Introduction
Spinal cord injury (SCI) is traumatic with extremely
high mortality and disability rates. The course of the trauma is
complex and variable. Besides, SCI can develop continuously in
different time stages, often resulting in the expansion of initial
injured area, and it has almost no self-repair ability and leads to
motor function loss (1). SCI can
lead to permanent paralysis, complete loss of sensation in affected
limbs and trunks, and loss of bowel, bladder and sexual function
(2). Besides, SCI also exerts
far-reaching impacts on individuals, families and the whole
society. High treatment costs and nursing expenditures have brought
huge economic burden to individuals, families and the society
(3). However, up to now, there is no
definite treatment method that is able to remarkably restore
neurological function after SCI (4).
After SCI, changes in local environment cause
changes in the physiological function of cells, such as neurons,
oligodendrocytes and astrocyte-spinal cords (ASCs) (5). These changes begin with the entry of
inflammatory cytokines into the injured spinal cord, releasing a
large number of cytokines and chemokines and thereby leading to
cell injury. Moreover, SCI triggers the reaction of ASCs and causes
their proliferation and hypertrophy (6,7), thus
facilitating the expression of proteins, such as glial fibrillary
acidic protein (GFAP), nestin and vimentin (8,9) and
finally causing scar formation by activating signaling pathways
such as STAT3 and transforming growth factor-β (TGF-β)/Smads
pathways (7,10,11).
Besides, ASCs also regulate their own activities in autocrine
manner (7,9).
Mesenchymal stem cells (MSCs) first found in bone
marrow can differentiate into osteoblasts, chondrocytes,
adipocytes, myoblasts and other functional cells in vitro
(12). According to related studies,
Basso, Beattie and Bresnahan (BBB) score system confirms that the
injection of bone marrow MSCs (BMSCs) in SCI rats can improve their
motor function and accelerate motor recovery during spinal
transplantation in mice with moderate SCI (13). Furthermore, BMSCs can also migrate to
the injured sites of spinal cord tissues to replace injured and
lost cells, thus enhancing motor function (14). Ankeny et al (15) found that BMSCs can stimulate the
regeneration of injured axons in the whole spinal cord and restore
motor control.
This study explored the effect of BMSCs on the motor
function repair of SCI rats and the role of the TGF-β/Smads
signaling pathway in the repair process, so as to provide
theoretical bases and potential therapeutic methods for SCI
repair-related research and clinical treatment of SCI.
Materials and methods
Main materials
Sprague Dawley (SD) rats, Dulbecco's modified
Eagle's medium (DMEM), fetal bovine serum (FBS), trypsin,
phosphate-buffered saline (PBS) and double-antibody (Gibco; Thermo
Fisher Scientific, Inc.), cluster of differentiation 34
(CD34)-phycoerythrin (PE), CD90-PE, CD105-PE, GFAP, TGF-β1, Smad2,
p-Smad2 and β-actin antibodies (Abcam), TGF-β1 (PeproTech) and 0.22
µm pinhole filter (EMD Millipore). This study was approved by the
Animal Ethics Committee of Jining No. 1 People's Hospital Animal
Center (Jining, China).
Establishment of rat SCI models
Rat SCI models were established using the Allen
method (16): After 10% chloral
hydrate (300 mg/kg) was injected intraperitoneally into the rats
(180-200 g), the skin was prepared locally, and the rats were fixed
on the operating Table in the prone position. No rat exhibited
signs of peritonitis after the administration of 10% chloral
hydrate. Then a 2-3 cm incision was made in the center of the back
with T10 spinous process as the center, so as to expose the T9-T11
spinous processes and vertebral plates, and the interspinous
ligament was cut off to expose T10 spinal cord. Subsequently, a 3.5
mm round plastic pad was placed on T10 spinal cord, and a 10 g iron
cone was used to hit against the pad in a free fall at a height of
3 cm to cause impact injury to the spinal cord. Thereafter, it was
observed that the rapid retraction and springing appeared in the
hind legs, and the tail gradually tilted and fell down. The
relaxation and collapse of the hind legs indicated successful
modeling. Next, the spinal cord was washed with warm normal saline,
and the incision was sutured. At 3 days after operation,
antibiotics were injected continuously, and the rats were fed
regularly. The movement of both hind limbs and tail were observed
and recorded daily.
Isolation and culture of BMSCs
Cervical dislocation (after being anesthetized using
10% chloral hydrate at a dose of 250 mg/kg) was used as the method
of euthanasia. No rat exhibited signs of peritonitis after the
administration of 10% chloral hydrate. We verified that the
experimental animals were properly sacrificed by observing the
indications of breathing and heartbeat. Then the rat tibia was
taken out under aseptic conditions and cleaned with PBS to open the
capitulum, and DMEM (with 1% double-antibody) containing 10% FBS
was used to wash the bone cavity through a syringe. Then the
harvested cell suspension was centrifuged at 4˚C, 1,050 x g for 10
min, and an appropriate amount of DMEM (with 1% double-antibody)
containing 10% FBS was added to suspend the precipitate again.
After counting, the cells were inoculated into a 100 mm culture
dish at the density of 1x105 cells/ml and cultured in an
incubator containing 5% CO2 at 37˚C. The solution was
replaced every other day until the cells covered 90% of the bottom
of the dish. After washing twice with PBS, the cells were digested
with 0.25% trypsin and subjected to passage at a density of
1:3.
Isolation, culture and purification of
ASCs
The spinal cord of SD rats was taken at 1-3 days
under aseptic conditions, the spinal membrane and vascular tissues
were removed in pre-cooled DMEM, and the remaining spinal cord
tissues were cut into 1 mm3 tissue blocks. After that,
an appropriate amount of 0.125% trypsin digestion solution was
added for digestion at 37˚C for 5 min, after which the tissues were
taken out and blown off. Then DMEM + 10% FBS culture solution was
added to terminate digestion, and the harvested cell suspension was
centrifuged at 4˚C, 1,050 x g for 10 min to collect precipitates.
After resuspension with DMEM + 10% FBS culture solution, the cells
were inoculated into a culture dish at 1x105 cells/ml
and cultured in the incubator containing 5% CO2 at 37˚C.
The solution was replaced every other day until the cells covered
90% of the bottom of the dish. Cell purification: Cells were
inoculated into a polylysine-coated culture dish at
1x105 cells/ml. Twenty-four hours later, a culture
solution containing 5 µg/ml of Ara-C was added and replaced with
DMEM + 10% FBS culture solution after 48 h. The solution was
replaced every 3 days, and the cells were continuously cultured for
~10 days.
Cell co-culture experiment
In vitro, ASCs and BMSCs were cultured and
proliferated, respectively. P3 ASCs and P4 BMSCs growing well were
taken for experiments and divided into 4 groups: ASCs culture group
(ASCs group), ASCs + 0.5 µg/l of TGF-β1 culture group (ASCs +
TGF-β1 group), ASCs + BMSCs co-culture group (ASCs + BMSCs group),
and ASCs + BMSCs + 0.5 µg/l of TGF-β1 culture group (ASCs + BMSCs +
TGF-β1 group). ASCs were inoculated into the lower layer of a
96-well Transwell plate at a density of 1x105 cells/ml,
while BMSCs were inoculated into the upper layer of the plate at a
density of 1x105 cells/ml. The basic culture solution
was DMEM + 10% FBS, and the cell proliferation was detected by
3-(4,5)-dimethylthiahiazo (-z-y1)-3,5-diphenyltetrazoliumbromide
(MTT) (Sigma-Aldrich; Merck KGaA) after 48 h of cell culture.
Subsequently, the cells were divided into two
groups, namely, ASCs group and ASCs + BMSCs group. ASCs were
inoculated into the lower layer of a 6-well Transwell plate at a
density of 1x105 cells/ml, whereas BMSCs were inoculated
into the upper layer of the plate at a density of 1x105
cells/ml. The basic culture solution was DMEM + 10% FBS, and the
protein expression of TGF-β1, Smad2 and p-Smad2 were examined by
western blotting (WB) after cell culture for 3 and 7 days,
respectively.
Cell injection
BMSCs were cultured and proliferated in
vitro, and P4 BMSCs in good growth status were collected, and
cell suspension was prepared at a density of 5x107
cells/ml. Subsequently, the rat models of SCI established above
were divided into SCI group and BMSCs transplantation group, and
sham operation (Sham) group was used as control. In BMSCs
transplantation group, 10 µl of cell suspension was injected into
the subarachnoid space at the spinal injury site at 30 min after
SCI modeling. In SCI group and Sham group, the same amount of
normal saline was injected.
MTT assay
The culture medium was discarded, BMSCs in the upper
layer was removed in ASCs + BMSCs group, with only ASCs in the
lower layer left. Then the remaining ASCs were washed with PBS 2-3
times, and 100 µl of fresh culture solution was added. Then, 20 µl
of MTT solution (5 mg/ml) was added to each well, and the culture
solution was removed from the well after continuous culture for 4
h. Thereafter, each well was added with 150 µl of dimethyl
sulfoxide (DMSO) and oscillated on a shaker at a low speed for 10
min, and the optical density at 490 nm in each well was measured
using enzyme-linked immunosorbent assay after the crystal was fully
dissolved.
Analysis via flow cytometry
Well-growing cells were analyzed via flow cytometry.
Then the cells were washed twice with PBS, digested with 0.25%
trypsin to form single cell suspension, and washed with PBS three
times to adjust the cell density to 3x105 cells/ml.
Thereafter, BMSCs were incubated with CD34-PE, CD90-PE and CD105-PE
flow antibodies at 4˚C in the dark, whereas ASCs were incubated
with CD34-PE flow antibody at 4˚C in the dark. After fixation, ASCs
were incubated with GFAP antibody and mouse anti-rabbit IgM/PE-Cy3.
Next, cell samples incubated with anti-rat IgG1-PE antibody were
used as the same type control. After 30 min, PBS was utilized for
washing 3 times, and flow cytometry (C6) was employed for detection
on the machine, followed by data analysis using CFlow Plus
software.
WB
Cells co-cultured in vitro or injured spinal
tissues were added with an appropriate amount of cell lysate for
lysis overnight at 4˚C, and the total protein was extracted by
centrifugation at 4˚C, 10,500 x g for 15 min, the concentration was
determined by bicinchoninic acid (BCA) (Pierce; Thermo Fisher
Scientific, Inc.). Then the protein was separated by 8% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis and then
transferred onto a polyvinylidene fluoride membrane (Roche
Diagnostics). After that, the membrane was sealed in Tris-buffered
saline with 0.1% Tween-20 and 5% skim milk powder, and TGF-β1,
Smad2, p-Smad2 and β-actin primary antibodies were used to gently
shake the membrane overnight at 4˚C. After the reaction with
primary antibody, horse radish peroxidase (HRP)-labeled secondary
antibody was applied for incubation. Finally, the exposed detection
protein was treated with electrochemiluminescence (ECL)
reagent.
BBB score
The spinal motor function of rats was scored by BBB
behavioral function evaluation method (17) (21 grades in total). The rats were
placed in an open basin, and the basin wall was knocked to make the
rats crawl. The movement of each joint part was observed and
recorded, and the motor function of each rat was evaluated
according to scoring rules. The observation time was 5 min each
time, and the final score was the average score of bilateral hind
limbs.
Statistical analysis
Statistical Product and Service Solutions (SPSS)
20.0 software (IBM, Corp.) was adopted to process the data. The
data in each group were expressed as mean ± standard deviation
(mean ± SD), and the intergroup comparison was conducted using
independent samples t-test. P<0.05 indicates that the difference
is statistically significant.
Results
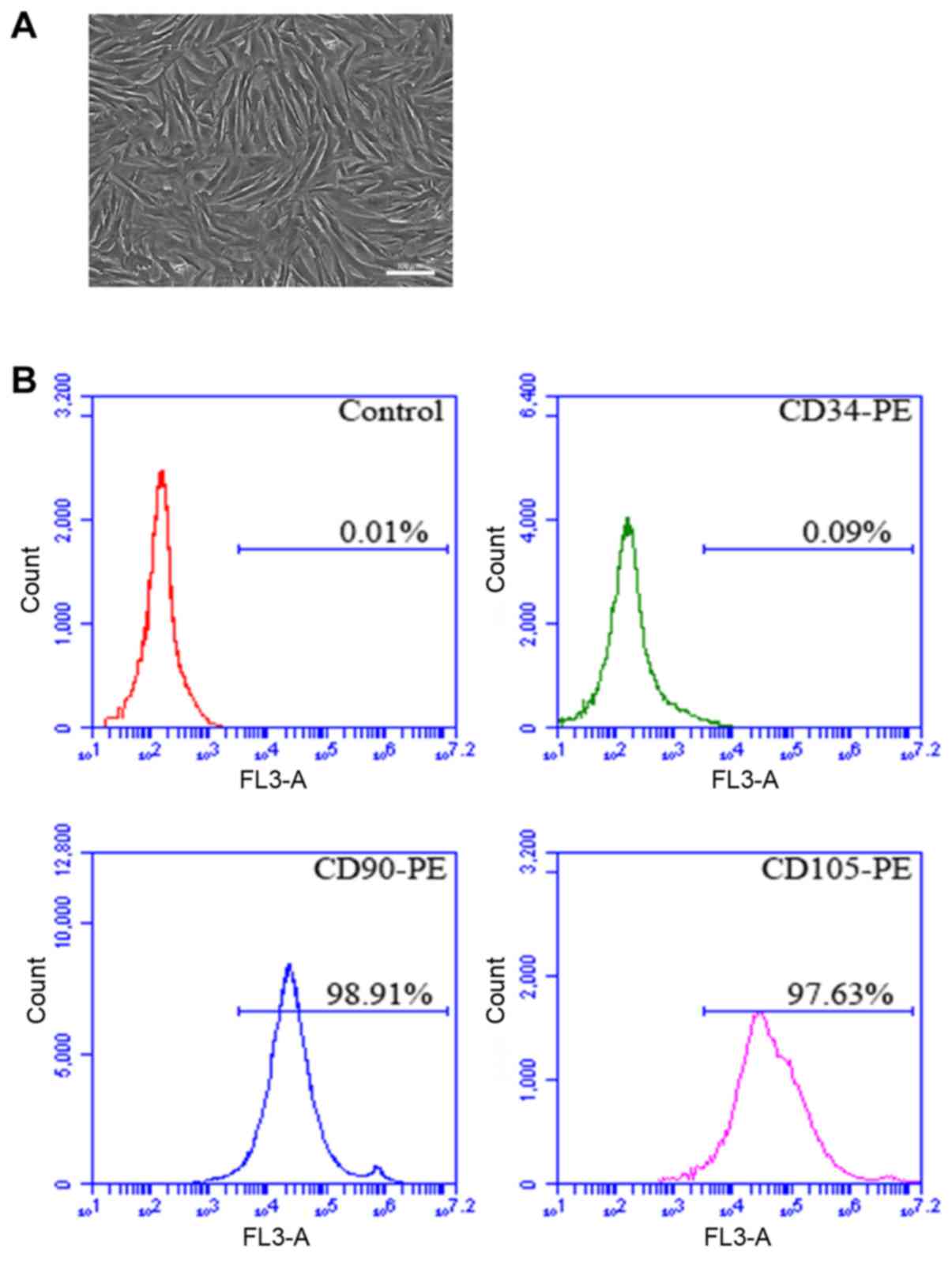
Isolation and culture of rat BMSCs in
vitro
The rat tibia was isolated in sterile environment,
the bone marrow was obtained, and BMSCs were isolated and cultured
in vitro. After amplification and culture, the cells
displayed plump morphology and were spindle-like growing in spiral
shape (Fig. 1A). In addition, flow
cytometry results manifested that the positive rates of CD90 and
CD105 in P4 cells were 98.91 and 97.63%, respectively, and CD45 was
not expressed (Fig. 1B).
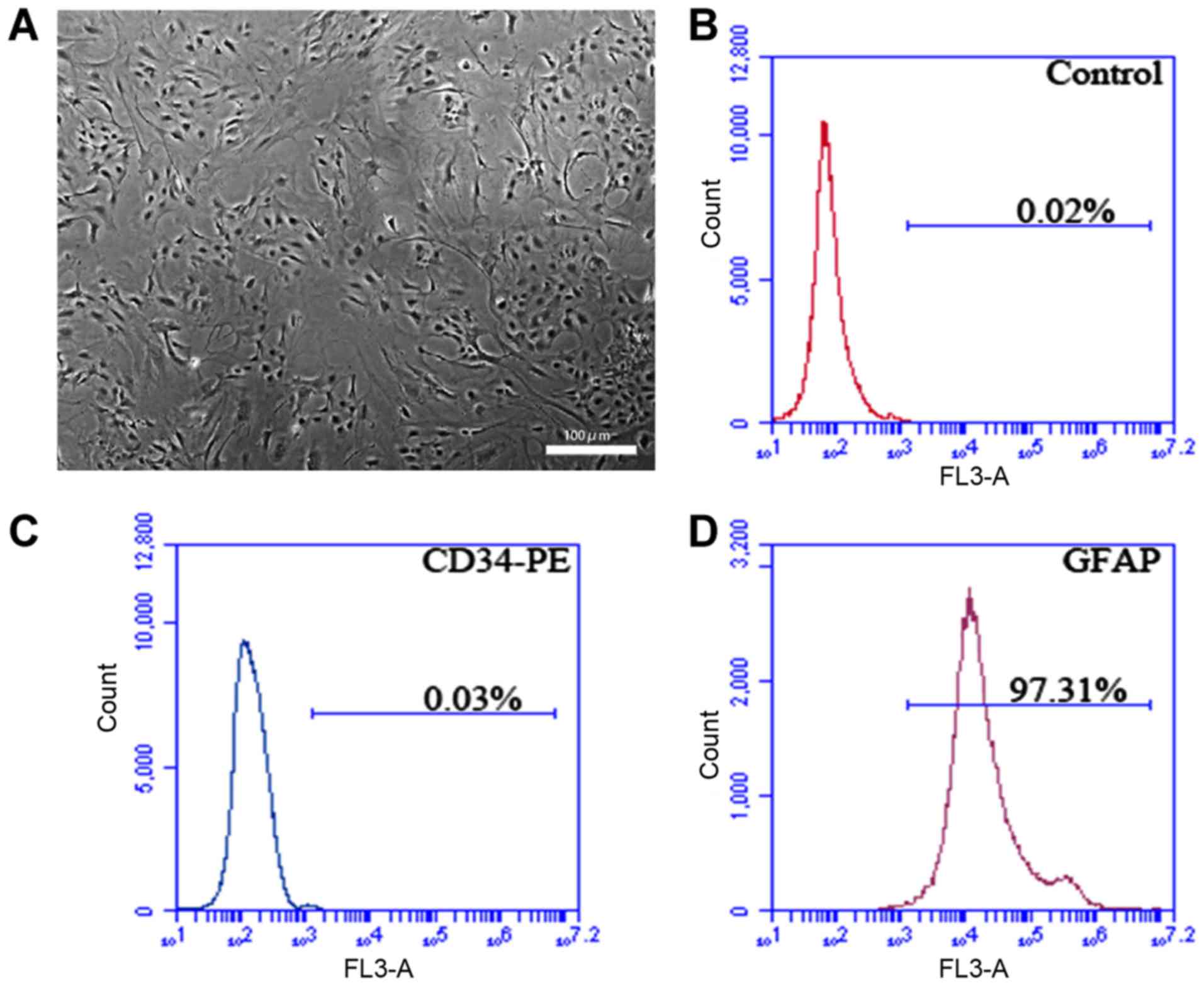
Isolation and culture of rat ASCs in
vitro
The rat spine was isolated in sterile environment,
and ASCs obtained were isolated and cultured in vitro,
showing normal morphology (Fig. 2A).
According to flow cytometry detection, the positive rate of the
specific marker of P3 ASCs GFAP was 97.31%, and CD34 was not
expressed (Fig. 2B-D).
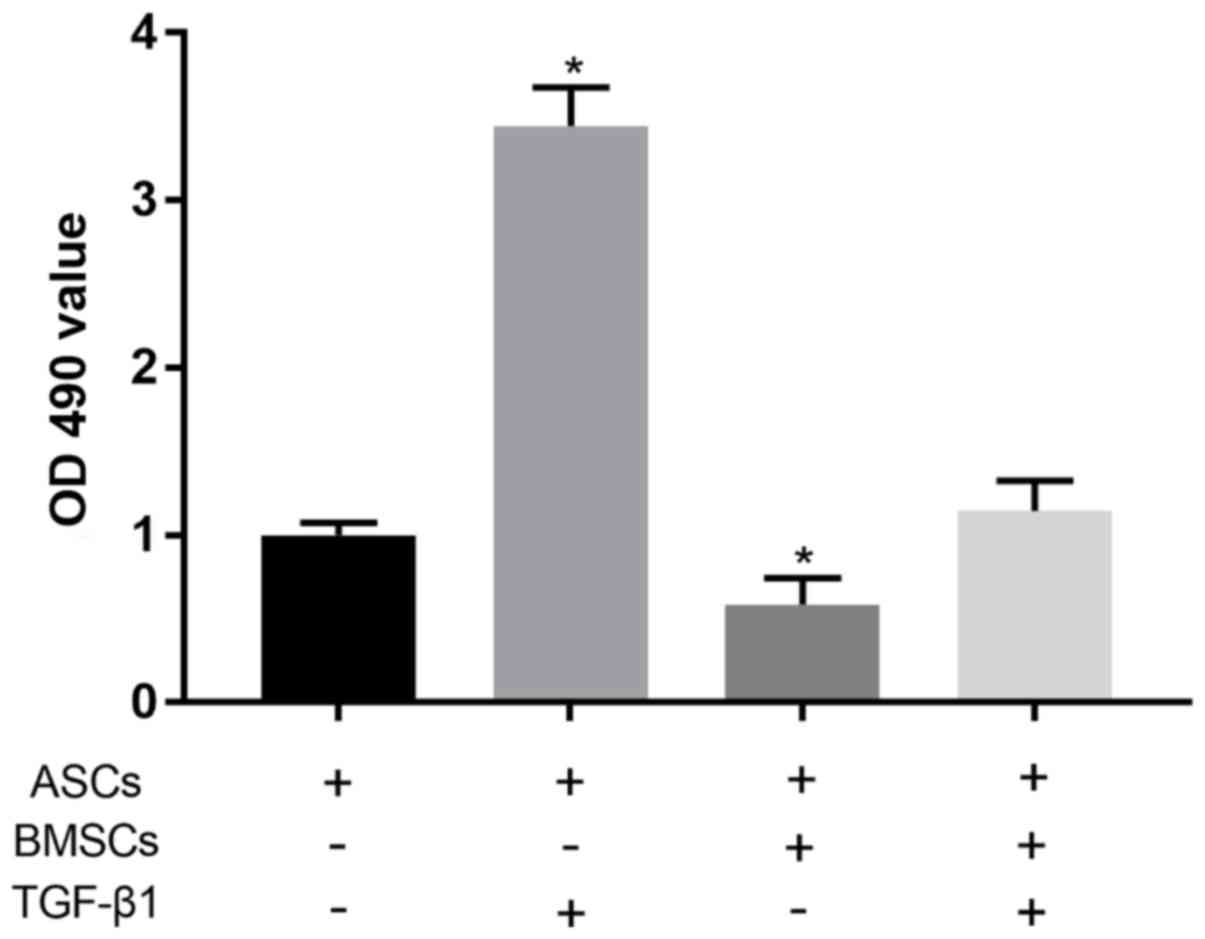
Co-culture experiment of ASCs
MTT assay showed that compared with normally
cultured ASCs, 0.5 µg/l TGF-β1 could significantly promote the
proliferation of ASCs (P<0.05). Co-culture of ASCs and BMSCs was
able to significantly reduce the proliferation level of ASCs
(P<0.05). After the co-culture of ASCs and BMSCs under the
effect of TGF-β1, there was no significant difference in the
proliferation level of ASCs compared with that in control group
(P>0.05) (Fig. 3).
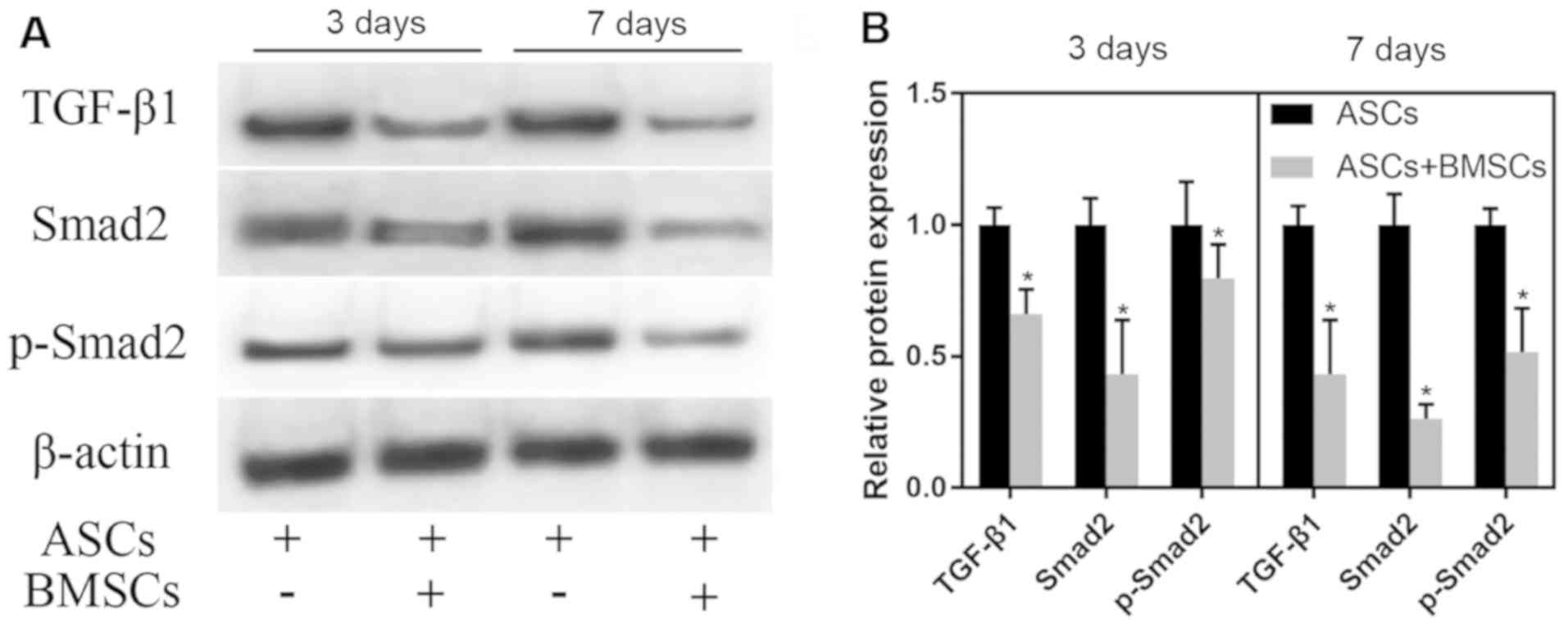
Effect of co-culture of ASCs and BMSCs
on the TGF-β/Smads pathway
ASCs were co-cultured with BMSCs in vitro.
With normally cultured ASCs as control, ASCs were cultured for 3
and 7 days. WB test results demonstrated that the co-culture
notably reduced the expression of TGF-β1, Smad2 and p-Smad2 in ASCs
(P<0.05). With the extension of co-culture, the degree of the
decrease in the expression of TGF-β1, Smad2 and p-Smad2 in ASCs
after 3 days of co-culture with BMSCs was lower than that in ASCs
after 7 days of co-culture with BMSCs (Fig. 4).
Effect of transplantation of BMSCs on
SCI in rats
Transplanted BMSCs were injected into the
subarachnoid space at the spinal injury site of SCI model rats. The
results manifested that BBB score in Sham group was 21 points, and
there were no differences at 1, 3 and 5 weeks. BBB score at 1, 3
and 5 weeks in SCI group was 0.33±0.47 points, 1.27±0.44 points and
3.33±0.47 points, respectively, that in SCI + BMSCs group was
0.73±0.57 points, 3.13±0.81 points and 8.00±2.56 points,
respectively, and that in SCI + PBS group were 0.40±0.49 points,
1.33±0.47 points and 2.80±0.40 points, respectively. It was found
that the BBB score in SCI + BMSCs group was evidently higher than
that in SCI group and SCI + PBS group at 3 and 5 weeks (P<0.05),
but no obvious difference was found in BBB score between SCI group
and SCI + PBS group (P>0.05) (Fig.
5).
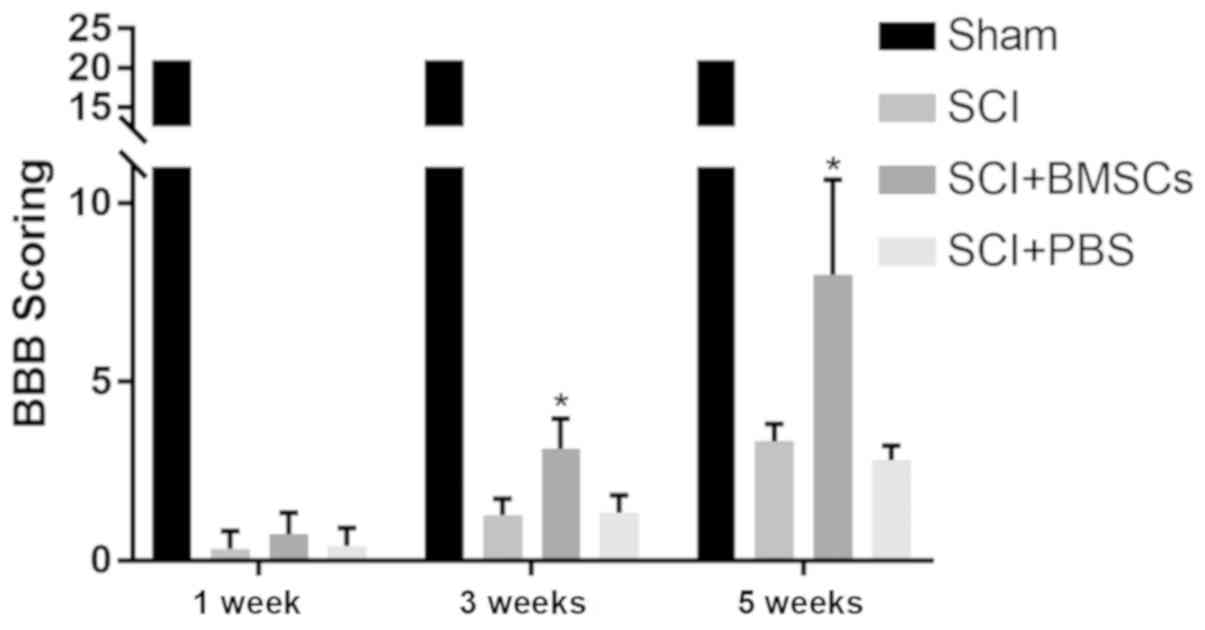 | Figure 5BBB score. At 3 and 5 weeks in SCI +
BMSCs group, BBB score is predominantly higher than that in SCI
group and SCI + PBS group (P<0.05), but no obvious difference is
found in BBB score between SCI group and SCI + PBS group
(P>0.05). *P<0.05, obvious difference is detected
in comparison among SCI + BMSCs group, SCI group and SCI + PBS
group (P<0.05). Sham: sham operation group, SCI: SCI model
group, SCI + BMSCs: SCI + BMSCs injection group, SCI + PBS: SCI +
PBS injection group. BMSCs, bone marrow mesenchymal stem cells;
BBB, Basso, Beattie and Bresnahan; SCI, spinal cord injury; PBS,
phosphate-buffered saline. |
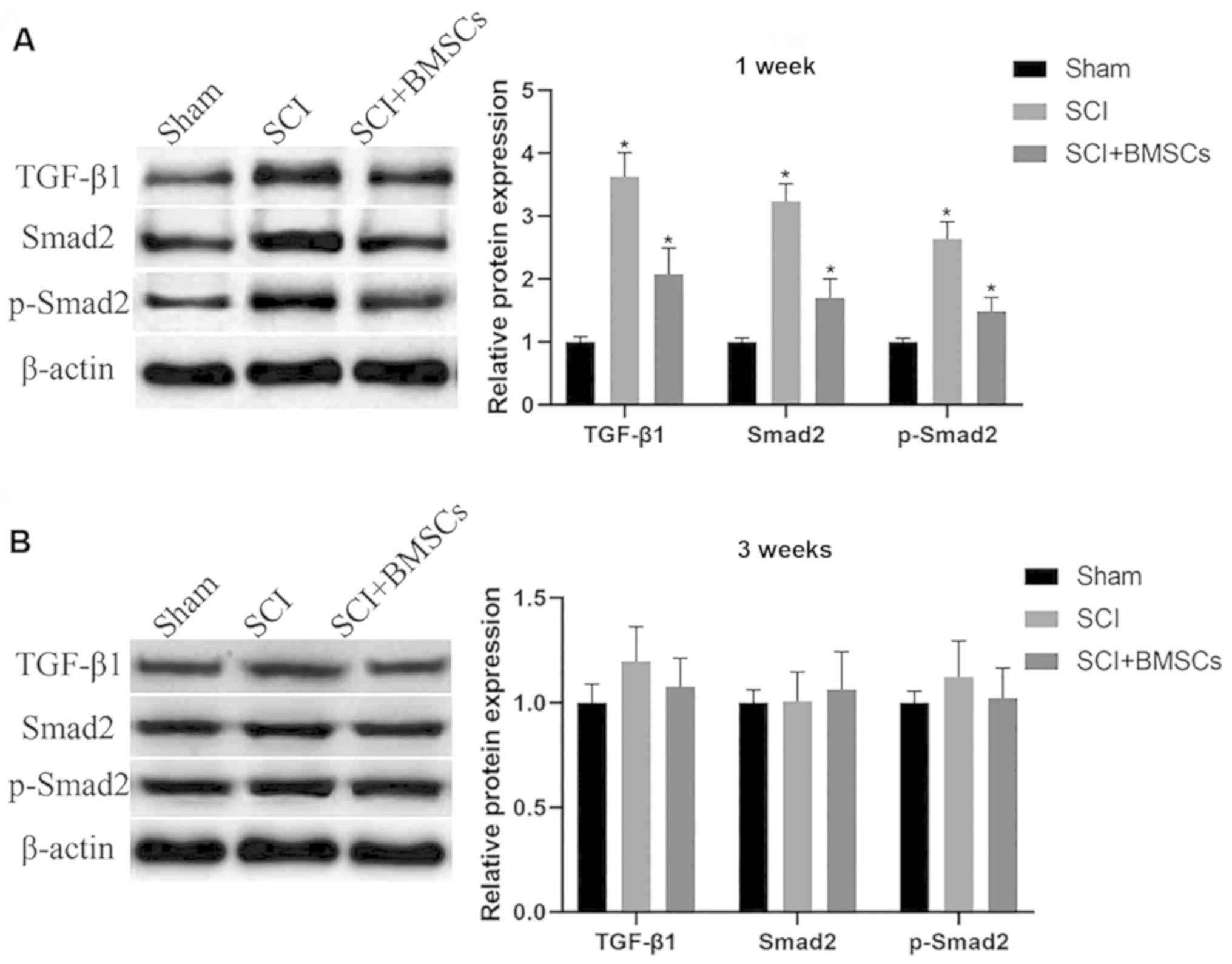
Influence of the transplantation of
BMSCs on the TGF-β/Smads pathway at the spinal injury site of
rats
Transplanted BMSCs were injected into the
subarachnoid space at the spinal injury site of SCI model rats. WB
was used to detect protein expression at the spinal injury site
after 1 and 3 weeks, and the results demonstrated that the
expression of TGF-β1, Smad2 and phosphorylated (p)-Smad2 in SCI
group were obviously upregulated compared with those in Sham group
at 1 weeks (P<0.05), and injection of BMSCs markedly
downregulated the expression (P<0.05). After 3 weeks, there was
no significant difference in the expression of TGF-β1, Smad2 and
p-Smad2 among the groups (P>0.05) (Fig. 6).
Discussion
SCI mainly leads to nerve conduction interruption,
spinal cord structure injury and neuron necrosis, thus resulting in
tissue function loss. ASCs are the main glial cell population in
the mammalian nervous system, playing an important role. When
injuries or diseases such as trauma, neurodegeneration or ischemia
occur, ASCs are in a reactive state, manifested as cell
hypertrophy, gene expression changes, cell proliferation and other
characteristics (9,10). The proliferation of ASCs triggers
increased production of extracellular matrix, such as chondroitin
sulfate proteoglycan (CSPG) and class I ethylene glycol conjugate
(18). CSPG overexpression is
closely associated with the formation of glial scars, thus
hindering the regeneration and growth of axons (6). After culture in vitro, the
proliferation ability of ASCs was evidently enhanced in the
presence of TGF-β1, suggesting that ASCs can be stimulated by TGF-β
accordingly. Studies have shown that TGF-β increases the expression
of calcium ion channel of ASCs as well as the expression of GFAP
and CSPGs, thus facilitating the increased number of proliferative
ASCs (19).
The TGF-β/Smads signaling pathway mainly consists of
TGF-β, TGF-β receptors, Smads and various proteins in the nucleus.
TGF-β activates TGF-β receptors, and the activated receptor further
phosphorylates Smad2 and other proteins. Then the p-Smad2 combines
with other proteins into a complex and enters the nucleus to
further function (20). The
TGF-β/Smads signaling pathway exerts crucial effects in the process
of fibrosis of tissues and organs. When SCI occurs, microglia
activation and macrophage invasion are the first reactions. These
activated cells release large amounts of cytokines and growth
factors, including TGF-β, basic fibroblast growth factor,
interleukin-6 and interleukin-1(21). The produced TGF-β further modulates
smooth muscle cells, fibroblasts and reactive ASCs (22-24).
Moreover, the proliferation ability of ASCs was notably enhanced
through the culture of ASCs with TGF-β1.
The TGF-β/Smads signaling pathway plays a vital role
in the development process of hypertrophic scars, in which it
promotes tissue scar formation and fibrosis. The main physiological
mechanism is the high expression of various cytokines in this
pathway and the inhibition on the expression of anti-Smads
(25). A study of Chin et al
(25) revealed that the expression
levels of TGF-β1 and TGF-β2 ligands in scar fibroblasts are higher,
the expression of TGF-β receptors (type I and II) is increased, and
the phosphorylation of Smad3 is raised. O'Brien et al
(26) found that ASCs produce
complex reactions within 24 h after SCI, and extracellular TGF-β
with high concentration is produced immediately after injury.
Subsequently, intracellular TGF-β production is observed in the
nucleus and cytoplasm of ASCs, intramedullary and extramedullary
capillary endothelial cells, and motor neurons. In this study, the
upregulated protein expression of TGF-β1, Smad2 and p-Smad2 were
detected at the SCI site of SCI rats, consistent with previous
research results. It can be concluded that a large quantity of
extracellular TGF-β is produced after SCI, which further activates
the TGF-β/Smads signaling pathway of ASCs, thus causing massive
proliferation of ASCs and generation of hyperplastic scars and
hindering axonal regeneration and growth of injured spinal cords
(6).
BMSCs have been applied to the treatment of various
diseases. After transplanting BMSCs into the injured spinal cord of
rats, the function of BMSCs can be improved moderately (27). According to studies, BMSCs can
release anti-inflammatory cytokines and neurotrophic factors to
protect neurons (28,29). In this study, it was discovered from
co-culture of BMSCs and ASCs that BMSCs weakened the proliferation
ability of ASCs and reduced the expression levels of TGF-β1, Smad2
and p-Smad2, possible because BMSCs attenuate the effect of TGF-β1
on ASCs through interaction with TGF-β1. Furthermore, SCI rats were
treated by transplantation of BMSCs. BBB score displayed that the
transplantation improved the recovery of injured spinal cords in
animal models, and reduced the protein expression levels of TGF-β1,
Smad2 and p-Smad2 in the TGF-β/Smads signaling pathway.
In conclusion, the results of this study verify that
the transplantation of BMSCs is able to improve the spinal function
of ι SCI rats probably by inhibiting the TGF-β/Smads signaling
pathway and reducing the proliferation of ASCs.
Acknowledgements
Not applicable.
Funding
National Natural Science Foundation of China (NSFC)
(no. 81801906); Natural Science Foundation of Shandong Province
(no. ZR2015YL034).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
CL and KG designed the study and performed the
experiments, CL and TZ collected the data, KG and KL analyzed the
data, CL and KG prepared the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Animal Ethics
Committee of Jining No. 1 People's Hospital Animal Center (Jining,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Himes BT, Neuhuber B, Coleman C, Kushner
R, Swanger SA, Kopen GC, Wagner J, Shumsky JS and Fischer I:
Recovery of function following grafting of human bone
marrow-derived stromal cells into the injured spinal cord.
Neurorehabil Neural Repair. 20:278–296. 2006.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Li JW, Kuang Y, Chen L and Wang JF: LncRNA
ZNF667-AS1 inhibits inflammatory response and promotes recovery of
spinal cord injury via suppressing JAK-STAT pathway. Eur Rev Med
Pharmacol Sci. 22:7614–7620. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Priebe MM, Chiodo AE, Scelza WM, Kirshblum
SC, Wuermser LA and Ho CH: Spinal cord injury medicine. Economic
and societal issues in spinal cord injury. Arch Phys Med Rehabil.
88 (Suppl 1):S84–S88. 2007.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Chhabra HS and Arora M: Demographic
profile of traumatic spinal cord injuries admitted at Indian Spinal
Injuries Centre with special emphasis on mode of injury: A
retrospective study. Spinal Cord. 50:745–754. 2012.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Karimi-Abdolrezaee S and Billakanti R:
Reactive astrogliosis after spinal cord injury-beneficial and
detrimental effects. Mol Neurobiol. 46:251–264. 2012.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Fitch MT and Silver J: Glial cell
extracellular matrix: Boundaries for axon growth in development and
regeneration. Cell Tissue Res. 290:379–384. 1997.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Fitch MT and Silver J: CNS injury, glial
scars, and inflammation: Inhibitory extracellular matrices and
regeneration failure. Exp Neurol. 209:294–301. 2008.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Sofroniew MV and Vinters HV: Astrocytes:
Biology and pathology. Acta Neuropathol. 119:7–35. 2010.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Sofroniew MV: Molecular dissection of
reactive astrogliosis and glial scar formation. Trends Neurosci.
32:638–647. 2009.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Silver J and Miller JH: Regeneration
beyond the glial scar. Nat Rev Neurosci. 5:146–156. 2004.PubMed/NCBI View
Article : Google Scholar
|
|
11
|
Herrmann JE, Imura T, Song B, Qi J, Ao Y,
Nguyen TK, Korsak RA, Takeda K, Akira S and Sofroniew MV: STAT3 is
a critical regulator of astrogliosis and scar formation after
spinal cord injury. J Neurosci. 28:7231–7243. 2008.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ji M, Bai C, Li L, Fan Y, Ma C, Li X and
Guan W: Biological characterization of sheep kidney-derived
mesenchymal stem cells. Exp Ther Med. 12:3963–3971. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Lin GL, Wang H, Dai J, Li X, Guan M, Ding
Q, Wang HX and Fang H: Upregulation of UBAP2L in bone marrow
mesenchymal stem cells promotes functional recovery in rats with
spinal cord injury. Curr Med Sci. 38:1081–1089. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Syková E, Jendelová P, Urdzíková L, Lesný
P and Hejcl A: Bone marrow stem cells and polymer hydrogel - two
strategies for spinal cord injury repair. Cell Mol Neurobiol.
26:1113–1129. 2006.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ankeny DP, McTigue DM and Jakeman LB: Bone
marrow transplants provide tissue protection and directional
guidance for axons after contusive spinal cord injury in rats. Exp
Neurol. 190:17–31. 2004.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Namjoo Z, Mortezaee K, Joghataei MT,
Moradi F, Piryaei A, Abbasi Y, Hosseini A and Majidpoor J:
Targeting axonal degeneration and demyelination using combination
administration of 17β-estradiol and Schwann cells in the rat model
of spinal cord injury. J Cell Biochem. 119:10195–10203.
2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Basso DM, Beattie MS and Bresnahan JC: A
sensitive and reliable locomotor rating scale for open field
testing in rats. J Neurotrauma. 12:1–21. 1995.PubMed/NCBI View Article : Google Scholar
|
|
18
|
McKeon RJ, Jurynec MJ and Buck CR: The
chondroitin sulfate proteoglycans neurocan and phosphacan are
expressed by reactive astrocytes in the chronic CNS glial scar. J
Neurosci. 19:10778–10788. 1999.PubMed/NCBI
|
|
19
|
Yu Z, Yu P, Chen H and Geller HM: Targeted
inhibition of KCa3.1 attenuates TGF-β-induced reactive astrogliosis
through the Smad2/3 signaling pathway. J Neurochem. 130:41–49.
2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Leask A and Abraham DJ: TGF-beta signaling
and the fibrotic response. FASEB J. 18:816–827. 2004.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Hausmann ON: Post-traumatic inflammation
following spinal cord injury. Spinal Cord. 41:369–378.
2003.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wang H, Katagiri Y, McCann TE, Unsworth E,
Goldsmith P, Yu ZX, Tan F, Santiago L, Mills EM, Wang Y, et al:
Chondroitin-4-sulfation negatively regulates axonal guidance and
growth. J Cell Sci. 121:3083–3091. 2008.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Schönherr E, Järveläinen HT, Sandell LJ
and Wight TN: Effects of platelet-derived growth factor and
transforming growth factor-beta 1 on the synthesis of a large
versican-like chondroitin sulfate proteoglycan by arterial smooth
muscle cells. J Biol Chem. 266:17640–17647. 1991.PubMed/NCBI
|
|
24
|
Heimer R, Bashey RI, Kyle J and Jimenez
SA: TGF-beta modulates the synthesis of proteoglycans by myocardial
fibroblasts in culture. J Mol Cell Cardiol. 27:2191–2198.
1995.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Chin GS, Liu W, Peled Z, Lee TY,
Steinbrech DS, Hsu M and Longaker MT: Differential expression of
transforming growth factor-beta receptors I and II and activation
of Smad 3 in keloid fibroblasts. Plast Reconstr Surg. 108:423–429.
2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
O’Brien MF, Lenke LG, Lou J, Bridwell KH
and Joyce ME: Astrocyte response and transforming growth
factor-beta localization in acute spinal cord injury. Spine.
19:2321–2330. 1994.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Ohta M, Suzuki Y, Noda T, Ejiri Y, Dezawa
M, Kataoka K, Chou H, Ishikawa N, Matsumoto N, Iwashita Y, et al:
Bone marrow stromal cells infused into the cerebrospinal fluid
promote functional recovery of the injured rat spinal cord with
reduced cavity formation. Exp Neurol. 187:266–278. 2004.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Neuhuber B, Timothy Himes B, Shumsky JS,
Gallo G and Fischer I: Axon growth and recovery of function
supported by human bone marrow stromal cells in the injured spinal
cord exhibit donor variations. Brain Res. 1035:73–85.
2005.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Dai G, Liu X, Zhang Z, Yang Z, Dai Y and
Xu R: Transplantation of autologous bone marrow mesenchymal stem
cells in the treatment of complete and chronic cervical spinal cord
injury. Brain Res. 1533:73–79. 2013.PubMed/NCBI View Article : Google Scholar
|