HMGA2 is a driver of tumor metastasis. Cancer Res. 73:4289–4299.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gao X, Dai M, Li Q, Wang Z, Lu Y and Song
Z: HMGA2 regulates lung cancer proliferation and metastasis.
Thoracic Cancer. 8:501–510. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Dong J, Wang R, Ren G, Li X, Wang J, Sun
Y, Liang J, Nie Y, Wu K, Feng B, et al: HMGA2-FOXL2 axis regulates
metastases and epithelial-to-mesenchymal transition of
chemoresistant gastric cancer. Clin Cancer Res. 23:3461–3473. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hawsawi O, Henderson V, Burton LJ, Dougan
J, Nagappan P and Odero-Marah V: High mobility group A2 (HMGA2)
promotes EMT via MAPK pathway in prostate cancer. Biochem Biophys
Res Commun. 504:196–202. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ping H, Guo L, Xi J and Wang D:
Angiotensin II type 2 receptor-interacting protein 3a inhibits
ovarian carcinoma metastasis via the extracellular HMGA2-mediated
ERK/EMT pathway. Tumour Biol. 39:10104283177133892017.PubMed/NCBI
|
|
57
|
Kao CY, Yang PM, Wu MH, Huang CC, Lee YC
and Lee KH: Heat shock protein 90 is involved in the regulation of
HMGA2-driven growth and epithelial-to-mesenchymal transition of
colorectal cancer cells. PeerJ. 4:e16832016. View Article : Google Scholar : PubMed/NCBI
|