Introduction
Peripheral nerve (PN) injury (PNI) is a common
condition, occurring in a range of diseases and after a variety of
injuries, that can lead to sensory and motor dysfunction or even
permanent disability (1). When the
peripheral nerve is severely damaged, a series of metamorphoses and
phagocytic processes, such as axonal necrosis and degradation of
the myelin sheath, occur in distal nerves, in a process referred to
as Wallerian degeneration (2).
Structural and sensory problems, such as innervated muscle atrophy
and localized sensory loss of innervation, also occur after PNI
(3). Peripheral nerve (PN)
regeneration is a complex process associated with the elimination
of inflammation, release of neurotrophic factors, axonal growth and
neuron survival (3). Successful
peripheral nerve regeneration after injury depends on dynamic
changes in Schwann cells (SCs) (4,5). After
PNI, SCs begin to divide and proliferate, forming a cord-like
channel, known as a Büngner band, surrounded by basal lamina
(6). SCs synthesize and release
neurotrophic and cell adhesion factors that stimulate the
regeneration of myelin and axons and provide the necessary
attachment surfaces for regenerating axons (7,8).
However, the regeneration of injured nerves is slow, due to the
complexity of the repair process, and prolonged denervation of the
proximal nerve and neural mismatch may result in irreversible
atrophy of the associated organ (9,10). For
this reason, PN repair remains a considerable clinical
challenge.
Currently the gold standard treatment for PNI is
autologous nerve transplantation, however, this treatment has a
number of limitations, such as shortage of donor sources, the need
for an extra incision, nerve matching problems, and possible loss
of neurological function at the donor site (11,12).
Due to advances in tissue engineering,
tissue-engineered nerve scaffolds provide a promising new strategy
for the treatment of PNI (13). This
method of tissue engineered nerve grafting involves the use of seed
cells that are cultured in vitro and anchored to a natural
or synthetic biocompatible conduit to form a composite nerve
scaffold. This scaffold can be implanted in vivo to bridge
gaps between injured nerves. Cells attached to scaffolds have shown
some positive effects in nerve regeneration (14). However, SCs, which are the central
factors required for PN regeneration, are difficult to obtain and
expand to sufficient numbers. Tissue availability is limited, due
to problems with morbidity of the donor site and the risk of
sacrifice of one or more normal nerves leading to a loss of
sensation (15,16). Furthermore, SCs are terminal cells
(17). Although techniques to
increase the acquired number and survival rates of SCs in
vitro have been developed (18,19),
further studies are required to make these a viable option. In the
last decade, the field of regenerative medicine has been focused on
stem cells, which may be useful in PNI treatment as they can be
differentiated into Schwann (SC)-like cells through physical and
chemical induction methods (20,21).
Adipose-derived mesenchymal stem cells (ADSCs) are
adult mesenchymal stem cells (MSCs) with self-renewal ability and
multidirectional differentiation potential (20). Compared with bone marrow-derived MSCs
(BMSCs), ADSCs have various advantages, such as the ease of their
extraction, their abundance in adipose tissues and their rapid rate
of proliferation (22). A number of
studies have confirmed that PNI can be alleviated after ADSC
transplantation into nerve injury sites (23,24). A
number of studies have previously demonstrated that ADSC
transplantation has a regenerative effect on sciatic nerve injury
in a rat model (25,26). These studies found that only a small
number of ADSCs survived in vivo and that the rate of ADSC
differentiation into SC-like cells at the site of injury was low.
Therefore, the success of PNI treatment with undifferentiated ADSCs
is not guaranteed (27). Several
investigators have induced ADSC differentiation into SC-like cells
in vitro and then transplanted these cells in vivo,
confirming that SC-like cells differentiated from ADSCs can promote
peripheral nerve regeneration (28,29).
However, these differentiation procedures have a number of
disadvantages, including cumbersome protocols, high cost and low
differentiation rate (30,31). The results of previous studies
suggested that SCs secrete neurotrophic factors, which are released
by injured nerve endings during PNI, that can promote axon growth
and protect neurons (32). Based on
these findings, injured nerve endings were placed in culture medium
to generate a neurotrophic substance-rich nerve leachate (33–35). A
previous study showed that nerve leachate induced the in
vitro differentiation of ADSCs into SC-like cells, which
expressed specific SC markers (36).
This differentiation method is convenient, economical and simple.
However, whether nerve leachate-treated ADSCs can influence PNI
repair has yet to be confirmed in an in vivo study. In the
present study, a 1-cm length of sciatic nerve was removed, in order
to establish a model of PNI, and nerve leachate-differentiated
ADSCs on acellular nerve scaffolds were transplanted to bridge this
defect. The repair effects of nerve leachate-treated ADSCs on rat
sciatic nerve injury were determined through investigation of
indicators including nerve regeneration and gastrocnemius
recovery.
Materials and methods
Animals
Two groups of male Sprague-Dawley (S-D) rats (6;
age, 3-weeks; weight, 45–55 g; and 21; age, 8-weeks; weight range,
180–210 g) were obtained from the Experimental Animal Center of
Zhengzhou University Medical College (Zhengzhou, China).
Three-week-old rats were used to isolate and culture ADSCs, whereas
eight-week-old rats were used for in vivo experiments. The
animals were housed at 20–26°C with 50–60% humidity, under a 12-h
light/dark cycle and had free access to water and food. Experiments
were designed to minimize animal suffering and reduce the number of
experimental animals used. All animal care and experimental
protocols were conducted in accordance with university policies on
the use and care of animals and were approved by the Institutional
Animal Experiment Committee of Henan University of Science and
Technology (Henan, China).
Isolation and culture of rat
ADSCs
After the sacrifice of 6 3-week old rats, adipose
tissue was isolated from the inguinal region and minced under
aseptic conditions. This isolated adipose tissue was digested at
37°C for 60 min with 0.1% collagenase type I (Gibco; Thermo Fisher
Scientific, Inc.). The resulting suspension was centrifuged at 175
× g (1,000 rpm) for 10 min at room temperature to separate the
ADSCs from the stromal vascular fraction (36). The precipitated cells were
resuspended and cultured at 37°C in 5% CO2 in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.). Cells that did not adhere to tissue
culture plastic after 24 h were removed when the medium was
replaced. Rat ADSCs were passaged 3–5 times before use.
Verification of ADSC multi-lineage
differentiation and expression of specific surface markers
Using the method reported by Fu et al
(16), passage 3 rat ADSCs were
seeded into six-well plates at a density of 5×105
cells/ml. When the ADSCs were 80% confluent, normal medium was
replaced with medium to induce their differentiation. ADSCs were
cultured for 14 days in adipogenic differentiation medium (DMEM
supplemented with 10% FBS, 0.5 mM isobutyl-methylxanthine, 1 µM
dexamethasone, 10 µM insulin and 200 µM indomethacin;
Sigma-Aldrich; Merck KGaA) (36).
The cells were fixed by 4% paraformaldehyde for 30 min at room
temperature, then washed with PBS for two times and the lipid
droplets present in the cells were stained using oil red O for 30
min at room temperature (Sigma-Aldrich; Merck KGaA). Rat ADSCs were
cultured for 21 days in osteogenic differentiation medium (DMEM
supplemented with 10% FBS, 100 nM dexamethasone, 50 µM
ascorbate-2-phosphate, and 10 mM β-glycerophosphate) (37). The cells were fixed by 4%
paraformaldehyde for 30 min at room temperature, then washed two
times with PBS before intracellular calcium deposition in the cells
was assessed by von Kossa staining for 30 min at room temperature
(all reagents are from Sigma-Aldrich; Merck KGaA).
The immunophenotype of ADSCs was determined by flow
cytometry. Passage 3 rat ADSCs were washed with PBS and then the
cells were fixed by 4% paraformaldehyde for 30 min at room
temperature. The fixed cells were incubated with FITC-coupled
antibodies against rat CD31 (1:200; cat. no. MCA1334G), CD44
(1:200; cat. no. MCA643), CD45 (1:200; cat. no. MCA43) and CD90
(1:200; cat. no. MCA47; Bio-Rad Laboratories, Inc.) in the dark at
room temperature for 30 min and analyzed by a flow cytometer
(FACSCalibur™; Becton, Dickinson and Company) using the FlowJo
Software (version 7.6.1; FlowJo LLC).
Preparation of the nerve leachate
According to a previously reported method (36), rat sciatic nerves were removed from 9
male 8-weeks old rats under aseptic conditions, and cut into 2-cm
segments. After washing the nerve segments three times with PBS
containing 0.2% penicillin-streptomycin (Gibco; Thermo Fisher
Scientific, Inc.), they were transferred into 100 ml DMEM medium
containing 10% FBS, following which the medium containing the
sciatic nerve was stored at 4°C for 3 days. The culture medium was
removed on the third day and filtered with a 0.22 µm microporous
membrane filter. The nerve leachate was prepared prior to this
study.
Induction of rat ADSCs into SC-like
cells
Passage 3 ADSCs were used in all experiments.
Briefly, culture medium was removed from sub-confluent cultures of
ADSCs and replaced with nerve leachate medium, which was in turn
refreshed once every 1–2 days. Cells were then incubated for 5 days
at 37°C under 5% CO2. Morphological changes were
observed using a light microscope (Olympus Corporation) after ADSCs
were inducted for 96 h using the nerve leachate medium.
Preparation of the tissue-engineered
nerve scaffold
Acellular nerve scaffolds were prepared using the
method of Zhang et al (37,38) from
five 8-week-old male rats, which were prepared prior to this study.
Sciatic nerve segments isolated from rats were placed in distilled
water and underwent 3 cycles of freezing at −80°C for 60 min and
thawing at room temperature for 20 min per cycle. The nerve
segments were digested with 40 U/ml DNase (Sigma-Aldrich; Merck
KGaA) and RNase (Sigma-Aldrich, Merck KGaA) at 37°C for 2 h. They
were then immersed in 4% sodium hydroxide solution at 4°C for 24 h.
After washing with PBS, the nerve segments were lyophilized,
sterilized and stored at 4°C.
Suspensions of untreated ADSCs and nerve
leachate-treated ADSCs containing 5×107 cells were
prepared and placed in centrifuge tubes. A total of 2 ml collagen
(Sigma-Aldrich; Merck KGaA) was added to the centrifuge tubes at
4°C and the suspensions were mixed rapidly. Collagen with or
without cells was slowly injected into the end of the acellular
nerve scaffolds. The composite nerve scaffolds that contained cells
and collagen were then transferred to a Petri dish, incubated at
37°C under 5% CO2 in 5 ml DMEM containing 10% FBS for 4
days.
Surgical transplantation
The S-D rats were randomly divided into the
following four groups (n=3 per group): Scaffold only, untreated
ADSCs + scaffold, nerve leachate-treated ADSCs + scaffold, and
autograft. The rats were anesthetized with 1% pentobarbital sodium
(50 mg/kg) by intraperitoneal injection. The fur of the right hind
limb was shaved and the skin was sterilized. A skin incision was
made along the femoral axis of the right hind leg, and the thigh
muscles were separated. The sciatic nerve was exposed through the
thigh muscle interval, and a 1-cm nerve segment was removed
(39). An acellular nerve scaffold,
an acellular nerve scaffold combined with ADSCs, an acellular
scaffold combined with nerve leachate-treated ADSCs or an autograft
nerve were transplanted into this 1-cm segment to bridge the nerve
gap. The muscle layers and skin were sutured with 4-0 sutures.
After surgery, rat mental state, physical activity, diet, wound
healing and foot ulcer formation were checked regularly. After two
months, all rats were sacrificed by cervical dislocation following
intraperitoneal injection with 3% sodium pentobarbital (50
mg/kg).
Electrophysiological assessment
Electrophysiological analysis was performed on the
rats at 2 months post-surgery. Under anesthesia, the bilateral
sciatic nerves of the rats were exposed. Electrical stimuli were
then applied to the proximal portion of the nerve trunk, and the
nerve action potential (NAP) was measured at a distance of 10 mm
from the electrical stimulation site. The nerve conduction velocity
(NCV) was analyzed as a function of distance and time.
Histological assessment
Two months after the surgery, the rats were
sacrificed by cervical dislocation under anaesthesia by
intraperitoneal injection with 3% sodium pentobarbital (50 mg/kg).
The rat sciatic nerve was re-exposed and the regenerated nerve was
harvested. The nerve segment was fixed with 2% glutaraldehyde at
4°C for 1 h and post-fixed with 1% osmium tetroxide at 4°C for 1 h
before it was embedded in epoxy resin. The embedded samples were
cut into thin sections (0.5–1.0 µm). These thin sections were
stained with 1% toluidine blue at 60°C for 5 min and the density of
myelinated nerve fibers was observed under a light microscope
(Olympus Corporation). The nerve segments were fixed in 2% (v/v)
glutaraldehyde in a 0.1-M phosphate buffer (pH 7.2) at 4°C for ≥2 h
and washed three times with PBS at room temperature. The samples
were then post-fixed with 1% (w/v) osmium tetroxide at 4°C for 2 h
followed by a further three-time PBS wash at room temperature. The
samples were dehydrated by using an alcohol gradient, replaced with
acetone and embedded in epoxy resin for 2 h at 45°C and polymerized
at 60°C for 48 h. Ultrathin sections (50 nm) were then lifted onto
formvar-coated grids, post-stained with lead citrate for 15 min and
uranyl acetate for 40 min at room temperature, and observed by
transmission electron microscopy (TEM; Hitachi, Ltd.). The total
number of myelinated nerve fibers and myelin thickness and area,
specifically the TEM data, were analyzed using HPIAS-1000-type
high-resolution color graphic pathological analysis system
(HPIAS-1000-type; Tongji Medical University, Wuhan, China).
Two months after surgery, the gastrocnemius tissue
was removed and embedded in paraffin and cut into thin sections
(5–10 µm). The sections were stained using Masson's trichrome
(40). The results were observed
under a light microscope and the cross-sectional area of the
gastrocnemius muscle was measured.
Statistical analysis
The data were analyzed with SPSS 18.0 software
(SPSS, Inc.). Each experiment was performed at least three times.
All quantitative data are presented as the mean ± standard
deviation. Differences between the groups were compared by one-way
analysis of variance and Duncan's multiple comparison tests.
P<0.05 was considered statistically significant.
Results
Morphology and multidirectional
differentiation of rat ADSCs
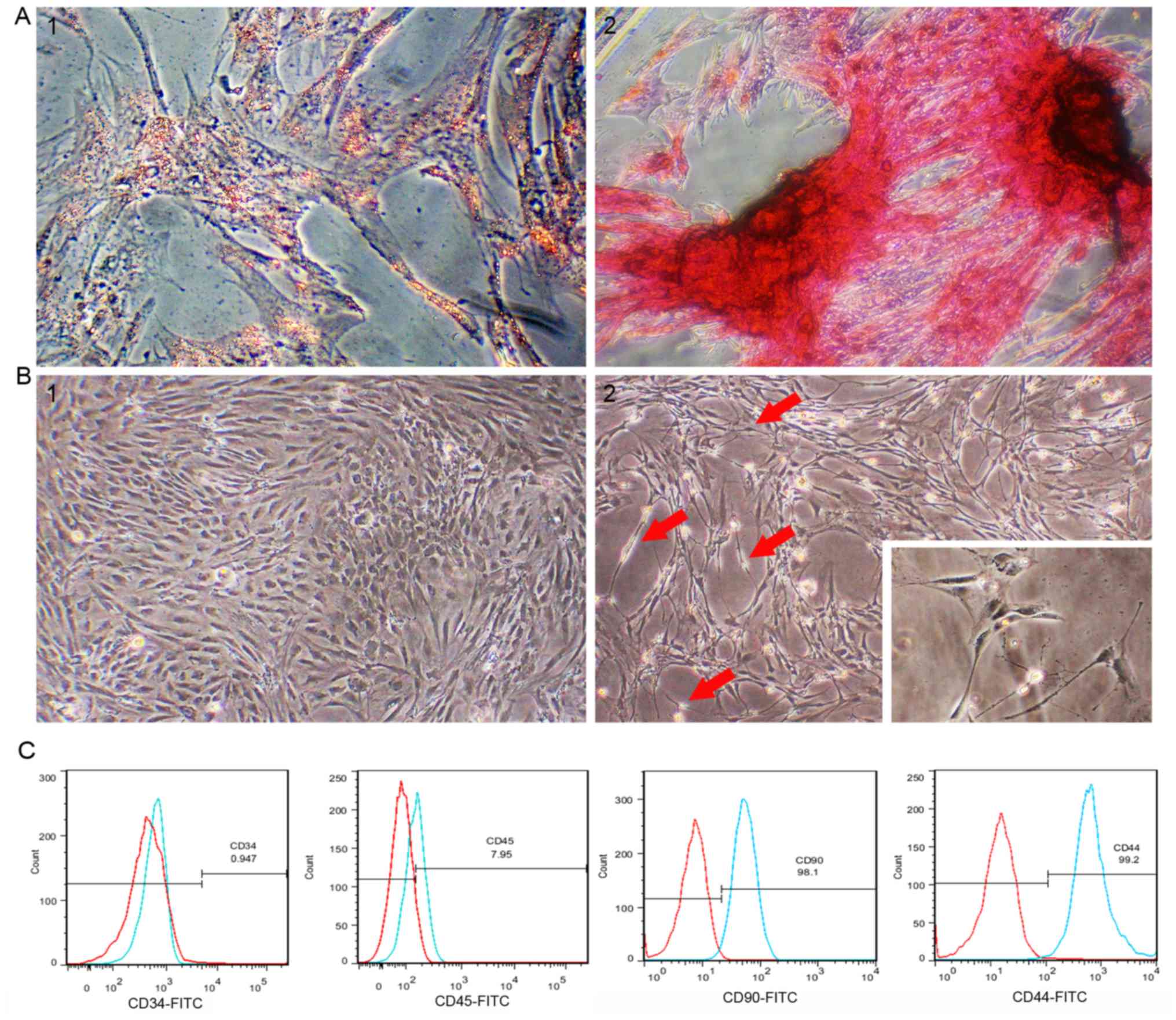
Passage 3 rat ADSCs exhibited a regular spindle
shape (Fig. 1B-1). ADSCs were
treated with induction medium to investigate whether cells isolated
from adipose tissue have the multidirectional differentiation
ability of MSCs. After rat ADSCs were incubated in adipogenic
differentiation medium for 2 weeks, lipid droplets in the cells
accounted for approximately 80% of the cell volume, and adipocyte
differentiation was confirmed by lipid droplet staining with oil
red O (Fig. 1A-1). After 21 days of
culture in osteogenic differentiation medium, osteogenic
differentiation was confirmed by the presence of calcium deposits
stained with von Kossa staining (Fig.
1A-2). Flow cytometry analysis of passage 3 rat ADSCs showed
that the cultured ADSCs were positive for CD44 and CD90, and
negative for CD34 and CD45 (Fig.
1C).
Nerve leachate induces morphological
changes in ADSCs
The cells began to change the cytoplasm from a flat
to contractile shape after treating with nerve leachate for 24 h.
After 48 h, the cells showed a mixture of morphologies, with the
cells changing from long fusiform to a bi- or tri-polar elongated
spindle shapes. After the cells were treated for 96 h, the cells
had morphologies similar to those of SCs, namely cytoplasmic
extension and large nuclei (Fig. 1B-1
and B-2).
Rat health observations
The rats in each group had a good mental state after
surgery and no infections occurred in the wounds. One month after
surgery, the incisions and ulcers of the rats in the nerve
leachate-treated ADSCs + scaffold and autograft groups recovered
better than those of the rats in the scaffold only group. Two
months after surgery, the rats in the nerve leachate-treated ADSCs
+ scaffold and autograft groups were able to walk normally. When
all rats were sacrificed to re-expose the sciatic nerve, the
damaged nerves had regained connectivity in all groups.
Histological assessment of regenerated
nerves
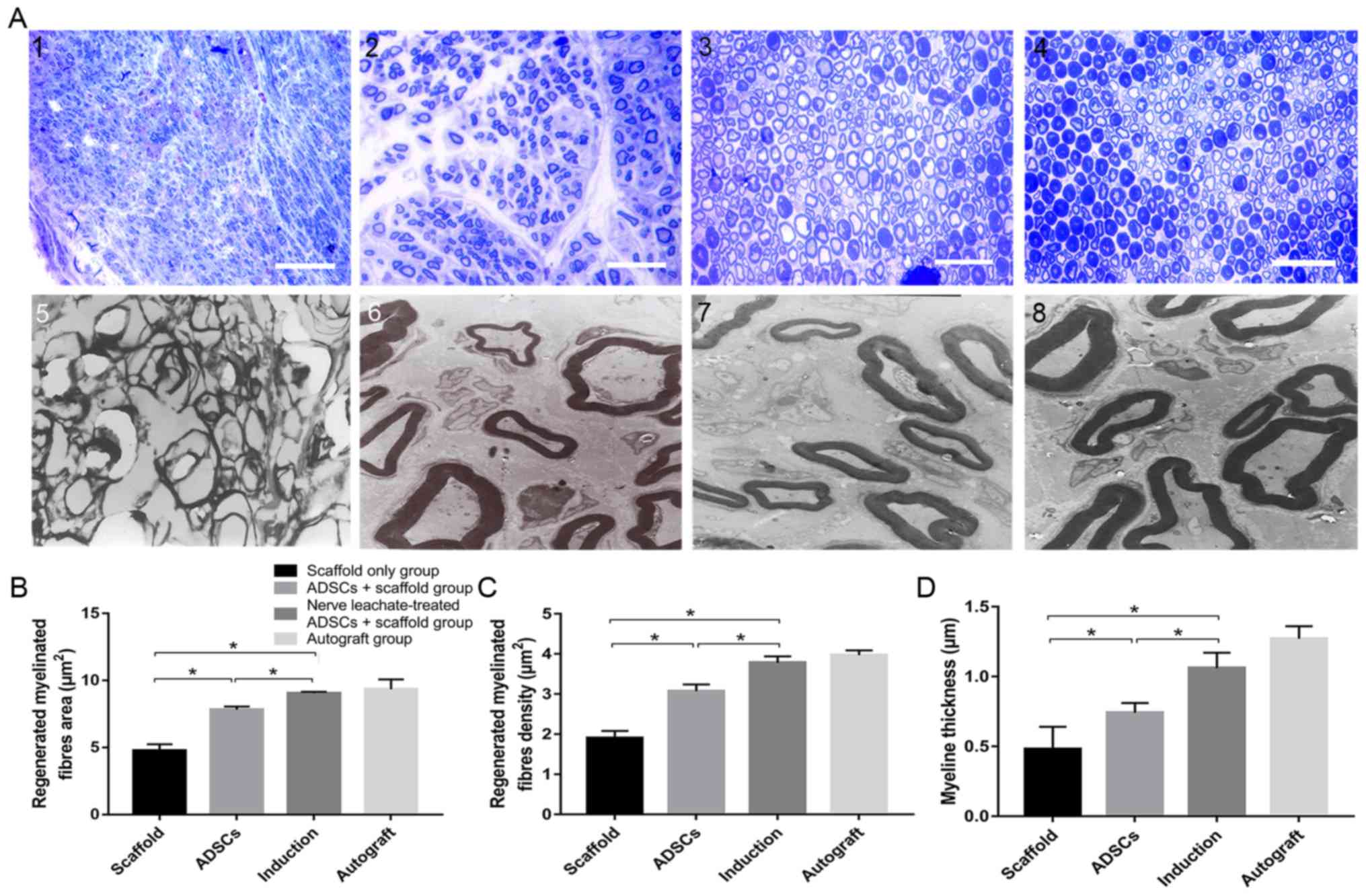
TEM and toluidine blue staining results indicated
that a smaller number of regenerated nerve fibers formed in the
scaffold only group compared with the untreated ADSCs and the nerve
leachate-treated ADSCs + scaffold group, which were irregularly
arranged (Fig. 2A). Several
regenerated nerve fibers were observed in the ADSCs + scaffold
group, but they were less abundant than those in the nerve
leachate-treated ADSCs + scaffold and autograft groups. The largest
numbers of regenerated nerve fibers, which were dispersed in
clusters with thick myelin sheaths, were observed in the nerve
leachate-treated ADSCs + scaffold and autograft groups. Based on
the results of image analysis, the average area of the nerve fibers
in the nerve leachate-treated ADSCs + scaffold group (15.86%), the
nerve fiber density (23.13%), and the nerve fiber thickness
(43.24%) increased significantly compared with the untreated ADSCs
+ scaffold group (Fig. 2B-D).
Electrophysiological assessment of
regenerated nerves
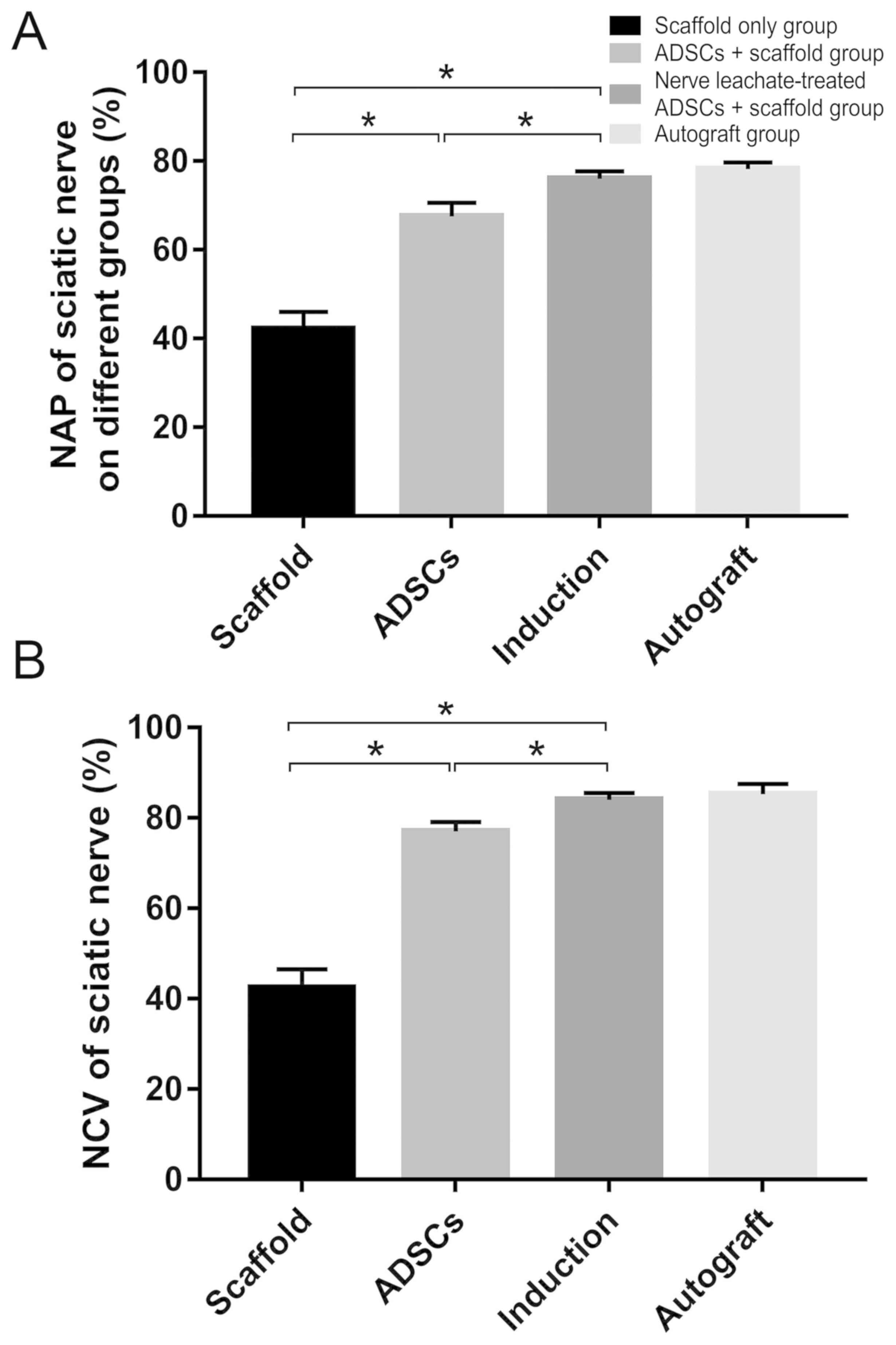
Electrophysiological examination of the regenerated
nerves in each group at 2 months after the operation showed that
the recovery rates of NAP and NCV were significantly reduced in the
scaffold only group compared with those in the untreated ADSCs and
the nerve leachate-treated ADSCs + scaffold group. In addition,
they were higher in the untreated ADSCs + scaffold group compare
with those in the scaffold only group. The rates of NAP and NCV in
the nerve leachate-treated ADSCs + scaffold group were
significantly greater than those in the untreated ADSCs + scaffold
and scaffold only groups. There was no significant difference
between the nerve leachate-treated ADSCs + scaffold group and the
autograft groups (Fig. 3A and
B).
Assessment of the gastrocnemius
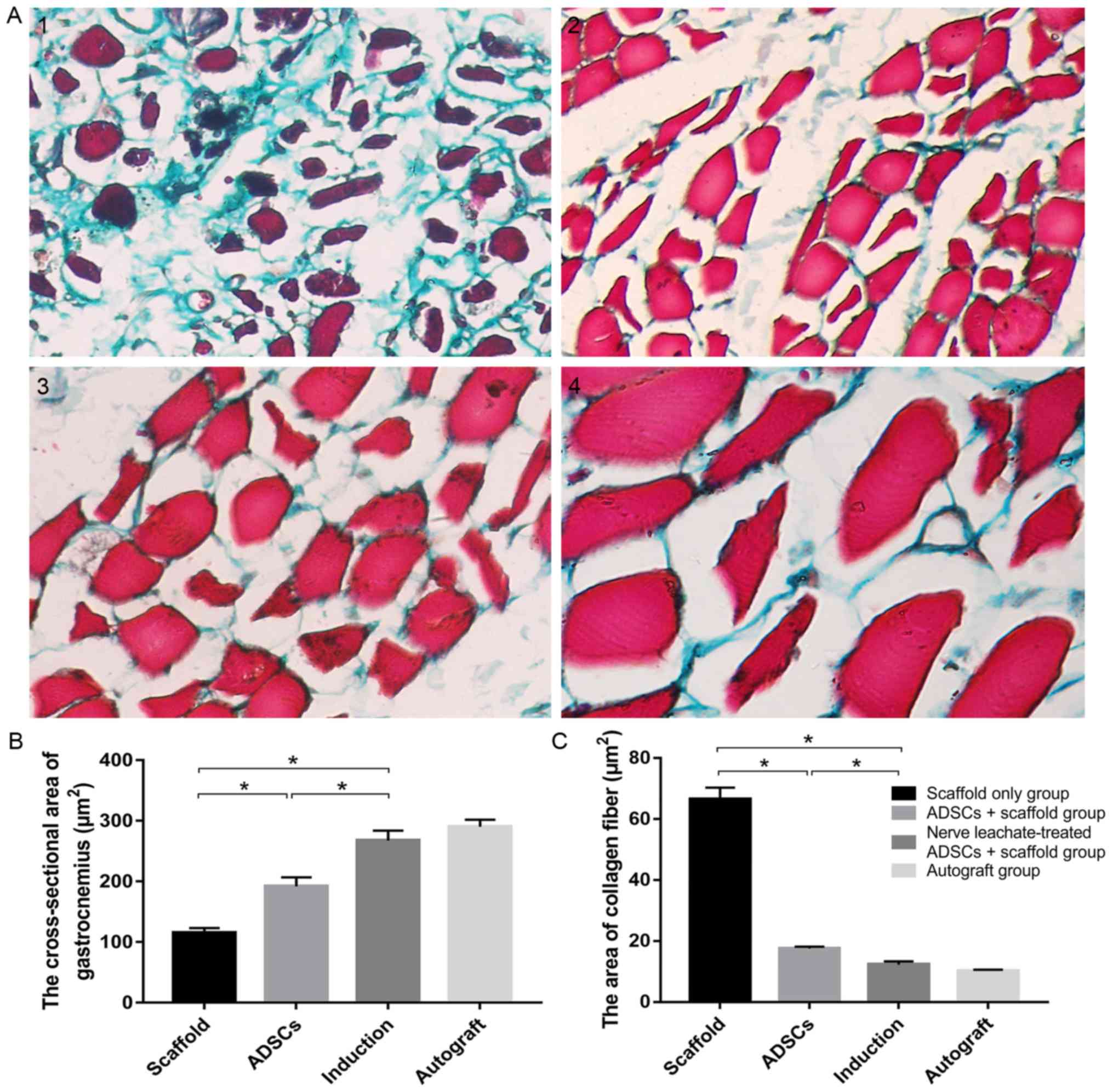
Masson's trichrome stained hyperplastic collagen
fibers green in muscle bundles (Fig.
4A). The number of hyperplastic fibers in the scaffold only
group was markedly increased compared with the untreated ADSCs and
the nerve leachate-treated ADSCs + scaffold group and a lower
number of hyperplastic fibers were observed in the untreated ADSCs
+ scaffold group compared with the scaffold only group.
Hyperplastic fibers were not clearly observed in the nerve
leachate-treated ADSCs and autograft groups. The results of the
image analysis system indicated that the cross-sectional area of
the gastrocnemius and the area of collagen fibers in the scaffold
group were 115.11 and 66.56 µm2, respectively. Compared
with the scaffold only group, the cross-sectional area of
gastrocnemius increased by 66.78% and the area of collagen fibers
decreased by 73.65% in the untreated ADSCs + scaffold group
(Fig. 4B and C). Compared with the
untreated ADSCs + scaffold group, the cross-sectional area of
gastrocnemius, and the area of collagen fibers in the nerve
leachate-treated ADSC + scaffold group increased by 39.28% and
decreased by 29.87%, respectively (Fig.
4B and C). There were no significant differences between the
autograft and the nerve leachate-treated ADSCs + scaffold
groups.
Discussion
In contrast to the central nervous system, the
peripheral nervous system has excellent regeneration and
self-repair abilities (41),
particularly in favorable microenvironments (42). When PNI occurs, SCs undergo
morphological changes to guide axonal regeneration, and trophic
factors and adhesion factors released by SCs promote nerve repair
(43,44). However, the injury site does not
provide a suitable microenvironment, hindering the self-repair
process of long-gap nerve defects (33).
The rapid development of tissue engineering
technology provides opportunities for the repair of PNI. Composite
nerve grafts not only guide axonal regeneration, but also create a
favorable microenvironment to ensure the effective treatment of
nerve endings with various nutrient factors (14). A mature tissue-engineered nerve
scaffold, which can create a better regenerative microenvironment
following PNI, should have the following three components: A
biological scaffold, seed cells and nutrient factors. Biological
scaffolds and seed cells play a crucial role in tissue engineering.
MSCs have been investigated as key seed cells in tissue-engineered
scaffold transplantation due to their multidirectional
differentiation and self-renewal properties (45). BMSCs were the first discovered MSCs
and have been shown to differentiate into SC-like cells under
certain conditions (46,47). ADSCs have similar properties to BMSCs
and are easier to acquire (48).
Previous studies have indicated that ADSCs express CD29, CD44, CD90
and CD105 but are negative for the hematopoietic cell lineage
markers CD34, CD45, CD31 and CD106 (49,50). In
the present study, flow cytometry analysis showed that the isolated
cells were positive for CD44/CD90 but not CD34/CD45, which was
consistent with the phenotypic characteristics of ADSCs. A number
of studies have shown that ADSCs can be differentiated into SC-like
cells using various methods, such as chemical (51,52) and
co-culture methods (53). In both of
these methods ADSCs differentiation into SC-like cells is induced
by neurotrophic factors, including nerve growth factor and
brain-derived neurotrophic factor. These are added to
differentiation medium or directly secreted by SCs (54). Fundamentally, neurotrophic factors
are the key to promoting the differentiation of stem cells into
neuron-like cells (55). In a
previous study, the differentiation of ADSCs into SC-like cells was
induced in vitro by nerve leachate (36).
In the present study, the regeneration of the
sciatic nerve was assessed after a 2-month healing period. This
length of time was chosen as compared with crushing injuries,
transection injuries require more than 3–4 weeks for recovery
(56). In addition, studies have
shown that stem cells are still present and survive at the site of
injury after transplantation of composite nerve conduits at 6 weeks
after PNI (57). The present study
used electrophysiological assessment to indicate the continuity of
the regenerated nerve. The NAP and NCV of the sciatic nerve reached
76.08 and 84.07%, respectively, in the nerve leachate-treated ADSCs
+ scaffold group. These results suggested that a 1-cm injury to the
sciatic nerve was repaired by regenerated nerve and that the
conduction function of the nerve in the nerve leachate treated ADSC
+ scaffold group was superior to that in the untreated ADSC +
scaffold group. Morphological analysis is commonly used to assess
the quality of a regenerative nerve (58). In the present study, the results of
TEM suggested that the number of nerve fibers was greater and that
the myelin sheath was thicker in the nerve leachate-treated ADSCs +
scaffold group. The same result was observed with toluidine blue
staining. When peripheral nerves are damaged, muscle atrophy occurs
in the gastrocnemius muscle (10),
which is controlled by the sciatic nerve. Thus, the recovery of
peripheral nerves was evaluated by observing pathological changes
in the gastrocnemius muscle stained with Masson's trichrome
(59). No obvious pathological
changes were seen in the gastrocnemius muscle tissue in the nerve
leachate-treated ADSCs + scaffold group. Moreover, the
cross-sectional area of the gastrocnemius muscle in the nerve
leachate-treated ADSCs + scaffold group was larger compared with
the untreated ADSCs and the scaffold only group, indicating that
the degree of gastrocnemius atrophy in the rats from this group was
slight. The gastrocnemius muscles in the scaffold only group had a
large area of collagen fibers, and muscle atrophy in the scaffold
only group was more obvious compared the nerve leachate-treated
ADSCs + scaffold and the untreated ADSCs group. This demonstrated
that somatic function dominated by the sciatic nerve recovered
effectively in the nerve leachate-treated ADSCs + scaffold
group.
Autograft was chosen as a positive control because
autografting is the gold standard therapeutic method for peripheral
nerve repair in the clinic (59).
The results of the present study indicated that the recovery level
in the nerve leachate-treated ADSCs + scaffold group was similar to
that in the autograft group. As expected, the composite nerve
scaffolds containing nerve leachate-treated ADSCs resulted in
better functional and structural recovery during the treatment of
PNI than the scaffold only and the untreated ADSCs + scaffold
group. ADSCs treated with nerve leachate may have therapeutic
potential for PNI.
Previous studies have suggested that SCs undergo
dedifferentiation, and their secretion of factors, such as
neurotrophic factors, brain-derived neurotrophic factor (60,61),
neuronal growth factor (62) and
cell adhesion factors, increases after PNI. These factors regulate
the structure and function of the nervous system and play a key
role in the repair of PNI (63). In
the present study, rat sciatic nerves were cut and used to prepare
a nerve leachate. Results suggested that ADSCs induced with this
leachate may play an effective therapeutic role in PNI. This
leachate may contain various neurotrophic factors released by the
nerve ends that induce the differentiation of ADSCs. In previous
work, nerve leachate was prepared from the sciatic nerves of
animals, such as rabbits, dogs and frogs, and was also shown to
induce the differentiation of ADSCs into SC-like cells (unpublished
data). In addition, research revealed that nerve leachate from the
sciatic nerves of cattle induced the neural differentiation of rat
PC12 cells (60). The above results
indicated that there is no species limitation for the source of
nerve leachate that can be used to induce the differentiation of
ADSCs. In addition, the nerves from different animals can be
obtained for the preparation of nerve leachate from animal feed
houses and slaughterhouses, such as barns and pig farms. During the
preparation of nerve leachate, in order to ensure the consistent
quality, animals of the same species with similar body weight were
selected, where the sciatic nerve segment of the same length and
diameter were taken and the preparation method of the nerve
leachate was also kept consistent. However, although it was
demonstrated in the present study to exert positive effects,
differences in the nerve leachate from different species were not
determined. The composition of the nerve is complex due to the
presence of a variety of positive nerve factors (32,60,61). Any
future research should focus on the study of characterizing and
comparing the components of the nerve leachate from different
species. Co-culture and chemical induction methods to induce the
differentiation of ADSCs into SC-like cells exist, but both methods
have limitations. For example, co-culture is cumbersome, and some
of the neurotrophic factors for chemical induction are expensive.
In the present study, preparation of nerve leachate was simple and
used abundant biological materials.
Acknowledgements
No applicable.
Funding
This work was supported by grants from the National
Natural Science Foundation of China (grant nos. U1504325 and
31101779) and the Undergraduate Training Programs in Henan
University of Science and Technology (grant no. 2018380).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL, RD and ZZ conceived and designed the study. RD,
CZ, YY, YX, HW, MZ, JZ, YW and YS performed the experiments. YL and
RD wrote the manuscript. The manuscript was reviewed and edited by
ZZ. All authors have read and approved this manuscript.
Ethics approval and consent to
participate
All animal care and experimental protocols were
conducted according to the University Policies on the Use and Care
of Animals and were approved by the Institutional Animal Experiment
Committee of Henan University of Science and Technology (Henan,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ADSCs
|
adipose-derived mesenchymal stem
cells
|
|
MSCs
|
mesenchymal stem cells
|
|
PNI
|
peripheral nerve injury
|
|
SCs
|
Schwann cells
|
References
|
1
|
Campbell WW: Evaluation and management of
peripheral nerve injury. Clin Neurophysiol. 119:1951–1965. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Conforti L, Gilley J and Coleman MP:
Wallerian degeneration: An emerging axon death pathway linking
injury and disease. Nat Rev Neurosci. 15:394–409. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Faroni A, Mobasseri SA, Kingham PJ and
Reid AJ: Peripheral nerve regeneration: Experimental strategies and
future perspectives. Adv Drug Deliv Rev. 82-83:160–167. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang X, Luo E, Li Y and Hu J: Schwann-like
mesenchymal stem cells within vein graft facilitate facial nerve
regeneration and remyelination. Brain Res. 1383:71–80. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dubey N, Letourneau PC and Tranquillo RT:
Guided neurite elongation and schwann cell invasion into
magnetically aligned collagen in simulated peripheral nerve
regeneration. Exp Neurol. 158:338–350. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen ZL, Yu WM and Strickland S:
Peripheral regeneration. Annu Rev Neurosci. 30:209–233. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mirsky R, Jessen KR, Brennan A, Parkinson
D, Dong Z, Meier C, Parmantier E and Lawson D: Schwann cells as
regulators of nerve development. J Physiol Paris. 96:17–24. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Matsuyama T, Mackay M and Midha R:
Peripheral nerve repair and grafting techniques: A review. Neurol
Med Chir (Tokeyo). 40:187–199. 2000. View Article : Google Scholar
|
|
9
|
Griffin JW, Hogan MV, Chhabra AB and Deal
DN: Peripheral nerve repair and reconstruction. J Bone Joint Surg
Am. 95:2144–2151. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brull R, Hadzic A, Reina MA and Barrington
MJ: Pathophysiology and etiology of nerve injury following
peripheral nerve blockade. Reg Anesth Pain Med. 40:479–490. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Scheib J and Hoke A: Advances in
peripheral nerve regeneration. Nat Rev Neurol. 9:668–676. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Trehan SK, Model Z and Lee SK: Nerve
repair and nerve grafting. Hand Clin. 32:119–125. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brooks DN, Weber RV, Chao JD, Rinker BD,
Zoldos J, Robichaux MR, Ruggeri SB, Anderson KA, Bonatz EE,
Wisotsky SM, et al: Processed nerve allografts for peripheral nerve
reconstruction: A multicenter study of utilization and outcomes in
sensory, mixed, and motor nerve reconstructions. Microsurgery.
32:1–14. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sarker MD, Naghieh S, McInnes AD, Schreyer
DJ and Chen X: Regeneration of peripheral nerves by nerve guidance
conduits: Influence of design, biopolymers, cells, growth factors,
and physical stimuli. Prog Neurobiol. 171:125–150. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hayashi A, Moradzadeh A, Tong A, Wei C,
Tuffaha SH, Hunter DA, Tung TH, Parsadanian A, Mackinnon SE and
Myckatyn TM: Treatment modality affects allograft-derived Schwann
cell phenotype and myelinating capacity. Exp Neurol. 212:324–336.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fu X, Tong Z, Li Q, Niu Q, Zhang Z, Tong
X, Tong L and Zhang X: Induction of adipose-derived stem cells into
Schwann-like cells and observation of Schwann-like cell
proliferation. Mol Med Rep. 14:1187–1193. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shen M, Tang W, Cao Z, Cao X and Ding F:
Isolation of rat Schwann cells based on cell sorting. Mol Med Rep.
16:1747–1752. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rodriguez Sanchez DN, de Lima Resende LA,
Boff Araujo Pinto G, de Carvalho Bovolato AL, Possebon FS, Deffune
E and Amorim RM: Canine adipose-derived mesenchymal stromal cells
enhance neuroregeneration in a rat model of sciatic nerve crush
injury. Cell Transplant. 28:47–54. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu H, Lv P, Wu H, Zhang K, Xu F, Zheng L
and Zhao J: The proliferation enhancing effects of salidroside on
schwann cells in vitro. Evid Based Complement Alternat Med.
2017:46732892017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou W, Stukel JM, Cebull HL and Willits
RK: Tuning the mechanical properties of poly(Ethylene Glycol)
microgel-based scaffolds to increase 3D schwann cell proliferation.
Macromol Biosci. 16:535–544. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Uz M, Buyukoz M, Sharma AD, Sakaguchi DS,
Altinkaya SA and Mallapragada SK: Gelatin-based 3D conduits for
transdifferentiation of mesenchymal stem cells into Schwann
cell-like phenotypes. Acta Biomater. 53:293–306. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tabatabaei Qomi R and Sheykhhasan M:
Adipose-derived stromal cell in regenerative medicine: A review.
World J Stem Cells. 9:107–117. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mazini L, Rochette L, Amine M and Malka G:
Regenerative capacity of adipose derived stem cells (ADSCs),
comparison with mesenchymal stem cells (MSCs). Int J Mol Sci.
20:E25232019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
di Summa PG, Kingham PJ, Raffoul W, Wiberg
M, Terenghi G and Kalbermatten DF: Adipose-derived stem cells
enhance peripheral nerve regeneration. J Plast Reconstr Aesthet
Surg. 63:1544–1552. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kolar MK and Kingham PJ: Regenerative
effects of adipose-tissue-derived stem cells for treatment of
peripheral nerve injuries. Biochem Soc Trans. 42:697–701. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schilling BK, Schusterman MA II, Kim DY,
Repko AJ, Klett KC, Christ GJ and Marra KG: Adipose-derived stem
cells delay muscle atrophy after peripheral nerve injury in the
rodent model. Muscle Nerve. 59:603–610. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Erba P, Mantovani C, Kalbermatten DF,
Pierer G, Terenghi G and Kingham PJ: Regeneration potential and
survival of transplanted undifferentiated adipose tissue-derived
stem cells in peripheral nerve conduits. J Plast Reconstr Aesthet
Surg. 63:e811–e817. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Georgiou M, Golding JP, Loughlin AJ,
Kingham PJ and Phillips JB: Engineered neural tissue with aligned,
differentiated adipose-derived stem cells promotes peripheral nerve
regeneration across a critical sized defect in rat sciatic nerve.
Biomaterials. 37:242–251. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hei WH, Kim S, Park JC, Seo YK, Kim SM,
Jahng JW and Lee JH: Schwann-like cells differentiated from human
dental pulp stem cells combined with a pulsed electromagnetic field
can improve peripheral nerve regeneration. Bioelectromagnetics.
37:163–174. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bayat N, Ebrahimi-Barough S, Ardakan MM,
Ai A, Kamyab A, Babaloo H and Ai J: Differentiation of human
endometrial stem cells into schwann cells in fibrin hydrogel as 3D
culture. Mol Neurobiol. 53:7170–7176. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tohill M, Mantovani C, Wiberg M and
Terenghi G: Rat bone marrow mesenchymal stem cells express glial
markers and stimulate nerve regeneration. Neurosci Lett.
362:200–203. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jessen KR and Mirsky R: The repair Schwann
cell and its function in regenerating nerves. J Physiol.
594:3521–3531. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dowsing BJ, Morrison WA, Nicola NA,
Starkey GP, Bucci T and Kilpatrick TJ: Leukemia inhibitory factor
is an autocrine survival factor for Schwann cells. J Neurochem.
73:96–104. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Meier C, Parmantier E, Brennan A, Mirsky R
and Jessen KR: Developing Schwann cells acquire the ability to
survive without axons by establishing an autocrine circuit
involving insulin-like growth factor, neurotrophin-3, and
platelet-derived growth factor-BB. J Neurosci. 19:3847–3859. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Weiner JA and Chun J: Schwann cell
survival mediated by the signaling phospholipid lysophosphatidic
acid. Proc Natl Acad Sci USA. 96:5233–5238. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu Y, Zhang Z, Qin Y, Wu H, Lv Q, Chen X
and Deng W: A new method for Schwann-like cell differentiation of
adipose derived stem cells. Neurosci Lett. 551:79–83. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Y, Luo H, Zhang Z, Lu Y, Huang X,
Yang L, Xu J, Yang W, Fan X, Du B, et al: A nerve graft constructed
with xenogeneic acellular nerve matrix and autologous
adipose-derived mesenchymal stem cells. Biomaterials. 31:5312–5324.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Luo H, Zhang Y, Zhang Z and Jin Y: The
protection of MSCs from apoptosis in nerve regeneration by TGFbeta1
through reducing inflammation and promoting VEGF-dependent
angiogenesis. Biomaterials. 33:4277–4287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gu X, Ding F and Williams DF: Neural
tissue engineering options for peripheral nerve regeneration.
Biomaterials. 35:6143–6156. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang Y, Li D, Wang G, Chen L, Chen J, Liu
Z, Zhang Z, Shen H, Jin Y and Shen Z: The effect of
co-transplantation of nerve fibroblasts and Schwann cells on
peripheral nerve repair. Int J Biol Sci. 13:1507–1519. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cattin AL and Lloyd AC: The multicellular
complexity of peripheral nerve regeneration. Curr Opin Neurobiol.
39:38–46. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lehmann HC and Hoke A: Use of engineered
Schwann cells in peripheral neuropathy: Hopes and hazards. Brain
Res. 1638:97–104. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
La Noce M, Mele L, Tirino V, Paino F, De
Rosa A, Naddeo P, Papagerakis P, Papaccio G and Desiderio V: Neural
crest stem cell population in craniomaxillofacial development and
tissue repair. Eur Cells Mater. 28:348–357. 2014. View Article : Google Scholar
|
|
44
|
Lee JK, Choi IS, Oh TI and Lee E:
Cell-surface engineering for advanced cell therapy. Chemistry.
24:15725–15743. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Orbay H, Uysal AC, Hyakusoku H and Mizuno
H: Differentiated and undifferentiated adipose-derived stem cells
improve function in rats with peripheral nerve gaps. J Plast
Reconstr Aesthet Surg. 65:657–664. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ladak A, Olson J, Tredget EE and Gordon T:
Differentiation of mesenchymal stem cells to support peripheral
nerve regeneration in a rat model. Exp Neurol. 228:242–252. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cai S, Tsui YP, Tam KW, Shea GK, Chang RS,
Ao Q, Shum DK and Chan YS: Directed differentiation of human bone
marrow stromal cells to fate-committed schwann cells. Stem Cell
Reports. 9:1097–1108. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li X, Wang M, Jing X, Guo W, Hao C, Zhang
Y, Gao S, Chen M, Zhang Z, Zhang X, et al: Bone marrow- and adipose
tissue-derived mesenchymal stem cells: Characterization,
differentiation, and applications in cartilage tissue engineering.
Crit Rev Eukaryot Gene Expr. 28:285–310. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Huang SJ, Fu RH, Shyu WC, Liu SP, Jong GP,
Chiu YW, Wu HS, Tsou YA, Cheng CW and Lin SZ: Adipose-derived stem
cells: Isolation, characterization, and differentiation potential.
Cell Transplant. 22:701–709. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Dubey NK, Mishra VK, Dubey R, Deng YH,
Tsai FC and Deng WP: Revisiting the advances in isolation,
characterization and secretome of adipose-derived stromal/stem
cells. Int J Mol Sci. 19:E22002018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lei Z, Yongda L, Jun M, Yingyu S, Shaoju
Z, Xinwen Z and Mingxue Z: Culture and neural differentiation of
rat bone marrow mesenchymal stem cells in vitro. Cell Biol Int.
31:916–923. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kingham PJ, Mantovani C and Terenghi G:
Notch independent signalling mediates Schwann cell-like
differentiation of adipose derived stem cells. Neurosci Lett.
467:164–168. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li X, Liao D, Gong P, Yuan Q and Tan Z:
Neural differentiation of adipose-derived stem cells by indirect
co-culture with Schwann cells. Arch Biol Sci. 61:703–711. 2009.
View Article : Google Scholar
|
|
54
|
Jiang L, Jones S and Jia X: Stem cell
transplantation for peripheral nerve regeneration: Current options
and opportunities. Int J Mol Sci. 18:E942017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Pan Y and Cai S: Current state of the
development of mesenchymal stem cells into clinically applicable
Schwann cell transplants. Mol Cell Biochem. 368:127–135. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yousefi F, Lavi Arab F, Nikkhah K, Amiri H
and Mahmoudi M: Novel approaches using mesenchymal stem cells for
curing peripheral nerve injuries. Life Sci. 221:99–108. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Sun AX, Prest TA, Fowler JR, Brick RM,
Gloss KM, Li X, DeHart M, Shen H, Yang G, Brown BN, et al: Conduits
harnessing spatially controlled cell-secreted neurotrophic factors
improve peripheral nerve regeneration. Biomaterials. 203:86–95.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Cui Y, Yao Y, Zhao Y, Xiao Z, Cao Z, Han
S, Li X, Huan Y, Pan J and Dai J: Functional collagen conduits
combined with human mesenchymal stem cells promote regeneration
after sciatic nerve transection in dogs. J Tissue Eng Regen Med.
12:1285–1296. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kemp SW, Cederna PS and Midha R:
Comparative outcome measures in peripheral regeneration studies.
Exp Neurol. 287:348–357. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhang Z, Liu Y, Zhu X, Wei L, Zhu J, Shi
K, Wang G and Pan L: Sciatic nerve leachate of cattle causes
neuronal differentiation of PC12 cells via ERK1/2 signaling
pathway. J Vet Sci. 19:512–518. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Shakhbazau A, Martinez JA, Xu QG, Kawasoe
J, van Minnen J and Midha R: Evidence for a systemic regulation of
neurotrophin synthesis in response to peripheral nerve injury. J
Neurochem. 122:501–511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Nurcombe V, Hill MA, Eagleson KL and
Bennett MR: Motor neuron survival and neuritic extension from
spinal cord explants induced by factors released from denervated
muscle. Brain Res. 291:19–28. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Jessen KR, Mirsky R and Lloyd AC: Schwann
cells: Development and role in nerve repair. Cold Spring Harb
Perspect Biol. 7:a0204872015. View Article : Google Scholar : PubMed/NCBI
|