Introduction
Osteosarcoma (OS) is derived from primitive bone
mesenchymal cells and is the most common malignant bone tumor
(1). OS primarily occurs in children
and young adults between 10-20 years of age and is characterized by
lung metastasis and a poor prognosis, posing a serious threat to
health (2). Multiple factors,
including epigenetic alterations, environmental ionizing radiation
and lesions, have been demonstrated to be associated with OS
formation and progression (3).
However, the complex molecular mechanisms associated with OS
development remain unclear. Currently, surgical resection in
combination with preoperative and postoperative chemotherapy is a
standard treatment for patients with OS (4). Despite years of development in OS
diagnosis and treatment, therapeutic outcomes have not improved
(5). Therefore, an understanding of
OS pathogenesis is required so that more specific therapeutic
targets can be identified for patients with this fatal malignant
tumor.
Recent studies have demonstrated that microRNAs
(miRNAs or miRs) serve crucial roles in different types of human
cancer (6-8).
miRNAs are a series of non-coding, short RNA molecules that
negatively regulate gene expression via sequence-specific
interactions with the 3'-untranslated regions (UTR) of their target
gene (9). These interactions
suppress translation and/or induce the degradation of mRNAs
(10). The dysregulation of miRNAs
has been demonstrated in nearly all types of human malignancy,
including bladder cancer (11),
colorectal cancer (12), lung cancer
(13) and cervical cancer (14). A variety of miRNAs are differentially
expressed in OS and their deregulation is involved in OS genesis
and development (15,16). miRNAs serve oncogenic or tumor
suppressive roles, and participate in the regulation of a variety
of pathological processes, including cell proliferation, the cell
cycle, apoptosis, metastasis and epithelial-mesenchymal transition
(17-19).
Identifying the roles of dysregulated miRNAs in OS may contribute
to the development of techniques for improved diagnosis, therapy
and prognosis.
miR-652 is an miRNA that has been studied in
pancreatic cancer (20) and
non-small cell lung cancer (21).
However, the specific role and underlying mechanism of miR-652 in
OS has not yet been elucidated. In the current study, the
expression of miR-652 was examined in OS tissues and cell lines.
The effects of miR-652 on the biological role of OS cells were
examined, and the molecular mechanisms underlying the activity of
miR-652 in OS cells were also assessed.
Materials and methods
Tumor specimen collection
A total of 29 pairs of OS and adjacent normal
tissues (2 cm away from tumor tissues) were obtained from 29
patients (age range, 12-27 years; 17 males, 12 females) between May
2015 and March 2017 from The First Affiliated Hospital of Henan
University (Kaifeng, China). No patients were subjected to
chemotherapy, radiotherapy or other treatments prior to surgical
resection and none were suffering from any other disease. All
tissues were snap-frozen in liquid nitrogen and stored at -80˚C.
The current study was approved by the Ethics Committee of Henan
University. Written informed consent was provided by all
participants or their legal guardians prior to enrollment in the
current study.
Cell lines
A total of three human OS cell lines (U2OS, MG-63
and HOS) and a normal human osteoblast (hFOB1.19) were purchased
from the Type Culture Collection of the Chinese Academy of
Sciences. All cell lines were cultured in DMEM supplemented with
10% FBS and 1% penicillin/streptomycin mixture (all, Gibco; Thermo
Fisher Scientific, Inc.), and grown at 37˚C in a humidified
incubator supplied with 5% CO2.
Oligonucleotides, small interfering
RNA (siRNA) and cell transfection
miR-652 mimics, miRNA mimic negative controls
(miR-NC), miR-652 inhibitors, negative control (NC) inhibitors,
siRNA against the expression of kruppel-like factor 9 (KLF9) and NC
siRNA were all produced by Shanghai GenePharma Co., Ltd. U2OS and
HOS cells were plated into six-well plates at a density of
6x105 cells/well 1 day prior to transfection at 37˚C.
The following sequences were used: miR-652 mimics,
5'AAUGGCGCCACUAGGGUUGUG3'; miR-NC, 5'UUCUCCGAACGUGUCACGUTT3';
miR-652 inhibitor, 5'UUACCGCGGUGAUCCCAACAC3'; NC inhibitor,
5'CAGUACUUUUGUGUAGUACAA3'; KLF9 siRNA, 5'CAGUUCCGCUGUCCGCUGU3' and
NC siRNA, 5'UUCUCCGAACGUGUCACGUTT3'. Cells were subsequently
transfected transiently with 100 pmol miR-652 mimic, 100 pmol
miR-NC, 100 pmol miR-652 inhibitor, 100 pmol NC inhibitor, 100 pmol
KLF9 siRNA or 100 pmol NC siRNA using Lipofectamine 2000 (cat. no.
11668019; Invitrogen; Thermo Fisher Scientific, Inc.), according to
manufacturer's protocol. FBS-free DMEM was used during
transfection. At 6 h after transfection, culture medium was
replaced with DMEM containing 10% FBS. Reverse
transcription-quantitative (RT-q)PCR and a transwell invasion assay
were performed at 48 h post-transfection. A Cell Counting Kit-8
(CCK-8) assay and western blotting were performed at 24 and 72 h
after transfection.
Reverse transcription-quantitative PCR
(RT-qPCR)
RT-qPCR analysis was performed to detect miR-652 and
KLF9 mRNA expression form OS tissues, adjacent normal tissues, OS
cell lines (U2OS, MG-63 and HOS) and a normal human osteoblast
(hFOB1.19). The TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to isolate total RNA from tissue
specimens or cells following the manufacturer's protocol. To
measure mRNA expression, total RNA was converted to cDNA using a
TaqMan miRNA Reverse Transcription kit and subjected to qPCR which
was performed using a TaqMan miRNA PCR kit (all, Applied
Biosystems; Thermo Fisher Scientific, Inc.). The temperature
protocol for RT was as follows: 16˚C for 30 min, 42˚C for 30 min
and 85˚C for 5 min. The thermocycling conditions for PCR were: 50˚C
for 2 min; 95˚C for 10 min; 40 cycles of denaturation at 95˚C for
15 sec and annealing/extension at 60˚C for 60 sec. To quantify KLF9
mRNA expression, cDNA was prepared from total RNA using a
PrimeScript RT Reagent kit (Takara Bio, Inc.). qPCR was
subsequently performed using a SYBR Premix Ex Taq™ II kit (Takara
Bio, Inc.). The temperature protocol for RT was: 37˚C for 15 min
and 85˚C for 5 sec. qPCR was then performed with the following
thermocycling conditions: 5 min at 95˚C; 40 cycles of 95˚C for 30
sec and 65˚C for 45 sec. miR-652 expression was normalized to that
of U6 small nuclear RNA and KLF9 mRNA was normalized to that of
GAPDH. Relative gene expression was quantified using the
2-ΔΔCq method (22). The
primer sequences were as follows: miR-652 forward,
5'-ACACTCCAGCTGGGCAACCCTAGGAGAGGGTGC-3' and reverse,
5'-GTGTCGTGGAGTCGGCAATTC-3'; U6 forward,
5'-GCTTCGGCAGCACATATACTAAAAT-3' and reverse,
5'-CGCTTCACGAATTTGCGTGTCAT-3'; KLF9 forward,
5'-ACAGTGGCTGTGGGAAAGTC-3' and reverse, 5'-TCACAAAGCGTTGGCCAGCG-3';
GAPDH forward 5'-TGGTATCGTGGAAGGACTC-3' and reverse,
5'-AGTAGAGGCAGGGATGATG-3'.
CCK-8 assay
Following transfection, U2OS and HOS cells were
incubated for 24 h at 37˚C in a humidified incubator containing 5%
CO2. Cells were collected and subsequently seeded into
96-well plates with an initial density of 2x103
cells/well. After incubations at 37˚C for 0, 24, 48 and 72 h, a
CCK-8 assay (Dojindo Molecular Technologies, Inc.) was performed to
determine cell proliferation according to the manufacturer's
protocol. A total of 10 µl CCK-8 solution was added into each well
prior to incubation at 37˚C for an additional 2 h. Absorbance was
detected at a wavelength of 450 nm using an Enzyme Immunoassay
Analyzer (Bio-Rad Laboratories, Inc.).
Transwell invasion assay
Transfected U2OS and HOS cells were incubated at
37˚C with 5% CO2 for 48 h, harvested and resuspended in
FBS-free DMEM. In total, 5x104 transfected cells
suspended in FBS-free DMEM medium were plated into the upper
compartments of a transwell chamber that were precoated with
Matrigel (all, BD Biosciences). The precoating was conducted in a
37˚C incubator for 4-5 h. The lower compartments were coated with
500 µl DMEM containing 20% FBS. After 24 h of incubation at 37˚C,
cells that had invaded through the membrane were fixed with 4%
paraformaldehyde at 37˚C for 20 min and stained with 0.05% crystal
violet at 37˚C for 30 min. Non-invading cells were scraped off
using a cotton swab. The number of invading cells was counted with
a CKX41 inverted light microscope (magnification x200; Olympus
Corporation).
Bioinformatics analysis
The putative targets of miR-652 were predicted using
TargetScan Human 7.1 (http://www.targetscan.org/vert_71/) and microRNA.org (http://www.microrna.org/microrna/).
Luciferase reporter assay
The 3'-UTR regions of the human KLF9 gene containing
the putative wild-type or mutant miR-652 binding sequences were
amplified by Shanghai GenePharma Co., Ltd., cloned into a pGL3
luciferase vector (Promega Corporation) and respectively named
pGL3-KLF9-3'-UTR wild-type and pGL3-KLF9-3'-UTR mutant. U2OS and
HOS cells were inoculated into 24-well plates at a density of
1.5x105 cells/well and after 24 h incubation at 37˚C,
the recombined luciferase reporter plasmids and miR-652 inhibitor
or NC inhibitor were transfected into cells using Lipofectamine
2000®. At 48 h after transfection, luciferase activity
was determined using a dual-luciferase Reporter Assay system
(Promega Corporation). Firefly luciferase activities were
normalized to that of Renilla luciferase activity.
Western blot analysis
After washing twice with cold PBS, total protein was
extracted from transfected U2OS and HOS cells using RIPA lysis
buffer (Upstate Biotechnology, Inc.). Total protein was
subsequently detected using a BCA assay kit (Beyotime Institute of
Biotechnology). Equal quantities of protein (30 µg/lane) were
loaded for SDS-PAGE on 10% polyacrylamide gels and transferred to
PVDF membranes (EMD Millipore). After blocking with 5% fat-free
milk at room temperature for 2 h, the membranes were washed three
times with Tris-buffered saline containing 0.1% Tween-20 and
incubated overnight at 4˚C with the following primary antibodies:
Rabbit anti-human KLF9 (1:1,000; cat. no. ab227920; Abcam) and
rabbit anti-human GAPDH (1:1,000; cat. no. ab128915; Abcam). GAPDH
was used as an internal reference to confirm equal protein loading.
Membranes were then further probed with horseradish
peroxidase-conjugated goat anti-rabbit IgG secondary antibodies
(1:5,000; cat. no. ab6721; Abcam) at room temperature for 2 h.
Signals were visualized using an enhanced chemiluminescence
detection system (Pierce; Thermo Fisher Scientific, Inc.), and were
analyzed with Quantity One software version 4.62 (BioRad
Laboratories, Inc.).
Statistical analysis
All data were presented as the mean ± standard
deviation. SPSS software version 17.0 (SPSS Inc.) was used to
perform statistical analysis. A paired Student's t-test (for
parametric data) or a Wilcoxon signed-rank test (for non-parametric
data) was used to analyze data between two groups, while a one-way
ANOVA followed by a Tukey's post hoc test was used to compare the
differences between three or more groups. The association between
miR-652 and KLF9 mRNA levels in OS tissue was assessed using
Spearman's correlation analysis. P<0.05 was considered to
indicate a statistically significant result.
Results
miR-652 is highly expressed in human
OS tissues and cell lines
To reveal the expression pattern of miR-652 in OS,
RT-qPCR analysis was performed using 29 pairs of OS tissue and
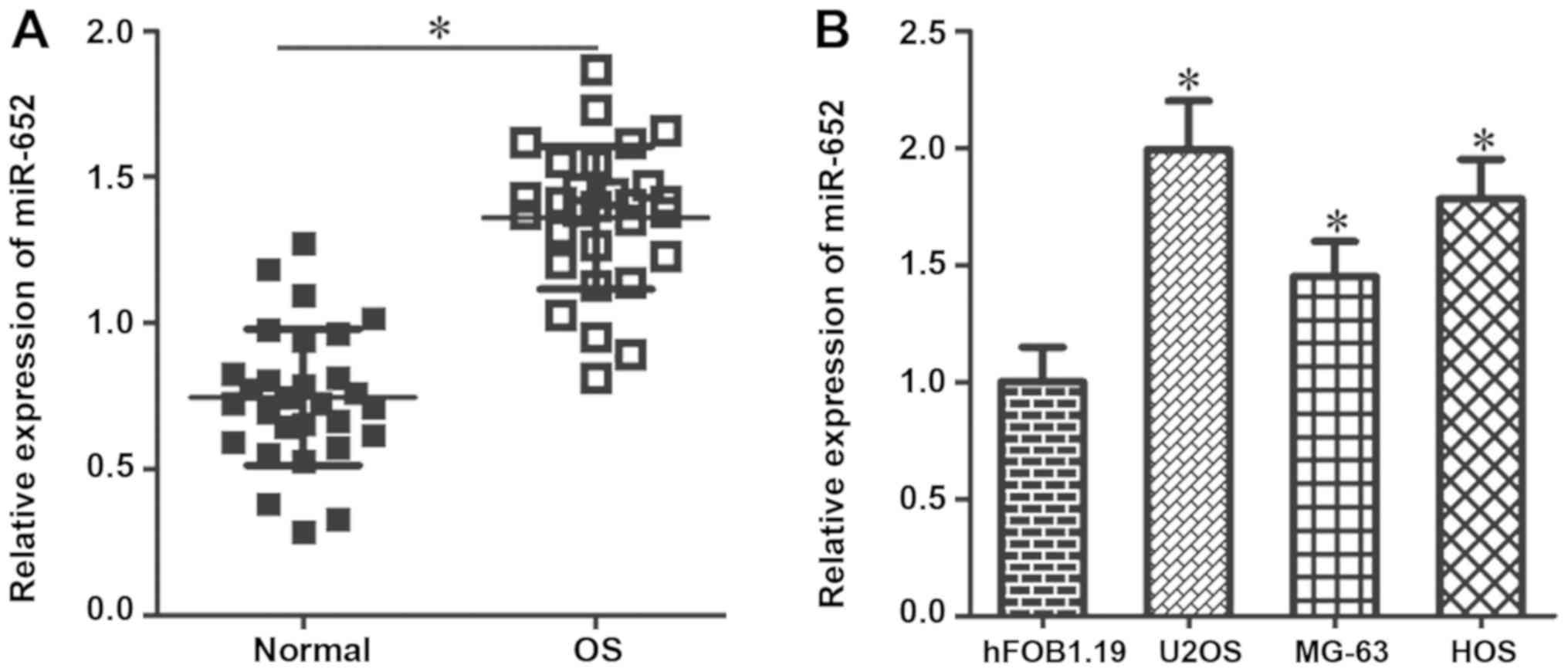
adjacent normal tissue. The results revealed that miR-652
expression was significantly higher in OS tissues compared with
adjacent normal tissues (Fig. 1A;
P<0.05). The expression of miR-652 in three human OS cell lines
(U2OS, MG-63 and HOS) and a normal human osteoblast cell line
(hFOB1.19) was also assessed. The results demonstrated that miR-652
expression was significantly higher in all three OS cell lines
compared with hFOB1.19 cells (Fig.
1B; P<0.05). The results indicated that the upregulation of
miR-652 may serve an important role in the pathogenesis of OS.
miR-652 inhibition impairs OS cell
proliferation and invasion
To determine the functional roles of miR-652 in OS,
U2OS and HOS cells were transfected with a miR-652 inhibitor or NC
inhibitor. These cell lines were selected as they exhibited a
relatively higher miR-652 expression than the other cell lines
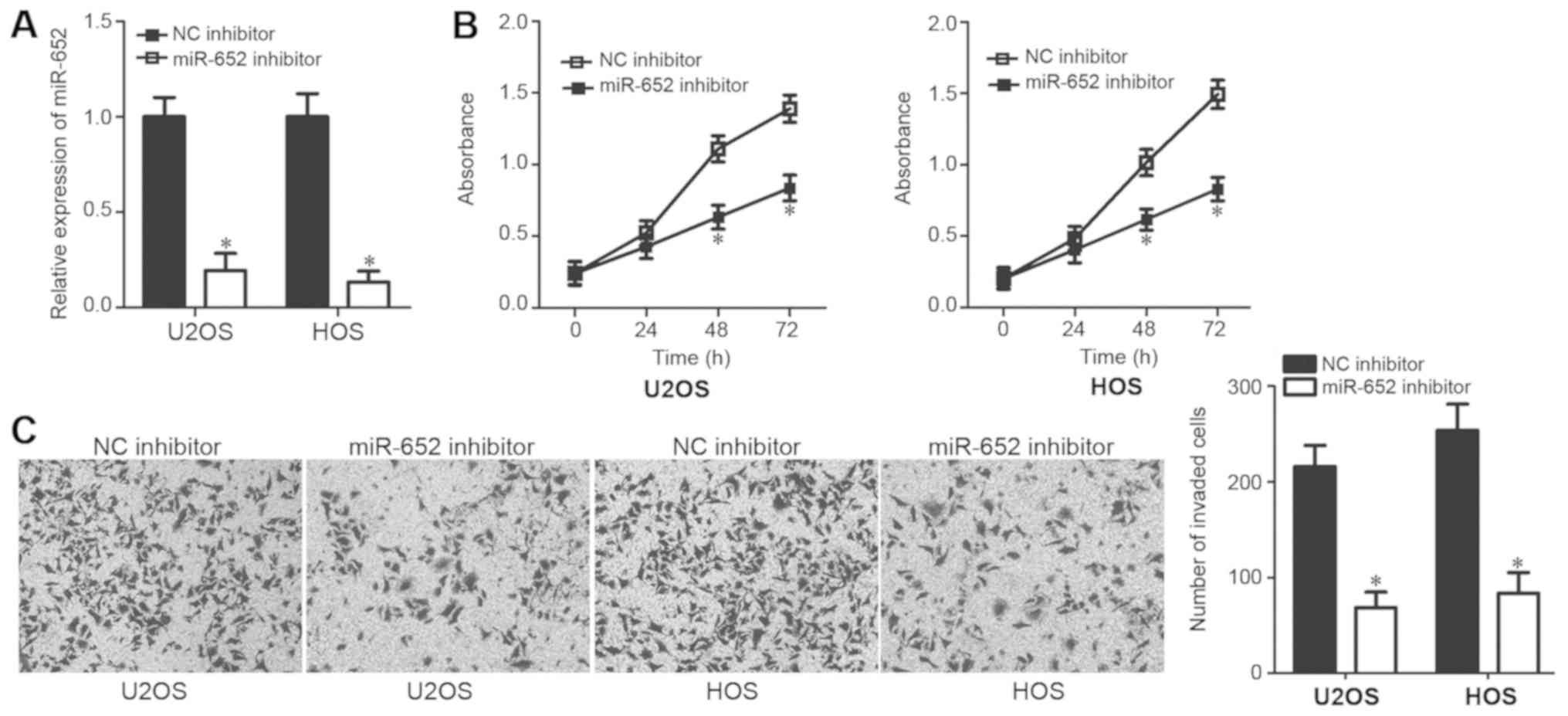
assessed. RT-qPCR analysis revealed that miR-652 expression was
significantly downregulated in U2OS and HOS cells after
transfection with the miR-652 inhibitor (Fig. 2A; P<0.05). The regulatory effect
of miR-652 on OS cell proliferation was assessed via a CCK-8 assay.
The inhibition of miR-652 expression in U2OS and HOS cells resulted
in the suppression of cell proliferation at 48 and 72 h (Fig. 2B; P<0.05). In addition, a
transwell invasion assay was performed to assess the effect of
miR-652 in OS cell invasion. The invasive ability of U2OS and HOS
cells was significantly inhibited following treatment with the
miR-652 inhibitor (Fig. 2C;
P<0.05). The results demonstrated that miR-652 functions as an
oncogene in OS by regulating cell proliferation and invasion.
KLF9 is a direct target of miR-652 in
OS cells
The oncogenic role of miR-652 in OS cells was
assessed using bioinformatics analysis to determine potential
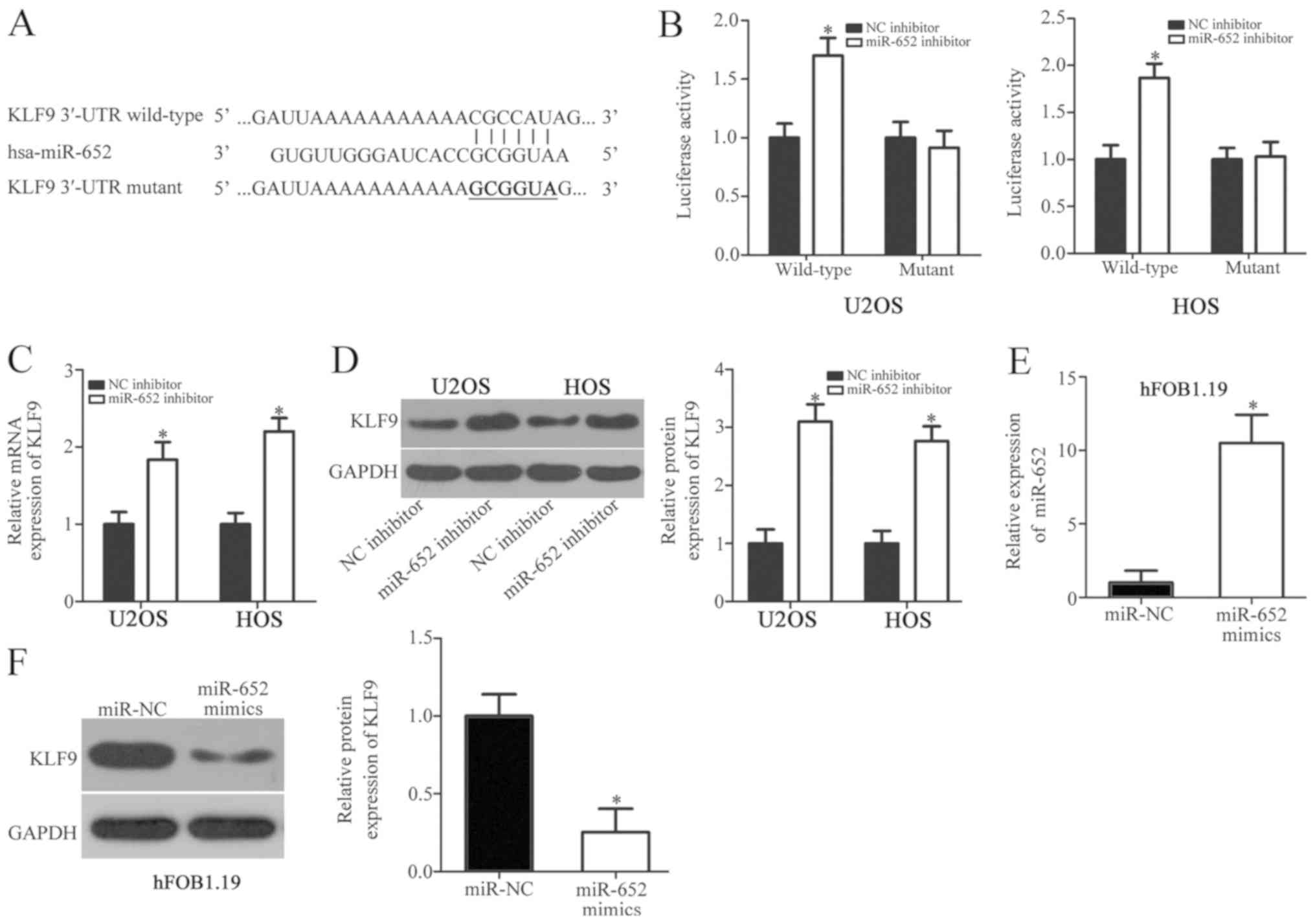
miR-652 targets. A combination of target sequences were located
between the 3'-UTR of KLF9 and miR-652 (Fig. 3A). Subsequently, KLF9 was selected
for further experimental identification due to its association with
OS formation and progression (23).
Luciferase reporter plasmids were constructed and co-transfected
with a miR-652 inhibitor or NC inhibitor into U2OS and HOS cells.
At 48 h after transfection, luciferase activity was determined. The
results revealed that the downregulation of miR-652 significantly
increased the luciferase activity of the plasmid carrying the
wild-type 3'-UTR of KLF9 (Fig. 3B;
P<0.05). However, the luciferase activity of the plasmid
carrying the mutant 3'-UTR was not significantly altered,
indicating that miR-652 directly binds to the 3'-UTR of KLF9
(Fig. 3B). In subsequent RT-qPCR and
western blot analysis, the expression of KLF9 mRNA (Fig. 3C; P<0.05) and protein (Fig. 3D; P<0.05) were significantly
upregulated in U2OS and HOS cells transfected with the miR-652
inhibitor. To determine whether miR-652 was able to regulate KLF9
expression in hFOB1.19, cells were transfected with miR-652 mimics
or an miR-NC. RT-qPCR analysis demonstrated that miR-652 was
upregulated in hFOB1.19 cells after transfection with miR-652
mimics (Fig. 3E; P<0.05). KLF9
protein levels were also downregulated in miR-652 mimic-transfected
hFOB1.19 cells compared with miR-NC transfected cells (Fig. 3F; P<0.05). The results
demonstrated that KLF9 is a direct target gene of miR-652 in OS and
normal osteoblast cells.
KLF9 expression is downregulated in OS
tissue samples and is inversely correlated with the expression of
miR-652
To evaluate the association between miR-652 and KLF9
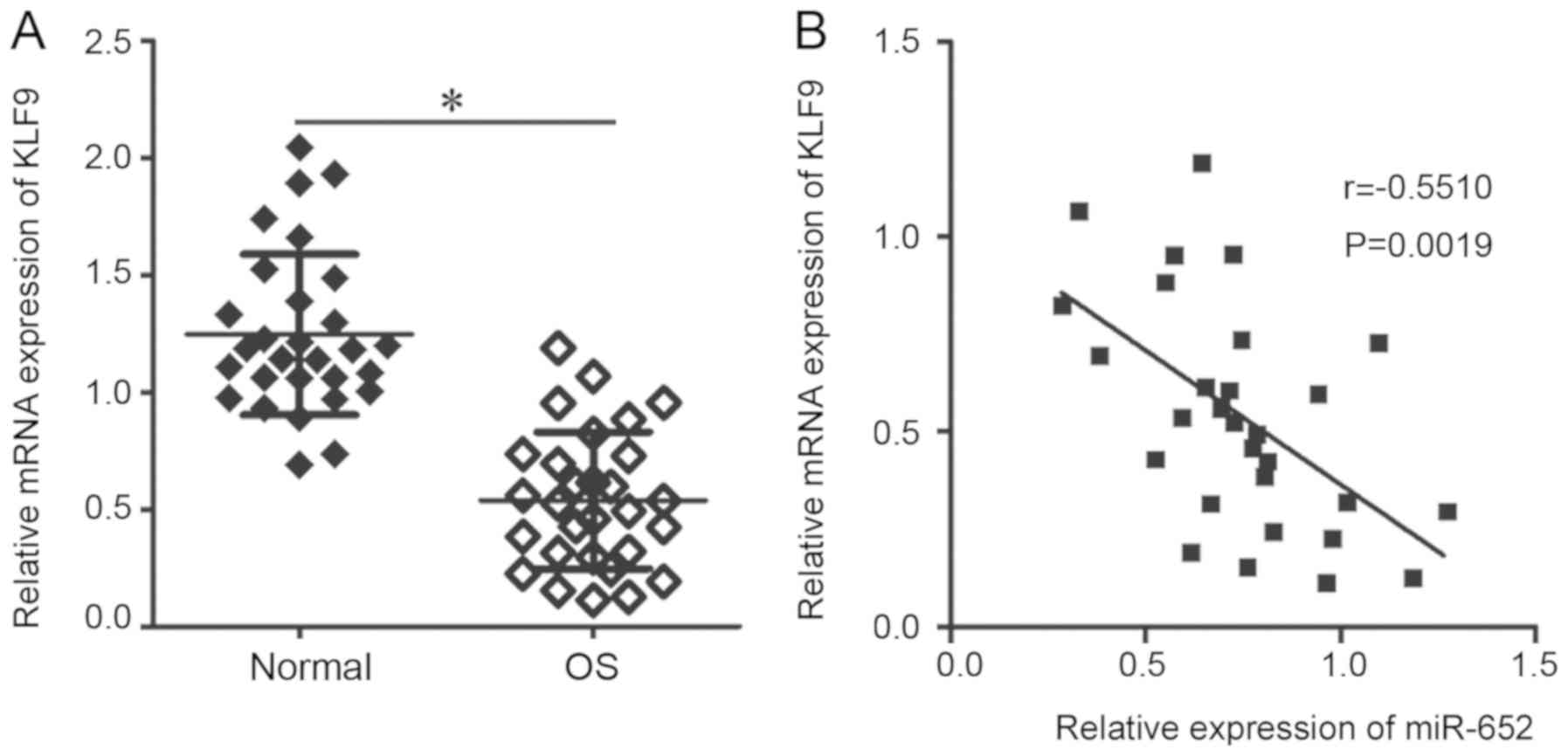
in OS, KLF9 expression was assessed in 29 pairs of OS and adjacent
normal tissue. RT-qPCR analysis revealed that KLF9 mRNA expression
was significantly lower in OS tissues compared with adjacent normal
tissues (Fig. 4A; P<0.05).
Furthermore, Spearman's correlation analysis identified an inverse
correlation between miR-652 and KLF9 mRNA in OS tissues (Fig. 4B; r=-0.5510; P=0.0019). The results
indicated that the downregulation of KLF9 in OS tissue is at least,
in part, induced by the overexpression of miR-652.
KLF9 knockdown counteracts the miR-652
inhibitor-induced suppression of OS cell proliferation and
invasion
As aforementioned, miR-652 was implicated in the
regulation of OS cell proliferation and invasion and KLF9 was
validated as a direct target of miR-652. Therefore, whether KLF9
mediated the effects of miR-652 was subsequently assessed.
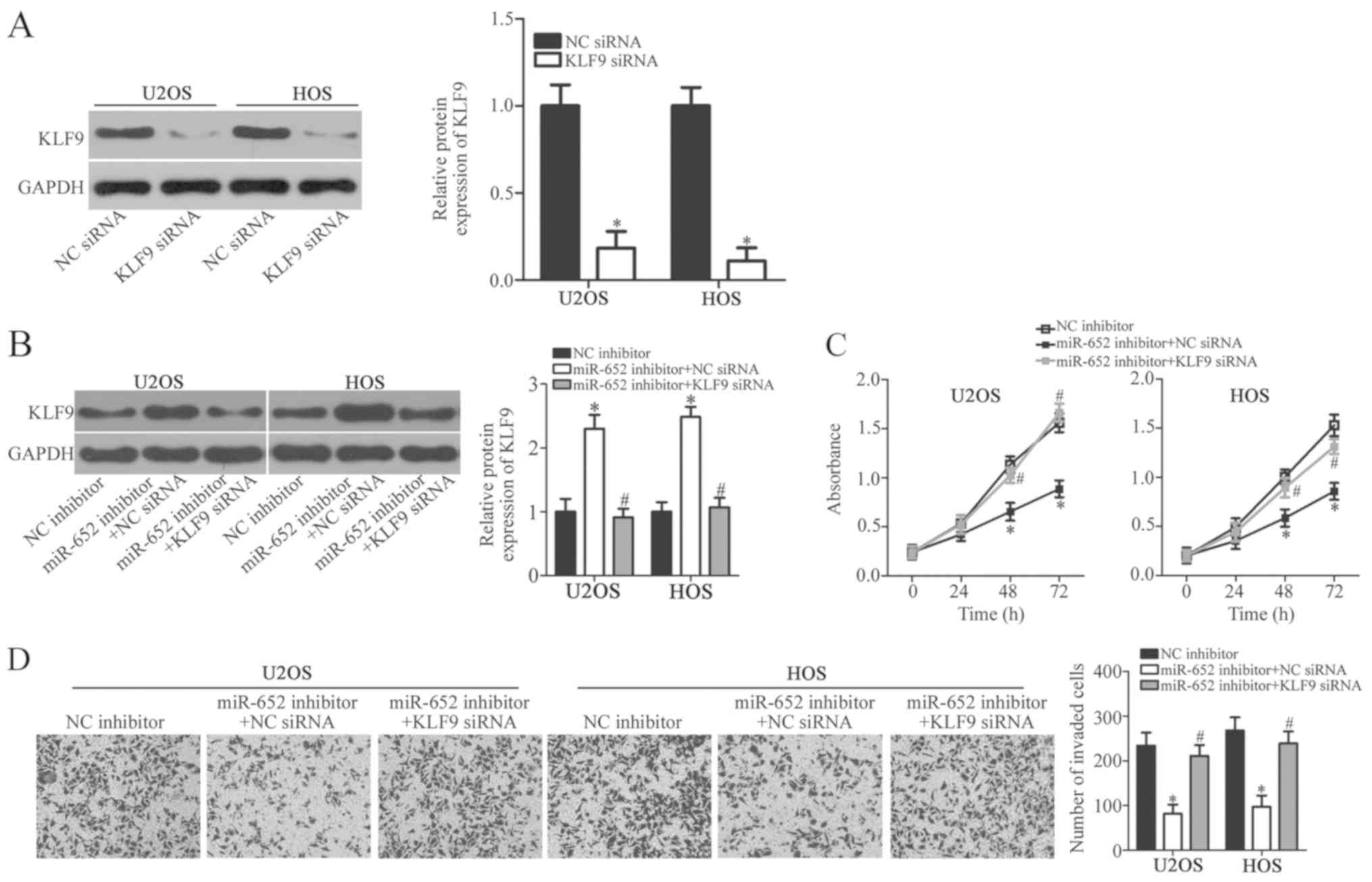
siRNA-mediated knockdown was confirmed in U2OS and Hos cells
(Fig. 5A) The siRNA-mediated
knockdown of KLF9 was performed in miR-652 inhibitor-transfected
U2OS and HOS cells. Western blot analysis demonstrated that the
upregulation of KLF9 protein expression in U2OS and HOS cells
induced by the miR-652 inhibitor was recovered by co-transfection
with KLF9 siRNA (Fig. 5B;
P<0.05). CCK-8 and transwell invasion assays indicated that
recovered KLF9 expression counteracted the impaired proliferation
of U2OS and HOS cells (Fig. 5C;
P<0.05) and invasion (Fig. 5D;
P<0.05) induced by the miR-652 inhibitor at 48 and 72 h. The
results indicated that KLF9 mediated the functional roles of
miR-652 in OS cell proliferation and invasion.
Discussion
Previous studies have determined that a variety of
miRNAs are abnormally expressed in OS and that their abnormal
expression serves a key role in malignant development (15,16,24).
Understanding the mechanism responsible for OS carcinogenesis and
progression is therefore critical for the development of effective
therapeutic strategies for this disease (25). An in-depth understanding of the
functional roles and underlying mechanisms of dysregulated miRNAs
in OS may be significant for cancer research and important for the
identification of novel therapeutic targets. To the best of our
knowledge, the current study assessed the expression status of
miR-652 in OS and determined the detailed roles of miR-652 in OS
progression for the first time. The molecular mechanisms of miR-652
activity in OS cells were also assessed.
miR-652 has been revealed to be downregulated in
pancreatic cancer tissues and cell lines. A decreased expression of
miR-652 was significantly correlated with tumor stage, lymphatic
invasion and metastasis in patients with pancreatic cancer
(20). In contrast, miR-652 is
highly expressed in non-small cell lung cancer tissue and cell
lines. The upregulation of miR-652 was associated with lymph node
metastasis, TNM stage and prognosis in patients with non-small cell
lung cancer (21). These
inconsistent results indicate that the expression of miR-652
exhibits tissue specificity in human malignancies. However, the
expression status of miR-652 in OS remains unclear. In the current
study, RT-qPCR was performed to detect miR-652 and the results
revealed that it was significantly upregulated in OS tissues and
cell lines. The results also demonstrated that miR-652 may be used
as a potential biomarker in the diagnosis of OS.
The dysregulation of miR-652 contributes to the
malignant phenotype of human cancer. miR-652 has been identified as
a tumor suppressor in pancreatic cancer (20). The upregulation of miR-652 expression
abolishes the acidity-induced epithelial-mesenchymal transition of
pancreatic cancer cells via the negative regulation of zinc finger
E-Box binding homeobox 1(20).
miR-652 also serves oncogenic roles in the progression of non-small
cell lung cancer and directly targets lethal giant larvae 1 to
affect cell proliferation, apoptosis, migration and invasion
(21). However, the roles of miR-652
in OS progression are unknown. In the current study, CCK-8 and
transwell invasion assays demonstrated that miR-652 inhibition
resulted in a significant suppression of OS cell proliferation and
invasion. The results indicated that miR-652 may be a promising
therapeutic target for anticancer therapy.
The identification of the direct target genes of
miR-652 is important for understanding its role in carcinogenesis
and progression (26). The results
of the present study demonstrated that KLF9 was a direct target
gene of miR-652 in OS cells. Bioinformatic predictions indicated
KLF9 as a putative target of miR-652. Subsequently, a luciferase
reporter assay, RT-qPCR and western blot analysis revealed that
miR-652 regulated the expression of KLF9 by binding to its 3'-UTR
in OS cells. KLF9 was also downregulated in OS tissues and this
downregulation was inversely correlated with miR-652 expression.
Furthermore, the downregulation of KLF9 decreased the miR-652
inhibitor-induced suppression of OS cell proliferation and
invasion. These results may provide sufficient evidence to support
the hypothesis that KLF9 is a direct target gene of miR-652 in OS
cells.
KLF9 is a member of the KLF family (27) and has been previously reported to be
downregulated in various types of human malignancy, including
pancreatic ductal adenocarcinoma (28), hepatocellular carcinoma (29), colorectal cancer (30) and prostate cancer (31). KLF9 serves as a tumour suppressor at
tumour onset and development, affecting cell proliferation,
apoptosis, metastasis and tumorigenicity (32-34).
Small quantities of KLF9 are expressed in OS and this low
expression level may modulate OS aggression (23). The present study is, to the best of
our knowledge, the first to demonstrate that miR-652 regulated KLF9
expression and may therefore inhibit the progression of OS. The
results of the current study revealed that miR-652 inhibition or
KLF9 restoration may be an effective therapeutic technique to treat
patients with OS in the future.
In the current study, the correlation between
miR-652 expression and clinicopathological factors in patients with
OS was not determined. Therefore, in following investigations, the
collection of more tissues and the determination of clinical
significance should be assessed. The influence of miR-652
overexpression in the malignant development of OS cells was also
not clarified. miR-652 mimics should be utilized in future studies
to increase endogenous miR-652 expression. In addition, a series of
functional experiments should be performed to evaluate the effects
of miR-652 overexpression in OS cells. The regulatory effects of
miR-652 in the apoptosis and migration of OS cells was not assessed
in the present study, and as such, flow cytometry analysis and
transwell migration assay should be applied in future studies to
determine this.
In conclusion, miR-652 was upregulated in OS tissues
and cell lines. The downregulation of miR-652 also inhibited the
proliferation and invasion of OS cells by targeting KLF9 directly.
All the results obtained in the current study may provide a basis
for the identification of novel therapeutic targets for the
prevention of OS and for OS therapy.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL designed the current study, wrote the manuscript
and performed the luciferase reporter assay. YJ and LY performed
reverse transcription-quantitative PCR, the cell counting kit-8
assay, the transwell invasion assay and western blot analysis. All
authors have read and approved the final draft.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Henan University and was performed in accordance with
the Declaration of Helsinki and the guidelines of the Ethics
Committee of The First Affiliated Hospital of Henan University.
Written informed consent was provided by all participations prior
to their enrollment.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ando K, Mori K, Verrecchia F, Marc B,
Redini F and Heymann D: Molecular alterations associated with
osteosarcoma development. Sarcoma. 2012(523432)2012.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Picci P: Osteosarcoma (osteogenic
sarcoma). Orphanet J Rare Dis. 2(6)2007.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Tan ML, Choong PF and Dass CR:
Osteosarcoma: Conventional treatment vs. Gene therapy. Cancer Biol
Ther. 8:106–117. 2009.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Ferrari S, Palmerini E, Staals EL, Mercuri
M, Franco B, Picci P and Bacci G: The treatment of nonmetastatic
high grade osteosarcoma of the extremity: Review of the italian
rizzoli experience. Impact on the future. Cancer Treat Res.
152:275–287. 2009.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Pan Y, Lu L, Chen J, Zhong Y and Dai Z:
Identification of potential crucial genes and construction of
MicroRNA-MRNA negative regulatory networks in osteosarcoma.
Hereditas. 155(21)2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
To KK, Tong CW, Wu M and Cho WC: MicroRNAs
in the prognosis and therapy of colorectal cancer: From bench to
bedside. World J Gastroenterol. 24:2949–2973. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Yuan HL, Wang T and Zhang KH: MicroRNAs as
potential biomarkers for diagnosis, therapy and prognosis of
gastric cancer. Onco Targets Ther. 11:3891–3900. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Sharma N and Baruah MM: The microRNA
signatures: Aberrantly expressed miRNAs in prostate cancer. Clin
Transl Oncol. 21:126–144. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Han K, Chen X, Bian N, Ma B, Yang T, Cai
C, Fan Q, Zhou Y and Zhao TB: MicroRNA profiling identifies MiR-195
suppresses osteosarcoma cell metastasis by targeting CCND1.
Oncotarget. 6:8875–8889. 2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chen X, Jia C, Jia C, Jin X and Gu X:
MicroRNA-374a Inhibits aggressive tumor biological behavior in
bladder carcinoma by suppressing wnt/β-Catenin signaling. Cell
Physiol Biochem. 48:815–826. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Tao Y, Ma C, Fan Q, Wang Y, Han T and Sun
C: MicroRNA-1296 facilitates proliferation, migration and invasion
of colorectal cancer cells by targeting SFPQ. J Cancer.
9:2317–2326. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Yang JZ, Bian L, Hou JG and Wang HY:
MiR-550a-3p promotes non-small cell lung cancer cell proliferation
and metastasis through down-regulating TIMP2. Eur Rev Med Pharmacol
Sci. 22:4156–4165. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Wu Y, Huang J, Xu H and Gong Z:
Over-Expression of MiR-15a-3p enhances the radiosensitivity of
cervical cancer by targeting tumor protein D52. Biomed
Pharmacother. 105:1325–1334. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Leichter AL, Sullivan MJ, Eccles MR and
Chatterjee A: MicroRNA expression pat terns and signalling pathways
in the development and progression of childhood solid tumours. Mol
Cancer. 16(15)2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Kim YH, Goh TS, Lee CS, Oh SO, Kim JI,
Jeung SH and Pak K: Prognostic value of microRNAs in osteosarcoma:
A meta-analysis. Oncotarget. 8:8726–8737. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Wang Z, Zheng C, Jiang K, He J, Cao X and
Wu S: MicroRNA-503 suppresses cell proliferation and invasion in
osteosarcoma via targeting insulin-like growth factor 1 receptor.
Exp Ther Med. 14:1547–1553. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Ma C, Han J, Dong D and Wang N:
MicroRNA-152 suppresses human osteosarcoma cell proliferation and
invasion by targeting E2F transcription factor 3. Oncol Res.
26:765–773. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Liu K, Sun X, Zhang Y, Liu L and Yuan Q:
MiR-598: A tumor suppressor with biomarker significance in
osteosarcoma. Life Sci. 188:141–148. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Deng S, Li X, Niu Y, Zhu S, Jin Y, Deng S,
Chen J, Liu Y, He C, Yin T, et al: MiR-652 inhibits acidic
microenvironment-induced epithelial-mesenchymal transition of
pancreatic cancer cells by targeting ZEB1. Oncotarget.
6:39661–39675. 2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Yang W, Zhou C, Luo M, Shi X, Li Y, Sun Z,
Zhou F, Chen Z and He J: MiR-652-3p is upregulated in non-small
cell lung cancer and promotes proliferation and metastasis by
directly targeting Lgl1. Oncotarget. 7:16703–16715. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Peng N, Miao Z, Wang L, Liu B, Wang G and
Guo X: MiR-378 promotes the cell proliferation of osteosarcoma
through down-regulating the expression of kruppel-like factor 9.
Biochem Cell Biol. 96:515–521. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Sampson VB, Yoo S, Kumar A, Vetter NS and
Kolb EA: MicroRNAs and potential targets in osteosarcoma: Review.
Front Pediatr. 3(69)2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Smolle MA, Leithner A, Posch F, Szkandera
J, Liegl-Atzwanger B and Pichler M: MicroRNAs in different
histologies of soft tissue sarcoma: A comprehensive review. Int J
Mol Sci. 18(E1960)2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Yao J, Zhang P, Li J and Xu W:
MicroRNA-215 acts as a tumor suppressor in breast cancer by
targeting AKT serine/threonine kinase 1. Oncol Lett. 14:1097–1104.
2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
McConnell BB and Yang VW: Mammalian
kruppel-like factors in health and diseases. Physiol Rev.
90:1337–1381. 2010.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Mao Z, Fan X, Zhang J, Wang X, Ma X,
Michalski CW and Zhang Y: KLF9 is a prognostic indicator in human
pancreatic ductal adenocarcinoma. Anticancer Res. 37:3795–3799.
2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Sun J, Wang B and Liu Y, Zhang L, Ma A,
Yang Z, Ji Y and Liu Y: Transcription factor KLF9 suppresses the
growth of hepatocellular carcinoma cells vivo and positively
regulates p53 expression. Cancer Lett. 355:25–33. 2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Kang L, Lu B, Xu J, Hu H and Lai M:
Downregulation of kruppel-like factor 9 in human colorectal cancer.
Pathol Int. 58:334–338. 2008.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Shen P, Sun J, Xu G, Zhang L, Yang Z, Xia
S, Wang Y, Liu Y and Shi G: KLF9, a transcription factor induced in
flutamide-caused cell apoptosis, inhibits AKT activation and
suppresses tumor growth of prostate cancer cells. Prostate.
74:946–958. 2014.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Bai XY, Li S, Wang M, Li X, Yang Y, Xu Z,
Li B, Li Y, Xia K, Chen H and Wu H: Kruppel-like factor 9
down-regulates matrix metalloproteinase 9 transcription and
suppresses human breast cancer invasion. Cancer Lett. 412:224–235.
2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Huang S, Wang C, Yi Y, Sun X, Luo M, Zhou
Z, Li J, Cai Y, Jiang X and Ke Y: Kruppel-like factor 9 inhibits
glioma cell proliferation and tumorigenicity via downregulation of
miR-21. Cancer Lett. 356:547–555. 2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Zhang QH, Dou HT, Tang YJ, Su S and Liu
PS: Lentivirus-mediated knockdown of Kruppel-like factor 9 inhibits
the growth of ovarian cancer. Arch Gynecol Obstet. 291:377–382.
2015.PubMed/NCBI View Article : Google Scholar
|