Introduction
Gastric cancer is one of the primary causes of
cancer-associated mortality worldwide and is responsible for over
700,000 fatalities per year (1). It
is the second most common type of cancer and the third leading
cause of fatality amongst patients with cancer in China (2). The gastric cancer incidence in China
and Japan account for more than 40% of the worldwide occurrences
(3).
A wide range of cytokines, chemokines and growth
factors, as well as the extracellular matrix can affect the
carcinogenesis and progression of gastric cancer (4). Recently, it has been suggested that the
interaction between cancer cells and the surrounding tumor
microenvironment serves a pivotal role during tumor progression
(5). As one of the cytokines
secreted by T helper 17 cells in the tumor microenvironment,
interleukin (IL)-22 is a cytokine that structurally associated with
IL-10 and produced predominantly by activated lymphocytes in
chronically inflamed tissues (6).
IL-22 exerts its biological actions via the IL-22 receptor (IL-22R)
(7). IL-22R is a heterodimeric
receptor consisting of two chains: IL-22R1 and IL-10R2. IL-10R2 is
ubiquitously expressed in various organs, whereas IL-22R1 is
restricted to epithelial cells in the skin, pancreas, kidney, liver
and gastrointestinal tract (8). It
has been reported that the expression of IL-22 is elevated in
several types of gastrointestinal cancer (9,10) and
that increased IL-22 expression is associated with cancer
development (8).
JNK, a member of the mitogen-activated protein
kinase (MAPK) family, can respond to a variety of environmental
stresses, including cytokines, ultraviolet irradiation and heat
shock, and has been implicated in multiple cellular events,
including apoptosis and autophagy (11). There are three JNK genes, namely
JNK1, JNK2 and JNK3, which encode 2-4 JNK isoforms (12). JNK1 has been revealed to be involved
in apoptosis, neurodegeneration, cell differentiation and
proliferation, as well as inflammatory conditions (13-16).
It can also regulate several important cellular functions,
including cell growth, differentiation, survival and apoptosis
(17,18). It has been documented that IL-22 can
trigger the nuclear factor-κB, MAPK and PI3K/Akt/mTOR signaling
pathways (19). IL-22-mediated
signaling enhances the expression of genes with anti-inflammatory,
mitogenic, proliferative and anti-apoptotic effects, which are
cellular effects that promote local tissue regeneration and host
defense (20).
The aim of the present study was to analyze the role
of IL-22 in gastric cancer cell progression and explore its
underlying molecular mechanism. The effects of IL-22-plasmid and
IL-22-short hairpin (sh)RNA on the viability of gastric cancer
cells were therefore investigated.
Materials and methods
Cell culture
The gastric cancer cell line AGS and the human
normal gastric epithelial cell line GES-1 were obtained from the
American Type Culture Collection. GES-1 cells were maintained in
Dulbecco's modified Eagle's medium (DMEM; Hyclone; GE Healthcare
Life Sciences). AGS cells were maintained in Roswell Park Memorial
Institute 1640 (Gibco; Thermo Fisher Scientific, Inc.). All media
were supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin (Sigma-Aldrich; Merck KGaA)
and 100 µg/ml streptomycin (Sigma-Aldrich; Merck KGaA). Cell lines
were cultured at 37˚C in a humidified incubator with 5%
CO2.
Cell transfection
IL-22-shRNA, control shRNA and IL-22-plasmids
control plasmid were designed and constructed by Genechem
Corporation. IL-22-shRNA, IL-22-plasmid and the corresponding
control were transfected into AGS cells using Lipofectamine™ 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. 1 µl shRNA or 0.5 µg plasmid was
diluted with 50 µl serum-free Opti-MEM (Gibco; Thermo Fisher
Scientific, Inc.) respectively, gently mixed and incubated at room
temperature for 5 min. 1 µl Lipofectamine™ 2000 reagent
(Invitrogen; ThermoFisher Scientific Inc.) was diluted with 50 µl
serum-free Opti-MEM, lightly mixed and incubated at room
temperature for 5 min. Then, the above mixtures were mixed again
gently and incubated at room temperature for 20 min. The mixture of
shRNA-Lipofectamine™ 2000 or plasmid-Lipofectamine™ 2000 was added
into 400 µl medium and incubated for 6 h. The medium was replaced
with fresh medium at 37˚C for 48 h. Untreated cells served as the
control group. Following incubation for 48 h, the cells were
subjected to subsequent experiments. Transfection efficiency was
determined using reverse transcription-quantitative PCR
(RT-qPCR).
Western blot analysis
The anti-ERK monoclonal antibody (mab; MAB1230;
1:1,000), anti-phosphorylated (p)-ERK mab (MAB18251; 1:1,000),
anti-JNK mab (AF1387; 1:2,000), anti-p-JNK mab (AF1205; 1:500),
anti-STAT3 mab (MAB1799; 1:1,000), anti-p-STAT3 mab (AF4607;
1:500), anti-Bcl-2 mab (AF810; 1:1,000), anti-GAPDH mab (MAB5718;
1:2,000) and mouse or rabbit horseradish peroxidase
(HRP)-conjugated secondary antibodies (HAF007, HAF008; 1:1,000)
were purchased from R&D Systems, Inc.
Cells were lysed in radioimmunoprecipitation assay
buffer containing phenylmethane sulfonyl fluoride and protease
inhibitors (Beyotime Institute of Biotechnology). The protein
concentration of each sample was determined using a BCA protein
assay kit (Beyotime Institute of Biotechnology). Following this,
30-50 µg of whole-cell protein from each sample was loaded and
separated using 10% SDS-PAGE and then transferred to polyvinylidene
difluoride membranes (EMD Millipore). The membranes were blocked
with 5% non-fat milk or bovine serum albumin at room temperature
for 1 h, and incubated with primary antibodies as mentioned above
overnight at 4˚C, according to the manufacturer's instructions,
followed by HRP-conjugated secondary antibody at room temperature
for 2 h. An enhanced chemiluminescence detection system (GE
Healthcare) was used for the visualization of the target proteins.
GAPDH was used as the loading control. Relative quantification of
protein expression was conducted using Image-Pro Plus 6.0 (Media
Cybernetics, Inc.).
Cell viability assay
Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc.) was used to assess cell viability. Cells were
seeded onto 96-well culture plates (2x103 cell/well),
then transfected with IL-22-plasmid, IL-22-shRNA and corresponding
negative control as described above. Cells were cultured for a
further 24, 48, 72 h, allowing for transcription, translation and
secretion. Subsequently, CCK-8 was added to the culture media. The
optical density value of each well was measured at a wavelength of
450 nm using a plate reader (Infinite 200 PRO NanoQuant; Tecan
Austria GmbH).
RT-qPCR
Total RNA was extracted from the cells using the
TRIzol Plus RNA Purification kit (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions. A
reaction mixture (20 µl) containing total RNA (1 µg) was
transcribed to cDNA at 37˚C for 15 min and 85˚C for 5 sec using
PrimeScript RT-polymerase (GE Healthcare Life Sciences).
qPCR was performed using the SYBR Green qPCR kit
(Takara Bio, Inc.), and the reaction system was established
according to the manufacturer's instructions. qPCR was performed at
95˚C for an initial 10 min followed by 40 cycles of denaturation
for 30 sec at 95˚C, annealing at 60˚C for 30 sec and extension at
72˚C for 30 sec. GAPDH was used as the internal control. All
reactions were performed on the CFX96 Touch™ System (Bio-Rad
Laboratories, Inc.). Relative expression levels were calculated
using the 2-ΔΔCq method (21). The following primers were used: GAPDH
forward, 5'-ATTGATGGATGCTAFGAGTATT-3' and reverse,
5'-AGTCTTCTGGGTGGCAGTGAT-3'; and IL-22 forward
5'-GCTAAGGAGGCTAGCTTG-3' and reverse, 5'-CAGCAAATCCAGTTCTCC-3'.
Statistical analysis
Statistical analysis was performed using SPSS 13.0
software (SPSS, Inc.). Experimental values are expressed as the
mean ± SEM of at least three independent experiments performed in
triplicate. Data were analyzed using one-way analysis of variance
followed by Dunnett's post hoc test for multiple comparisons or a
Student's t-test for comparisons between two groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
IL-22 is overexpressed in the AGS cell
line
In order to explore the role of IL-22 in gastric
cancer, the expression level of IL-22 in GES-1 and AGS cells was
compared. As depicted in Fig. 1, the
expression of IL-22 was significantly higher in AGS cells compared
with GES-1 cells at the protein and mRNA levels. Therefore, AGS was
chosen as the target cell line in the present study and subjected
to further analysis.
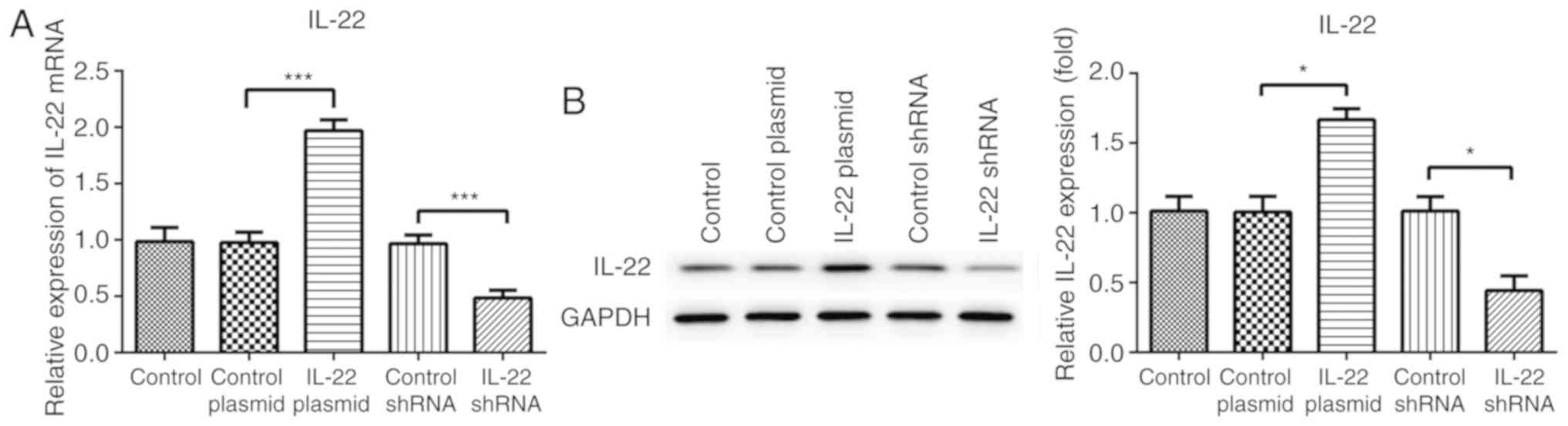
IL-22 promotes AGS cell viability
To investigate the effect of IL-22 on cell
viability, AGS cells were transfected with control plasmid, IL-22
plasmid, control shRNA or IL-22 shRNA. The expression level of
IL-22 in each treatment group was detected using western blot
analysis and RT-qPCR. As indicated in Fig. 2, significantly higher IL-22
expression and an enhanced transcriptional level of IL-22 mRNA was
observed in the IL-22 plasmid treatment group compared with the
control plasmid treatment group. Furthermore, shRNA IL-22 reduced
the translation of IL-22 in AGS cells, as the mRNA and protein
expression levels of IL-22 were knocked down significantly compared
with those in the control shRNA group.
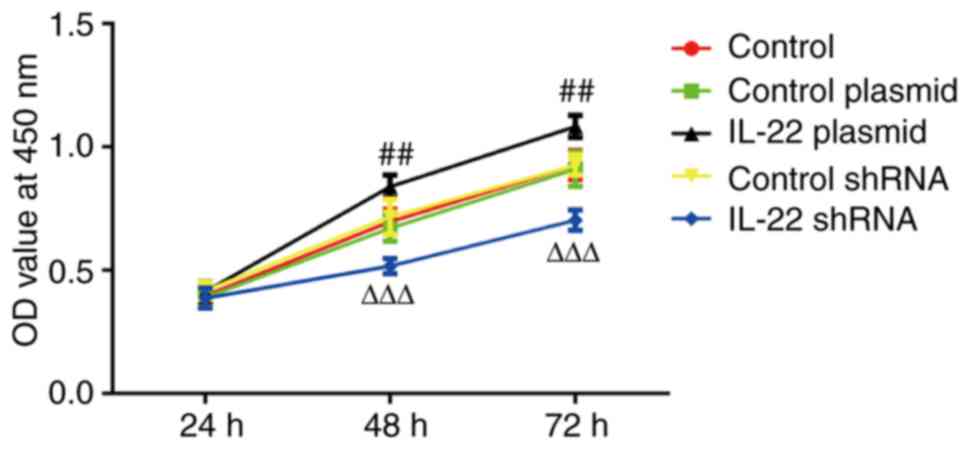
The viability of AGS cells at different time points
following transfection for 48 h was examined using the CCK-8 assay.
As indicated in Fig. 3, the
overexpression of IL-22 significantly increased AGS cell viability
compared with that in the control plasmid group. In addition, IL-22
knockdown significantly decreased the viability of AGS cells when
compared with the control shRNA group.
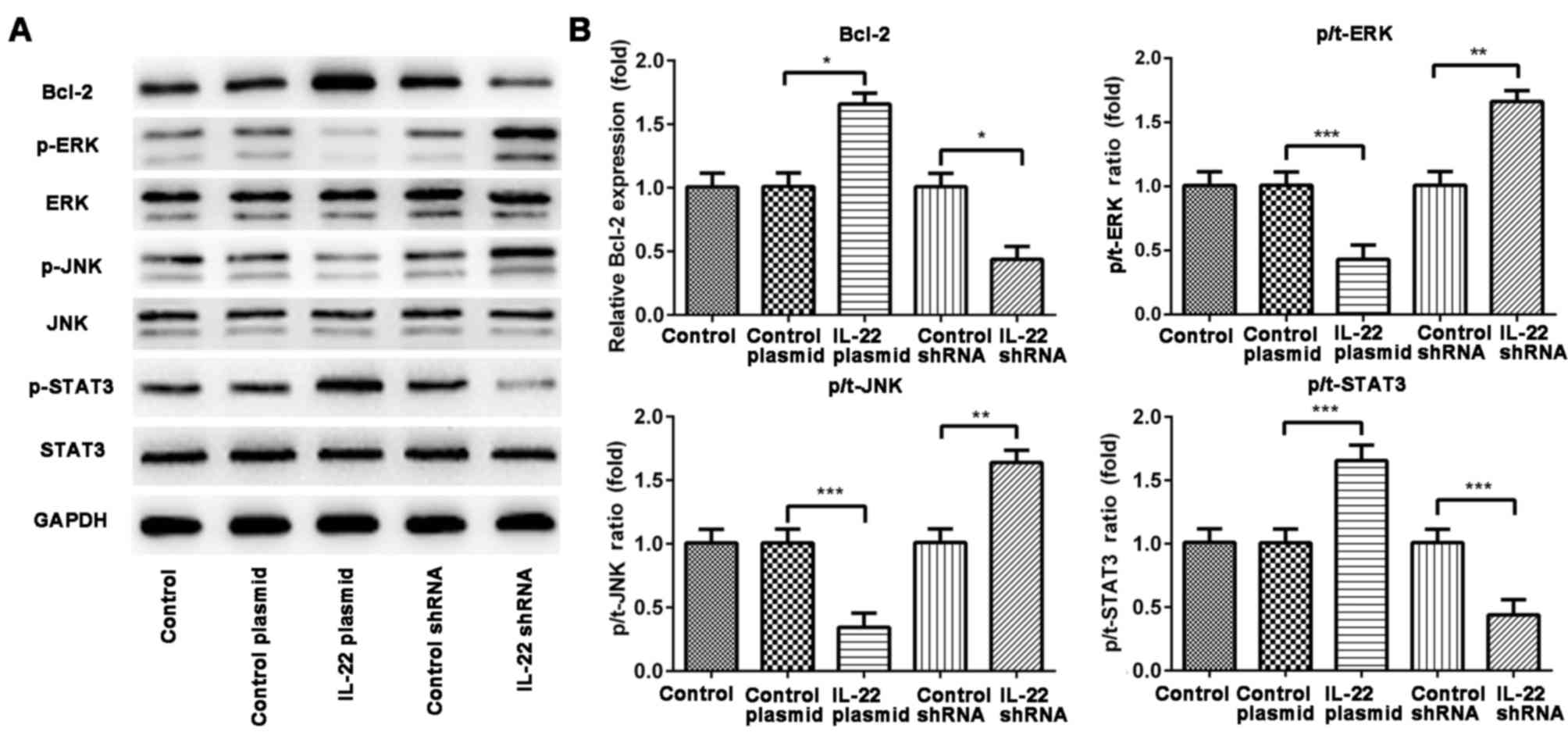
MAPK signaling pathway is involved in
the effects of IL-22 on AGS cells
To investigate the molecular mechanism underlying
the effects of IL-22, activation of the MAPK signaling pathway in
AGS cells was investigated following transfection with IL-22
plasmid and IL-22 shRNA (Fig. 4).
Western blot analysis revealed that the IL-22 plasmid decreased the
phosphorylation of ERK and JNK, increased the phosphorylation of
p-STAT3 and increased the expression of Bcl-2, when compared with
the control plasmid. By contrast, transfection with IL-22 shRNA
increased the phosphorylation of ERK and JNK, decreased the
phosphorylation of STAT3 and decreased the expression of Bcl-2 when
compared with the control shRNA.
Discussion
The incidence of gastric cancer is gradually
declining in developed countries; however, it remains one of the
most common types of cancer worldwide (22). Although the overall survival of
patients with gastric cancer has been prolonged in the last
decades, due to an increase in early detection and a wider use of
radical surgery (23), the prognosis
of this advanced type of cancer remains poor as safe and effective
treatment options are limited (24).
It is urgently necessary to explore the molecular mechanism of
gastric cancer in order to aid the progress of clinical practice
and drug development.
While it is clear that various cytokines are
associated with the proliferation of human cancer cells, the
mechanisms of cytokine action remain largely unknown. IL-22 and
other members of the IL-20 subfamily (IL-20, IL-24 and IL-26)
belong to the larger IL-10 family of cytokines that have important
roles in tissue remodeling and wound healing (25). Previous studies have demonstrated
that IL-22 is released by infiltrating immune cells and by the
tumor microenvironment, thereby promoting the progression of tumor
cells. As these functions are also central to several types of
cancer, it is not surprising that IL-22 has been implicated in
cancer development and progression (26-28).
Increased numbers of blood-circulating and intratumoral
IL-22-secreting cells have been reported in lung, gastric,
colorectal, pancreatic and hepatocellular carcinomas (20). Although IL-22 has been well
documented in the aforementioned cancer types, the underlying
mechanism of IL-22 regulation of gastric cancer cell is not clear.
In the present study, the pro-survival effect of IL-22 and its
underlying molecular mechanism of action in AGS cells were revealed
for the first time. It was identified that IL-22 promoted AGS cell
proliferation.
The signaling pathway underlying the effect of IL-22
on AGS cells was also investigated in the present study. IL-22 has
various functions in multiple cell types by activating a wide range
of signaling cascades, such as the Janus kinase (JAK)/STAT, ERK,
JNK, p38 and PI3K/Akt/mTOR pathways (29-32).
Several important signaling pathways can be regulated downstream of
IL-22R1 activation. For instance, IL-22 can stimulate the PI3K/AKT
pathway in epidermal keratinocytes (31). It can also activate the JAK/STAT
signaling pathway in human hepatocellular carcinoma cells (33) and ERK1/2, p38 and JNK in H4IIE cells
(29). Furthermore, IL-22 induces
matrix metalloproteinase-9 production via activation of IL-22R1 and
AKT, further confirming the involvement of IL-22R1/AKT signaling in
the IL-22-mediated migration and invasion of gastric cancer cells
(34). JNKs have a critical role in
death receptor-initiated extrinsic and mitochondrial intrinsic
apoptotic pathways, and can activate apoptotic signaling through
the upregulation of pro-apoptotic genes via the transactivation of
specific transcription factors upon stimulation (18). Therefore, it may be hypothesized that
IL-22 could protect AGS cells from apoptosis through the JNK
signaling pathway. In the present study, western blot analysis and
RT-qPCR demonstrated that IL-22 downregulated the phosphorylation
of ERK and JNK simultaneously, which indicated multiple inhibition
of MAPK activity in AGS cells. The activated MAPK pathway has been
detected in many types of cancer, including breast, prostate, lung
and gastric cancer (35). In
addition, the present findings showed that IL-22 induced a robust
activation of STAT3. The expression of Bcl-2, an anti-apoptotic
protein, was also increased in AGS cells transfected with
IL-22-plasmid, which may indicate a potential anti-apoptotic effect
of IL-22, in accordance with a previous study (36). However, further experiments such as
flow cytometry and TUNEL staining are necessary to verify the
anti-apoptotic effect of IL-22, and whether alternation of JNK
activity affects the role of IL-22 in cell apoptosis is worth
exploring.
In conclusion, the present study demonstrated that
IL-22 acts as an endogenous resistance factor in human gastric
cancer cell lines. By modulating the IL-22 expression levels with
IL-22 plasmid or IL-22 shRNA, it was demonstrated that IL-22
enhanced the viability of AGS cells, which may involve the JNK
signaling pathway.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HD designed the research. HD and FZ performed the
experiments. SJ and JT analyzed the data. FZ drafted the manuscript
and analyzed data. SJ and JT collected and interpreted data and
revised the final manuscript. HD wrote the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108.
2005.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Larsson SC, Orsini N and Wolk A: Processed
meat consumption and stomach cancer risk: A meta-analysis. J Natl
Cancer Inst. 98:1078–1087. 2006.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Cirri P and Chiarugi P:
Cancer-associated-fibroblasts and tumour cells: A diabolic liaison
driving cancer progression. Cancer Metastasis Re. 31:195–208.
2012.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Xing F, Saidou J and Watabe K: Cancer
associated fibroblasts (CAFs) in tumor microenvironment. Front
Biosci (Landmark Ed). 15:166–179. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
6
|
Brockmann L, Giannou AD, Gagliani N and
Huber S: Regulation of TH17 cells and associated
cytokines in wound healing, tissue regeneration, and
carcinogenesis. Int J Mol Sci. 18(pii: E1033)2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Vivier E, Spits H and Cupedo T:
Interleukin-22-producing innate immune cells: New players in
mucosal immunity and tissue repair? Nat Rev Immunol. 9:229–234.
2009.PubMed/NCBI View
Article : Google Scholar
|
|
8
|
Jiang R, Wang H, Deng L, Hou J, Shi R, Yao
M, Gao Y, Yao A, Wang X, Yu L and Sun B: IL-22 is related to
development of human colon cancer by activation of STAT3. BMC
Cancer. 13(59)2013.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Xu X, Tang Y, Guo S, Zhang Y, Tian Y, Ni B
and Wang H: Increased intratumoral interleukin 22 levels and
frequencies of interleukin 22-producing CD4+ T cells correlate with
pancreatic cancer progression. Pancreas. 43:470–477.
2014.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Petanidis S, Anestakis D, Argyraki M,
Hadzopoulou-Cladaras M and Salifoglou A: Differential expression of
IL-17, 22 and 23 in the progression of colorectal cancer in
patients with K-ras mutation: Ras signal inhibition and crosstalk
with GM-CSF and IFN-γ. PLoS One. 8(e73616)2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Kyriakis JM, Banerjee P, Nikolakaki E, Dai
T, Rubie EA, Ahmad MF, Avruch J and Woodgett JR: The
stress-activated protein kinase subfamily of c-Jun kinases. Nature.
369:156–160. 1994.PubMed/NCBI View
Article : Google Scholar
|
|
12
|
Auladell C, de Lemos L, Verdaguer E,
Ettcheto M, Busquets O, Lazarowski A, Beas-Zarate C, Olloquequi J,
Folch J and Camins A: Role of JNK isoforms in the kainic acid
experimental model of epilepsy and neurodegeneration. Front Biosci
(Landmark Ed). 22:795–814. 2017.PubMed/NCBI View
Article : Google Scholar
|
|
13
|
Sabapathy K, Jochum W, Hochedlinger K,
Chang L, Karin M and Wagner EF: Defective neural tube morphogenesis
and altered apoptosis in the absence of both JNK1 and JNK2. Mech
Dev. 89:115–124. 1999.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ribas VT, Gonçalves BS, Linden R and
Chiarini LB: Activation of c-Jun N-terminal kinase (JNK) during
mitosis in retinal progenitor cells. PLoS One.
7(e34483)2012.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Nanni M, Ranieri D, Rosato B, Torrisi MR
and Belleudi F: Role of fibroblast growth factor receptor 2b in the
cross talk between autophagy and differentiation: Involvement of
Jun N-terminal protein kinase signaling. Mol Cell Biol. 38(pii:
e00119-18)2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Song N, Liu ZS, Xue W, Bai ZF, Wang QY,
Dai J, Liu X, Huang YJ, Cai H, Zhan XY, et al: NLRP3
phosphorylation is an essential priming event for inflammasome
activation. Mol Cell. 68:185–197.e6. 2007.
|
|
17
|
Lawan A and Bennett AM: Mitogen-activated
protein kinase regulation in hepatic metabolism. Trends Endocrinol
Metab. 28:868–878. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Dhanasekaran DN and Reddy EP: JNK
signaling in apoptosis. Oncogene. 27:6245–6251. 2008.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Markota A, Endres S and Kobold S:
Targeting interleukin-22 for cancer therapy. Hum Vaccin Immunother.
14:2012–2015. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Lim C and Savan R: The role of the
IL-22/IL-22R1 axis in cancer. Cytokine Growth Factor Rev.
25:257–271. 2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)). Methods. 25:402–408. 2001.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Neugut AI, Hayek M and Howe G:
Epidemiology of gastric cancer. Semin Oncol. 23:281–291.
1996.PubMed/NCBI
|
|
23
|
Tanizawa Y and Terashima M: Lymph node
dissection in the resection of gastric cancer: Review of existing
evidence. Gastric Cancer. 13:137–148. 2010.PubMed/NCBI View Article : Google Scholar
|
|
24
|
An JY, Ha TK, Noh JH, Sohn TS and Kim S:
Proposal to subclassify stage IV gastric cancer into IVA, IVB, and
IVM. Arch Surg. 144:38–45. 2009.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Rutz S, Wang X and Ouyang W: The IL-20
subfamily of cytokines-from host defence to tissue homeostasis. Nat
Rev Immunol. 14:783–795. 2014.PubMed/NCBI View
Article : Google Scholar
|
|
26
|
Rolla S, Alchera E, Imarisio C, Bardina V,
Valente G, Cappello P, Mombello C, Follenzi A, Novelli F and Carini
R: The balance between IL-17 and IL-22 produced by
liver-infiltrating T-helper cells critically controls NASH
development in mice. Clin Sci (Lond). 130:193–203. 2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Crome SQ, Nguyen LT, Lopez-Verges S, Yang
SY, Martin B, Yam JY, Johnson DJ, Nie J, Pniak M, Yen PH, et al: A
distinct innate lymphoid cell population regulates tumor-associated
T cells. Nat Med. 23:368–375. 2017.PubMed/NCBI View
Article : Google Scholar
|
|
28
|
Khosravi N, Caetano MS, Cumpian AM, Unver
N, De la Garza Ramos C, Noble O, Daliri S, Hernandez BJ, Gutierrez
BA, Evans SE, et al: IL22 Promotes Kras-Mutant lung cancer
by induction of a protumor immune response and protection of
stemness properties. Cancer Immunol Res. 6:788–797. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Lejeune D, Dumoutier L, Constantinescu S,
Kruijer W, Schuringa JJ and Renauld JC: Interleukin-22 (IL-22)
activates the JAK/STAT, ERK, JNK, and p38 MAP kinase pathways in a
rat hepatoma cell line. Pathways that are shared with and distinct
from IL-10. J Biol Chem. 277:33676–33682. 2002.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Andoh A, Zhang Z, Inatomi O, Fujino S,
Deguchi Y, Araki Y, Tsujikawa T, Kitoh K, Kim-Mitsuyama S,
Takayanagi A, et al: Interleukin-22, a member of the IL-10
subfamily, induces inflammatory responses in colonic subepithelial
myofibroblasts. Gastroenterology. 129:969–984. 2005.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Mitra A, Raychaudhuri SK and Raychaudhuri
SP: IL-22 induced cell proliferation is regulated by PI3K/Akt/mTOR
signaling cascade. Cytokine. 60:38–42. 2012.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Ikeuchi H, Kuroiwa T, Hiramatsu N, Kaneko
Y, Hiromura K, Ueki K and Nojima Y: Expression of interleukin-22 in
rheumatoid arthritis: Potential role as a proinflammatory cytokine.
Arthritis Rheum. 52:1037–1046. 2005.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Jiang R, Tan Z, Deng L, Chen Y, Xia Y, Gao
Y, Wang X and Sun B: Interleukin-22 promotes human hepatocellular
carcinoma by activation of STAT3. Hepatology. 54:900–909.
2011.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ji Y, Yang X, Li J, Lu Z, Li X, Yu J and
Li N: IL-22 promotes the migration and invasion of gastric cancer
cells via IL-22R1/AKT/MMP-9 signaling. Int J Clin Exp Pathol.
7:3694–3703. 2014.PubMed/NCBI
|
|
35
|
Wagner EF and Nebreda AR: Signal
integration by JNK and p38 MAPK pathways in cancer development. Nat
Rev Cancer. 9:537–549. 2009.PubMed/NCBI View
Article : Google Scholar
|
|
36
|
Leu CM, Wong FH, Chang C, Huang SF and Hu
C: Interleukin-6 acts as an antiapoptotic factor in human
esophageal carcinoma cells through the activation of both STAT3 and
mitogen-activated protein kinase pathways. Oncogene. 22:7809–7818.
2003.PubMed/NCBI View Article : Google Scholar
|