Introduction
Tuberous sclerosis complex (TSC) is a relatively
rare autosomal dominant disorder, which can involve multiple
organs, including brain, kidney, lung, skin and heart (1). The typical manifestations of TSC in the
kidneys include angiomyolipomas (AMLs), multiple renal cysts and
renal carcinomas (2). Renal AML
(RAML) is present in ~70-90% of patients with TSC, and it is the
leading cause of mortality in adult patients with the disorder
(3). The recommended first-line
therapy for TSC-RAML is the use of mTOR inhibitors. However,
~30-40% patients using mTOR inhibitors do not achieve clinical
remission, which is defined as a RAML volume reduction ≥50% and
severe adverse reactions may also occur in some patients (4,5). Despite
these issues, there are currently no other treatment options for
RAML (5). The critical role of
autophagy in the formation and development of tumors has become
increasingly recognized (6).
Autophagy influences the rearrangement of the plasma membrane,
leading to the sequestration of proteins and organelles intended
for delivery into lysosomes for degradation (7). Autophagy is a highly conserved
catabolic process of cellular proteins and organelles in which they
are sequestered into lysosomes for degradation to maintain cellular
homeostasis (8). There are three
types of autophagy, including microautophagy, macroautophagy and
chaperone-mediated autophagy (7), in
which macroautophagy is among the most researched. During
macroautophagy, the membrane expands to form phagophores, which
expand and bend continuously. The membrane then wraps around the
cargo to form complete autophagosomes, which is then delivered into
the lysosomes for degradation where the materials are then reused
(7). Human microtubule-associated
protein 1 light chain 3 (LC3) is an important protein for
autophagosome formation (7). LC3 is
normally spliced further into LC3-I and LC3-II, which is mainly
localized in the cytoplasm and on the autophagosomal double
membrane, respectively (9). The
levels of LC3-I and II can therefore be used to reflect the degree
of autophagy. Miyahara et al (9) previously demonstrated in the cerebral
cortical tubers of patients with TSC that the levels of LC3-I and
LC3-II were lower compared with those in normal cerebral tissues.
Autophagy may promote or inhibit tumorigenesis, depending on the
cellular context (6). Autophagic
dysfunction can lead to a variety of human pathologies, including
cancer, metabolic diseases and degenerative conditions (10). In Myc-induced lymphomas, chloroquine
may inhibit autophagy and the fusion of autophagosomes and
lysosomes, resulting in the suppression of lymphoma development
(11). Haploinsufficiency of the
autophagy gene, Beclin 1, promotes tumor formation in the
TSC2+/- mouse model (6) and allelic loss of Beclin 1 may be
associated with breast, ovarian and prostate cancers (12).
mTOR also participates in the regulation of
autophagy by inhibiting the formation of the autophagosome
(13). Ras homolog-enriched in brain
(Rheb) is a principal downstream target of the TSC1/TSC2 complex
and an upstream regulator of mTOR. When TSC1 or TSC2 mutates, Rheb
can be abnormally activated and promote mTOR activation to
downregulate autophagy (13). PTEN
is an upstream regulator of Akt that regulates the TSC1/TSC2
complex (14,15), but has also been revealed to
potentially regulate autophagy via a different signaling pathway
independent of TSC1/TSC2(15). The
current study indicated that the PTEN inhibitor,
hydroxyl(oxo)vanadium 3-hydroxypiridine-2-carboxylic acid
(VO-OHpic), could suppress the proliferation of
TSC2-/- murine embryonic
fibroblasts (MEFs) by excessively inhibiting autophagy via the
PTEN/proline-rich Akt substrate 40 kDa (PRAS40) pathway.
Materials and methods
Cell culture
The TSC2-/- and
TSC2+/+ MEF cell lines, as
aforementioned (16), were donated
by Professor Hongbing Zhang, Chinese Academy of Medical Sciences
and Peking Union Medical College (Beijing, China). The cells were
cultured in DMEM supplemented with 10% FBS, 100 U/ml penicillin and
100 mg/ml streptomycin at 37˚C in a humidified 5% CO2
atmosphere. The TSC2-/- MEFs were
used as two groups: i) With VO-OHpic treatment at 1 µM as
previously described (17); or ii)
without VO-OHpic treatment. All cultured
TSC2-/- and
TSC2+/+ MEFs were harvested at 24,
48 and 72 h, respectively, for further analysis.
Cell proliferation analysis
Cell proliferation assays were conducted using a
Cell Counting kit-8 (CCK-8; Beijing Solarbio Science &
Technology Co., Ltd.; cat. no CA1210) according to the
manufacturer's protocol. The harvested cells (5,000 cells/well)
were seeded in 96-well plates at 100 µl per well and cultured in
growth medium at 37˚C and 5% CO2. A total of 10 µl of
CCK-8 were added to each well at 24, 48 and 72 h. Subsequently, the
cells were incubated at 37˚C for 3 h and the optical density was
measured at 450 nm.
Western blot analysis
The cells were stored at -80˚C, then proteins were
subsequently extracted with protein lysis buffer (Beijing Solarbio
Science & Technology Co., Ltd.; cat. no. R0020). The protein
concentration was standardized using a BCA protein assay kit
(Beijing Solarbio Science & Technology Co. Ltd.; cat. no.
PC0020). A total of 30 µg protein extracts from the various groups
(with or without VO-OHpic treatment, at 24, 48 or 72 h,
respectively) were loaded into a 15% SDS-PAGE gel and transferred
onto PVDF membranes. The membranes were blocked with 5% skimmed
milk in Tris-buffered saline with Tween 20 (0.01 M Tris-HCl, pH
7.6; 0.15 M NaCl; 0.05% Tween 20) for 2 h at room temperature. The
membranes were incubated with rabbit anti-LC3 antibodies Abs
(Sigma-Aldrich; Merck KGaA; cat. no. L8918; 1:2,000), rabbit
anti-phosphorylated (p)-PTEN Abs (Abcam; cat. no. ab31392;
1:1,000), rabbit anti-PTEN Abs (Abcam; cat. no. ab32199; 1:1,000),
rabbit anti-p-PRAS40 Abs (Abcam; cat. no. ab134084; 1:1,000) or
rabbit anti-PRAS40 Ab (Abcam; cat. no. ab151719; 1:1,000) overnight
at 4˚C. GAPDH antibodies were used as an internal control (Beijing
Zhongshan Jinqiao Biotechnology Co., Ltd.; cat. no. TA-08;
1:1,000). The bound primary Ab was detected with horseradish
peroxidase-linked goat anti-rabbit immunoglobulin G (IgG; Beijing
Zhongshan Jinqiao Biotechnology Co., Ltd.; cat. no. ZB-2301;
1:10,000). The secondary incubation was conducted at room
temperature for 1 h. Subsequently, the antigen-antibody complexes
were detected using ECL (Tanon Science & Technology Co. Ltd.).
Gray values were calculated using Image J 1.50i software (National
Institutes of Health).
Statistical analysis
All of the data were analyzed using SPSS version
19.0.0 (IBM Corp.) and GraphPad Prism version 6.0 (GraphPad
Software, Inc.). The results are presented as the mean ± SEM.
Differences between groups were analyzed using two-way ANOVAs
followed by post-hoc Tukey's tests. P<0.05 was considered to
indicate a statistically significant difference. All experiments
were repeated at least three times.
Results
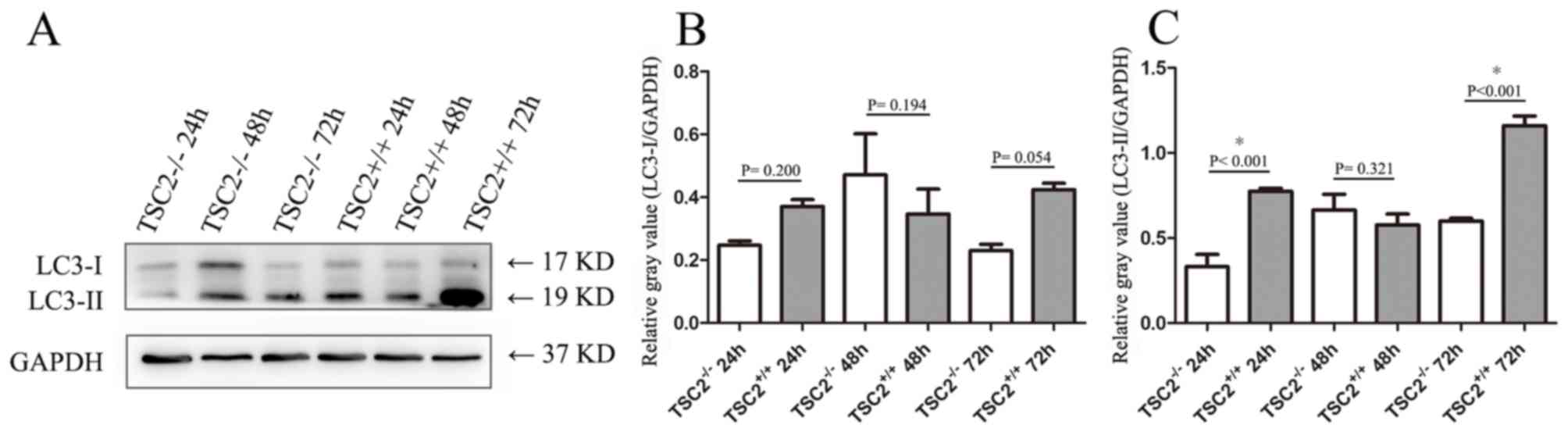
To confirm the autophagic conditions in
TSC2-/- MEFs, the expression
levels of LC3 were analyzed.
TSC2-/- and
TSC2+/+ MEFs were harvested at 24,
48 and 72 h, and western blot analysis was performed (Fig. 1). Overall, LC3 expression in
TSC2-/- MEFs was reduced compared
with that in TSC2+/+ MEFs at 24
and 72 h. Two-way ANOVA analysis indicated that neither
TSC2+/+/TSC2-/-
nor the time point of collection, had a significant effect on LC3-I
expression levels (P=0.245 and P=0.291, respectively; data not
shown). Considering the effect of
TSC2+/+/TSC2-/-,
no statistically significant differences were found (All P>0.05;
Fig. 1B). By contrast, both
TSC2+/+/TSC2-/-
and the collection time had a significant effect on LC3-II
expression (both P<0.001). When considering the effect of
TSC2+/+/TSC2-/-,
except for the cells at 48 h, LC3-II was expressed at a
significantly lower level in
TSC2-/- MEFs (24 h, P<0.001; 72
h, P<0.001; Fig. 1C).
Subsequently, a PTEN inhibitor, VO-OHpic, was added
to TSC2-/- MEFs and the effect on
cell proliferation and growth was observed. At various time points,
the cells that underwent VO-OHpic treatment demonstrated a reduced
volume and a decrease in their density (Fig. 2). The MEFs displayed larger cellular
bodies and nuclei, with more obvious processes, and the cells grew
at a greater density when VO-OHpic was not applied. VO-OHpic
treatment led to atrophy of the cell bodies and nuclei of MEFs and
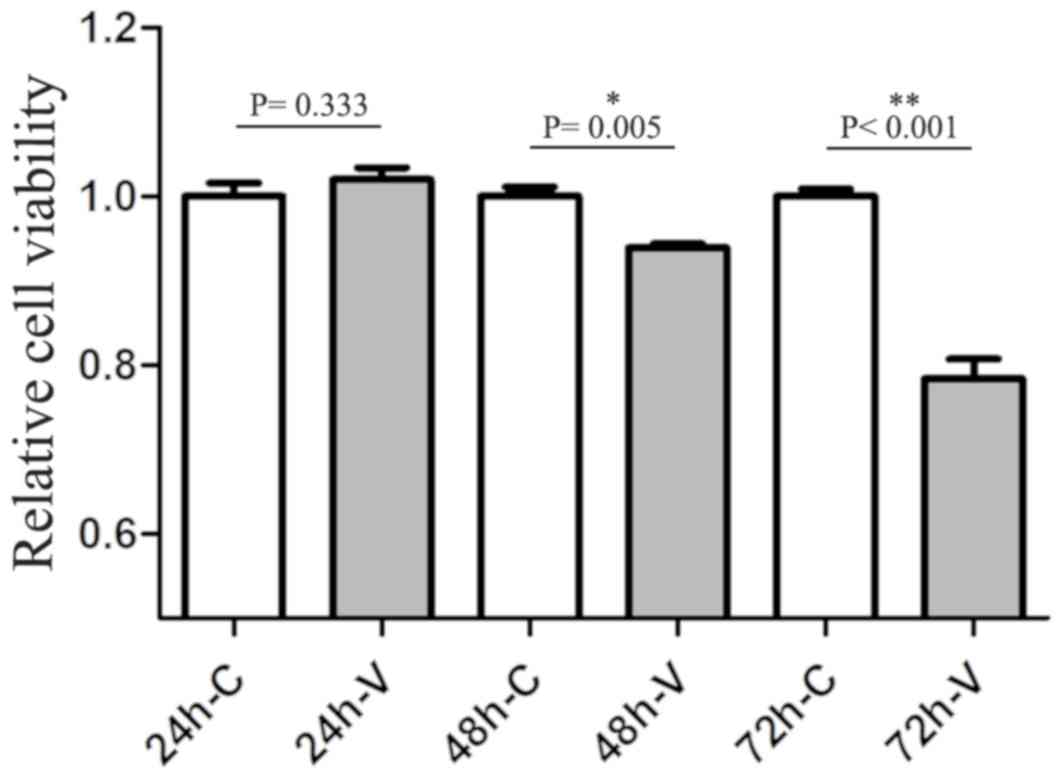
a reduction in the size and number of processes. CCK-8 assays were
also performed for quantitative analysis. A two-way ANOVA revealed
that both the intervention and the time points of collection had a
significant effect on the extent of cell viability (both,
P<0.001). When considering the effect of the intervention,
VO-OHpic led to a reduction in cell viability (with vs. without
VO-OHpic treatment: 24 h, P=0.333; 48 h, P=0.005; 72 h, P<0.001;
Fig. 3).
To further analyze the effects of the PTEN inhibitor
on autophagy of TSC2-/- MEFs, the
LC3 levels of cells treated with or without VO-Ohpic, were examined
(Fig. 4). Two-way ANOVA analysis
showed that the intervention had a significant effect on LC3-I
expression levels (overall, P<0.001; intervention vs. control:
24 h, P<0.001; 48 h, P=0.015; and 72 h, P=0.001); however, this
did not change with time (P=0.800). Similar results were indicated
for LC3-II expression levels (overall intervention, P<0.001;
overall time, P=0.644; intervention vs. control: 24 h, P=0.020; 48
h, P<0.001; and 72 h, P=0.007).
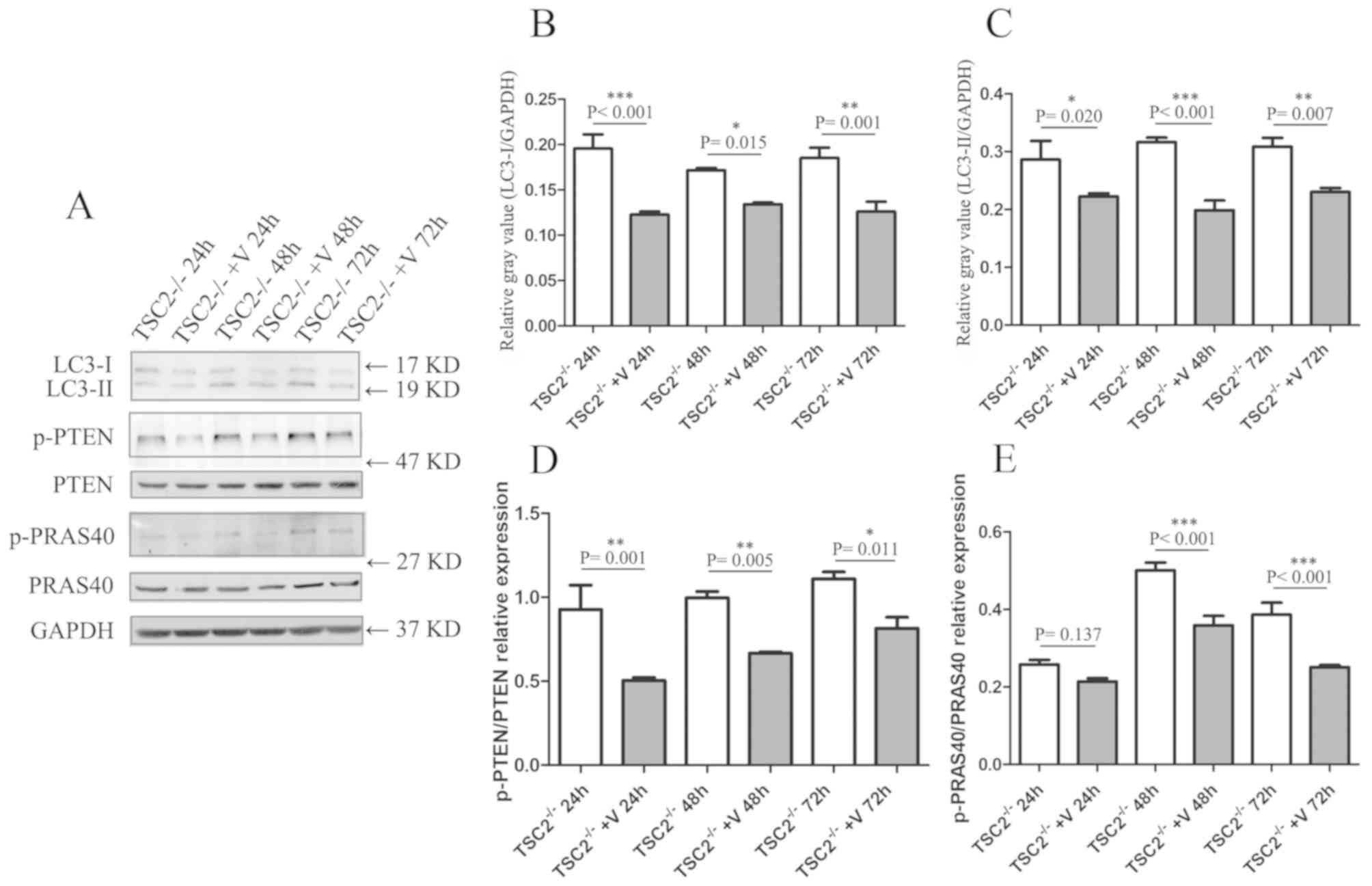 | Figure 4The autophagy levels and related
molecular pathways assessed in
TSC2-/- MEFs that received
VO-OHpic treatment. (A) Western blot assays of LC3, p-PTEN and
p-PRAS40 in TSC2-/- MEFs, with or
without VO-OHpic treatment. Quantitative analysis of (B) LC3-I and
(C) LC3-II expression levels in
TSC2-/- MEFs, with or without
VO-OHpic treatment. Quantitative analysis of (D) p-PTEN/PTEN and
(E) p-PRAS40/PRAS40 relative expression levels in
TSC2-/- MEFs, with or without
VO-OHpic treatment. *P<0.05, **P<0.01
and ***P<0.001. VO-OHpic, hydroxyl(oxo)vanadium
3-hydroxypiridine-2-carboxylic acid; +V, VO-OHpic treatment; MEF,
murine embryonic fibroblasts; p, phosphorylated; TSC, tuberous
sclerosis complex; LC3, human microtubule-associated protein 1
light chain 3; PRAS40, proline-rich Akt substrate 40 kDa. |
The potential pathways which allowed for this change
were subsequently investigated. p-PTEN and PTEN levels were
examined in TSC2-/- MEFs under
various conditions (with or without VO-OHpic treatment at 24, 48 or
72 h). The relative expression ratios of p-PTEN/PTEN were then
calculated. Two-way ANOVA analysis revealed that both interventions
and time points had a significant effect on the p-PTEN/PTEN
expression ratio (interventions, P<0.001; time points, P=0.013).
When considering the effect of the PTEN inhibitor intervention, the
relative expression ratio of p-PTEN/PTEN declined in
TSC2-/- MEFs which were treated
with VO-OHpic (Fig. 4), reflecting
the decrease in the activity of the cells (intervention vs.
control: 24 h, P=0.001; 48 h, P=0.005; and 72 h, P=0.011; Fig. 4). The activity of PRAS40, which is a
downstream regulator of PTEN, was also affected by treatment with
VO-OHpic. Both interventions and time point measured had a
significant effect on p-PRAS40/PRAS40 expression ratio, as
indicated by two-way ANOVA analysis (both, P<0.001). When
considering the effect of the intervention, the relative expression
ratio of p-PRAS40/PRAS40 deceased in
TSC2-/- MEFs treated with VO-OHpic
(intervention vs. control: 24 h, P=0.137; 48 and 72 h, P<0.001;
Fig. 4).
Discussion
TSC-associated RAML is a life-threatening disease in
adult patients (3). Recently, mTOR
inhibitors such as everolimus are the preferred treatment option
for TSC-RAML (4). However, in
addition to severe adverse reactions associated with these
treatment options, ≤40% patients don't respond effectively to mTOR
inhibitors (4,5). Therefore, finding an alternative
therapeutic strategy for TSC-RAML is of urgent importance. The
present study revealed that PTEN inhibitors can suppress
TSC2-/- MEFs proliferation by
inhibition of autophagy through the PTEN/PRAS40/mTOR pathway,
bypassing the PTEN/Akt/Rheb/mTOR pathway. These observations
provide a research basis for further feasibility study of PTEN
inhibitor in TSC treatment.
mTORC1 is a complex, which contains mTOR, and the
Rheb-mTORC1 pathway activates the translation of mRNA (18). The excessive activation of mTORC1 has
been revealed to participate in a number of different diseases,
including lung cancer, diabetes, neurodegeneration and age-related
diseases (19). Overactive mTORC1
can perturb ribosome synthesis and reduce the accuracy of
translation, therefore leading to the production and accumulation
of abnormal proteins (19). Hamartin
and tuberin negatively regulate mTORC1 and GAP negatively regulates
Rheb (18). In a study on patients
with TSC and in TSC animal models, the overactive mTORC1 pathway
was revealed to be inhibited using rapamycin (20,21).
Rapamycin and its homologs are used in the clinical treatment for
TSC associated RAML, subependymal giant cell astrocytoma and
lymphangioleiomyomatosis (4,6,22-24).
However, rapamycin is not a cytotoxic agent, but a cytostatic
agent; thus, cannot be used to eliminate RAML. The tumors will
recover following the withdrawal of treatment; therefore,
alternative medical strategies are required (4,5).
In a previous study investigating TSC and autophagy,
Parkhitko et al (6) reported
that chloroquine, which may function by preventing the fusion of
autophagosomes and lysosomes, could prevent the proliferation of
TSC2-/- MEFs. It was also
demonstrated that in TSC2-/-
xenograft tumors or renal tumors from
TSC2+/- mice, rapamycin combined
with chloroquine presented a stronger inhibitory effect than either
single drug. Beclin 1 knockdown has also been demonstrated to
inhibit the development of the tumors (6). It is hypothesized that there is a
balance between the inhibition and activation of autophagy in TSC
tumors (6). The inhibition of
autophagy leads to the accumulation of P62/SQSTM1 in
TSC2-/- MEFs, which initiates the
development of TSC tumors (6).
Autophagy cannot be induced adequately with numerous metabolic
defects existing in TSC2-/- MEFs;
therefore, further suppression of autophagy may exhibit an
inhibitory effect on tumor development. Therefore, the development
of TSC tumors may also depend on autophagy.
PTEN is a highly conserved dual-function protein
tyrosine phosphatase, which can dephosphorylate both inositol
lipids and proteins. PTEN can dephosphorylate the lipid second
messengers phosphatidylinositol 3,4,5-trisphosphate and
phosphatidylinositol 3,4-bisphosphate to produce
phosphatidylinositol 4,5-bisphosphate and phosphatidylinositol
4-phosphate, respectively, leading to the inhibition of Akt
activation (14). VO-OHpic is a
specific and reversible small molecule inhibitor of PTEN (14). PRAS40 is a component of the mTORC1
complex and is also a downstream target of Akt (25). However, p70 ribosome S6 kinase and
4E-binding protein 1, substrates of mTORC1, can combine with
mTORC1, thereby competing with PRAS40(26). The inhibition of PRAS40 increases the
frequency of interactions between mTORC1 and its substrates, and
suppresses autophagy by upregulating mTOR-p70S6K, resulting in
further mTORC1 activation and autophagy inhibition (26).
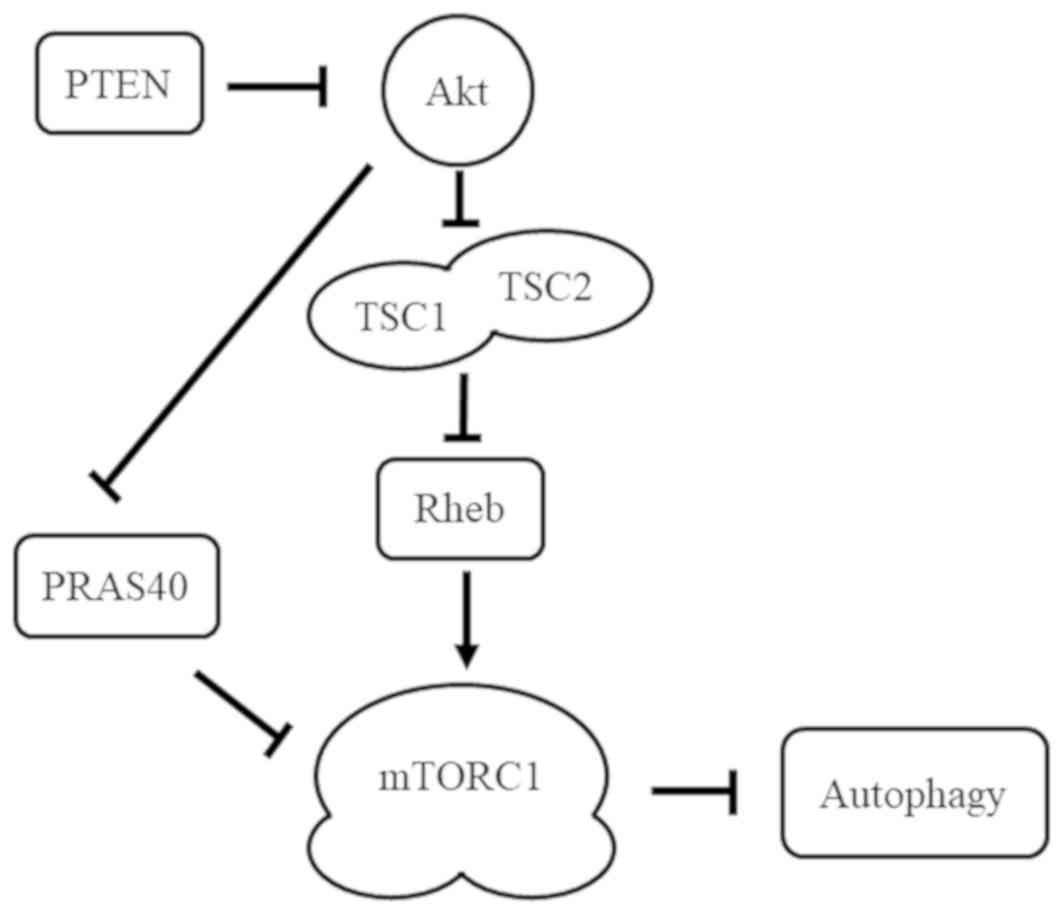
In a study by Nardella et al (15) on prostate cancer, PTEN deficiencies
promoted prostate tumor formation in Rheb overexpressing mice. The
aforementioned study indicated that PTEN not only regulates
autophagy through the Akt/Rheb/mTOR pathway, but also through the
Akt/PRAS40 pathway (Fig. 5). In the
present study, the effect of PTEN inhibitors on autophagy in
TSC2-/- MEFs was investigated. It
was revealed that the PTEN inhibitor, VO-OHpic, could suppress
autophagy, cell proliferation and cell growth in
TSC2-/- MEFs. The expression of
p-PTEN and p-PRAS40 also declined, revealing that VO-OHpic may
regulate autophagy via the PTEN-PRAS40 pathway. The results are
based on in vitro experiments, and further in vivo
studies are required to validate these results. The further
downstream proteins also require investigation.
In conclusion, the PTEN inhibitor, VO-OHpic, may
suppress the proliferation of
TSC2-/- MEFs, potentially through
the excessive inhibition of autophagy via the PTEN/PRAS40 pathway,
and may be a potential therapeutic target for the treatment of
TSC-RAML.
Acknowledgements
We thank Professor Hongbing Zhang (Institute of
Basic Medical Sciences and School of Basic Medicine, Chinese
Academy of Medical Sciences and Peking Union Medical College,
Beijing, China) for the donation of
TSC2-/- and
TSC2+/+ MEF cell lines.
Funding
The present study was supported by National Natural
Science Foundation of China (grant nos. 81670611 and 81800590).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author's contributions
YZ and HL conceived and designed the experiments.
WW, XW and HG performed the cellular experiments. YC provided and
performed the cell culture. WW produced the manuscript. WW and YZ
conducted data analysis. All the authors have read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sasongko TH, Ismail NF and Zabidi-Hussin
Z: Rapamycin and rapalogs for tuberous sclerosis complex. Cochrane
Database Syst Rev. 7(CD011272)2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Lam HC, Siroky BJ and Henske EP: Renal
disease in tuberous sclerosis complex: Pathogenesis and therapy.
Nat Rev Nephrol. 14:704–716. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Vekeman F, Magestro M, Karner P, Duh MS,
Nichols T, van Waalwijk van Doorn-Khosrovani SB and Zonnenberg BA:
Kidney involvement in tuberous sclerosis complex: The impact on
healthcare resource use and costs. J Med Econ. 18:1060–1070.
2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Bissler JJ, Kingswood JC, Radzikowska E,
Zonnenberg BA, Frost M, Belousova E, Sauter M, Nonomura N,
Brakemeier S, de Vries PJ, et al: Everolimus for renal
angiomyolipoma in patients with tuberous sclerosis complex or
sporadic lymphangioleiomyomatosis: Extension of a randomized
controlled trial. Nephrol Dial Transplant. 31:111–119.
2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Cai Y, Guo H, Wang W, Li H, Sun H, Shi B
and Zhang Y: Assessing the outcomes of everolimus on renal
angiomyolipoma associated with tuberous sclerosis complex in China:
A two years trial. Orphanet J Rare Dis. 13(43)2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Parkhitko A, Myachina F, Morrison TA,
Hindi KM, Auricchio N, Karbowniczek M, Wu JJ, Finkel T, Kwiatkowski
DJ, Yu JJ and Henske EP: Tumorigenesis in tuberous sclerosis
complex is autophagy and p62/sequestosome 1 (SQSTM1)-dependent.
Proc Natl Acad Sci USA. 108:12455–12460. 2011.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Parzych KR and Klionsky DJ: An overview of
autophagy: Morphology, mechanism, and regulation. Antioxid Redox
Signal. 20:460–473. 2014.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Wang CW and Klionsky DJ: The molecular
mechanism of autophagy. Mol Med. 9:65–76. 2003.PubMed/NCBI
|
|
9
|
Miyahara H, Natsumeda M, Shiga A, Aoki H,
Toyoshima Y, Zheng Y, Takeuchi R, Murakami H, Masuda H, Kameyama S,
et al: Suppressed expression of autophagosomal protein LC3 in
cortical tubers of tuberous sclerosis complex. Brain Pathol.
23:254–262. 2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Wirawan E, Vanden Berghe T, Lippens S,
Agostinis P and Vandenabeele P: Autophagy: For better or for worse.
Cell Res. 22:43–61. 2012.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Amaravadi RK, Yu D, Lum JJ, Bui T,
Christophorou MA, Evan GI, Thomas-Tikhonenko A and Thompson CB:
Autophagy inhibition enhances therapy-induced apoptosis in a
Myc-induced model of lymphoma. J Clin Invest. 117:326–336.
2007.PubMed/NCBI View
Article : Google Scholar
|
|
12
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kim YC and Guan KL: mTOR: A pharmacologic
target for autophagy regulation. J Clin Invest. 125:25–32.
2015.PubMed/NCBI View
Article : Google Scholar
|
|
14
|
Pulido R: PTEN inhibition in human disease
therapy. Molecules. 23(E285)2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Nardella C, Chen Z, Salmena L, Carracedo
A, Alimonti A, Egia A, Carver B, Gerald W, Cordon-Cardo C and
Pandolfi PP: Aberrant Rheb-mediated mTORC1 activation and Pten
haploinsufficiency are cooperative oncogenic events. Genes Dev.
22:2172–2177. 2008.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zhang H, Bajraszewski N, Wu E, Wang H,
Moseman AP, Dabora SL, Griffin JD and Kwiatkowski DJ: PDGFRs are
critical for PI3K/Akt activation and negatively regulated by mTOR.
J Clin Invest. 117:730–738. 2007.PubMed/NCBI View
Article : Google Scholar
|
|
17
|
Spinelli L, Lindsay YE and Leslie NR: PTEN
inhibitors: An evaluation of current compounds. Adv Biol Regul.
57:102–111. 2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Saran U, Foti M and Dufour JF: Cellular
and molecular effects of the mTOR inhibitor everolimus. Clin Sci
(Lond). 129:895–914. 2015.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Yun YS, Kim KH, Tschida B, Sachs Z,
Noble-Orcutt KE, Moriarity BS, Ai T, Ding R, Williams J, Chen L, et
al: mTORC1 coordinates protein synthesis and immunoproteasome
formation via PRAS40 to prevent accumulation of protein stress. Mol
Cell. 61:625–639. 2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Kenerson HL, Aicher LD, True LD and Yeung
RS: Activated mammalian target of rapamycin pathway in the
pathogenesis of tuberous sclerosis complex renal tumors. Cancer
Res. 62:5645–5650. 2002.PubMed/NCBI
|
|
21
|
Kenerson H, Dundon TA and Yeung RS:
Effects of rapamycin in the Eker rat model of tuberous sclerosis
complex. Pediatr Res. 57:67–75. 2005.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Bissler JJ, Kingswood JC, Radzikowska E,
Zonnenberg BA, Frost M, Belousova E, Sauter M, Nonomura N,
Brakemeier S, de Vries PJ, et al: Everolimus for angiomyolipoma
associated with tuberous sclerosis complex or sporadic
lymphangioleiomyomatosis (EXIST-2): A multicentre, randomised,
double-blind, placebo-controlled trial. Lancet. 381:817–824.
2013.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Franz DN, Belousova E, Sparagana S, Bebin
EM, Frost M, Kuperman R, Witt O, Kohrman MH, Flamini JR, Wu JY, et
al: Efficacy and safety of everolimus for subependymal giant cell
astrocytomas associated with tuberous sclerosis complex (EXIST-1):
A multicentre, randomised, placebo-controlled phase 3 trial.
Lancet. 381:125–132. 2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Bissler JJ, McCormack FX, Young LR, Elwing
JM, Chuck G, Leonard JM, Schmithorst VJ, Laor T, Brody AS, Bean J,
et al: Sirolimus for angiomyolipoma in tuberous sclerosis complex
or lymphangioleiomyomatosis. N Engl J Med. 358:140–151.
2008.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Xiong X, Xie R, Zhang H, Gu L, Xie W,
Cheng M, Jian Z, Kovacina K and Zhao H: PRAS40 plays a pivotal role
in protecting against stroke by linking the Akt and mTOR pathways.
Neurobiol Dis. 66:43–52. 2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Chong ZZ: Targeting PRAS40 for multiple
diseases. Drug Discov Today. 21:1222–1231. 2016.PubMed/NCBI View Article : Google Scholar
|