Introduction
Lung cancer is a respiratory disease, which is
associated with global air contamination (1). Non-small cell lung cancer (NSCLC) is a
type of cancer that initiates in the non-small cells of the lung
and occupies the highest occurrence rate among all cancer cases
(2). NSCLC, including squamous cell
carcinoma, large cell carcinoma and adenocarcinoma, accounts for
>80% of all lung cancer cases (3–5).
Although studies have provided therapeutic improvements for NSCLC,
the poor survival rate (overall 5-year survival rate, <15%) of
patients with NSCLC is primarily attributed to critical clinical
problems, including tumor cell diffusion and metastasis (4,6,7). Migration and invasion of NSCLC is the
primary cause of the poor survival rate during treatment and
recurrence for patients with NSCLC (8,9).
Therefore, investigating effective agents that inhibit migration
and invasion and providing individualized medication for NSCLC has
become the focus for treating patients with cancer (10,11).
Lenvatinib is a multi-targeted tyrosine kinase
inhibitor targeting fibroblast growth factor receptor (FGFR)1-4,
platelet derived growth factor receptor (PDGFR)-β, Ret, vascular
endothelial growth factor receptor (VEGFR)1–3 and Kit and is used
to treat patients (12,13). PDGFR-β, Ret, FGFR1-4, VEGF and
Kit-mediated angiogenesis have been demonstrated to target
molecules associated with the progression and neoplasm metastasis
of NSCLC (14). In addition, the
antitumor activities of lenvatinib against various human tumors
have been assessed in clinical and preclinical experiments to
confirm efficacy (15,16). Target agents that suppressed the VEGF
signal pathway have demonstrated beneficial outcomes in NSCLC,
which suggests that the VEGF signal pathway is important and
lenvatinib is an effective anticancer drug (17). Furthermore, lenvatinib has also
demonstrated clinical benefits via the targeting of PDGFR-β, Ret,
VEGFR1-3, FGFR1-4 and Kit pathways in metastatic renal cell
carcinoma (18,19). A previous study indicated that
lenvatinib administration with carboplatin and paclitaxel extended
the overall survival in patients with NSCLC (14). However, the association between
lenvatinib and migration and invasion remains unknown.
Dexamethasone is commonly applied as an anti-emetic
drug in combination with chemotherapy. A previous study evaluated
the pharmacokinetics and safety of drugs combined with
dexamethasone in patients with multiple myeloma and renal
impairment (20). Han et al
(21) reported that dexamethasone
inhibited transforming growth factor (TGF)-β1-induced cell
migration by regulating the extracellular signal-regulated kinases
(ERK) and protein kinase B (AKT) pathways in human colon cancer
cells via the cysteine-rich angiogenic inducer 61 (CYR61). CYR61 is
a member of the CYR61/connective tissue growth
factor/nephroblastoma overexpressed protein family, which is
mediated in cellular adhesion, survival, migration, mitogenesis,
differentiation, proliferation, invasion, survival and angiogenesis
and the metastasis of cancer cells (22). CYR61 may have an essential role as an
oncogene and a tumor suppressor for suppressing angiogenesis by
supplying oxygen and nutrients to tumor cells (23).
The aim of the present study was to elucidate the
molecular mechanism of migration and invasion in NSCLC progression
and investigate the synergistic effects of TGF and dexamethasone on
NSCLC for improved therapy. In addition, the therapeutic outcomes
and molecular mechanism were investigated via cooperative treatment
with lenvatinib and dexamethasone, which inhibited human NSCLC
migration and invasion via mediated EKR/AKT and VEGF signaling
pathways.
Materials and methods
Cell culture
H1975 and H358 cells were purchased from American
Type Culture Collection (Manassas, VA, USA). H1975 and H358 cells
were cultured in RPMI 1640 medium supplemented with 10% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) at 37°C in an atmosphere containing 5% CO2 and
reasonable humidity (45–60%).
MTT assay for viability
H1975 and H358 cells were cultured in 96-well plates
to form a ~90% monolayer. Subsequently, dexamethasone, TGF-β1 (20,
40 and 100 mg/ml), lenvatinib (20, 40 and 100 mg/ml) or
dexamethasone (20, 40 and 100 mg/ml plus TGF-β1 (20, 40 and 100
mg/ml) were added into cells for 12 h at 37°C (all Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany). A total of 10 µl MTT at a
concentration of 5 mg/ml (Amresco LLC, Solon, OH, USA) was added to
the cells and incubated for 4 h at 37°C. Subsequently, dimethyl
sulfoxide was added for incubation for 30 min at 37°C to dissolve
the precipitate, following the removal of the supernatant. The
results were determined using a spectrophotometer (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) at 570 nm.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from H1975 and H358 cells
prior to or following treatment with TGF-β1, lenvatinib or
dexamethasone using an RNAeasy Mini kit (Qiagen Sciences, Inc.,
Gaithersburg, MD, USA). Total RNA (1 µg) was reverse transcribed
into cDNA using an RT kit (Qiagen Sciences, Inc.) and the quality
was confirmed by 2% agarose gel electrophoresis. Template cDNA (10
ng) was subjected to qPCR using a SYBR Green Master Mix (Bio-Rad
Laboratories, Inc.). PCR was performed using the following
conditions: Preliminary denaturation at 94°C for 2 min, 45 cycles
of 94°C for 30 sec, the annealing temperature was reduced to 56°C
for 30 sec and a final step of 72°C for 10 min. All the forward and
reverse primers were synthesized by Invitrogen (Table I) (Thermo Fisher Scientific, Inc.).
Relative mRNA expression changes were calculated according to the
2−ΔΔCq method (24).
Results were expressed as the n-fold β-actin compared with the
relative control.
 | Table I.Sequences of primers used in the
study. |
Table I.
Sequences of primers used in the
study.
| Gene | Sequence reverse
(5′-3′) | Forward |
|---|
| CYR61 |
ATCGAGATCTGGAGAAGGCGGAG GGCGCGG |
ATCGAGATCTAATGGAGCCAGGGGAGGCG |
| VEGFR1 |
AAGAGAGCTTCCGTAAGGCG |
GCATCCTCTTCAGTTACGTCC |
| VEGFR2 |
GGAAGCTCCTGAAGATCTGT |
GAGGATATTTCGTGCCGCGC |
| VEGFR3 |
AGTCACACGTCATCGACACC |
ATTGGGACAGCTTGGATCAC |
| FGFR1 |
CACATCGAGGTGAACGGGAGTAAG |
CGCATCCTCAAAGGAGACATTCC |
| FGFR2 |
GGAAAGTGTGGTCCCATCTGA |
TCCAGGTGGTACGTGTGATTG |
| FGFR3 |
CATTGGAGGCATAAGCTG |
AGCACGGTAACGTAGGGTGT |
| FGFR4 |
GCAACTCCATCGGCCTTTCCTACCAG |
AGAACCAGTGAGCCTGATACATACAG |
| Vimentin
E-cadherin |
TGGAGGAATTCTTGCTTTGC |
CGTACATGTCAGCCAGCTTC |
| Slug |
TATTTGGTTGGTCAGCACAGG |
GACGCAATCAATGTTTACTCG |
| MMP-1 |
TTCCACAGGTCCCACAAC |
GCATTCCTCACAGCCAAC |
| MMP-9 |
TGGGCTACGTGACCTATGACAT |
GTATGGTCGTGGCTCTAAGC |
| CPI |
GGGATTCCCTGGACCTAAAG |
GGAACACCTCGCTCTCCA |
| Fibronectin |
CCAGGCAGTACAATGTGGGT |
TGGAATAGAGCTCCCAGGCT |
| Cytochalasin-D |
GGTATTCAGCCAAACGACCA |
CCTCTCACTCGGTTCTCGAT |
| G-actin |
CCAGGGCTTTTCAAAAATGA |
CCGATCCAATCTGTTCTGGT |
| β-actin |
CGGAGTCAACGGATTTGGTC |
AGCCTTCTCCATGGTCGTGA |
Cell viability assay
H1975 and H358 cells were cultured in 24-well plates
at a concentration of 1×106/ml and a total column volume
of 500 µl. Following overnight culture, H1975 and H358 cells were
treated with TGF-β1 (20, 40 and 100 mg/ml), lenvatinib (20, 40 and
100 mg/ml) or dexamethasone (20, 40 and 100 mg/ml plus TGF-β1 (20,
40 and 100 mg/ml) or PBS as a control. These concentrations were
selected as they maximally inhibited cell growth, as confirmed as
aforementioned. The viability of H1975 and H358 cells was examined
using a Cell Counting kit-8 according to manufacturer's protocol
(Dojindo Molecular Technologies, Inc., Kumamoto, Japan) following
12 h.
Cell invasion and migration
assays
H1975 cells were treated with TGF-β1, lenvatinib or
dexamethasone and PBS-treated cells were used as control. For the
invasion assay, cells were suspended at a density of
1×105 in 500 µl serum-free Dulbecco's modified Eagle's
medium (Gibco; Thermo Fisher Scientific, Inc.). H1975 cells were
treated with TGF-β1, lenvatinib or dexamethasone and inserted into
the top of a BD BioCoat Matrigel Invasion Chamber (BD Biosciences,
Franklin Lakes, NJ, USA) according to the manufacturer's protocol.
For the migration assay, H1975 cells were inoculated with TGF-β1,
lenvatinib or dexamethasone for 12 h at 37°C and a control insert
(BD Biosciences) instead of a Matrigel Invasion Chamber. Cells were
fixed with 4% paraformaldehyde for 30 min and then stained with
0.2% crystal violet for 30 min at 37°C (Sigma-Aldrich; Merck KGaA).
The tumor cells that had successfully invaded and migrated were
counted in at least three randomly selected stained-fields using
light microscopy (Olympus BX51; Olympus Corporation, Tokyo, Japan)
for each membrane at magnification, ×40.
Scratch wound healing assays
H1975 and H358 cells were cultured in RPMI 1640
medium supplemented with 10% FBS until a 50–60% monolayer of cells
was observed in the culture dishes. A sterile pipette tip was
scratched across the surface of the culture dish to disturb the
monolayer. TGF-β1 (20 ng/ml), dexamethasone (20 µM) and combined
treatment (20 ng/ml TGF-β1 and 20 µM dexamethasone) were added into
the RPMI-1640 medium following surface scratching. Images of H1975
and H358 cells were captured 12 h post-treatment using a digital
camera (DSC-RXO; Sony Corporation, Tokyo, Japan). ImageJ software
(version 3.60; National Institutes of Health, Bethesda, MA, USA)
was used to analyze the scratches.
Western blot analysis
H1975 and H358 cells were treated with lenvatinib
(40 ng/ml), TGF-β1 (40 ng/ml), dexamethasone (20 µM) and combined
treatment for 12 h. H1975 and H358 cells were harvested by
scraping, lysed in RIPA buffer and homogenized at 4°C for 10 min.
Proteins (20 µg) were separated by 12% SDS-PAGE assays followed by
transfer to polyvinylidene fluoride membranes (EMD Millipore
Billerica, MA, USA). The membranes were blocked with 5% bovine
serum albumin (Sigma-Aldrich; Merck KGaA) at 4°C for 12 h and then
incubated with ERK (cat. no. ab196883), AKT (cat. no. ab8805),
Snail (cat. no. ab180714), VEGFR1 (cat. no. ab36844), VEGFR2 (cat.
no. ab2349), VEGFR3 (cat. no. ab27278), FGFR1-4 (cat. no. ab10646),
FGFR2 (cat. no. ab10648), FGFR3 (cat. no. ab137084), FGFR4 (cat.
no. ab178396),(all dilution 1:1,000) and β-actin (dilution 1:2,000;
cat. no. ab8226) primary antibodies for 12 h at 4°C. All primary
antibodies were purchased from Abcam (Cambridge, MA, USA). The
membranes were subsequently incubated with secondary rabbit
anti-mouse antibodies (dilution 1:5,000; cat. no. PI-1000; Vector
Laboratories, Inc.) for 2 h at 37°C. Protein expression was
analyzed using a biotin-labeled chemi-luminescence detection system
(cat. no. Z370398; Sigma-Aldrich; Merck KGaA). Densitometric
quantification of the immunoblot data was performed using
Quantity-One 1.1 (Bio-Rad Laboratories, Inc.) software.
Apoptosis assay
H1975 (1×106) and H358 (1×106)
cells were treated with lenvatinib (40 ng/ml), TGF-β1 (40 ng/ml),
dexamethasone (20 µM) or combined treatment for 12 h at 37°C. The
cells were incubated with PBS and the apoptosis of the suspended
cells was analyzed by flow cytometry. Cells were collected and
suspended with Annexin V-FITC and PI (Annexin V-FITC/PI Apoptosis
Detection kit; BD Biosciences) for 30 min at 4°C according to the
manufacturer protocol. Fluorescence was measured using a
fluorescence-activated cell sorting flow cytometer (FCS
Express™ 4 IVD, BD Biosciences) and analyzed using
Quantity One software (version 3.0; Bio-Rad Laboratories,
Inc.).
Animal study
A total of 80 specific pathogen-free female C57BL/6
(6-week-old; 26–32 g) mice were purchased from Shanghai SLAC
Laboratory Animal Co., Ltd. (Shanghai, China). All mice were
treated in accordance with the Guide for the Care and Use of
Laboratory Animals of Tianjin Chest Hospital (Tianjin, China) and
the study was approved by the Ethics Committee of Tianjin Chest
Hospital (Tianjin, China). All mice were housed under controlled
temperatures (23±2°C) in 50% humidity, in a 12 h light/dark cycle
with free access to food and water. Mice were subcutaneously
implanted with H1975 tumor cells (1×107) using a syringe
and divided into four groups (n=20 per group). Treatment was
initiated on day 7 following tumor implantation when the tumor
diameter reached 5–6 mm. Xenograph mice were intravenously injected
with 500 ng lenvatinib, 500 ng dexamethasone or synergistic
treatment with PBS as control. Treatment was continued once a day
for 14 days. Tumor volumes were calculated according to a previous
study (25).
Statistical analysis
All data were presented as the mean ± standard error
of the mean from triplicate experiments. All data were analyzed
using SPSS Statistics 19.0 (IBM Corp., Armonk, NY, USA). Unpaired
data were determined by Student's t-test and comparisons of data
between multiple groups were analyzed via analysis of variance.
Kaplan-Meier was used to estimate the risk of relapse and
re-treatment during 120-day treatment. P<0.05 was considered to
represent statistically significant differences.
Results
Dexamethasone inhibits TGF-β1 induced
migration and invasion in NSCLC
It has been demonstrated that the expression of
TGF-β1 is associated with the prognosis of human cancer and TGF-β1
enhanced CYR61 expression, which leads to the migration and
invasion of tumor cells (26).
Although the role of TGF-β1 in NSCLC is not understood,
dexamethasone has been demonstrated to inhibit migration and
invasion in human colon cancer cells in a previous study (21). To investigate the suppressive effects
of dexamethasone in human NSCLC H1975 and H358 cells, tumor cell
viability and migration were evaluated via MTT and scratch-wound
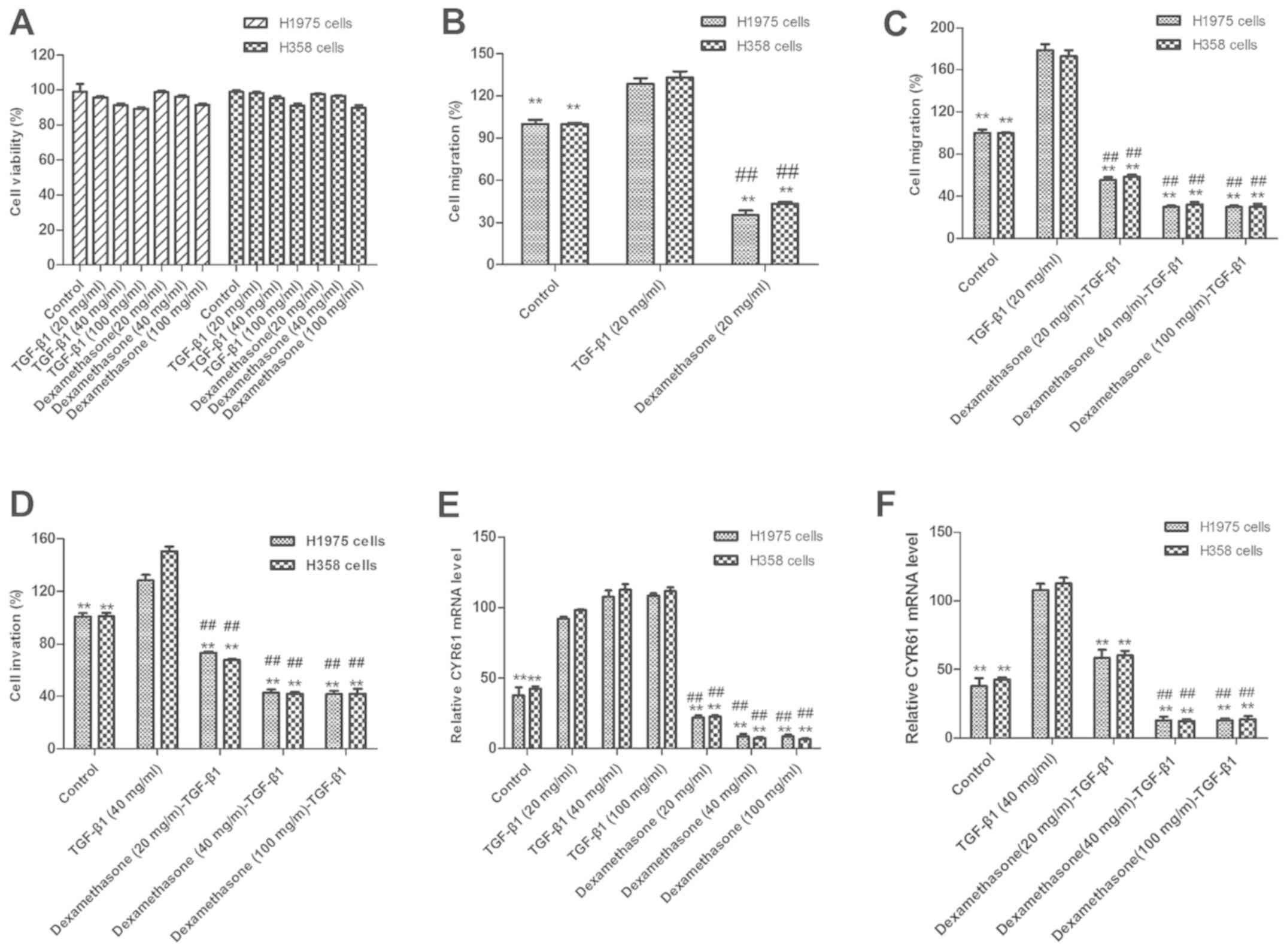
assays, respectively. The results in Fig. 1A indicate that there were changes in
the viability of cells treated with TGF-β1 and dexamethasone (20,
40 and 100 mg/ml) compared with non-treated control cells. As
presented in Fig. 1B, dexamethasone
treatment (20 mg/ml) significantly suppressed H1975 and H358 cell
migration following 12-h exposure, as compared with untreated and
TGF-β1-treated cells (P<0.01). The results revealed that 20
mg/ml of TGF-β1 and 20 mg/ml of dexamethasone exhibited the maximal
inhibitory effect on H1975 and H358 cells. Therefore, 20 mg/ml of
TGF-β1 in combination with 20 mg/ml of dexamethasone was selected
as the final dose.
Notably, migration data also demonstrated that TGF-β
promoted the migration and invasion of NSCLC H1975 and H358 cells.
As presented in Fig. 1C, 20 ng/ml
TGF-β1 significantly enhanced the migration of H1975 and H358
cells, as compared with control cells (P<0.01). By contrast,
dexamethasone (20 mg/ml) significantly inhibited the migration of
H1975 and H358 cells when administered with 20 ng/ml TGF-β1
(P<0.01).
In order to confirm the effects of dexamethasone on
the migration of H1975 cells, Transwell assays were used to further
assess the invasion of H1975 and H358 cells. The results in
Fig. 1D indicate that dexamethasone
significantly inhibited the tumor cell invasion induced by TGF-β1
(P<0.01). In conclusion, these results suggest that
dexamethasone not only affects tumor cell viability but also
inhibits TGF-β1-dependent tumor cell migration and invasion. To
investigate the influence of dexamethasone on tumor cell migration
through CYR61 modulation, CYR61 was analyzed. The results in
Fig. 1E and F indicate that CYR61
expression was significantly increased following treatment with
TGF-β1 and significantly decreased following dexamethasone
treatment (P<0.01).
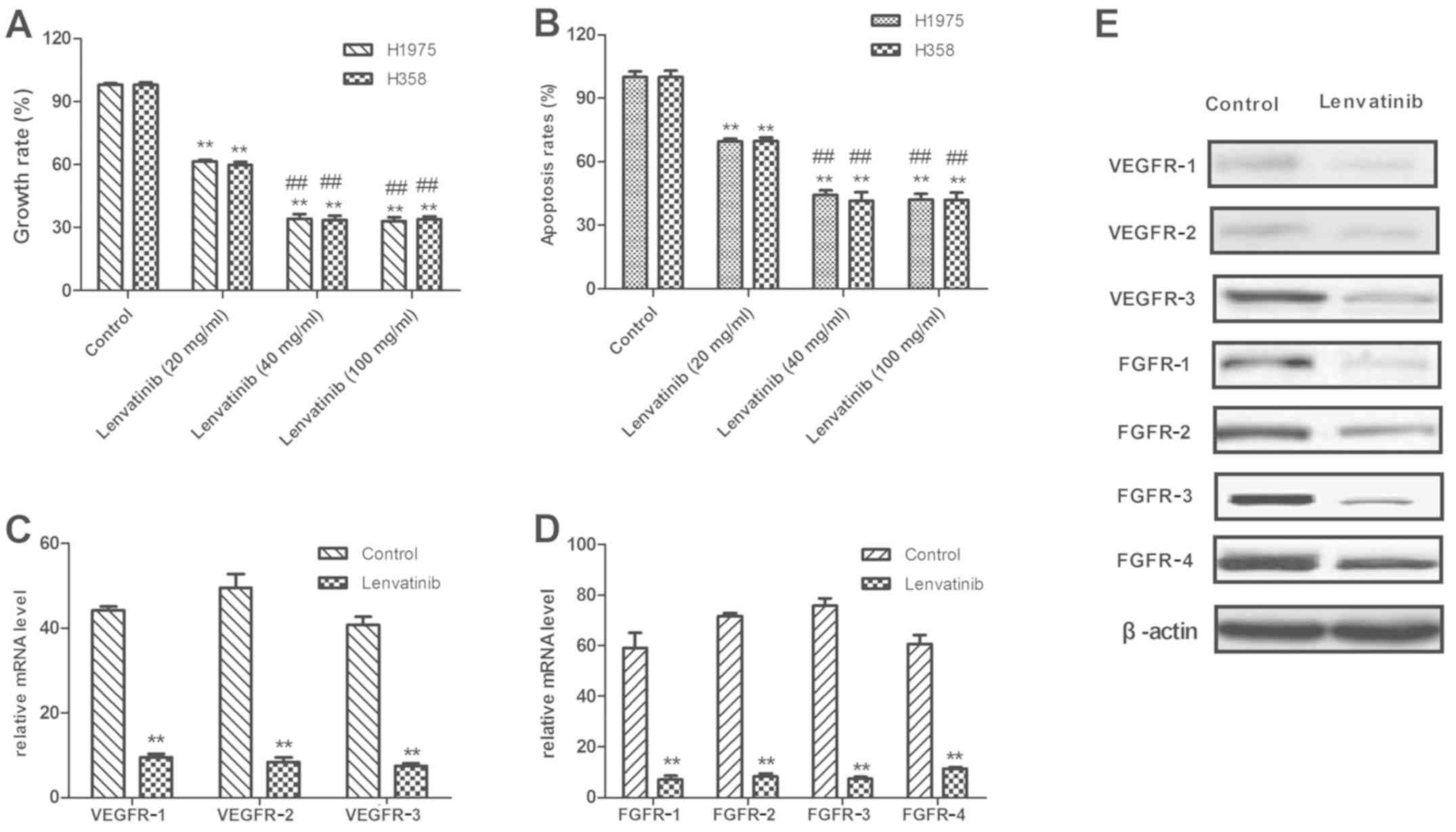
Lenvatinib suppresses the growth of
NSCLC by binding with VEGFR1-3
Lenvatinib was reported to be an adjuvant therapy
combined with carboplatin and paclitaxel in patients with NSCLC
(14). In order to analyze the
cell-killing effects of lenvatinib, the current study used an MTT
assay to assess the inhibitory effects on the viability of H1975
and H358 cells. In Fig. 2A, growth
of H1975 and H358 cells is demonstrated to be inhibited following
treatment with 40 mg/ml lenvatinib for 48 h. It was also
demonstrated that lenvatinib at a concentration of >40 mg/ml was
enough to inhibit tumor cell growth. For further analysis, the
apoptosis of two human NSCLC cell lines, H1975 and H358, was
examined by FACS following lenvatinib treatment FACS. As presented
in Fig. 2B, the apoptosis rate of
H1975 and H358 was significantly decreased following treatment with
lenvatinib at a concentration of 40 mg/ml compared with a
concentration of 10 mg/ml. It was also observed that a lenvatinib
dose ≥20 mg/ml was enough to significantly reduce the rate of
apoptosis in H1975 and H358 cells compared with the untreated
control.
Previous reports indicate that treatment with
lenvatinib exhibited an effective outcome in patients with cancer
(14). However, few studies
analyzing the use of lenvatinib for NSCLC have been completed
(14,27). In the current study, the expression
of VEGFR1-3 and FGFR1-4 protein in H1975 and H358 was analyzed,
compared with normal lung cells. As presented in Fig. 2C, the mRNA expression level of
VEGFR1-3 in control H1975 cells was significantly higher than
lenvatinib-treated cells (P<0.01). RT-qPCR demonstrated that
FGFR1-4 mRNA expression was significantly decreased in
lenvatinib-treated H1975 cells compared with the non-treated
control (Fig. 2D; P<0.01). The
results in Fig. 2E demonstrated that
VEGFR1-3 protein expression was decreased in lenvatinib-treated
H1975 cells compared with untreated cells. These results indicated
that the growth of H1975 and H358 cells was suppressed following
treatment with lenvatinib.
Dexamethasone inhibits the
epithelial-mesenchymal transition (EMT) process via the AKT/ERK
signaling pathways
In consideration of the inhibitive effects of
dexamethasone on TGF-β1-induced cell migration and invasion, the
present study hypothesized that the TGF-β1-induced EMT signal
pathway was inhibited by dexamethasone. Therefore, the underlying
mechanism of dexamethasone on the expression of important EMT
markers, including Vimentin, E-cadherin and Slug was assessed. The
results in Fig. 3A demonstrate that
the expression levels of Vimentin, E-cadherin and Slug were
significantly elevated when treated with TGF-β1 in H1975 cells,
whereas dexamethasone significantly decreased this elevated
expression (P<0.01), compared with a control. Furthermore, to
confirm that dexamethasone inhibited the EMT process via the
AKT/ERK signaling pathway, protein expression of EKR, AKT and Snail
induced by TGF-β was markedly promoted and eliminated this effect
following dexamethasone treatment (Fig.
3B).
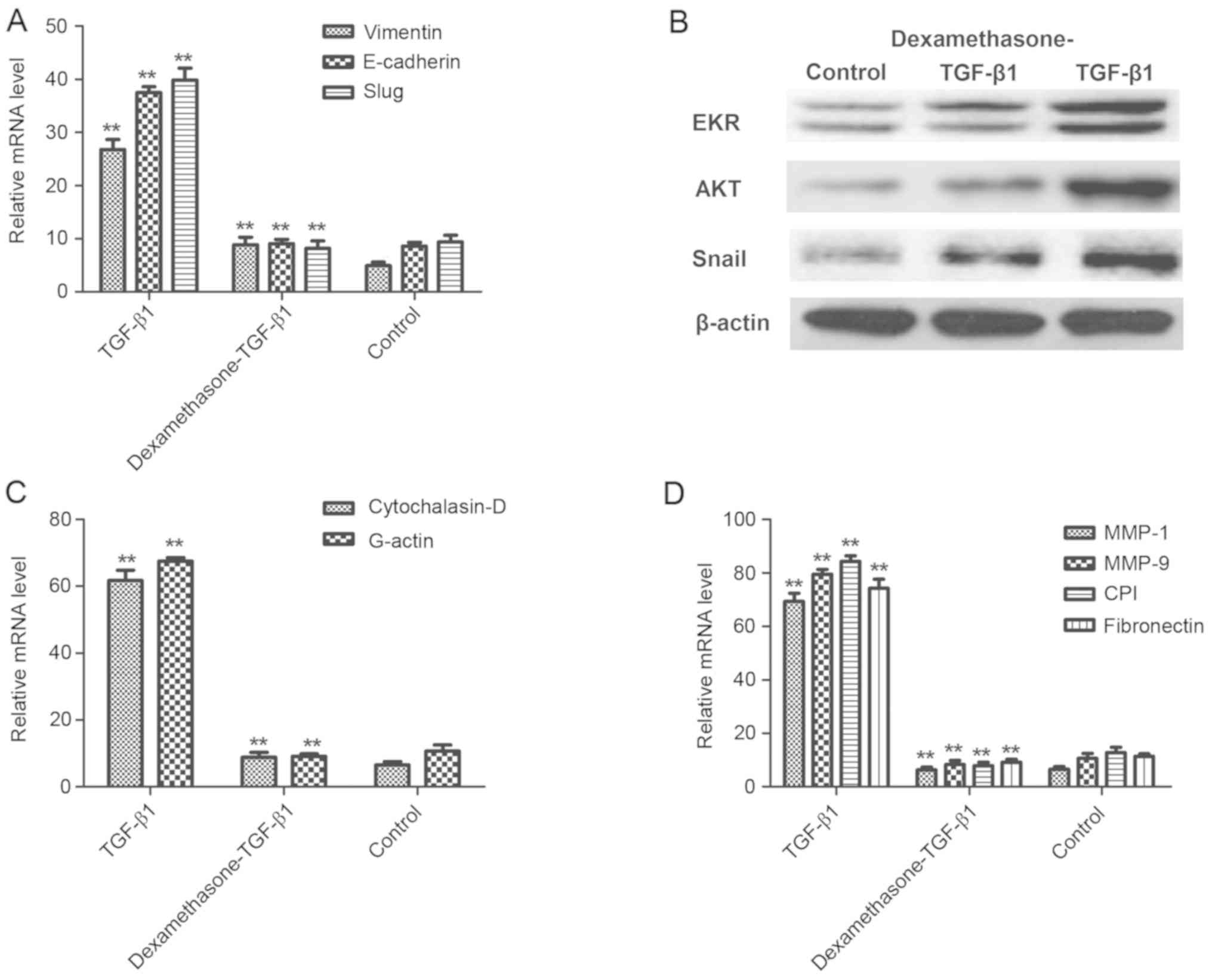 | Figure 3.Dexamethasone inhibited the EMT
process via the AKT/ERK signaling pathways. (A) TGF-β1 increased
the mRNA expression of Vimentin, E-cadherin and Slug, and
dexamethasone eliminated increasing expression in H1975 cells. (B)
TGF-β1 upregulated protein expression of EKR, AKT and Snail and
dexamethasone eliminated increasing expression in H1975 cells. (C)
TGF-β1 increased Cytochalasin-D and G-actin but dexamethasone
canceled-out this effect of regulation. (D) Migration-related
MMP-1, MMP-9, CPI and fibronectin expression were increased
following treatment with TGF-β1 and this was canceled-out by 40
mg/ml dexamethasone. **P<0.01 vs. control. Data are presented as
mean ± standard error of the mean. EMT, epithelial-mesenchymal
transition; AKT, protein kinase B; ERK, extracellular
signal-regulated kinases; TGF-β1, transforming growth factor-β1;
MMP, matrix metalloproteinase; CPI, collagen type I. |
A previous study indicated that degradation of the
extracellular matrix (ECM) by MMP proteins is associated with tumor
cell migration (28). The results
presented in Fig. 3C indicate that
Cytochalasin-D and G-actin were significantly upregulated in
TGF-β1-treated tumor cells, whereas dexamethasone canceled-out this
effect (P<0.01), compared with a control. However, the
expression levels of MMP-1, MMP-9, collagen type I and fibronectin,
which are important ECM proteins with a vital role in cell
migration, were enhanced following TGF-β1 treatment and were
significantly decreased after treatment with TGF-β1 and
dexamethasone in H1975 cells, compared with a control (P<0.01;
Fig. 3D). These results suggested
that dexamethasone suppressed the migration-promoting proteins and
promoted the migration-inhibiting proteins in TGF-β1-induced EMT
process, which may be beneficial in the treatment of cancer cell
migration and invasion in the EKR/AKT pathway.
Efficacy of synergistic treatment of
dexamethasone and lenvatinib in NSCLC-bearing mice
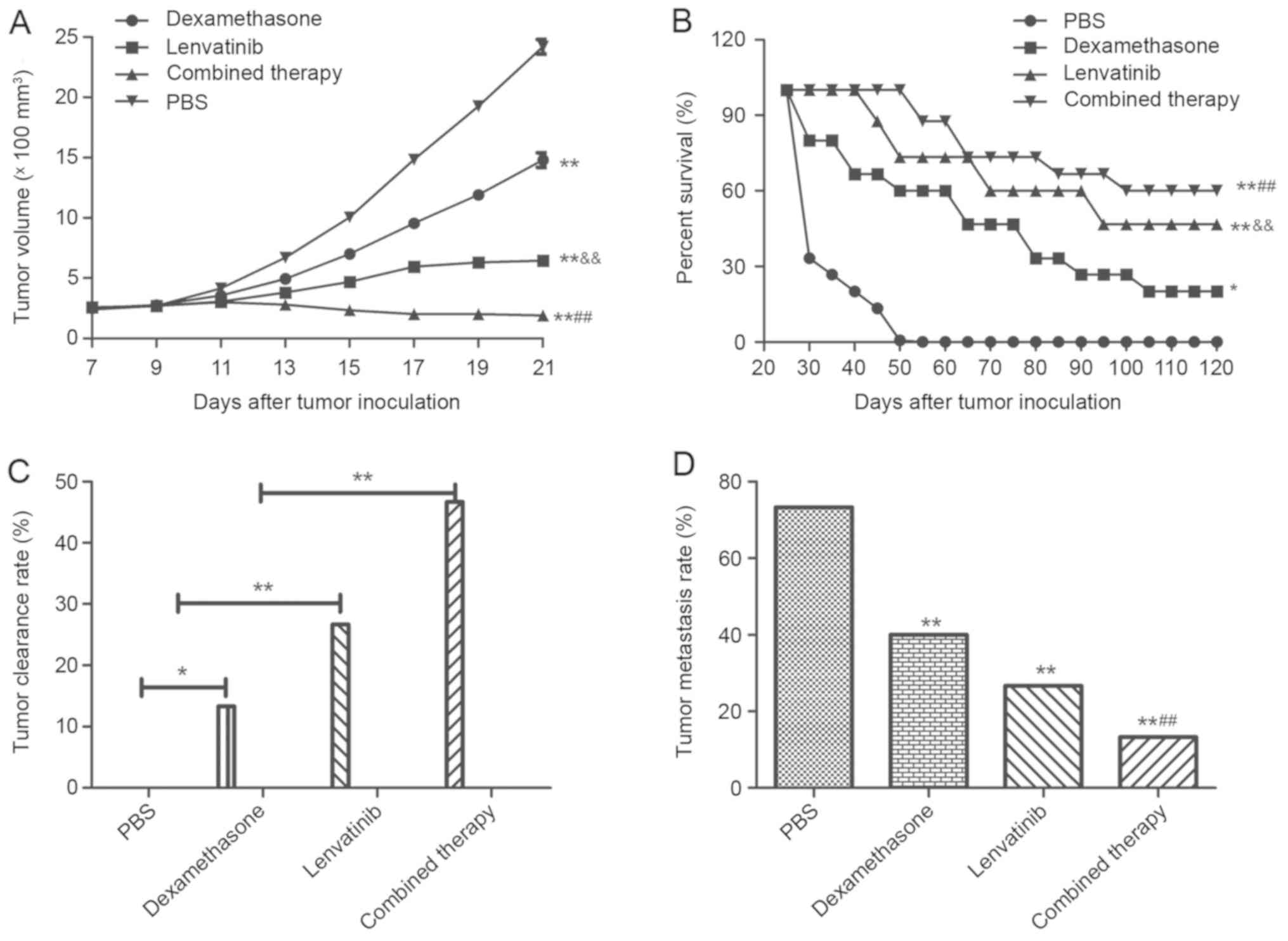
To investigate whether the synergistic treatment of
dexamethasone and lenvatinib are effective agents for NSCLC in
vivo, NSCLC-bearing mice were established for further analysis.
The anti-tumor efficacy of dexamethasone and lenvatinib was
assessed in the NSCLC mouse model. As presented in Fig. 4A, tumor size was significantly
inhibited in xenograph mice treated with dexamethasone and
lenvatinib compared with the mice treated with PBS or a single
agent (P<0.01). In addition, 120-day long-term survival analysis
revealed that the combined treatment of dexamethasone and
lenvatinib significantly increased the survival rate compared with
single dexamethasone or lenvatinib treatment (Fig. 4B). As expected, the results in
Fig. 4B demonstrated that
synergistic treatment (n=15 in each group) prolonged the survival
of NSCLC-bearing mice compared with single agent and control mice.
Fig. 4C also indicated that
synergistic treatment of dexamethasone and lenvatinib against NSCLC
was effective enough to partially protect the animals and
significantly increased the percentage of tumors eliminated in the
combined treatment group compared with the dexamethasone,
lenvatinib and PBS groups, which translated into long-term survival
and tumor-free living. Furthermore, Fig.
4D indicates that the inhibition of tumor metastasis in
NSCLC-bearing mice was significantly increased in the combined
therapy group compared with the dexamethasone, lenvatinib and PBS
groups (P<0.01). Results showed that combined therapy group
demonstrated significant difference compared to dexamethasone or
lenvatinib group (P<0.01). However, there was no significant
difference between the dexamethasone group and the lenvatinib
group.
Discussion
An increased occurrence of cancer has been
associated with industrial pollution and destruction of the
ecological environment in the last century (29). The increased rate of morbidity and
mortality from lung cancer has become a particular problem and an
increase in diagnosis has occurred in recent years (30). NSCLC is the type of primary lung
cancer and is difficult to detect in the early stages. Thus, the
majority of NSCLC patients are diagnosed with advanced stage lung
cancer (31). As conventional
radiation and chemotherapy exhibits little efficacy for NSCLC, more
frequent recurrence and metastasis has occurred. In addition, a
poor survival rate (<15%) of patients with NSCLC was determined
in over a 5-year observation (32).
Therefore, investigating novel agents to target
metastasis-promoting factors has become a focus for the treatment
of patients with NSCLC.
A previous study reported that CYR61 regulated
migration in the EMT process and induced apoptosis in various
cancer cells (33). In addition,
CYR61 mediated Src signaling in triple negative breast cancer cells
and was modulated by phosphoinositide 3-kinase/AKT signaling in
prostate cancer (34,35). TGF-β1 has been demonstrated to
upregulate CYR61 expression, which leads to migration and invasion
in colon cancer cells and osteosarcoma cells (21,36). In
the current study, CYR61 expression was demonstrated to be
superfluous in H1975 and H358 cells compared with normal lung
cells. The CYR61 signaling pathway was indispensable in the EMT
process. In addition, dexamethasone inhibited TGF-β1-induced CYR61
expression and regulated N-cadherin, fibronectin, vimentin and
E-cadherin expression, which was similar to a previous study
(21). Furthermore, dexamethasone
treatment significantly suppressed H1975 cells migration at dose of
20 µM following 12-h exposure compared with untreated cells in
vitro and inhibited tumor metastasis in xenograph mice in
vivo.
Evidence of targeting VEGF-mediated pathways in a
number of cancer tissues or tumors has demonstrated available
treatment options (15,37,38).
Lenvatinib is a multi targeted anti-cancer target-therapeutic drug
and studies have demonstrated that it is an efficient anticancer
agent in patients with advanced solid tumors (39). However, single-agent therapy has to
improve targeted treatment to overcome a low response rate and
metastasis for patients with cancer (40). Therefore, combined treatment may be
an ideal therapeutic regimen. In the current study, VEGFR1-3 and
FGFR1-4 expression was detected and cytotoxic effects on H1975 and
H358 cells were assessed. Notably, the migration of H1975 and H358
cells was observed to be suppressed following treatment with
lenvatinib in vitro and in vivo.
Preclinical observation with combination therapy
conferred relative advantages to single drug treatment and further
study is required to evaluate the overall efficacy (41). The current exploratory analysis of
tumor treatment and long-term survival indicated that treatment
with lenvatinib and dexamethasone once daily exhibited a clear
tumor regression and resulted in a survival rate of 80% in a
120-day period in xenograph mice. Furthermore, the median
metastasis-limit survival during 120-day observation in NSCLC mice
model reached 66.67%. The results of the current study indicated
that synergistic treatment of dexamethasone and lenvatinib against
NSCLC is effective enough to partially protect the animals and
eliminate the tumors in experimental mice, which translated into
long-term survival and tumor-free living.
In conclusion, two anticancer agents, dexamethasone
and lenvatinib, were administered once daily in the present study.
Notably, the data is suggestive of a NSCLC mice model, although
dexamethasone downregulated the expression of EKR and AKT, and
lenvatinib downregulated VEGFR and FGFR, which led to the reversal
of TGF-β1-induced cell migration. Dexamethasone and lenvatinib
inhibited the migration and invasion of NSCLC by regulating the
EKR/AKT and VEGF signal pathways. The current study suggests that
the beneficial effects of cellular targeted therapy may further
elucidate the mechanisms and this regimen is clinically applicable,
but requires further study.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
DZ designed and wrote this study. YZ analyzed data
and prepared the materials. ZC conducted some of the experiments.
YT and ZH conducted the data analysis and experimental design, and
proofed this manuscript.
Ethics approval and consent to
participate
The animal use protocol was approved by the
Committee on the Ethics of Animal Experiments of Tianjin Chest
Hospital.
Patient consent for publication
Not applicable.
Competing interests
All authors declare that they have no competing
interests.
References
|
1
|
Fenton-Ambrose L and Kazerooni EA:
Preventative care: Lung-cancer screens now worth the cost. Nature.
514:352014. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kong R, Feng J, Ma Y, Zhou B, Li S, Zhang
W, Jiang J, Zhang J, Qiao Z, Zhang T, et al: Silencing NACK by
siRNA inhibits tumorigenesis in non-small cell lung cancer via
targeting Notch1 signaling pathway. Oncol Rep. 35:2306–2314. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brody H: Lung cancer. Nature. 513:S12014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moro-Sibilot D, Smit E, de Castro Carpeño
J, Lesniewski-Kmak K, Aerts JG, Villatoro R, Kraaij K, Nacerddine
K, Dyachkova Y, Smith KT, et al: Non-small cell lung cancer
patients with brain metastases treated with first-line
platinum-doublet chemotherapy: Analysis from the European FRAME
study. Lung Cancer. 90:427–432. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Barnett SA, Downey RJ, Zheng J, Plourde G,
Shen R, Chaft J, Akhurst T, Park BJ and Rusch VW: Utility of
routine PET imaging to predict response and survival after
induction therapy for non-small cell lung cancer. Ann Thorac Surg.
101:1052–1059. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xie FJ, Lu HY, Zheng QQ, Qin J, Gao Y,
Zhang YP, Hu X and Mao WM: The clinical pathological
characteristics and prognosis of FGFR1 gene amplification in
non-small-cell lung cancer: A meta-analysis. Onco Targets Ther.
9:171–181. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lim SH, Sun JM, Lee SH, Ahn JS, Park K and
Ahn MJ: Pembrolizumab for the treatment of non-small cell lung
cancer. Expert Opin Biol Ther. 16:397–406. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Müller B, Bovet M, Yin Y, Stichel D, Malz
M, González-Vallinas M, Middleton A, Ehemann V, Schmitt J, Muley T,
et al: Concomitant expression of far upstream element (FUSE)
binding protein (FBP) interacting repressor (FIR) and its splice
variants induce migration and invasion of non-small cell lung
cancer (NSCLC) cells. J Pathol. 237:390–401. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao Q, Yue J, Zhang C, Gu X, Chen H and
Xu L: Inactivation of M2 AChR/NF-κB signaling axis reverses
epithelial-mesenchymal transition (EMT) and suppresses migration
and invasion in non-small cell lung cancer (NSCLC). Oncotarget.
6:29335–29346. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang H, Zhu X, Li N, Li D, Sha Z, Zheng X
and Wang H: miR-125a-3p targets MTA1 to suppress NSCLC cell
proliferation, migration, and invasion. Acta Biochim Biophys Sin
(Shanghai). 47:496–503. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Roth MT, Ivey JL, Esserman DA, Crisp G,
Kurz J and Weinberger M: Individualized medication assessment and
planning: Optimizing medication use in older adults in the primary
care setting. Pharmacotherapy. 33:787–797. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Solimando DA Jr and Waddell JA: Lenvatinib
and palbociclib. Hosp Pharm. 50:578–582. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Oikonomopoulos G, Aravind P and Sarker D:
Lenvatinib: A potential breakthrough in advanced hepatocellular
carcinoma? Future Oncol. 12:465–476. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nishio M, Horai T, Horiike A, Nokihara H,
Yamamoto N, Takahashi T, Murakami H, Yamamoto N, Koizumi F, Nishio
K, et al: Phase 1 study of lenvatinib combined with carboplatin and
paclitaxel in patients with non-small-cell lung cancer. Br J
Cancer. 109:538–544. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hutson TE: Targeted therapies for the
treatment of metastatic renal cell carcinoma: Clinical evidence.
Oncologist. 16 (Suppl 2):S14–S22. 2011. View Article : Google Scholar
|
|
16
|
Matsui J and Funahashi Y: Preclinical
biomarker research and patient stratification of molecular target
agents: The anti-angiogenic inhibitor Lenvatinib mesylate (E7080).
Nihon Yakurigaku Zasshi. 142:162–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Okamoto K, Kodama K, Takase K, Sugi NH,
Yamamoto Y, Iwata M and Tsuruoka A: Antitumor activities of the
targeted multi-tyrosine kinase inhibitor lenvatinib (E7080) against
RET gene fusion-driven tumor models. Cancer Lett. 340:97–103. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kuznar W: Lenvatinib extends survival in
metastatic renal-cell carcinoma. Am Health Drug Benefits.
8:182015.(In Japanese). PubMed/NCBI
|
|
19
|
Molina AM, Hutson TE, Larkin J, Gold AM,
Wood K, Carter D, Motzer R and Michaelson MD: A phase 1b clinical
trial of the multi-targeted tyrosine kinase inhibitor lenvatinib
(E7080) in combination with everolimus for treatment of metastatic
renal cell carcinoma (RCC). Cancer Chemother Pharmacol. 73:181–189.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Berdeja J, Jagannath S, Zonder J, Badros
A, Kaufman JL, Manges R, Gupta M, Tendolkar A, Lynch M, Bleickardt
E, et al: Pharmacokinetics and safety of elotuzumab combined with
lenalidomide and dexamethasone in patients with multiple myeloma
and various levels of renal impairment: Results of a phase Ib
study. Clin Lymphoma Myeloma Leuk. 16:129–138. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Han S, Bui NT, Ho MT, Kim YM, Cho M and
Shin DB: Dexamethasone inhibits TGF-β1-induced cell migration by
regulating the ERK and AKT pathways in human colon cancer cells via
CYR61. Cancer Res Treat. 48:1141–1153. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chijiiwa M, Mochizuki S, Kimura T, Abe H,
Tanaka Y, Fujii Y, Shimizu H, Enomoto H, Toyama Y and Okada Y: CCN1
(Cyr61) is overexpressed in human osteoarthritic cartilage and
inhibits ADAMTS-4 (Aggrecanase 1) activity. Arthritis Rheumatol.
67:1557–1567. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Grazioli S, Gil S, An D, Kajikawa O,
Farnand AW, Hanson JF, Birkland T, Chen P, Duffield J, Schnapp LM,
et al: CYR61 (CCN1) overexpression induces lung injury in mice. Am
J Physiol Lung Cell Mol Physiol. 308:L759–L765. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhuang T, Djemil T, Qi P, Magnelli A,
Stephans K, Videtic G and Xia P: Dose calculation differences
between Monte Carlo and pencil beam depend on the tumor locations
and volumes for lung stereotactic body radiation therapy. J Appl
Clin Med Phys. 14:40112013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang YT, Lan Q, Ponsonnet L, Blanquet M,
Christofori G, Zaric J and Rüegg C: The matricellular protein CYR61
interferes with normal pancreatic islets architecture and promotes
pancreatic neuroendocrine tumor progression. Oncotarget.
7:1663–1674. 2016.PubMed/NCBI
|
|
27
|
Nagashima S, Matsuo S, Takahashi M,
Umemoto Y, Hirano T, Enomoto K, Sakurai K and Amano S:
Effectiveness of lenvatinib for thyroid cancer with lung
metastases-report of a case. Gan To Kagaku Ryoho. 43:2121–2123.
2016.(In Japanese). PubMed/NCBI
|
|
28
|
Li Z, Xu X, Bai L, Chen W and Lin Y:
Epidermal growth factor receptor-mediated tissue transglutaminase
overexpression couples acquired tumor necrosis factor-related
apoptosis-inducing ligand resistance and migration through c-FLIP
and MMP-9 proteins in lung cancer cells. J Biol Chem.
286:21164–21172. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee YT, Liu CJ, Hu YW, Teng CJ, Tzeng CH,
Yeh CM, Chen TJ, Lin JK, Lin CC, Lan YT, et al: Incidence of second
primary malignancies following colorectal cancer: A distinct
pattern of occurrence between colon and rectal cancers and
association of co-morbidity with second primary malignancies in a
population-based cohort of 98,876 patients in Taiwan. Medicine
(Baltimore). 94:e10792015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Charvat H, Sasazuki S, Inoue M, Iwasaki M,
Sawada N, Shimazu T, Yamaji T and Tsugane S; JPHC Study Group, :
Prediction of the 10-year probability of gastric cancer occurrence
in the Japanese population: The JPHC study cohort II. Int J Cancer.
138:320–331. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim DS, Park KM, Won YS, Kim JY, Lee JK,
Kim JG, Oh ST, Jung SS and Kang WK: Occurrence and prognosis of
symptomatic venous thromboembolism in colorectal cancer surgery
patients. Vasc Specialist Int. 30:49–55. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gold M, Dunn LB, Phoenix B, Paul SM,
Hamolsky D, Levine JD and Miaskowski C: Co-occurrence of anxiety
and depressive symptoms following breast cancer surgery and its
impact on quality of life. Eur J Oncol Nurs. 20:97–105. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhu X, Song Y, Huo R, Zhang J, Sun S, He
Y, Gao H, Zhang M, Sun X, Zhai T, et al: Cyr61 participates in the
pathogenesis of rheumatoid arthritis by promoting proIL-1β
production by fibroblast-like synoviocytes through an AKT-dependent
NF-κB signaling pathway. Clin Immunol. 157:187–197. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sánchez-Bailón MP, Calcabrini A,
Mayoral-Varo V, Molinari A, Wagner KU, Losada JP, Ciordia S, Albar
JP and Martín-Pérez J: Cyr61 as mediator of Src signaling in triple
negative breast cancer cells. Oncotarget. 6:13520–13538. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee YJ, Lee DM and Lee SH: Production of
Cyr61 protein is modulated by extracellular acidification and
PI3K/Akt signaling in prostate carcinoma PC-3 cells. Food Chem
Toxicol. 58:169–176. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen J, Song Y, Yang J, Gong L, Zhao P,
Zhang Y and Su H: The up-regulation of cysteine-rich protein 61
induced by transforming growth factor beta enhances osteosarcoma
cell migration. Mol Cell Biochem. 384:269–277. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Villanueva MT: Targeted therapies:
Congratulations, you are still in the running towards becoming
ovarian-next-top treatment. Nat Rev Clin Oncol. 8:5702011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ngeow J, Tan IB and Choo SP: Targeted
therapies in the treatment of gastric cancer. Asia Pac J Clin
Oncol. 7:224–235. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Boss DS, Glen H, Beijnen JH, Keesen M,
Morrison R, Tait B, Copalu W, Mazur A, Wanders J, O'Brien JP, et
al: A phase I study of E7080, a multitargeted tyrosine kinase
inhibitor, in patients with advanced solid tumours. Br J Cancer.
106:1598–1604. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tsuruoka A, Matsui J, Suzuki T, Koyama N,
Watanabe T and Funahashi Y: Preclinical and clinical researches of
lenvatinib mesylate (Lenvima capsule), a novel antitumor agent
approved for thyroid cancer treatment. Nihon Yakurigaku Zasshi.
146:283–290. 2015.(In Japanese). View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nakagawa T, Matsushima T, Kawano S,
Nakazawa Y, Kato Y, Adachi Y, Abe T, Semba T, Yokoi A, Matsui J, et
al: Lenvatinib in combination with golvatinib overcomes hepatocyte
growth factor pathway-induced resistance to vascular endothelial
growth factor receptor inhibitor. Cancer Sci. 105:723–730. 2014.
View Article : Google Scholar : PubMed/NCBI
|