Introduction
Polycystic ovary syndrome (PCOS) is the most common
endocrine disorder in women of reproductive age and is the leading
cause of female infertility (1).
Although the clinical and biochemical characteristics of PCOS are
heterogeneous, abnormal folliculogenesis is still considered an
important clinical feature of PCOS (2). During follicular development, a large
number of follicles undergo atresia, a process tightly controlled
by the fine balance between survival and apoptotic factors
(3,4). The process is also regulated by
endocrine, autocrine and paracrine factors (5). Normal follicle development depends on
the balance between proliferation and apoptosis. Alterations in the
ovarian microenvironment that present follicular cysts could alter
the normal processes of ovarian cell proliferation and programmed
cell death leading to a variety of fertility problems including
PCOS (6,7). It has been speculated that abnormal
folliculogenesis and degeneration of the granulosa cell layers is
caused by the abnormal proliferation and/or apoptosis of follicular
granulosa cells, resulting in failure in further development
(8,9). However, the mechanism underlying
abnormal folliculogenesis is not fully understood.
MicroRNAs (miRNAs) are highly conserved, ~18–25
nucleotide non-coding RNA molecules that post-transcriptionally
regulate mRNA expression by binding to their 3′untranslated regions
(UTRs) (10). miRNAs have been
implicated in various biological and cellular processes including
cell proliferation, differentiation and apoptosis (11,12).
Evidence has demonstrated that certain miRNAs are involved in the
regulation of ovarian granulosa cell proliferation and apoptosis
(13,14). miR-30d-5p has been studied in several
cancers including cervical cancer, non-small cell lung cancer,
prostate cancer, gallbladder carcinoma and human colon cancer
(15–19). miR-30d-5p has also been reported to
serve critical roles in acute ischemic stroke-induced,
autophagy-mediated brain injury (20). A previous study reported that
miR-30d-5p is significantly increased during follicle stimulating
hormone (FSH)-mediated progesterone secretion of cultured granulosa
cells (21). However, the effect of
miR-30d-5p on ovarian granulosa cell apoptosis and its potential
mechanism has not been fully elucidated.
The transforming growth factor-β signaling pathway
participates in various cellular processes, including cell growth,
differentiation, apoptosis and homeostasis, and is mediated by a
complex of membrane-bound type I and type II receptors with Smad
proteins functioning as intracellular mediators (22,23).
Smad2 belongs to the receptor-activated Smad family and it serves a
key role in regulating cell proliferation and apoptosis (24). Abnormal proliferation and/or
apoptosis in granulosa cells have an important role in PCOS
(8,9). Therefore, Smad2 may be crucial to PCOS.
To the best of our knowledge, the role of Smad2 in POCS remains
largely unclear with the relationship between miR-30d-5p and Smad2
being unknown.
The present study investigated the role of
miR-30d-5p in ovarian granulosa cell proliferation and apoptosis to
elucidate the underlying molecular mechanisms and to reveal the
role of miR-30d-5p in PCOS. The results of this study indicated
that miR-30d-5p may be a new therapeutic target for the treatment
of PCOS.
Materials and methods
Cell culture
Rat ovarian granulosa cells (cat. no. CC-R050) were
purchased from Shanghai Mingjin Biotechnology Co., Ltd. Rat
granulosa cells were then cultured in DMEM/Ham's nutrient mixture
F-12 medium (Gibco; Thermo Fisher Scientific, Inc.) containing 10%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and 100
U/ml penicillin and 100 mg/ml streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) in a humidified atmosphere containing 5%
CO2 at 37°C.
Cell transfection
miR-30d-5p mimic (cat. no. miR10000461-1-5) or the
negative control (NC) of the miR-30d-5p mimic (NC scrambled
miR-30d-5p mimic; cat. no. miR01201-1-5) were purchased from
Guangzhou RiboBio Co., Ltd. Rat ovarian granulosa cells
(5×104 cells per well) were cultured in six-well plates
overnight at 37°C prior to transfection. Cells were then
transfected with 1 µg Smad2-plasmid (cat. no. sc-421525-ACT; Santa
Cruz Biotechnology, Inc.), 1 µg control plasmid (cat. no.
sc-437275; Santa Cruz Biotechnology, Inc.), 100 nM miR-30d-5p
mimics, 100 nM NC miR-30d-5p mimics, 100 nM miR-30d-5p mimics + 1
µg control plasmid (Mimics + plasmid) or 100 nM miR-30d-5p mimics +
1 µg Smad2-plasmid (Mimics + Smad2 plasmid) using Lipofectamine™
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Following 48-h of incubation at 37°C, the
transfection efficiency was detected by reverse
transcription-quantitative PCR (RT-qPCR).
RNA isolation and RT-qPCR
Following transfection as previously described,
total RNA from rat granulosa cells (24-well plates at a density of
2×105 cells per well) were extracted using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Total RNA was quantified
using a NanoDrop-1000 spectrophotometer (Thermo Fisher Scientific,
Inc.). RNA was reverse transcribed into cDNA using PrimeScript RT
reagent kit (Takara Bio, Inc.) according to the manufacturer's
protocol. The following reverse transcription conditions were used:
Initial denaturation at 37°C for 15 min, followed by 85°C for 5 sec
and 4°C for 5 min. The relative expression of miRNAs were
determined using TaqMan miRNA assay (Thermo Fisher Scientific,
Inc.) on an ABI 7500 Fast Instrument (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The following thermocycling conditions
were used for this qPCR: Initial denaturation at 95°C for 15 min,
followed by 40 cycles at 95°C for 10 sec and at 60°C for 60 sec.
The relative levels of mRNA were quantified using the SYBR Premix
Ex Taq (Takara Bio, Inc.) according to the manufacturer's protocol.
The following thermocycling conditions were used for this qPCR:
Initial denaturation at 95°C for 3 min, followed by 40 cycles at
95°C for 5 sec and at 60°C for 30 sec. β-actin and U6 were used for
mRNA and miRNA normalization, respectively. The primers utilized
were as follows: miR-30d-5p forward, 5′-CCTGTTGGTGCACTTCCTAC-3′ and
reverse, 5′-TGCAGTAGTTCTCCAGCTGC-3′; Smad2 forward,
5′-GTTCCTGCCTTTGCTGAGAC-3′ and reverse, 5′-TTCTCTTTGCCAGGAATGCT-3′;
Smad3 forward, 5′-GGAGGAGAAATGGTGCGAGAA-3′ and reverse,
5′-GCCACAGGCGGCAGTAGAT-3′; β-actin forward,
5′-CGAGCGTGGCTACAGCTTC-3′ and reverse, 5′-GTCACGCACGATTTCCCTCT-3′;
and U6 forward, 5′-ATGACGTCTGCCTTGGAGAAC-3′ and reverse,
5′-TCAGTGTGCTACGGAGTTCAG-3′. Relative gene expression was
quantified using the 2−ΔΔCq method (25). Experiments were repeated in
triplicate.
Dual-luciferase reporter assay
TargetScan (http://www.targetscan.org/vert_71/) was used to
predict the potential target genes of miR-30d-5p. The results
identified binding sites between miR-30d-5p and Smad2. A luciferase
reporter assay was subsequently performed to confirm the binding
sites between miR-30d-5p and Smad2 3′UTR. The 3′UTR of Smad2
containing the miR-30d-5p putative wild-type (WT) and mutant (MUT)
binding site were cloned into the psiCHECK-2 luciferase reporter
vector (Promega Corporation). Rat granulosa cells were plated
(5×104 per well) in 24-well plates and co-transfected
with miR-30d-5p mimic or NC and psiCHECK-2-Smad2-3′UTR-WT or
psiCHECK-2- Smad2-3′UTR-MUT using Lipofectamine™ 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). Following 48 h of incubation at
37°C, the luciferase activity was measured using a dual-luciferase
reporter assay system (Promega Corporation). Renilla
luciferase activity was used as an internal control. Experiments
were repeated in triplicate.
Western blot analysis
Following transfection as previously described,
total protein samples were extracted from rat granulosa cells
(6-well plates at a density of 4×105 cells per well)
following transfection as previously described using RIPA lysis
buffer (Gibco; Thermo Fisher Scientific, Inc.) containing
phenylmethylsulfonyl fluoride (Beyotime Institute of Biotechnology)
and phosphatase inhibitor cocktail (cat. no. ab201112; Abcam).
Protein concentrations were determined using the bicinchoninic acid
method. An equal quantity of protein (40 µg) obtained from cell
lysates were separated via 10% SDS-PAGE gel and then
electrophoretically transferred onto PVDF membranes (Immobilon; EMD
Millipore). The membranes were blocked with 5% non-fat dry milk for
1 h at room temperature, and incubated with the following primary
antibodies overnight at 4°C: Phosphorylated (p)-Smad2 (cat. no.
18338; 1:1,000; Cell Signaling Technology, Inc.), Smad2 (cat. no.
8685; 1:1,000; Cell Signaling Technology, Inc.), p-Smad3 (cat. no.
9520; 1:1,000; Cell Signaling Technology, Inc.), Smad3 (cat. no.
9523; 1:1,000; Cell Signaling Technology, Inc.) and β-actin (cat.
no. 4970; 1:1,000; Cell Signaling Technology, Inc.). Membranes were
then further incubated with horseradish peroxidase-conjugated
secondary antibodies (cat. no. 7074; 1:1,000; Cell Signaling
Technology, Inc.) at room temperature for 1 h. Proteins bands were
visualized using an enhanced chemiluminescence kit (Pierce; Thermo
Fisher Scientific, Inc.) and quantified using ImageJ software
(version 1.8.0; National Institutes of Health). Experiments were
repeated for three times.
MTT assay
Rat granulosa cells were seeded into 96-well plate
at 1×104 cells per well and cultured for 24 h at 37°C.
Cells were then transfected as previously described for 12, 24 or
48 h. Cells were incubated with 20 µl MTT (5 mg/ml; Sigma-Aldrich;
Merck KGaA) for 4 h at 37°C, after which the DMEM/Ham's nutrient
mixture F-12 medium was replaced with 150 µl DMSO to dissolve the
purple formazan product. The optical density at a wavelength of 490
nm was recorded using a microplate reader (Multiskan FC; Thermo
Fisher Scientific, Inc.). Experiments were repeated in
triplicate.
Flow cytometry analysis
An Annexin V-FITC/propidium iodide (PI) apoptosis
detection kit (Abcam) was used to evaluate cell apoptosis.
Following 48 h of transfection, rat granulosa cells were collected
and washed with cold PBS, after which cells were treated with 0.25%
trypsin to digest the cells. Cell pellets were collected,
centrifuged with 1,000 × g for 5 min at 20°C and suspended in PBS.
Subsequently, the supernatant was discarded and re-suspended with a
binding buffer containing Annexin V-FITC and PI for 15 min in the
dark at room temperature. Flow cytometry (FACSCalibur; BD
Biosciences) was used to evaluate cell apoptotic rate and the data
was analyzed using FlowJo software (version 7.6.1; FlowJo LLC).
Experiments were repeated in triplicate.
Statistical analysis
Statistical analysis was performed using SPSS 13.0
statistical software (SPSS, Inc.). Data were presented as mean ±
standard deviation of three independent experiments. A Student's
t-test was used to compare the differences between two groups.
One-way ANOVA followed by Tukey's post hoc test was used to analyze
the differences between more than two groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Smad2 is a target gene of
miR-30d-5p
Bioinformatics analysis predicted that miR-30d-5p
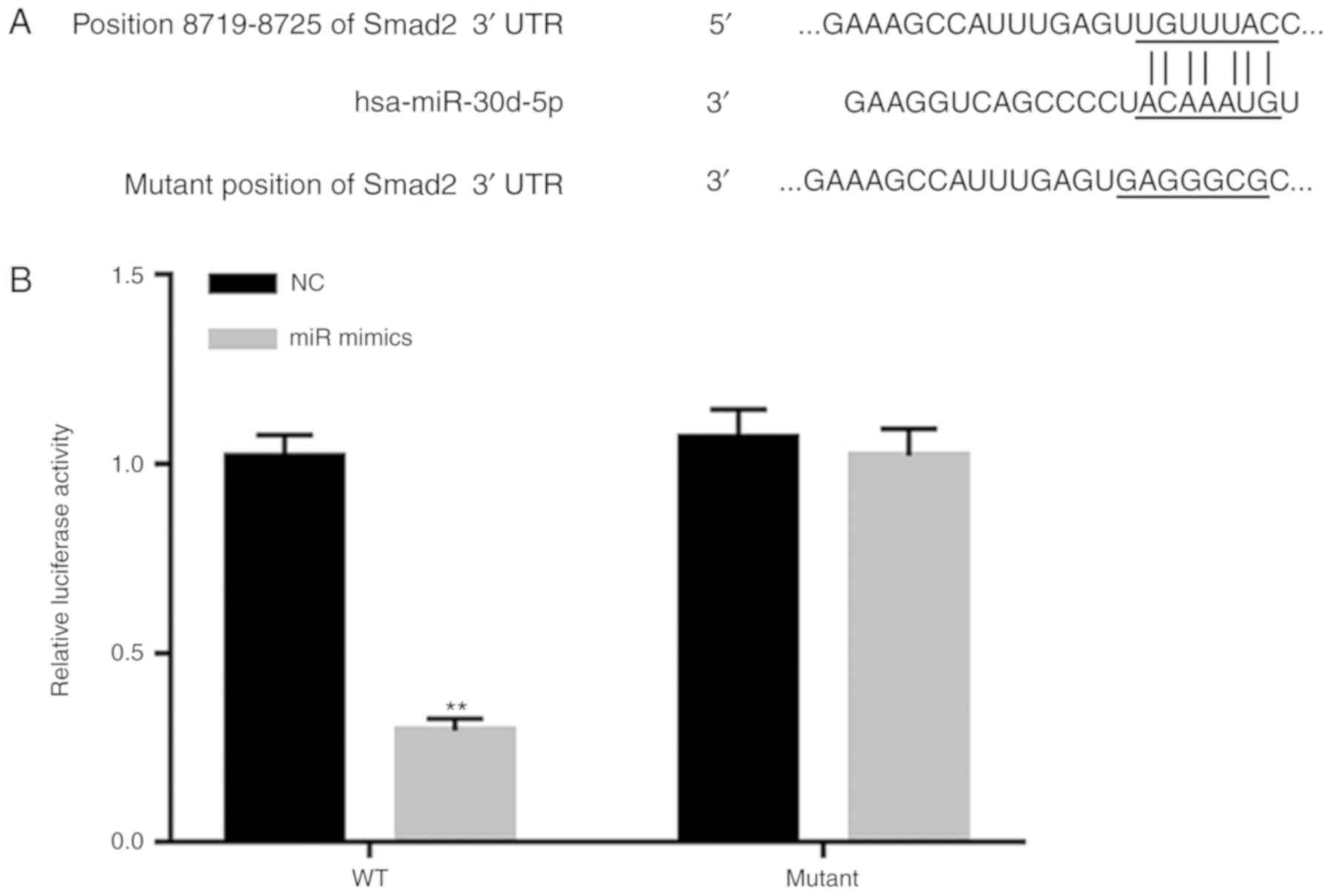
had hundreds of potential target genes including Smad2 (Fig. 1A). To confirm the relationship
between miR-30d-5p and Smad2, dual-luciferase reporter assay was
performed. As presented in Fig. 1B,
the luciferase activity of the miR-30d-5p mimic group following
transfection with the WT-Smad2 3′-UTR luciferase reporter vector
was significantly decreased compared with the NC group, whilst
there was no significant difference in miR-30d-5p mimics group
transfected with MUT-Smad2 3′-UTR luciferase reporter vector
compared with the NC group. The results indicated that Smad2 was
the target gene of miR-30d-5p.
Smad2 overexpression reduces apoptosis
and increases the viability of rat ovarian granulosa cells
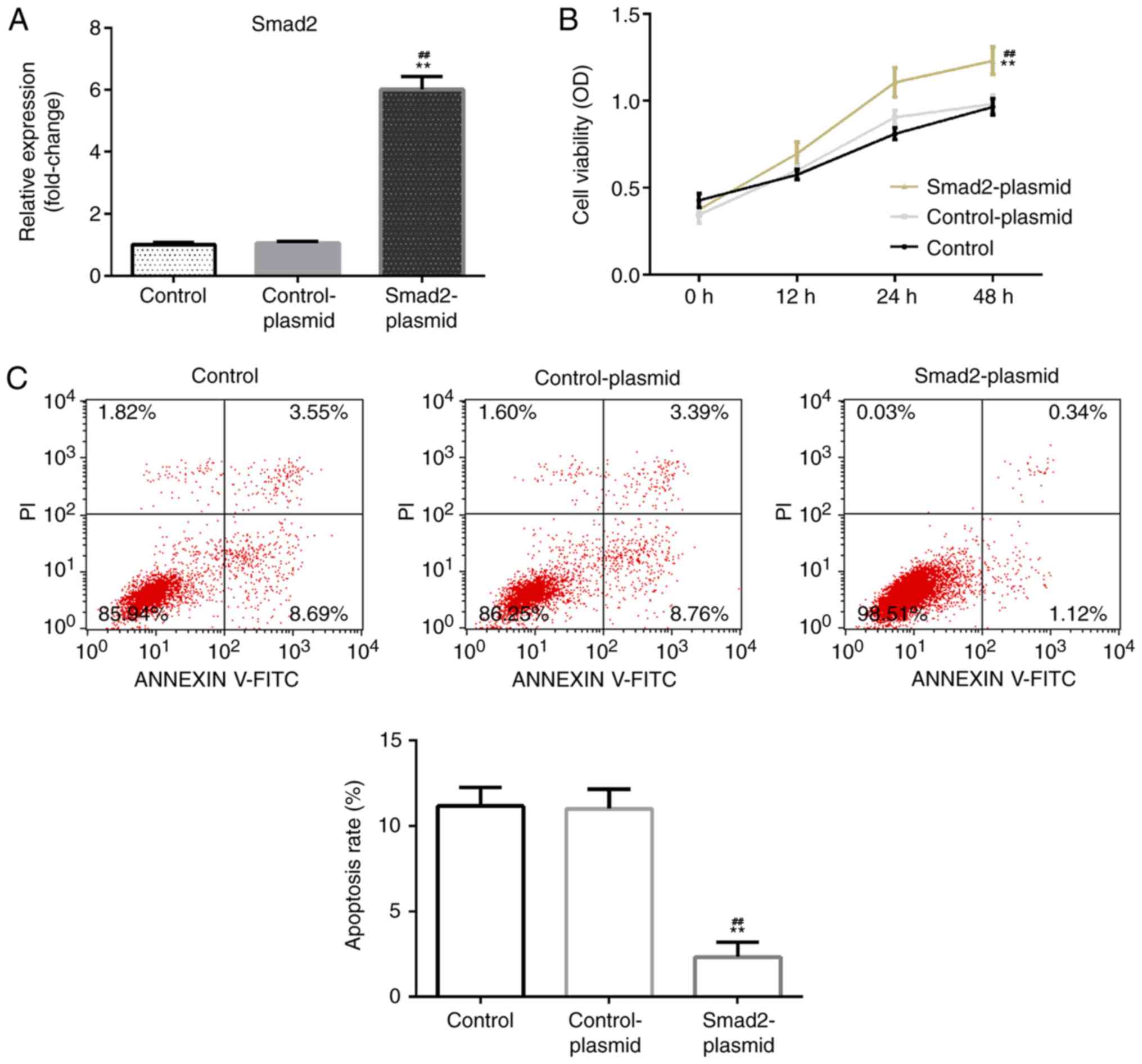
The effect of Smad2 overexpression on rat ovarian
granulosa cell viability and apoptosis was investigated. Rat
ovarian granulosa cells were transfected with Smad2 plasmid or
control-plasmid for 48 h. RT-qPCR was then performed to detect
transfection efficiency. It was determined that, compared with the
control group, the Smad2 plasmid significantly enhanced Smad2 mRNA
levels in rat ovarian granulosa cells (Fig. 2A). Further analysis indicated that
when compared with the control and control plasmid group, the Smad2
plasmid significantly enhanced rat ovarian granulosa cell viability
(Fig. 2B) and reduced cell apoptosis
(Fig. 2C).
miR-30d-5p decreases the viability of
rat ovarian granulosa cells by targeting Smad2
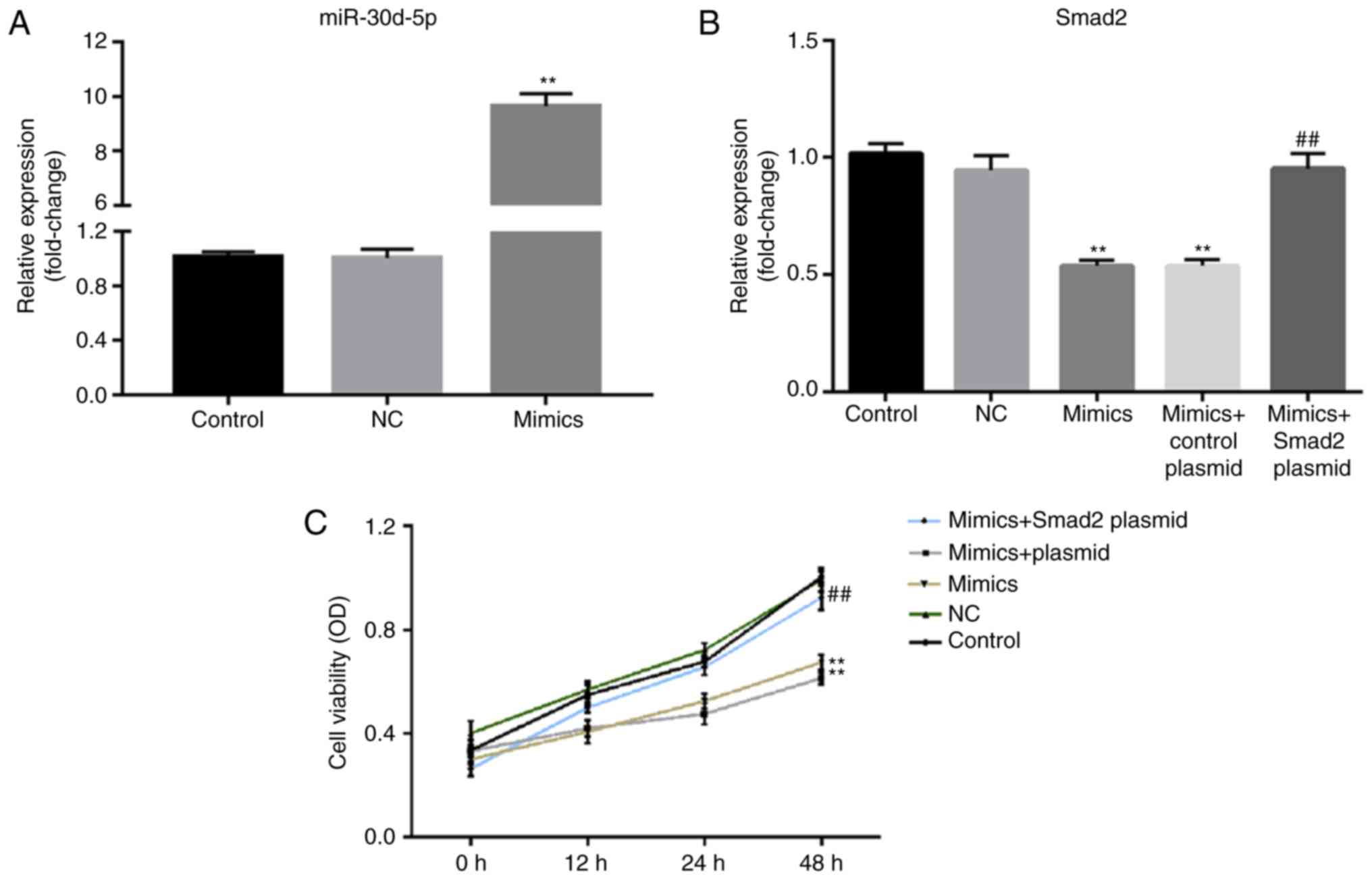
To further confirm whether Smad2 was involved with
miR-30d-5p in ovarian granulosa cells, rat ovarian granulosa cells
were transfected with miR-30d-5p mimics, NCs, miR-30d-5p mimics +
control plasmid or miR-30d-5p mimics + Smad2-plasmid for 48 h.
After transfection with miR-30d-5p mimics for 48 h, transfection
efficiency was detected via RT-qPCR, where the level of miR-30d-5p
significantly increased compared with the NC group (Fig. 3A). In addition, it was determined
that miR-30d-5p mimics significantly reduced the level of Smad2
mRNA in rat ovarian granulosa cells compared with the NC group.
This decrease was also significantly reversed following
Smad2-plasmid co-transfection (Fig.
3B). The viability of ovarian granulosa cells was examined to
confirm the biological role of miR-30d-5p in ovarian granulosa
cells. An MTT assay demonstrated that following transfection with
miR-30d-5p mimics, rat ovarian granulosa cell viability was
markedly decreased compared with the NC group. This decrease was
reversed following transfection with the Smad2-plasmid (Fig. 3C). The results indicated that
overexpression of Smad2 reversed the effects of miR-30d-5p on
ovarian granulosa cell proliferation.
miR-30d-5p promotes the apoptosis of
rat ovarian granulosa cells by targeting Smad2
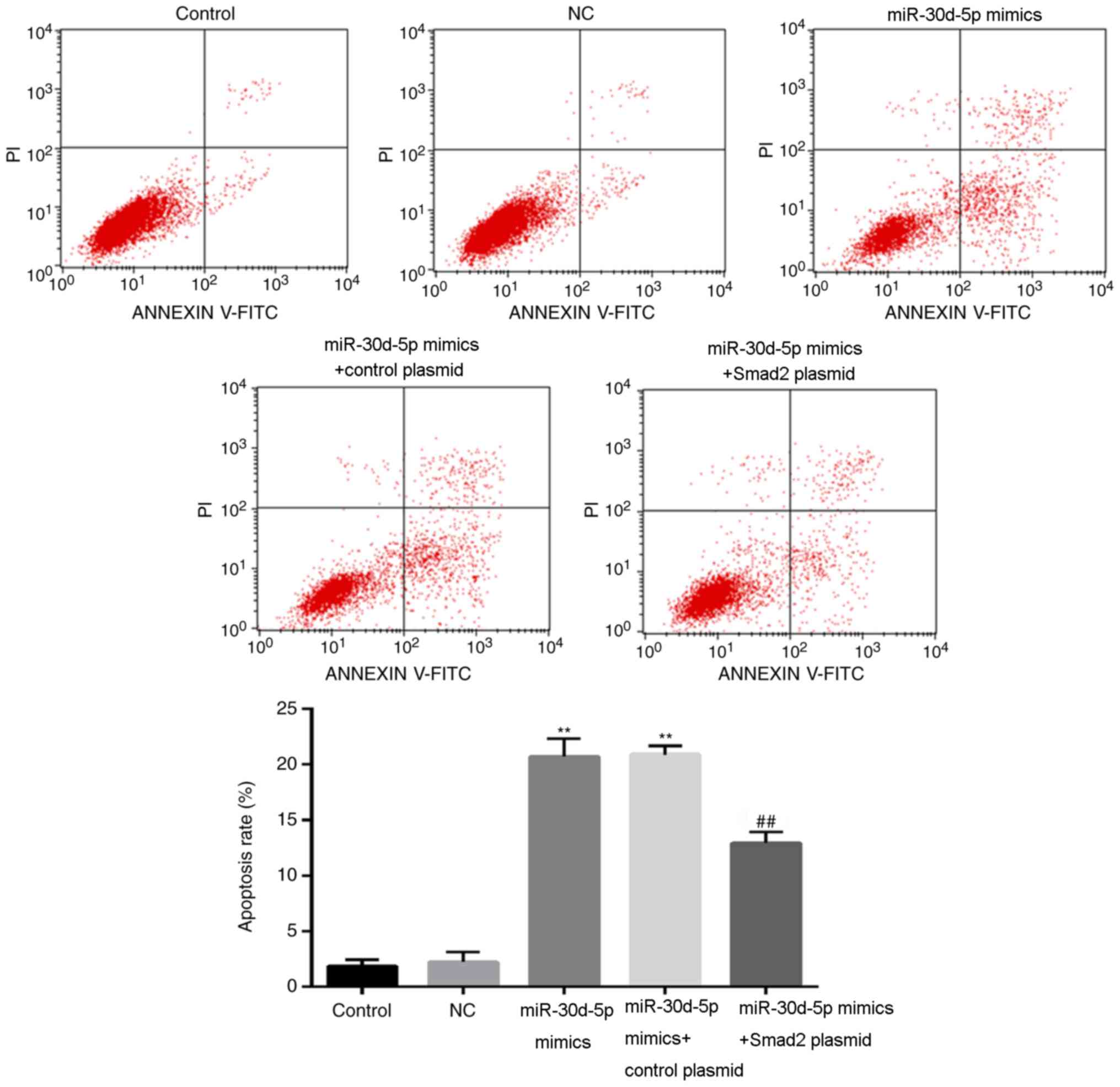
Flow cytometry results demonstrated that the
apoptotic rate of the miR-30d-5p mimics group was significantly
increased compared with the control group (Fig. 4). The apoptotic rate of the
miR-30d-5p mimic + Smad2 plasmid group was significantly lower
compared with the miR-30d-5p mimics transfection group (Fig. 4). These results indicated that
overexpression of Smad2 reversed the effects of miR-30d-5p on
ovarian granulosa cell apoptosis.
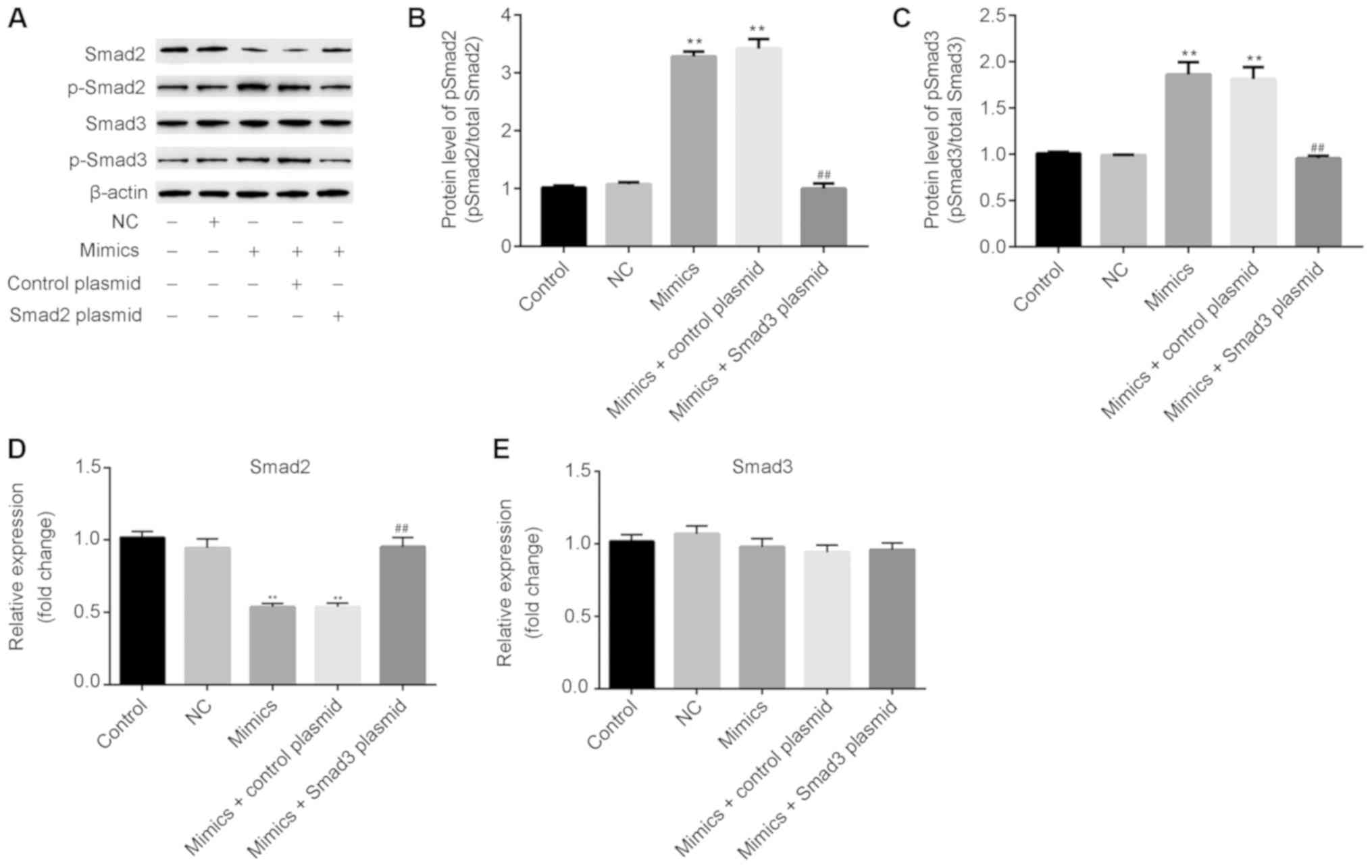
miR-30d-5p increases the ratio of
p-Smad2/Smad2 and p-Smad 3/Smad 3 in rat ovarian granulosa
cells
Proliferation can be regulated by the Smad protein
pathway (26). As presented in
Fig. 5, after transfection with
miR-30d-5p mimics, the protein (Fig. 5A
and B) and mRNA (Fig. 5D)
expression of Smad2 was significantly decreased, whilst the protein
(Fig. 5A) and mRNA (Fig. 5E) expression of Smad3 exhibited no
significant changes. In addition, the protein expression of the
phosphorylated (active) forms of Smad2 and Smad3 (Fig. 5A-C) were markedly increased in rat
ovarian granulosa cells following miR-30d-5p mimic transfection.
Notably, overexpression of Smad2 reversed the effects of miR-30d-5p
on the expression of p-Smad2/Smad2 and p-Smad3/Smad3 in ovarian
granulosa cells.
Discussion
PCOS is the most common metabolic and endocrine
disease in women of childbearing age, involving multiple factors
and a complicated etiology and pathophysiology (1). Researchers have identified that PCOS is
associated with disorders of multiple factors regulating the
ovaries. miR-30d-5p has been reported to serve important roles in
the regulation of cell proliferation, invasion and apoptosis in a
variety of tumor cells (27,28). A previous study also reported that
miR-30d-5p levels increased during cultured granulosa cell
secretion of FSH-mediated progesterone (21), indicating that it may be closely
associated with PCOS. In the present study, the results implied
that miR-30d-5p might regulate the proliferation and apoptosis of
ovarian granulosa cells by targeting Smad2, thus serving an
important role in PCOS.
Previous reports have indicated that miRNAs may be
involved in the pathogenesis of PCOS (29,30).
miR-324-3p levels have also been demonstrated to decrease in the
ovaries of PCOS rats (31).
Overexpression of miR-324-3p reduces the proliferation and induces
the apoptosis of granulosa cells by targeting Wnt family member 2B
(31). In addition, it has been
revealed that miR-141-3p is markedly decreased in the ovaries of
rat PCOS models and that apoptosis is inhibited in rat ovarian
granulosa cells by targeting death associated protein kinase 1
(32). A previous study demonstrated
that miR-30d-5p is markedly downregulated during cultured granulosa
cell secretion of FSH-mediated progesterone (21). As a member of Smad protein family,
Smad2 serves a key role in regulating cell proliferation and
apoptosis. Abnormal proliferation and/or apoptosis in granulosa
cells serves an important role in PCOS (8,9).
However, the relationship between miR-30d-5p and Smad2 remains
unclear. In the present study, a dual-luciferase reporter assay
confirmed that Smad2 was a target gene of miR-30d-5p and that Smad2
overexpression enhanced rat ovarian granulosa cell proliferation
and inhibited cell apoptosis. To understand the role of miR-30d-5p
in ovarian granulosa cells, samples were transfected with
miR-30d-5p mimics. The results demonstrated that miR-30d-5p
inhibited cell growth and promoted apoptosis, indicating that
miR-30d-5p could be involved in the regulation of rat ovarian
granulosa cell growth. Furthermore, Smad2 plasmid co-transfection
reversed all the inhibitory effects of miR-30d-5p on rat ovarian
granulosa cell viability and apoptosis.
Previous studies have reported that the endometrium
of women with PCOS exhibits decreases in the inhibitory activity of
the cell cycle from the G1 to S phase via the action of the Smad
protein, thereby inducing cell cycle progression (33,34).
Smad proteins constitute regulatory molecules of cellular
proliferation and apoptosis (35).
To better understand the mechanisms of miR-30d-5p used in the
regulation of ovarian granulosa cell survival, the participation of
the Smad pathway was evaluated in the present study. The results
demonstrated that the levels of Smad 2 significantly decreased,
whilst the p-Smad2 and p-Smad3 protein levels in the miR-30d-5p
mimic group markedly increased when compared with the control
group. Additionally, these effects could be reversed by Smad2
overexpression. The deregulation of Smad2 proteins may be
associated with the miR-30d-5p-induced apoptosis of ovarian
granulosa cells. However, further studies are required to
understand the role of miR-30d-5p and Smad2 in the pathogenesis of
PCOS.
In conclusion, the present results indicated that
Smad2 was a direct target of miR-30d-5p. miR-30d-5p was also
determined to promote ovarian granulosa cells apoptosis by
targeting Smad2. The results of this study indicated that
miR-30d-5p may be a new therapeutic target for PCOS treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MY wrote the manuscript and analyzed and interpreted
the data. JL designed the study and revised the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Norman RJ, Dewailly D, Legro RS and Hickey
TE: Polycystic ovary syndrome. Lancet. 370:685–697. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fux Otta C, Fiol de Cuneo M and Szafryk de
Mereshian P: Polycystic ovary syndrome: Physiopathology review. Rev
Fac Cien Med Univ Nac Cordoba. 70:27–30. 2013.(In Spanish).
PubMed/NCBI
|
|
3
|
McGee EA and Hsueh AJ: Initial and cyclic
recruitment of ovarian follicles. Endocr Rev. 21:200–214. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Craig J, Orisaka M, Wang H, Orisaka S,
Thompson W, Zhu C, Kotsuji F and Tsang BK: Gonadotropin and
intra-ovarian signals regulating follicle development and atresia:
The delicate balance between life and death. Front Biosci.
12:3628–3639. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hsueh AJ, Kawamura K, Cheng Y and Fauser
BC: Intraovarian control of early folliculogenesis. Endocr Rev.
36:1–24. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
de Melo AS, Dias SV, Cavalli Rde C,
Cardoso VC, Bettiol H, Barbieri MA, Ferriani RA and Vieira CS:
Pathogenesis of polycystic ovary syndrome: Multifactorial
assessment from the foetal stage to menopause. Reproduction.
150:R11–R24. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qiao J and Feng HL: Extra- and
intra-ovarian factors in polycystic ovary syndrome: Impact on
oocyte maturation and embryo developmental competence. Hum Reprod
Update. 17:17–33. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Onalan G, Selam B, Baran Y, Cincik M,
Onalan R, Gündüz U, Ural AU and Pabuccu R: Serum and follicular
fluid levels of soluble Fas, soluble Fas ligand and apoptosis of
luteinized granulosa cells in PCOS patients undergoing IVF. Hum
Reprod. 20:2391–2395. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shalev E, Goldman S and Ben-Shlomo I: The
balance between MMP-9 and MMP-2 and their tissue inhibitor (TIMP)-1
in luteinized granulosa cells: Comparison between women with PCOS
and normal ovulatory women. Mol Hum Reprod. 7:325–331. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y and Lee CG: MicroRNA and
cancer-focus on apoptosis. J Cell Mol Med. 13:12–23. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bueno MJ, Pérez de Castro I and Malumbres
M: Control of cell proliferation pathways by microRNAs. Cell Cycle.
7:3143–3148. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Carletti MZ, Fiedler SD and Christenson
LK: MicroRNA 21 blocks apoptosis in mouse periovulatory granulosa
cells. Biol Reprod. 83:286–295. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yao G, Yin M, Lian J, Tian H, Liu L, Li X
and Sun F: MicroRNA-224 is involved in transforming growth
factor-beta-mediated mouse granulosa cell proliferation and
granulosa cell function by targeting Smad4. Mol Endocrinol.
24:540–551. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zheng M, Hou L, Ma Y, Zhou L, Wang F,
Cheng B, Wang W, Lu B, Liu P, Lu W and Lu Y: Exosomal let-7d-3p and
miR-30d-5p as diagnostic biomarkers for non-invasive screening of
cervical cancer and its precursors. Mol Cancer. 18:762019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hosseini SM, Soltani BM, Tavallaei M,
Mowla SJ, Tafsiri E, Bagheri A and Khorshid HRK: Clinically
significant dysregulation of hsa-miR-30d-5p and hsa-let-7b
expression in patients with surgically resected non-small cell lung
cancer. Avicenna J Med Biotechnol. 10:98–104. 2018.PubMed/NCBI
|
|
17
|
Song Y, Song C and Yang S:
Tumor-suppressive function of miR-30d-5p in prostate cancer cell
proliferation and migration by targeting NT5E. Cancer Biother
Radiopharm. 33:203–211. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
He Y, Chen X, Yu Y, Li J, Hu Q, Xue C,
Chen J, Shen S, Luo Y, Ren F, et al: LDHA is a direct target of
miR-30d-5p and contributes to aggressive progression of gallbladder
carcinoma. Mol Carcinog. 57:772–783. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu X, Zhao J and He Y: Long non-coding RNA
PVT1 functions as an oncogene in human colon cancer through
miR-30d-5p/RUNX2 axis. J BUON. 23:48–54. 2018.PubMed/NCBI
|
|
20
|
Jiang M, Wang H, Jin M, Yang X, Ji H,
Jiang Y, Zhang H, Wu F, Wu G, Lai X, et al: Exosomes from
MiR-30d-5p-ADSCs reverse acute ischemic stroke-induced,
autophagy-mediated brain injury by promoting M2
microglial/macrophage polarization. Cell Physiol Biochem.
47:864–878. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yao N, Yang BQ, Liu Y, Tan XY, Lu CL, Yuan
XH and Ma X: Follicle-stimulating hormone regulation of microRNA
expression on progesterone production in cultured rat granulosa
cells. Endocrine. 38:158–166. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Massagué J: G1 cell-cycle control and
cancer. Nature. 432:298–306. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mishra L, Derynck R and Mishra B:
Transforming growth factor-beta signaling in stem cells and cancer.
Science. 310:68–71. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Syed V: TGF-β signaling in cancer. J Cell
Biochem. 117:1279–1287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Meran S, Luo DD, Simpson R, Martin J,
Wells A, Steadman R and Phillips AO: Hyaluronan facilitates
transforming growth factor-β1-dependent proliferation via CD44 and
epidermal growth factor receptor interaction. J Biol Chem.
286:17618–17630. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ye C, Yu X, Liu X, Dai M and Zhang B:
miR-30d inhibits cell biological progression of Ewing's sarcoma by
suppressing the MEK/ERK and PI3K/Akt pathways in vitro.
Oncol Lett. 15:4390–4396. 2018.PubMed/NCBI
|
|
28
|
Yao J, Liang L, Huang S, Ding J, Tan N,
Zhao Y, Yan M, Ge C, Zhang Z, Chen T, et al: MicroRNA-30d promotes
tumor invasion and metastasis by targeting Galphai2 in
hepatocellular carcinoma. Hepatology. 51:846–856. 2010.PubMed/NCBI
|
|
29
|
Hossain MM, Cao M, Wang Q, Kim JY,
Schellander K, Tesfaye D and Tsang BK: Altered expression of miRNAs
in a dihydrotestosterone-induced rat PCOS model. J Ovarian Res.
6:362013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Roth LW, McCallie B, Alvero R, Schoolcraft
WB, Minjarez D and Katz-Jaffe MG: Altered microRNA and gene
expression in the follicular fluid of women with polycystic ovary
syndrome. J Assist Reprod Genet. 31:355–362. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jiang YC and Ma JX: The role of MiR-324-3p
in polycystic ovary syndrome (PCOS) via targeting WNT2B. Eur Rev
Med Pharmacol Sci. 22:3286–3293. 2018.PubMed/NCBI
|
|
32
|
Li D, Xu D, Xu Y, Chen L, Li C, Dai X,
Zhang L and Zheng L: MicroRNA-141-3p targets DAPK1 and inhibits
apoptosis in rat ovarian granulosa cells. Cell Biochem Funct.
35:197–201. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen C, Sun MZ, Liu S, Yeh D, Yu L, Song
Y, Gong L, Hao L, Hu J and Shao S: Smad4 mediates malignant
behaviors of human ovarian carcinoma cell through the effect on
expressions of E-cadherin, plasminogen activator inhibitor-1 and
VEGF. BMB Rep. 43:554–560. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Goto N, Hiyoshi H, Ito I, Tsuchiya M,
Nakajima Y and Yanagisawa J: Estrogen and antiestrogens alter
breast cancer invasiveness by modulating the transforming growth
factor-β signaling pathway. Cancer Sci. 102:1501–1508. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bacallao K, Plaza-Parrochia F, Cerda A,
Gabler F, Romero C, Vantman D and Vega M: Levels of regulatory
proteins associated with cell proliferation in endometria from
untreated patients having polycystic ovarian syndrome with and
without endometrial hyperplasia. Reprod Sci. 23:211–218. 2016.
View Article : Google Scholar : PubMed/NCBI
|