Introduction
Damage to neurons or glial cells and demyelization
after traumatic brain or spinal cord injuries, strokes and
neurodegenerative diseases may lead to dysfunction of the nervous
system. This phenomenon is linked with serious neurological
deficits in patients. The strategies developed for treating
injuries or diseases of the central nervous system include
pharmacological neuroprotection and tissue engineering. In the
latter method, living scaffolds composed of defined cells seeded on
biomaterial are implanted to the site of injury (1–3). Several
types of scaffolds are used for the treatment of nervous system
injuries (1,3,4).
Scaffolds composed of extracellular matrix molecules
are thought to be involved in the stimulation of neurogenesis
(1,5), a process linked to the functional
recovery of a damaged brain. Huang et al (2) report that the implantation of scaffolds
containing collagen (Col) and glycosaminoglycans (GAG) provides a
good microenvironment for neurogenesis. This data is supported by
studies that show scaffolds composed of Col only or of Col with
chondroitin sulphate (CS) constitute good environments for the
entrapment and cultivation of embryonic nerve cells (6–9). Col
scaffolds supplemented with GAG were found to decrease cell
adhesion but increase the proliferation of human mesenchymal stem
cells (10). Expression of the α2β1
integrin was confirmed in mesenchymal stem cells seeded within the
Col scaffold (11). This integrin is
supposed to participate in the adhesion of the cell to Col
fibers.
In the present study, three types of Col-based
scaffolds were evaluated: Col sponges, bi-component Col-CS matrices
crosslinked with 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide
hydrochloride (EDC) and Col sponges modified using 2,3 DAC. It has
been shown that binary systems of collagen-chondroitin sulphate
(Col-CS) and collagen-dialdehyde cellulose (Col-DAC) have the
correct balance of properties to serve as a biomimetic niche: They
accommodate human induced pluripotent stem cells (hiPSC-NPs)
sustaining their ability to proliferate and differentiate into
neural lineages (12).
The environments of these scaffolds allow the
entrapment of nerve embryonic cells and increase their metabolic
activity (1,6). The aim of the present study was to
observe the responses of the brain to different biomaterial
implantations (Col, Col-DAC, Col-CS). Both acellular and populated
embryonic nerve cell scaffolds were tested through implantation.
The longevity of such scaffolds, specifically whether or not they
were subject to rapid degradation in the brain, was examined. The
study focused on the survival of microtubule-associated protein 2
(MAP2)- and glial fibrillary acidic protein (GFAP)-positive cells
within the scaffolds; it also was evaluated the expression of
integrins β1 and α2 on embryonic nerve cells, and the involvement
of the α2β1 integrin in cell attachment to the scaffold.
Materials and methods
Insight into 3D collagen-based
scaffolds
Col type I was derived from well-purified porcine
tendons by pepsin digestion and acetic acid dissolution to prepare
a 0.7% (w/v) dispersion. Porcine tendons were bought directly from
the slaughterhouse which had proper procedures and permission to
obtain this material. Tendons were collected by the company
Euroimplant S.A., which was a producer of collagen suspension.
Tendons were taken from the lower limbs from fully mature pigs
weighting approximately 100 kg, and free from the transmission
factors of the infection. Tendons were wrapped in polypropylene
foil and frozen at −18±2°C until to the preparation of dispersion.
All experiments were conducted in accordance with national and EU
ethical regulations regarding animal free from infections. The use
of collagen isolated from porcine tendons was approved by the Local
Commission of Ethics in Lodz, Poland (Resolution 4/LB
505/2010).
Three-dimensional (3D) sponge-shaped porous
scaffolds were obtained from a dispersion containing Col and CS
(purchased from Sigma-Aldrich; Merck KGaA) at a ratio of 100:18.
For comparison, sponges from pure Col were also formed. The sponge
matrices were formed by freezing the Col dispersion or a mixture of
Col with CS and then lyophilized. To improve the functional
properties of the scaffold, all sponges were modified according to
the methods previously described by Pietrucha (13). In brief, the sponges were immersed in
60% ethanol solution containing 35 mM 1-ethyl-3-(3-dimethyl
aminopropyl) carbodiimide hydrochloride (EDC) and 12 mM
N-hydroxysuccinimide (NHS) (EDC and NHS supplied from
Sigma-Aldrich; Merck KGaA). Following the reaction, these products
were thoroughly washed with Na2HPO4, NaCl and
deionized water. To prepare spongy 3D scaffolds, the matrices were
frozen at −45°C and lyophilized at −55°C.
The synthesis of the Col-DAC scaffold was performed
by cross-linking of Col samples using 2,3-dialdehyde cellulose
(DAC). A detailed description of the method has been provided
previously (14–16). Briefly, 3D porous Col sponges were
incubated in a solution of DAC (synthesized in own scope) at 25°C
for 24 h (pH 8.0). The crosslinked products were thoroughly washed
with deionized water and then refrigerated at −40°C and lyophilized
at −55°C. All of the constructed sponges before implantation were
sterilized using an electron beam (accelerator ELU6-Linac) with
dose of 18 kGy.
Comprehensive descriptions of the biochemical,
spectroscopic, morphological and structural properties of the
modified Col-based scaffolds can be found in our earlier
publications (14–16). Some results of the physicochemical
characterization of 3D Col-based sponges are summarized in Table I.
 | Table I.Characteristics of physicochemical
properties of collagen-based sponges. |
Table I.
Characteristics of physicochemical
properties of collagen-based sponges.
| Parameters | Col
non-crosslinked | Col-CS crosslinked
by EDC/NHS | Col crosslinked by
DAC |
|---|
| Temperature
denaturation (°C) | 98.0 | 119.0 | 104.0 |
| Degree of
crosslinking (%) | No
crosslinking | 53±2 | 49.0 |
| Porosity (%) | 97.00 | 95.01 | 86.49 |
| Average pore
diameter (µm) | 55.40 | 31.05 | 32.09 |
Analysis of the quantitative measurements revealed
that both types of investigated scaffolds, i.e., Col-CS and
Col-DAC, had an average pore size of approximately 32 µm. However,
the thermal stability and degree of crosslinking of Col-DAC sponges
were found to be slightly lower than the Col-CS sponges crosslinked
with EDC.
Animals and study design
Thirty male Wistar rats weighing 250±30 g and five
female (220±20 g) were housed with free access to autoclaved
commercial food (Murigran, Motycz, Poland) as well as tap water
ad libitum. The animals were kept in light (L)-dark (D)
conditions (L:D=12:12). The light was switched on at 07.00. The
experiments were approved by the Local Commission of Ethics in
Lodz, Poland (permission number 37/ŁB617/2012). All experiments
were performed according to humane guidelines of legal act from
January 15th 2015, devoted to protection of animals used for
scientific or educational purposes. The rats were divided into
three groups:
Group 1: Rats implanted with Col scaffold
Group 2: Rats implanted with Col-DAC material
Group 3: Rats implanted with Col-CS scaffold
Each group consisted of two subgroups of five rats
each. The rats in the first subgroup were treated with an acellular
scaffold, while the respective living biomaterial scaffold
containing embryonic nerve cells was implanted to the rats in the
second subgroup.
Female pregnant rats at 17 days of gestation were
used for embryo brain isolation.
Scaffold implantation to the rat
brain
Prior to the scaffold implantation rat was
anesthetised by intraperitoneal injection of pentobarbital at the
dose of 50 mg/kg−1. After anesthesia, the rat was
installed in the stereotactic frame, shaved and decontaminated with
ethanol. A linear 2–3 cm long skin incision was made and a
craniotomy of the left frontal bone of the skull was performed. The
respective scaffold was implanted into the frontal lobe of the
brain. The skin wound was closed with four sutures. Four weeks
after implantation the brains were collected for histological
analysis.
Isolation and culture of embryonic
nerve tissue
Prior to surgery and embryo collection, the rat was
anesthetized by exposure to carbon dioxide (70%) and oxygen
mixture. The rats were anesthetized in chambers (25×48×23 cm). Gas
flow was 5.6 dm3/min. Then, the animals were euthanized
by the spinal cord dislocation. Death of euthanized animals was
confirmed by absence of respiration, heartbeat and corneal reflex.
Embryonic rats were euthanized by decapitation before brain
isolation. The embryos at day 17 of gestation were isolated from
the uterus of the pregnant female rat in sterile cold PBS (7,8). Once
removed from the embryos, the brains were rinsed with PBS, and the
meninges and blood vessels were excised. Then, the brains were
incubated in a solution consisting of collagenase (1 mg/ml) and
dispase (2 mg/ml) for five minutes at room temperature. This step
aimed to remove the connective tissue. The samples were then
incubated with trypsin for five minutes at 37°C. After trypsin
neutralization, the samples were centrifuged at 1,000 rpm.
Following this, the cells were suspended with 100 ml of medium
(MACS NeuroBrew-21; MiltenyBiotec) and triturated three to five
times gently. Cell suspension was seeded on laminin-covered dishes.
For cell cultures, the medium composed of MACS NeuroBrew-21,
gentamycin (25 µg/ml) and fungizone (2.5 µg/ml) was applied. The
cells were packaged on the scaffold at a density of
9×103 cells/sample in 96 well plates (7). After three days of cultivation, the
scaffolds were implanted into the brains.
The isolated cells were identified in the earlier
studies (7,8). The embryonic nerve cells were nestin
negative but they were morphologically differentiated (7,8).
Flow cytometry experiments
To confirm the differentiation of the isolated
embryonic nerve cells into neurons (MAP 2 positive cells) as well
as the expression of integrins, flow cytometry experiments were
performed. After fixation (Fixation Buffer, BDCytofix), the cells
were permeabilized by BD Phosflow Perm Buffer III. Following this,
the cells were washed and centrifuged (5 min, 1,000 rpm) and then
stained (4°C-8°C for 30 min) in Stain Buffer (BD Pharmingen). After
washing and centrifugation (for 10 min at 2,000 rpm) the cells were
treated with the following antibodies: mouse anti-MAP2 conjugated
with Alexa Fluor 488, mouse IgG1 k isotype control conjugated with
Alexa Fluor 488 (BD Pharmingen) Hamster IgG2l1 isotype control
conjugated with FITC, Hamster anti-rat CD49b conjugated with FITC
(BD Pharmingen), Arm Hamster IgG isotype control conjugated with
PE, anti-mouse/rat CD29 conjugated with PE (eBioscience). The cells
were then analyzed using a FACScan analytical flow cytometer
(Becton Dickinson) and the results were compared with the isotype
control.
Investigation of the involvement of
integrin α2β1 in cell entrapment within the collagenous
scaffolds
The nerve embryonic cells were applied into the Col
scaffolds seeded into the 96-well plates at a density of
9×103. The cells were incubated for 30 min at 37°C with
either the medium only, the medium containing 0.1% of DMSO (the
TC-I15 solvent) or the medium containing the TC-I15 inhibitor of
the α2β1 integrin (10−5 M). After 30 min, the samples
were triturated 30 times to wash away the unbound cells. After that
time, the cells were stained with bisbensimide (1 µg/ml). The cell
nuclei were counted under a fluorescent microscope at
magnification, ×100. Each investigated group contained eight
samples.
Histopathology
Following dissection, the brains were fixed in
formalin. For further evaluation, coronal sections of each brain
with grossly visible alternations (cavity) were selected. In cases
with no macroscopic changes, the whole brain was submitted for
standard tissue processing. Sections with a thickness of 3 µm were
cut from blocks of formalin-fixed paraffin-embedded tissue, mounted
on microscopic slides and submitted for standard hematoxylin and
eosin staining and immunohistochemistry.
Immunohistochemical staining was performed to
identify neurons and dendrites (microtubule associated protein 2 a,
b, c, MAP2) and to show the astrocytes (glial fibrillary acidic
protein, GFAP). For antigen retrieval and deparaffinization, the
slides were heated for 20 min at 97°C in EnVision Flex Target
Retrieval Solution (high pH; Dako) in the PT-link instrument
(Dako). For the GFAP immunostaining process, the pre-treated slides
were loaded into the autostainer instrument (EnVision System; Dako)
and stained with the FLEX polyclonal rabbit anti- GFAP ready-to-use
primary antibody (IR524; Dako). For the MAP2 immunostaining,
following pre-treatment, the slides were incubated with Peroxidase
Blocking Reagent (Dako) for five minutes, and then with the primary
monoclonal mouse anti-microtubule associated protein 2 (MAP2,
MA5-12826, 1:200; Thermo Fisher Scientific, Inc.) for 16 h at 4°C.
The MAP2 staining was visualised using the Dako Real EnVision
Detection System (Dako). After visualisation, the slides were
counterstained with hematoxylin, then dehydrated, cleared in xylene
and mounted.
Results
Histological evaluation
Collagen cross-linked with EDC implanted into the
brain. Among the sustained collagenous fibers of the scaffold, an
inflammatory infiltrate was composed of mononuclear cells
(macrophages and lymphocytes), multinucleated giant cells (foreign
body type macrophages) and a few polymorphonuclear cells (Fig. 1A). Moreover, clusters of the
meningeal-like cells were found within the scaffolds. Blood vessels
(Fig. 1B) were seen in some
scaffolds implanted into the brain. Neither MAP2-positive (Fig. 1F) nor GFAP-positive cells (Fig. 1E) were found within the scaffold.
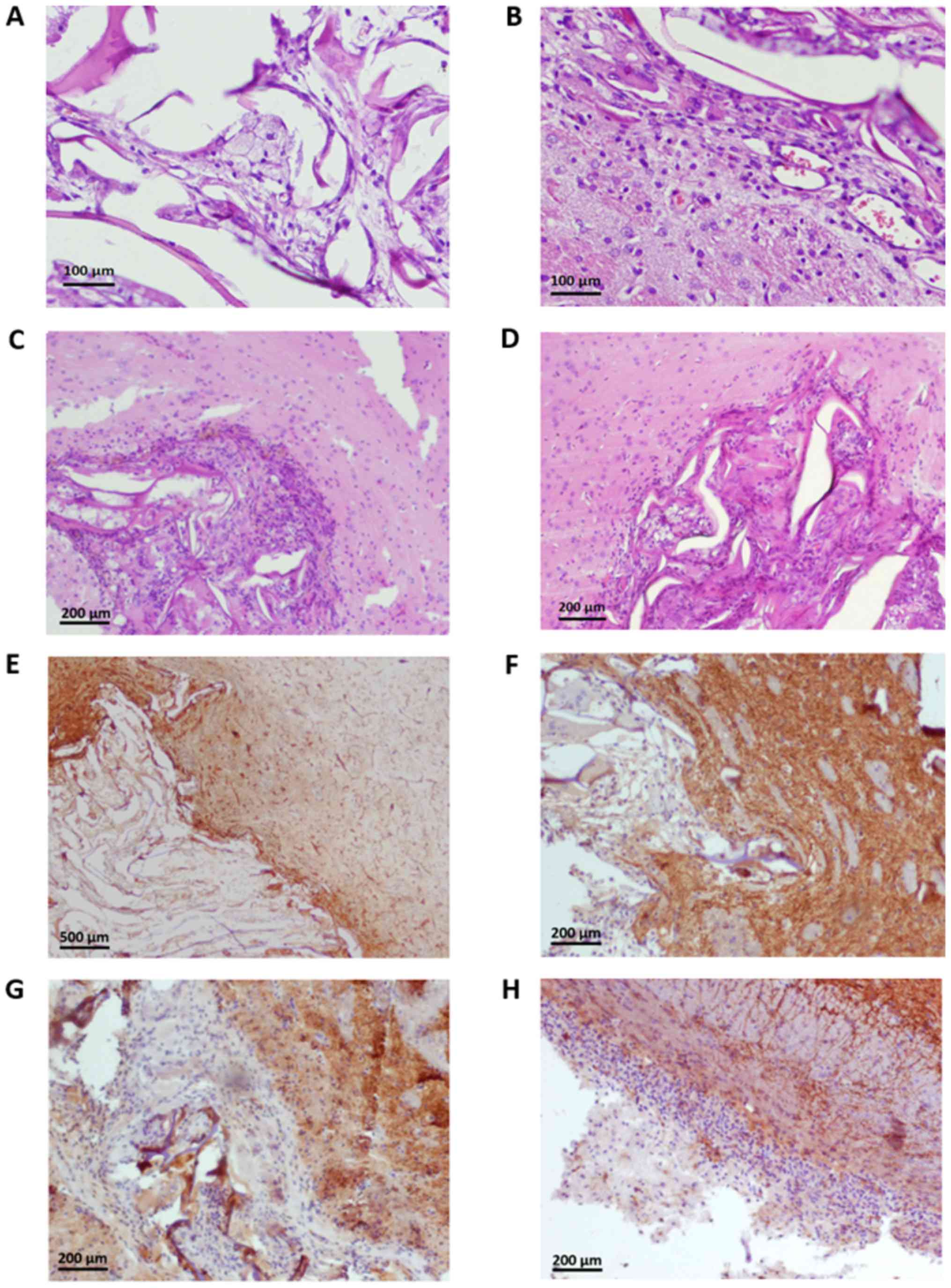 | Figure 1.(A) Col scaffolds grafted into the
brain. Among the sustained collagen fibers, multinucleated,
macrophages (cells with foamy cytoplasm) and mononuclear
inflammatory cells (lymphocytes) are observed. Staining with
hematoxylin and eosin. Magnification, ×200. (B) The brain adjacent
to the Col scaffolds. Multinucleated giant cells and thin-walled
blood vessels are present among the implanted collagen fibers
(right upper part of the figure). Mononuclear inflammatory cells
(lymphocytes) in the adjacent brain. Staining with hematoxylin and
eosin. Magnification, ×200. (C) The brain adjacent to the Col-DAC
scaffolds. The implanted Col-DAC scaffold (left lower part of the
figure) is infiltrated by multinucleated giant cells and
mononuclear cells (lymphocytes and macrophages). The reaction of
the adjacent nervous tissue to graft implantation is minimal and
comprises mononuclear cells (lymphocytes). Staining with
hematoxylin and eosin. Magnification, ×100. (D) The brain tissue
adjoined with the Col-CS scaffolds. The implanted Col-CS grafts
(right lower part of the figure) contain multinucleated giant cells
and mononuclear cells (lymphocytes and macrophages). The reaction
of the tissue to graft implantation is minimal and comprises
mononuclear cell infiltration. Staining with hematoxylin and eosin.
Magnification, ×100. (E) GFAP immunostaining of the brain adjacent
to the Col scaffolds. Cells present among the implanted collagen
fibers (left lower part of the figure) are GFAP-negative. The
positive GFAP reaction can be observed in the surrounding brain.
Magnification, ×40. (F) MAP2 immunostaining adjacent to the Col
scaffolds brain. The tissue surrounding the Col graft is normal and
indicated MAP2 immunoexpression. The mononuclear cells directly
adhered with the scaffold remnants and the multinucleated giant
cells within the graft are MAP2-negative. Magnification, ×100. (G)
MAP2 immunostaining of the brain adjacent to the Col-DAC scaffolds.
Cells directly adhered the Col-DAC scaffold and those situated
within the graft are MAP2 negative. The MAP2-positive reaction is
seen within the normal brain. Magnification, ×100. (H) MAP2
immunostaining of the brain adjacent to the Col-CS scaffolds. A
thin cell layer directly adhered the Col-CS scaffold and those
situated within the graft were revealed to be MAP2 negative. The
MAP2-positive reaction is observed within the encephalon. The
scaffold is not visible in the image. Magnification, ×100. Col,
collagen; Col-DAC, collagen-dialdehyde cellulose; Col-CS,
collagen-chondroitin sulphate; MAP2, microtubule-associated protein
2; GFAP, glial fibrillary acidic protein. |
In the adjacent area of the injury, brain
mononuclear cells (several lymphocytes, some hemosiderin-laden
macrophages and a few macrophages) and reactive astrocytes were
observed. Thick cerebral leptomeninges contained several
lymphocytes and macrophages, some of which were laden with
hemosiderin. In the deeper part of the brain, small basophilic
structures (dying neurons) were found (Fig. 1B). In the adjacent area, reactive
mononuclear cells (MAP2 negative) were observed, as well as an
increased number of GFAP positive (glial) cells (Fig. 1E).
Implantation of collagen cross-linked
by dialdehyde cellulose
Numerous cells were found to have infiltrated the
implant fibers; the inflammatory cells were thus comprised of
lymphocytes, multinucleated giant cells, macrophages and
neutrophils (Fig. 1C). Furthermore,
calcifications and eosinophilic structures were observed within the
scaffolds. Thin-walled blood vessels were shown in some implanted
scaffolds.
In the brain surrounding the implant, focal edema
and gliosis were observed. Moreover, hemosiderin deposits, amyloid
bodies, small calcifications, hemosiderin-laden macrophages and
dispersed infiltration of mononuclear cells (lymphocytes) were
detected (Fig. 1C). Fibrosis of the
cerebral leptomeninges was present. In the adjacent area of the
brain, a thin layer of MAP2 negative mononuclear cells was observed
(Fig. 1G).
Implantation of composite containing
collagen supplemented with CS cross-linked by EDC
Numerous cells were found among the fibers of the
implanted scaffold, these being mononuclear cells (macrophages,
hemosiderin-laden macrophages and lymphocytes) and multinucleated
giant cells (foreign body type' macrophages) (Fig. 1D). Thin-walled blood vessels were
seen among the collagen fibres of a few scaffolds implanted into
the brain. In the area adjacent to the implant layer, hyperaemia
was observed, as were small hemosiderin deposits and mononuclear
cells (hemosiderin-laden macrophages, macrophages and lymphocytes).
Moreover, cerebral leptomeninges, found adjacent to the cavity
containing the implant, showed features of fibrosis. In the deeper
parts of the brain area, small, oval, basophilic structures were
observed (Fig. 1D). In the adjacent
brain area, a thin layer of MAP2 negative mononuclear cells was
observed (Fig. 1H).
Examination of the adhesion of cells
to the scaffold
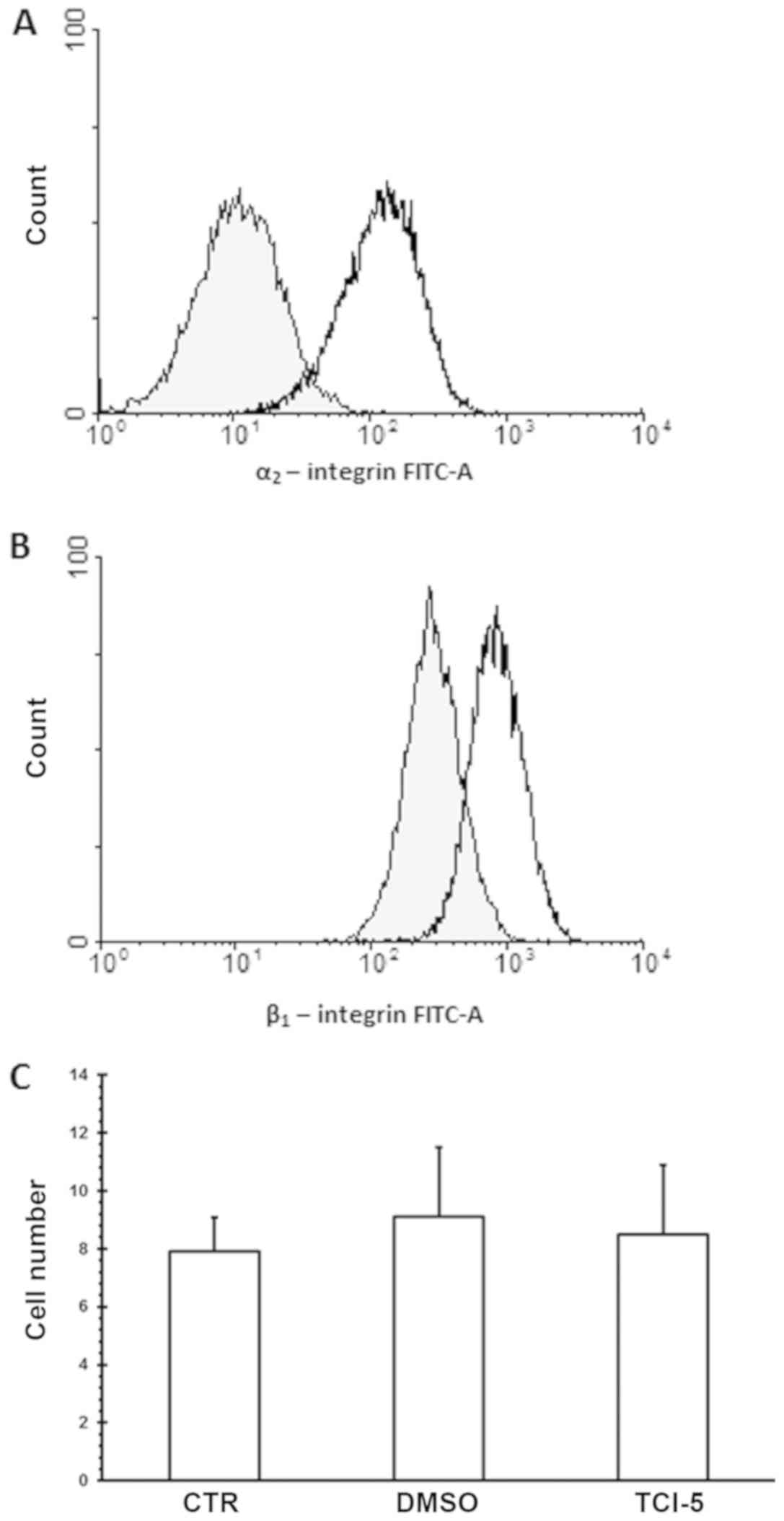
Expression of both integrin α2 (Fig. 2A) and integrin β1 (Fig. 2B) was detected on nerve embryonic
cells by flow cytometry studies. The same numbers of entrapped
cells were observed in the samples incubated with TC-I15 in the
collagenous scaffold (Fig. 2C), the
control group and the cells treated with TC-I15 solvent (0.1%
DMSO).
Discussion
Our results confirm the presence of Col fibers of
the inserted scaffold within the area of the brain injury, with the
morphological features of the scaffolds appearing to remain intact.
The scaffolds remained structurally intact for as long as four
weeks after implantation, suggesting that the material may serve as
a suitable template for cell migration. Moreover, the fibers of the
graft may be involved in the regulation of electrolytes and water
homeostasis of the tissue (17).
The implanted material induced no or minimal
inflammatory reaction in adjacent brain areas. The inflammatory
cells were found to accumulate around the scaffold as well as
infiltrated into the biomaterial (Fig.
1A-D). Implantation of the scaffolds induced granulomatous
tissue reaction comprising an accumulation of multinucleated
macrophages, mononuclear inflammatory cells and formation of blood
vessels. These results are comparable with other studies. The
inflammatory reaction was seen to be calmed by implantation of
gelatin-laminin materials (18),
while Vaysse et al did not find any inflammatory reaction
three months after implantation of micropatterned
polydimethylsiloxane with neuronal cells, with the graft remaining
intact (19). No excessive
inflammatory reaction was found to be associated with any of the
three types of applied implants during the present study (Fig. 1A-D).
Vascularization is a critical and important event in
tissue engineering. In all three types of tested biomaterials,
thin-walled blood vessels were found to be situated within the
scaffolds. These results suggest that Col alone, independent of the
cross-linking method, may support new blood vessel formation.
Hence, even without addition of angiogenic factors, the tested
samples constituted a suitable niche for promoting local
angiogenesis. Scaffolds with angiogenic properties are recommended
in the treatment of ischemic brain lesions (20). Some reports suggest a relationship
between neurogenesis and angiogenesis in brain lesions (21). Huang et al (5) report increased angiogenesis after
Col-GAG scaffold implantation into the brain; however, the types of
GAGs forming the scaffolds were not defined. Elsewhere,
implantation of such as scaffold into the brain lesion increased
proliferation of both smooth muscle cells and endothelial cells
(22). These phenomena were
accompanied with an increased concentration of angiogenic growth
factors (VEGF, FGF2, PDGF) in the tissue (2,5). To
fully clarify the angiogenic properties of the scaffolds, further
studies should employ immunohistochemical staining of CD 44.
Little development of the glial scar at the graft
periphery was observed, but no GFAP positive cells (cells with
glial differentiation) were found inside the scaffold (Fig. 1E); in addition, no excessive gliosis
was observed after any scaffold implantation. These results are
supported by those of experiments performed on other scaffolds
(2,18,23).
After implantation of the Col-GAGs scaffold into the brain,
astrocite accumulation was reported only in the lesion boundary
zone, and these cells did not proliferate (2). Jurga et al found that astrocites
mainly accumulated at the edges of the scaffolds (18). Although infiltration of astrocite
microglial and endothelial cells was reported following the
introduction of a gelatin-siloxane biomaterial implant into the
brain, no infiltration of neurons was reported (19).
Flow cytometry studies have found that the embryonic
nerve cells seeded within the scaffolds contained cells
differentiating into neurons (MAP2-positive cells) (7,8); these
are able to replace the damaged nervous tissue at the site of
trauma, and are hence, crucial for tissue reconstruction. Scaffolds
populated with embryonic nerve cells are also believed to be a
source of cytokines that may accelerate the regeneration process.
However, the clinical effects of cell-containing scaffold
implantations into the brain are very limited due to low cell
survival rate: the cells that populate the scaffolds usually die a
few days after implantation. The main causes of cell death are
inflammation, immune processing, oxidative stress and low levels of
trophic factors (23,24). The main mechanisms of neuron death
are necrosis and apoptosis (25).
Sortwell et al (26) found
that the critical period for apoptotic death is the first four days
post-implantation. In the present study, no MAP2-positive cells
were found within any scaffold at four weeks after implantation
(Fig. 1F-H). Furthermore, no
additional effects with regard to brain injury regeneration were
observed in the cell-seeded scaffold compared with the acellular
scaffold. This lack of difference may be linked to fact that these
cells populated within the scaffolds are believed to die just after
implantation.
Our present observations regarding cell survival
reflect a problem described earlier in the literature (25,26).
Although all tested scaffolds were well tolerated by injured brain
tissue, they were unable to shelter seeded embryonic nerve cells
from death. Applied scaffolds cannot support the survival of the
embryonic nerve cells or the ingrowth of MAP2- or GFAP-positive
cells from adjacent brain. Moreover, observations in vivo
revealed that meningeal cells and blood vessels may grow into the
scaffolds. Thus, the additional modification of the sponges are
postulated to create a niche supporting embryonic nerve cell
survival in vivo.
Some earlier studies were performed to protect the
cells populated within the scaffold from death. Marchionini et
al (27) tested the effects of
caspase inhibitors in protecting tyrosine hydroxylase
immunoreactive neurons from apoptosis, but found neuron survival
not to be prolonged. Despite this, a recent study by Vaysse et
al did achieve neuron survival following the implantation of
micropatterned polydimethylsiloxane biomaterial with neuronal cells
into the brain. This improvement in the neuronal cell survival
resulted in the addition of polylysine and laminin to the graft
(19). These compounds are believed
to reduce the traumatism of implantation. All grafts used in our
study displayed beneficial effects for embryonic nerve cell growth,
and have previously been reported in in vitro experiments
(7,8). However, the tested samples were unable
to improve the survival of the cells within the grafts after
implantation into the brain.
The interaction of the Col with embryonic nerve
cells may influence both the differentiation and the migration of
the cells (18,24,28,29). Our
findings are the first to demonstrate the expression of both α2 and
β1 integrins on the surface of the embryonic nerve cells (Fig. 2A and B). Integrin α2β1 is believed to
be involved in the interaction between mesenchymal stem cells and
Col scaffolds (11). The α2β1
integrin inhibitor TCI-15 did not modify cell entrapment within the
scaffolds. Although our findings fail to confirm the involvement of
α2β1 integrin in binding of collagen scaffolds, they nevertheless
suggest that other receptors for extracellular collagen (integrins
α1β1, α10β1 α11β1 and Discoidin Domain Receptors) should be
considered as potential targets for the interaction with
extracellular collagen.
Our findings confirm that the tested samples of Col,
either alone, with CS cross-linked by EDC or modified by DAC, are
well tolerated by rat tissue and may be implanted into the brain.
During the duration of the experiment, the Col fibers remained
intact and hence may support cell migration and development. None
of the tested biomaterials induced large glial scar formation.
Moreover, no additional effects were seen when scaffolds seeded
with embryonic nerve cells were implanted compared to the acellular
grafts. This may be due to the possible death of all embryonic
nerve cells after implantation of the scaffold. As our results
show, the tested biomaterials are worth considering in the
treatment of brain injury; however, additional modifications of the
material are postulated to improve survival of the cells in
vivo. This paper also confirmed the expression of integrins α2
and β1 on nerve embryonic cells.
Acknowledgements
The authors would like to thank Mrs Teresa
Staszewska (Department of Behavioral Pathophysiology, Medical
University of Lodz, ul. Zeligowskiego 9/7, 90–752 Lodz, Poland) for
her technical assistance.
Funding
The present study was supported by National Science
Centre (grant no. DEC-2011/03/B/ST8/05867). Publication of the
present study was supported by the Medical University of Lodz
(grant no. 503/6-103-02/503-061-001-18).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JD and KP contributed to the research concept and
design of the current study. JD, KJ, LP, BP, JS and MS collected
and assembled the data. JD, KJ, MS and JS analyzed and interpreted
the data. JD, KP and KJ wrote the manuscript. JD, KP, KJ, LP, BP,
MS and JS critically revised the article. JD, KP, KJ, LP, BP, JS
and MS approved the article.
Ethical approval and consent to
participate
The experiments were approved by the Local
Commission of Ethics in Lodz.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chwalek K, Tang-Schomer MD, Omenetto FG
and Kaplan DL: In vitro bioengineered model of cortical brain
tissue. Nat Protoc. 10:1362–1373. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huang KF, Hsu WC, Chiu WT and Wang JY:
Functional improvement and neurogenesis after collagen-GAG matrix
implantation into surgical brain trauma. Biomaterials.
33:2067–2075. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Teng YD, Lavik EB, Qu X, Park KI, Ourednik
J, Zurakowski D, Langer R and Snyder EY: Functional recovery
following traumatic spinal cord injury mediated by unique polymer
scaffold seeded with neural stem cells. Proc Natl Acad Sci USA.
99:3024–3029. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kowalska-Ludwicka K, Cala J, Grobelski B,
Sygut D, Jesionek-Kupnicka D, Kolodziejczyk M, Bielecki S and
Pasieka Z: Modified bacterial cellulose tubes for regeneration of
damaged peripheral nerves. Arch Med Sci. 9:527–534. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang KF, Hsu WC, Hsiao JK, Chen GS and
Wang JY: Collagen-Glycosaminoglycan Matrix implantation promotes
angiogenesis following surgical brain trauma. BioMed Res Int.
2014:6724092014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bandtlow CE and Zimmerman DR:
Proteoglycans in the developing brain: New conceptual insights for
old proteins. Physiol Rev. 80:1267–1290. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Drobnik J, Pietrucha K, Piera L, Szymański
J and Szczepanowska A: Collagenous scaffolds supplemented with
hyaluronic acid and chondroitin sulfate used for wound fibroblast
and embryonic nerve cell culture. Adv Clin Exp Med. 26:223–230.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pietrucha K, Szymański J and Drobnik J:
The behavior of embryonic neural cells within the 3D
micro-structured collagen-based scaffolds. IFMBE Proceedings.
45:549–552. 2015. View Article : Google Scholar
|
|
9
|
Sirko S, von Holst A, Wizenmann A, Gotz M
and Faissner A: Chondroitin sulphate glycosaminoglycans control
proliferation, radial glia cell differentiation and neurogenesis in
neural stem/progenitor cells. Development. 134:2727–2738. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li YY, Choy TH, Ho FC and Chan PB:
Scaffold composition affects cytoskeleton organization cell-matrix
interaction and the cellular fate of human mesenchymal stem cells
upon chondrogenic differentiation. Biomaterials. 52:208–220. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Milner R and Campbel IL: The integrin
family of cell adhesion molecules has multiple functions within
CNS. J Neurosci Res. 69:286–291. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pietrucha K, Zychowicz M, Podobinska M and
Buzanska L: Functional properties of different collagen scaffolds
to create a biomimetic niche for neurally committed human induced
pluripotent stem cells (iPSC). Folia Neuropathol. 55:110–123. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pietrucha K: Physicochemical properties of
3D collagen-CS scaffolds for potential use in neural tissue
engineering. Int J Biol Macromol. 80:732–739. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pietrucha K: Development of collagen
cross-linked with dialdehyde cellulose as a potential 3D scaffold
for neural tissue engineering. IFMBE Proceeding. 45:349–352. 2015.
View Article : Google Scholar
|
|
15
|
Pietrucha K and Safandowska M: Dialdehyde
cellulose-crosslinked collagen and its physicochemical properties.
Process Biochem. 50:2105–2111. 2015. View Article : Google Scholar
|
|
16
|
Pietrucha K, Marzec E and Kudzin M: Pore
structure and dielectric behaviour of the 3D collagen-DAC scaffolds
designed for nerve tissue repair. Int J Biol Macromol.
92:1298–1306. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rauch U: Extracellular matrix components
associated with remodeling process in brain. Cell Mol Life Sci.
61:2031–2045. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jurga M, Dainiak MB, Sarnowska A,
Jablonska A, Thripathi A, Plieva FM, Savina IN, Strojek L, Jungvid
H, Kumar A, et al: The performance of laminin-containing cryogel
scaffolds in neural tissue regeneration. Biomaterials.
32:3423–3434. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vaysse L, Beduer A, Sol JC, Vieu C and
Loubinoux I: Micropatterned bioimplant with guided neuronal cells
to promote tissue reconstruction and improve functional recovery
after primary motor cortex insult. Biomaterials. 58:46–53. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ju R, Wen Y, Gou R, Wang Y and Xu Q: The
experimental therapy on brain ischemia by improvement of local
angiogenesis with tissue engineering in the mouse. Cell Transplant.
23 (Suppl 1):S83–S95. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu S, Yao S, Wen Y, Wang Y, Wang H and Xu
Q: Angiogenic microspheres promote neural regeneration and motor
function recovery after spinal cord injury in rats. Sci Rep.
6:334282016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tapon-Bretaudière J, Drouet B, Matou S,
Mourão PA, Bros A, Letourner D and Fischer AM: Modulation of
vascular human endothelial rat smooth muscle cell growth by a
fucosylated chondroitin sulphate from echinoderm. Thromb Haemost.
84:332–337. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Deguchi K, Tsuru K, Hayashi T, Takaishi M,
Nagahara M, Nagotani S, Sehara Y, Jin G, Zhang H, Hayakawa S, et
al: Implantation of the new porous gelatin-siloxane hybrid into
brain lesions as a potential scaffod for tissue regeneration. J
Cereb Blood Flow Metab. 26:1263–1273. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Summitomo S, Muramatsu R, Fujii S and
Yamashita T: Vascular endothelial cells promote cortical neurite
outgrowth via an integrin β3-dependent mechanism. Biochem Biophys
Res Commun. 450:593–597. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Emgard M, Hallin U, Karlsson J, Bahr BA,
Brundin P and Blomgren K: Both apoptosis and necrosis occur early
after intracerebral grafting of ventral mesencephalic tissue: A
role for protease activation. J Neurochem. 86:1223–1232. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sortwell CE, Pitzer MR and Collier TJ:
Time course of apoptotic cell death within mesencephalic cell
suspension grafts: Implications for improving grafted dopamine
neuron survival. Exp Neurol. 165:268–277. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Marchionini DM, Collier TJ, Pitzer MR and
Sortwell CE: Reassessment of caspase inhibition to augment grafted
dopamine neuron survival. Cell Transplant. 13:273–282. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lanfer B, Hermann A, Kirsh M, Freudenberg
U, Reuner U, Werner C and Storch A: Directed growth of adult human
white matter stem cell-derived neurons on aligned fibrillar
collagen. Tissue Eng Part A. 16:1103–1113. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yoshinaga T, Hashimoto E, Ukai W, Ishii T,
Shirasaka T, Kigawa Y, Tateno M, Kaneta H, Watanabe K, Igarashi T,
et al: Effects of atelocollagen on neural stem cell function and
its migrating capacity into brain in psychiatric disease model. J
Neural Transm (Vienna). 120:1491–1498. 2013. View Article : Google Scholar : PubMed/NCBI
|