Introduction
Coronary heart disease is a major cause of death and
disability that causes serious debilitation in patients, with an
incidence rate that is increasing on a yearly basis (1,2). Acute
myocardial infarction (MI) is a coronary heart disease with high
worldwide incidence and mortality rates that leads to myocardial
tissue injury and ultimately disability or death (3). Hypoxia is one of the primary causes of
MI. Under hypoxic conditions, changes in energy metabolism occur
during mitochondrial respiration in which anaerobic glycolysis
results in numerous physiological and pathological reactions
leading to acidosis and necrosis (4). Therefore, understanding the
pathogenesis of MI may highlight potentially novel targets to limit
damage.
MicroRNAs (miRNAs or miRs) are small single-stranded
non-coding RNAs that are 22-25 nucleotides in length (5,6). miRNAs
post-transcriptionally regulate gene expression (7-9) and
mediate the degradation of or inhibit the translation of mRNAs by
directly binding to their 3' untranslated region (3'-UTR) (10,11).
Numerous studies have demonstrated that miRNAs serve an important
role in the pathogenesis of many types of cancer (12-14),
including miR-663b, indicating that they may be involved in the
occurrence and development of tumors (15).
Pan et al (16) demonstrated that miR-663b is
downregulated in gastric cancer and Jiao et al (17) revealed that expression was increased
in castration-resistant prostate cancer. Additionally, miR-663b is
upregulated in nasopharyngeal carcinoma (18). Liang et al (19) demonstrated that miR-663b promoted
proliferation by targeting tumor suppressor candidate 2 and that
the resulting upregulation was associated with advanced clinical
stage cancer and lymph node metastasis. miR-663b expression is also
upregulated in patients with MI (20). However, its role and function in MI
remain unknown.
The aim of the present study was to determine
whether miR-663b served a role in hypoxia-induced cardiomyocyte
injury in vitro and to study the specific underlying
molecular mechanisms. The present study may provide a theoretical
basis for the development of novel strategies for the treatment of
MI.
Materials and methods
Cell culture and transfection
Rat cardiomyocyte H9c2 cells were purchased from the
Cell Bank of the Type Culture Collection of the Chinese Academy of
Sciences. H9c2 cells were cultured in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) and incubated at 37˚C with 5% CO2. To
simulate hypoxic damage, cells were incubated for 48 h at 37˚C with
94% N2, 5% CO2 and 1% O2 (21). Cells in the control group were
incubated at 37˚C with 5% CO2.
Under hypoxic conditions, H9c2 cells were
transfected with 50 nM miR-663b inhibitor
(5'-GCGGUCCCGCGGCGCCCCGCCU-3'; Shanghai GenePharma Co., Ltd.), 50
nM inhibitor control (5'-UUGUACUACACAAAAGUACUG-3'; Shanghai
GenePharma Co., Ltd.), 50 nM miR-663b inhibitor + 1 µM
control-small interfering (si)RNA (cat. no. sc-36869; Santa Cruz
Biotechnology, Inc.) or 50 nM miR-663b inhibitor + 1 µM B-cell
lymphoma 2 like 1 (BCL2L1)-siRNA (cat. no. sc-29216; Santa Cruz
Biotechnology, Inc.) for 48 h using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Transfection efficiency was assessed using
reverse transcription quantitative PCR (RT-qPCR) 48 h after
experimentation.
RT-qPCR
Total RNA was extracted from H9c2 cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). RNA was reverse transcribed into cDNA using a cDNA Reverse
Transcription kit (Vazyme, Biotech Co., Ltd.) at a temperature of
42˚C for 60 min and 75˚C for 5 min. qPCR was performed using a
SYBR™ Green PCR Master kit (Vazyme Biotech Co., Ltd.). The
thermocycling conditions were as follows: Initial denaturation at
95˚C for 3 min; followed by 40 cycles of 95˚C for 30 sec, 56˚C for
30 sec and 72˚C for 30 sec. Primer sequences used for PCR were
listed as follows: miR-663b forward, 5'-CGCTAACAGTCTCCAGTC-3' and
reverse 5'-GTGCAGGGTCCGAGGT-3'; BCL2L1 forward,
5'-GACTGAATCGGAGATGGAGACC-3' and reverse
5'-GCAGTTCAAACTCGTCGCCT-3'; U6, 5'-GCTTCGGCAGCACATATACTAAAAT-3';
reverse, 5'-CGCTTCACGAATTTGCGTGTCAT-3'; GAPDH forward,
5'-CTTTGGTATCGTGGAAGGACTC-3' and reverse,
5'-GTAGAGGCAGGGATGATGTTCT-3'. The fold change of gene expression
was calculated using the 2-ΔΔCq method (22). U6 and GAPDH were used as the internal
controls for normalizing expression of miRNA and mRNA,
respectively. All experiments were performed in triplicate.
Dual-luciferase reporter assay
TargetScan analysis (version 7.2; http://www.targetscan.org/vert_72/) was performed
to examine the association between miRNA-663b and BCL2L1. The
results indicated that miR-663b binding sites were present in
BCL2L1. To confirm binding between miR-663 and BCL2L1, a dual
luciferase reporter assay was performed.
Luciferase reported plasmids (psi-CHECK2) containing
BCL2L1 wild-type (WT-BCL2L1) or mutant (MUT-BCL2L1) 3'-UTRs were
synthesized by TsingKe Biotech Co. H9c2 cells were co-transferred
with WT-BCL2L1 or MUT-BCL2L1 luciferase reporter plasmids and the
miR-663b mimic or mimic control, respectively, using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Cells were harvested 48 h after transfection and
luciferase activity was measured using a
Dual-Luciferase® Reporter Assay system (Promega
Corporation) according to the manufacturer's protocol. Firefly
luciferase was used to normalize data.
Western blotting
Total protein was extracted from H9c2 cells using
RIPA lysis buffer (Beijing Solarbio Science & Technology Co.,
Ltd.) at room temperature for 30 min, after which samples were
stored at -20˚C. A bicinchoninic acid assay kit (Pierce; Thermo
Fisher Scientific, Inc.) was used to quantify protein
concentration. Equal quantities of protein (30 µg/lane) were loaded
onto a 15% gel, resolved using SDS-PAGE and transferred onto PVDF
membranes (EMD Millipore). Membranes were blocked with 5% skimmed
milk in TBS containing 0.1% Tween at room temperature for 2 h.
Subsequently, the membranes were incubated with the following
primary antibodies: Anti-BCL2L1 (cat. no. sc-70418; 1:1,000; Santa
Cruz Biotechnology, Inc.), anti-cleaved-Caspase-3 (cat. no. ab2302;
1:1,000; Abcam) or anti-GAPDH (cat. no. 5174; 1:1,000; Cell
Signaling Technology, Inc.) at 4˚C overnight. The membranes were
incubated with horseradish peroxidase-conjugated goat anti-rabbit
immunoglobulin G secondary antibodies (cat. no. 7074; 1:2,000; Cell
Signaling Technology, Inc.) at room temperature for 2 h. Signals
were visualized using enhanced chemiluminescence reagent (EMD
Millipore) and GAPDH was used as the loading control.
Flow cytometry analysis
Transfected cells (1x106 cells) were
harvested, centrifuged at a low temperature at a high speed (1,000
x g; 4˚C; 5 min) and re-suspended in 200 µl binding buffer
containing 10 µl fluorescein isothiocyanate-annexin V. After
centrifugation, cells were re-suspended in 5 µl propidium iodide
(PI) and 300 µl PBS, and incubated for 30 min at room temperature
in the dark. Cells were then stained using annexin-V/PI Apoptosis
Detection kit (BD Biosciences) according to the manufacturer's
protocol. Fluorescence was assessed using a BD FACSCalibur™ flow
cytometer (Becton, Dickinson and Company) and flow data was
analyzed using FlowJo (version 7.6.1; FlowJo LLC).
MTT assay
H9c2 cells were transfected with miR-663b inhibitor,
inhibitor control, miR-663b inhibitor + control-siRNA or miR-663b
inhibitor + BCL2L1-siRNA for 48 h under hypoxic conditions.
Transfected cells were then plated in a 96-well plate at a density
of 5x103 cells/well. After 24 h of incubation at 37˚C,
20 µl MTT reagent (Sigma-Aldrich; Merck KGaA) was added to each
well. Following a further 4 h of incubation at 37˚C, 150 µl DMSO
was added to each well and agitated for 15 min in the dark. Optical
density values were measured at 490 nm using a microplate
reader.
ELISA
Levels of certain inflammatory cytokines, including
tumour necrosis factor α (TNF-α; cat. no. PT516), interleukin (IL)
1β (cat. no. PI303) and IL-6 (cat. no. PI328) in cell supernatants
were detected using specific ELISA kits (Beyotime Institute of
Biotechnology) according to the manufacturer's protocol.
Assessment of cellular injury
Following treatment, the release of various
cardiomyocyte injury biomarkers, including creatine
kinase-muscle/brain (CK-MB; cat. no. CSB-E14403r; Cusabio
Technology LLC) and cardiac troponin I (cTnI; cat. no. CSB-E08594r;
Cusabio Technology LLC) was determined using specific ELISA kits
according to the manufacturer's protocol. Mitochondrial viability
was measured using a mitochondrial viability staining kit (cat. no.
ab129732; Abcam) according to the manufacturer's protocol.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software (version 5; GraphPad Software, Inc.). Statistical
differences between groups were determined using a Student's t-test
or one-way ANOVA followed by Tukey's post hoc test. Data are
expressed as the mean ± standard deviation of at least three
independent experiments. P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-663b expression is significantly
increased in hypoxia-induced H9c2 cells
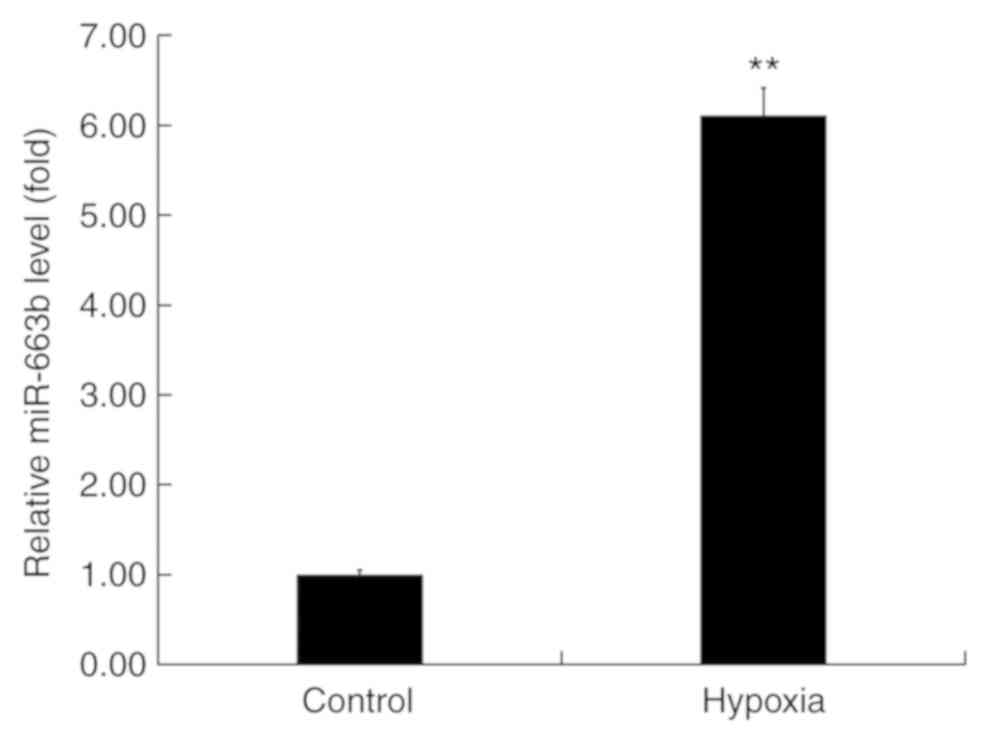
To examine the role of miR-663b in hypoxia-induced
H9c2 cells, miR-663b expression was assessed using RT-qPCR. The
results revealed that miR-663b expression was significantly
upregulated in H9c2 cells compared with the control group (Fig. 1), indicating that miR-663b may be
associated with the development of MI.
BCL2L1 is a direct target gene of
miR-663b
To determine the role and mechanism by which miR-663
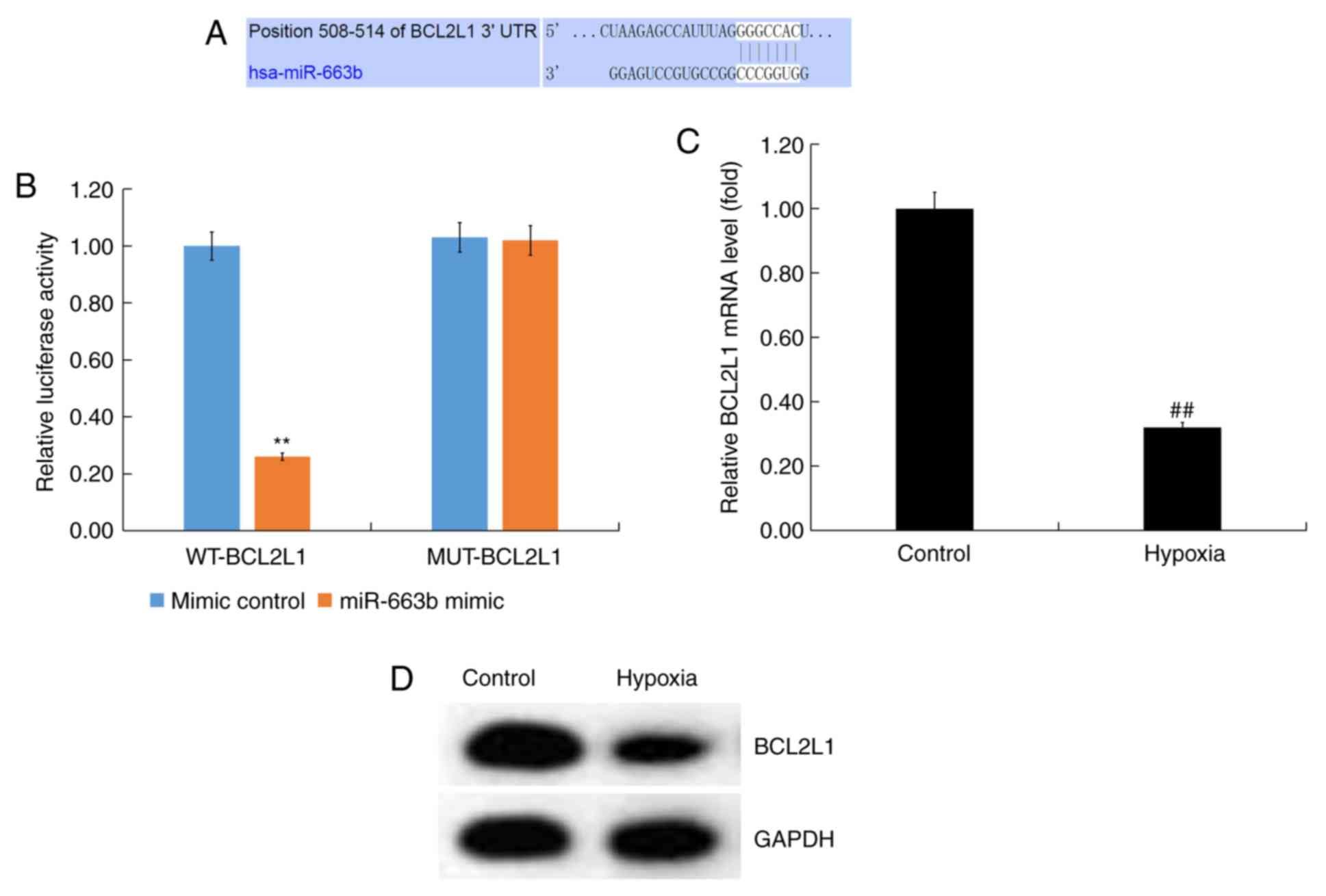
exerted its effects, bioinformatics analysis was performed using
TargetScan to predict miR-663b targets. BCL2L1 was predicted to be
a target gene of miR-663b (Fig.
2A).
The 3'-UTR (WT or MUT) of BCL2L1 was inserted into
the pmiR luciferase reporter and H9c2 cells were co-transfected
with the miR-663b mimic and luciferase reporter plasmid. The
results revealed that miR-663b mimic co-transfection with the
WT-BCL2L1 3'-UTR reporter inhibited luciferase activity while
miR-663b mimic had no effect on the reporter containing the
MUT-BCL2L1 3'-UTR (Fig. 2B). These
data suggested that BCL2L1 was a direct target gene of
miR-663b.
Subsequently, the expression of BCL2L1 in
hypoxia-induced H9c2 cells was determined. RT-qPCR and western
blotting demonstrated that BCL2L1 expression was downregulated in
hypoxia-induced H9c2 cells (Fig. 2C
and D).
Effect of miR-663b inhibitor on H9c2
cell injury induced by hypoxia
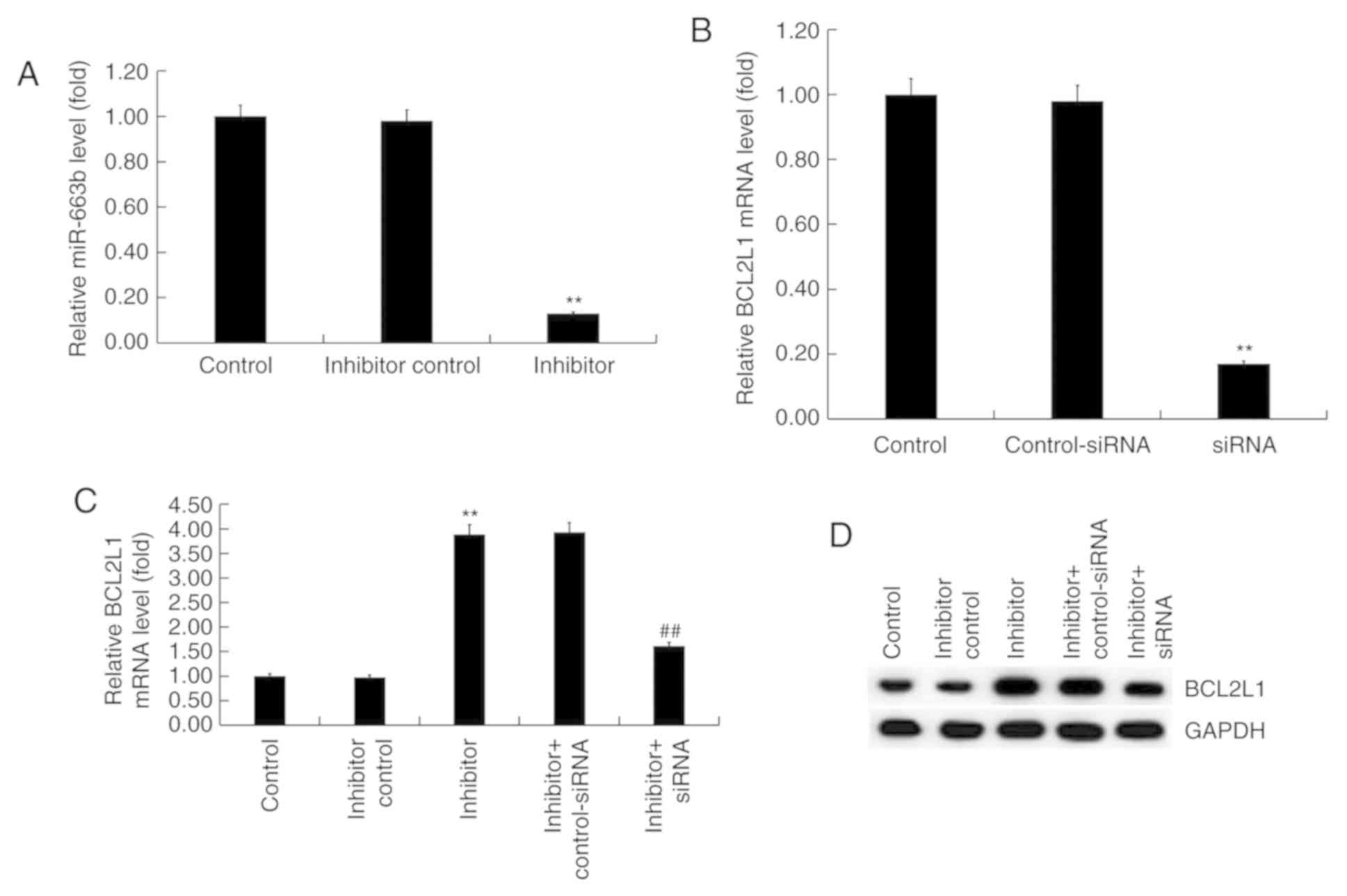
H9c2 cells were transfected with inhibitor control
or miR-663b inhibitor for 48 h. RT-qPCR revealed that the miR-663b
inhibitor significantly reduced miR-663 expression in H9c2 cells
compared with controls (Fig. 3A).
Additionally, H9c2 cells were transfected with BCL2L1-siRNA or
control-siRNA. RT-qPCR results revealed that BCL2L1-siRNA
significantly decreased the expression of BCL2L1 at the mRNA level
in H9c2 cells compared with controls (Fig. 3B). Subsequently, H9c2 cells were
co-transfected with miR-663b inhibitor + control-siRNA, or miR-663b
inhibitor + BCL2L1-siRNA. RT-qPCR and western blotting results
revealed that miR-663b inhibitor significantly increased the
expression of BCL2L1 at the mRNA level and that this increase was
reversed by BCL2L1-siRNA (Fig. 3C).
Similar results were observed at the protein level as determined
via western blotting (Fig. 3D).
The effect of miR-663b on hypoxia-induced cell
damage was determined by detecting the release of two biomarkers
associated with cardiomyocyte injury, CK-MB and cTnI. Compared with
the controls, hypoxia significantly increased the release of CK-MB
and cTnI whereas the miR-663b inhibitor significantly decreased
CK-MB (Fig. 3E) and cTnI release
(Fig. 3F). Transfection of
BCL2L1-siRNA abolished the aforementioned changes. Furthermore,
hypoxia also significantly reduced mitochondrial viability while
miR-663b inhibitor improved mitochondrial viability in H9c2 cells
(Fig. 3G). These changes were
reversed by transfection of BCL2L1-siRNA.
Effect of miR-663b inhibitor on cell
growth of hypoxia-induced H9c2 cells
Under hypoxic conditions, H9c2 cells were
transfected with inhibitor control, miR-663b inhibitor, miR-663b
inhibitor + control-siRNA or miR-663b inhibitor + BCL2L1-siRNA for
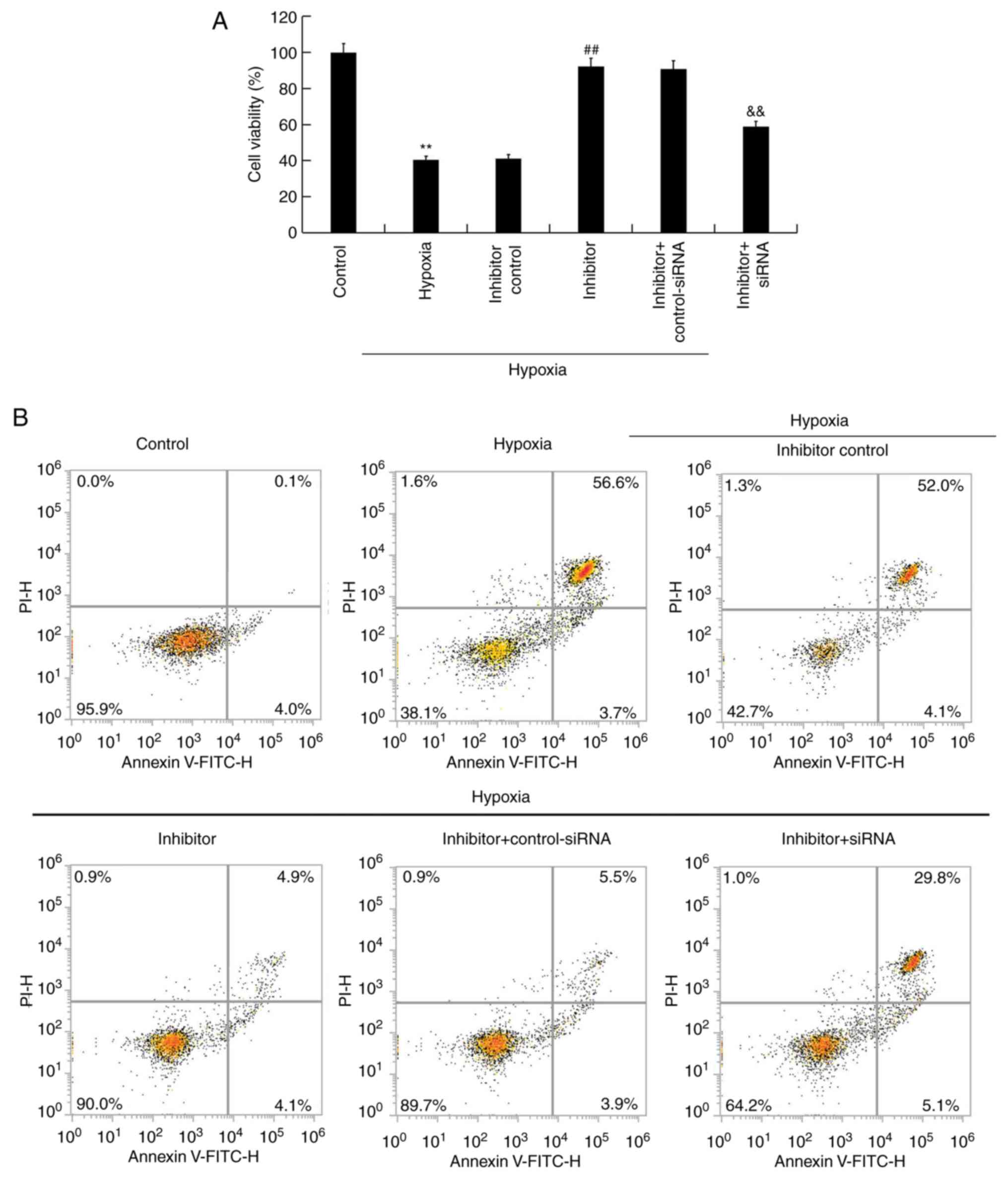
48 h, after which an MTT assay was performed to measure cell
viability. The results revealed that hypoxia significantly
decreased the viability of H9c2 cells, whereas miR-663b inhibitor
increased cell viability. This change was reversed by BCL2L1-siRNA
when compared with the controls (Fig.
4A). Flow cytometry was subsequently performed to detect the
proportion of apoptotic cells. The results revealed that hypoxia
induced cell apoptosis and that the miR-663b inhibitor decreased
cell apoptosis in the hypoxic cells compared with controls. These
changes were reversed following BCL2L1-siRNA transfection (Fig. 4B and C).
The protein expression of cleaved-Caspase-3 was
determined via western blotting. The results indicated that the
hypoxia-induced upregulation of cleaved-Caspase-3 protein
expression in H9c2 cells was significantly reduced in cells
transfected with miR-663b inhibitor. This reduction was reversed
following transfection with BCL2L1-siRNA (Fig. 4D). Together, the results indicated
that the miR-663b inhibitor increased the viability of
hypoxia-induced H9c2 cells and decreased cell apoptosis.
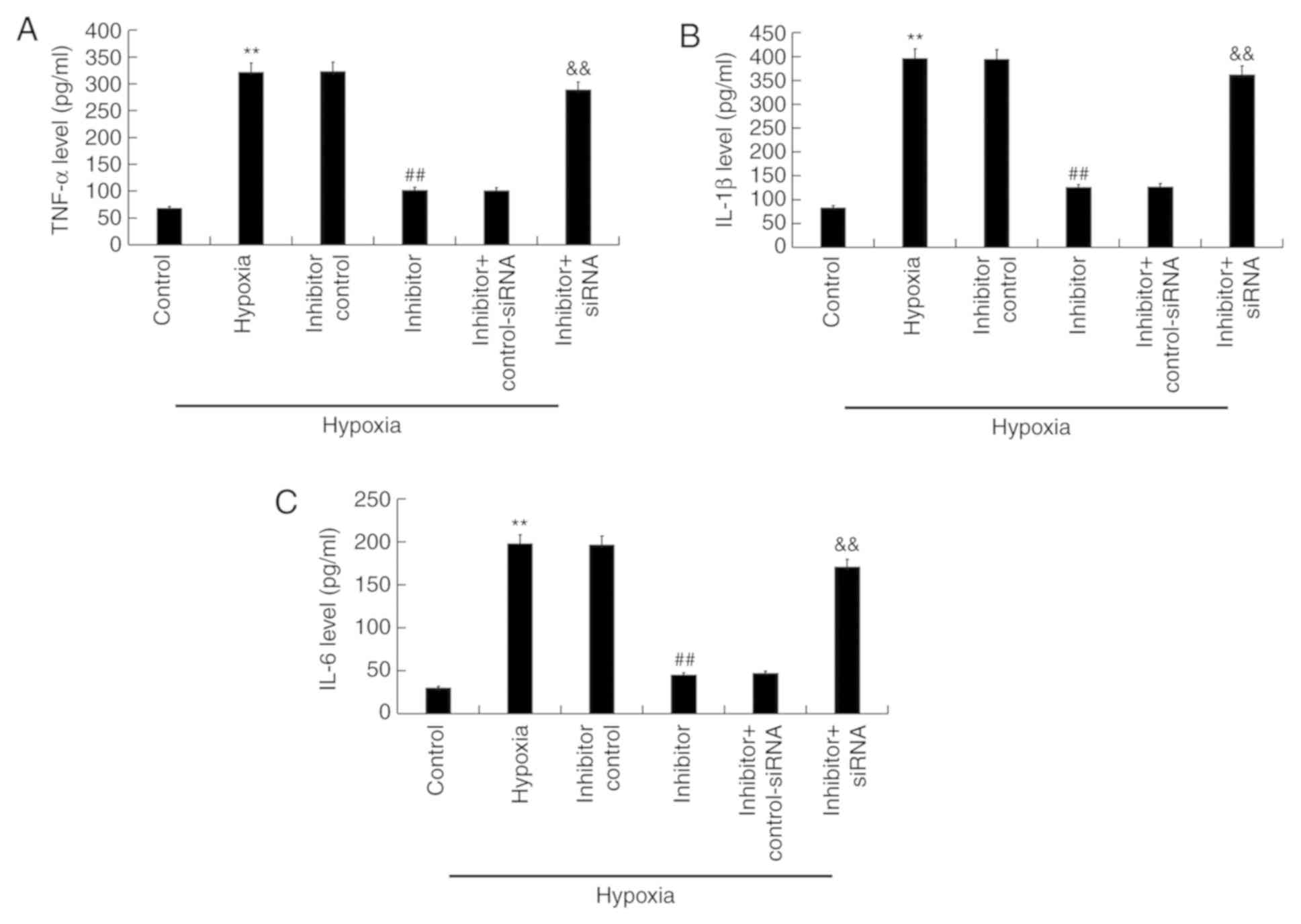
Effect of miR-663b inhibitor on the
secretion of inflammatory factors in hypoxia-induced H9c2
cells
ELISA was performed to detect the effects of
miR-663b inhibitor on the secretion of certain inflammatory factors
in hypoxia-induced H9c2 cells. Compared with the control group,
hypoxia increased the release of TNF-α, IL-1β and IL-6 while
miR-663b inhibitor transfection significantly decreased their
release (Fig. 5A-C). Knocking down
BCL2L1 expression using siRNA reversed these effects.
Discussion
Acute MI is characterized by ischemic injury and
cardiomyocyte cell death, which leads to cardiac dysfunction and
eventual heart failure (3).
Cardiomyocyte apoptosis is one of the primary types of cardiac cell
death during MI (23). Protecting
cardiomyocytes against hypoxia-induced damage may therefore be a
promising therapeutic target for the treatment of MI (24-26).
miRNAs serve an important role in the development of
MI. Xiao et al (27)
demonstrated that miR-24-3p decreased apoptosis in mouse
cardiomyocytes in response to ischemia/reperfusion (I/R) injury.
Zhang et al (2) also revealed
that miR-103a-3p serves a protective role in myocardial I/R injury.
Liu et al (28) demonstrated
that miR-199a-5p expression affects hypoxia/reoxygenation-induced
cytotoxicity in cardiomyocytes. Additionally, Peng et al
(20) demonstrated that miR-133,
miR-1291 and miR-663b expression was significantly increased in AMI
compared with controls. However, the role and function of miR-663b
in MI is yet to be fully elucidated. A previous study indicated
that miR-663b is a tumor-associated miRNA (29). Cai et al (30) also demonstrated that miR-663b exerts
a tumour suppressive effect by targeting insulin-like growth factor
in pancreatic cancer. It has been reported that miR-663b expression
was upregulated in osteosarcoma (31). In the present study, it was
demonstrated that the expression of miR-663b was upregulated in
hypoxia-induced H9c2 cells.
BCL2L1 was determined to be a direct target gene of
miR-663b. Bcl2 genes are a mammalian family of genes that encode
several proteins that directly regulate the activation of
caspase-mediated apoptosis (32). As
a member of this family, BCL2L1 acts as an antiapoptotic protein,
with decreased expression activating a caspase cascade that
ultimately promotes apoptosis. However, increased expression of
BCL2L1 inhibits cell apoptosis (33). Sun and Pei (34) revealed that BCL2L1 is a target of
miR-66 and increases vulnerability to propofol treatment in
developing astrocytes. Lin et al (35) indicated that BCL2L1 was a target gene
of miR-184 and that the miR-184 antagomir promoted apoptosis of
osteosarcoma cells.
In the present study, BCL2L1 expression was
downregulated at the mRNA and protein level in hypoxia-induced H9c2
cells. The effects of miR-663b on cell injury, viability, apoptosis
and the release of inflammatory factors in H9c2 cells were also
determined. Under hypoxic conditions, H9c2 cells were transfected
with miR-663b inhibitor, inhibitor control, miR-663b inhibitor +
control-siRNA or miR-663b inhibitor + BCL2L1-siRNA for 48 h. The
miR-663b inhibitor decreased the release of cardiomyocyte injury
biomarkers CK-MB and cTnI and increased mitochondrial viability and
cell viability while decreasing cell apoptosis and the secretion of
the inflammatory factors IL-1β, TNF-α and IL-6. These effects were
reversed by BCL2L1 knockdown.
In conclusion, the miR-663b inhibitor protected
cardiomyocytes against hypoxia-induced damage by targeting BCL2L1.
These data suggested that miR-663b may be a novel target for the
treatment of MI, exerting its effects by directly targeting
BCL2L1.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
FY and XZ designed the study, collected the data,
performed statistical analysis and data interpretation, and wrote
the manuscript. CS (qRT-PCR, MTT data collection), WX (flow
cytometry analysis and ELISA data collection) and JX (ELISA and MTT
data collection) collected the data and prepared the manuscript.
All authors read and approved the final version of this
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Henriksson R, Björklund F and Mooe T: The
introduction of ticagrelor is associated with lower rates of
recurrent ischemic stroke after myocardial infarction. PLoS One.
14(e0216404)2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Zhang C, Zhang C, Wang H, Qi Y, Kan Y and
Ge Z: Effects of miR-103a-3p on the autophagy and apoptosis of
cardiomyocytes by regulating Atg5. Int J Mol Med. 43:1951–1960.
2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Zeymer U: Diagnosis and initial management
of acute myocardial infarction. MMW Fortschr Med. 161:34–36.
2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Michiels C: Physiological and pathological
responses to hypoxia. Am J Pathol. 164:1875–1882. 2004.PubMed/NCBI View Article : Google Scholar : (In German).
|
|
5
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Poy MN, Eliasson L, Krutzfeldt J, Kuwajima
S, Ma X, Macdonald PE, Pfeffer S, Tuschl T, Rajewsky N, Rorsman P
and Stoffel M: A pancreatic islet-specifc microRNA regulates
insulin secretion. Nature. 432:226–30. 2004.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Wang Q, Liu N, Yang X, Tu L and Zhang X:
Small RNA-mediated responses to low- and high-temperature stresses
in cotton. Sci Rep. 6(35558)2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lim LP, Lau NC, Garrett-Engele P, Grimson
A, Schelter JM, Castle J, Bartel DP, Linsley PS and Johnson JM:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wang Y, Chen F, Zhao M, Yang Z, Zhang S,
Ye L, Gao H and Zhang X: MiR-107 suppresses proliferation of
hepatoma cells through targeting HMGA2 mRNA 3'UTR. Biochem Biophys
Res Commun. 480:455–460. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhu Q, Gong L, Wang J, Tu Q, Yao L, Zhang
JR, Han XJ, Zhu SJ, Wang SM, Li YH and Zhang W: miR-10b exerts
oncogenic activity in human hepatocellular carcinoma cells by
targeting expression of CUB and sushi multiple domains 1 (CSMD1).
BMC Cancer. 16(806)2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006.PubMed/NCBI View
Article : Google Scholar
|
|
14
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Liang S, Zhang N, Deng Y, Chen L, Zhang Y,
Zheng Z, Luo W, Lv Z, Li S and Xun T: Increased serum level of
MicroRNA-663 is correlated with poor prognosis of patients with
nasopharyngeal carcinoma. Dis Markers. 2016(7648215)2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Pan J, Hu H, Zhou Z, Sun L, Peng L, Yu L,
Sun L, Liu J, Yang Z and Ran Y: Tumor-suppressive mir-663 gene
induces mitotic catastrophe growth arrest in human gastric cancer
cells. Oncol Rep. 24:105–112. 2010.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Jiao L, Deng Z, Xu C, Yu Y, Li Y, Yang C,
Chen J, Liu Z, Huang G, Li LC and Sun Y: miR-663 induces
castration-resistant prostate cancer transformation and predicts
clinical recurrence. J Cell Physiol. 229:834–844. 2014.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yi C, Wang Q, Wang L, Huang Y, Li L, Liu
L, Zhou X, Xie G, Kang T, Wang H, et al: MiR-663, a microRNA
targeting p21 (WAF1/CIP1), promotes the proliferation and
tumorigenesis of nasopharyngeal carcinoma. Oncogene. 31:4421–4433.
2012.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Liang S, Zhang N, Deng Y, Chen L, Zhang Y,
Zheng Z, Luo W, Lv Z, Li S and Xu T: miR-663b promotes tumor cell
proliferation, migration and invasion in nasopharyngeal carcinoma
through targeting TUSC2. Exp Ther Med. 14:1095–1103.
2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Peng L, Chun-guang Q, Bei-fang L, Xue-zhi
D, Zi-hao W, Yun-fu L, Yan-ping D, Yang-gui L, Wei-guo L, Tian-yong
H and Zhen-wen H: Clinical impact of circulating miR-133, miR-1291
and miR-663b in plasma of patients with acute myocardial
infarction. Diagn Pathol. 9(89)2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Gong L, Chang H, Zhang J, Guo G, Shi J and
Xu H: Astragaloside IV protects rat cardiomyocytes from
hypoxia-Induced injury by down-regulation of miR-23a and miR-92a.
Cell Physiol Biochem. 49:2240–2253. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta c(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Thygesen K, Alpert JS, Jaffe AS, Simoons
ML, Chaitman BR and White HD: Writing Group on the Joint
ESC/ACCF/AHA/WHF Task Force for the Universal Definition of
Myocardial Infarction. Thygesen K, Alpert JS, White HD, et alThird
universal definition of myocardial infarction. Eur Heart J.
33:2551–2567. 2012.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Chen Y, Zhao Y, Chen W, Xie L, Zhao ZA,
Yang J, Chen Y, Lei W and Shen Z: MicroRNA-133 overexpression
promotes the therapeutic efficacy of mesenchymalstem cells on acute
myocardial infarction. Stem Cell Res Ther. 8(268)2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Hu G, Ma L, Dong F, Hu X, Liu S and Sun H:
Inhibition of microRNA-124-3p protects against acute myocardial
infarction by suppressing the apoptosis of cardiomyocytes. Mol Med
Rep. 20:3379–3387. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Luo Q, Guo D, Liu G, Chen G, Hang M and
Jin M: Exosomes from MiR-126-overexpressing adscs are therapeutic
in relieving acute myocardial ischaemic injury. Cell Physiol
Biochem. 44:2105–2116. 2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Xiao X, Lu Z, Lin V, May A, Shaw DH, Wang
Z, Che B, Tran K, Du H and Shaw PX: MicroRNA miR-24-3p reduces
apoptosis and regulates Keap1-Nrf2 pathway in mouse cardiomyocytes
responding to ischemia/reperfusion injury. Oxid Med Cell Longev.
2018(7042105)2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Liu DW, Zhang YN, Hu HJ, Zhang PQ and Cui
W: Downregulation of microRNA-199a-5p attenuates
hypoxia/reoxygenation-induced cytotoxicity in cardiomyocytes by
targeting the HIF-1α-GSK3β-mPTP axis. Mol Med Rep. 19:5335–5344.
2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Wang M, Jia M and Yuan K: MicroRNA-663b
promotes cell proliferation and epithelial mesenchymal transition
by directly targeting SMAD7 in nasopharyngeal carcinoma. Exp Ther
Med. 16:3129–3134. 2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Cai H, An Y, Chen X, Sun D, Chen T, Peng
Y, Zhu F, Jiang Y and He X: Epigenetic inhibition of miR-663b by
long non-coding RNA HOTAIR promotes pancreatic cancer cell
proliferation via up-regulation of insulin-like growth factor 2.
Oncotarget. 7:86857–86870. 2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhao H, Li M, Li L, Yang X, Lan G and
Zhang Y: MiR-133b is down-regulated inhuman osteosarcomaand
inhibits osteosarcoma cells proliferation, migrationand invasion,
and promotes apoptosis. PLoS One. 8(e83571)2013.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Fagundes DJ, Carrara FL, Teixeira WA,
Simões RS and Taha MO: The role of the exogenous supply of
adenosine triphosphate in the expression of Bax and Bcl2L1 genes in
intestinal ischemia and reperfusion in rats1. Acta Cir Bras.
33:889–895. 2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ureshino RP, Bertoncini CR, Fernandes MJ,
Abdalla FM, Porto CS, Hsu YT, Lopes GS and Smaili SS: Alteratons in
calcium signaling and a decrease in Bcl-2 expression: Possible
correlaton with apoptosis in aged striatum. J Neurosci Res.
88:438–447. 2010.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Sun WC and Pei L: rno-miR-665 targets
BCL2L1 (Bcl-xl) and increases vulnerability to propofol in
developing astrocytes. J Neurochem. 138:233–242. 2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Lin BC, Huang D, Yu CQ, Mou Y, Liu YH,
Zhang DW and Shi FJ: MicroRNA-184 modulates doxorubicin resistance
in osteosarcoma cells by targeting BCL2L1. Med Sci Monit.
22:1761–1765. 2016.PubMed/NCBI View Article : Google Scholar
|