Introduction
Bronchial asthma is an intractable pulmonary disease
associated with bronchial smooth muscle hyperresponsiveness, damage
of airway epithelium and continuous inflammation in the airways of
patients (1-3).
At the steady state, the intrapulmonary epithelium serves as a
barrier against inhaled microbes and pollutants and is essential
for maintaining lung homeostasis. Club cells are characterized as
abundant, discrete, electron-dense granules and represent important
secretory cells residing in the conducting epithelium that respond
to environmental insult through the release of biologically active
factors, including secretoglobin family 1A member 1 (Scgb1a1, also
called as CCSP), Scgb3a1, Scgb3a2, mucins (Muc), lactotransferrin,
cytokines and chemokines (4-7).
Club cells either exhibit antibacterial activity or recruit immune
cells to the airways to clear foreign substances (7).
Constant foreign challenge (either inhaled particles
or pathogens) are able to damage the pulmonary epithelium and
result in damage closely related to that of lung diseases (8). Epithelial shedding is a commonly
described event in bronchial asthma, respiratory infections,
chronic bronchitis, fibrosis and interstitial lung disease
(9-11).
After lung injury, rapid regenerating programs, including the
proliferation and differentiation of regionally distributed
epithelial progenitor cells, are necessary to maintain a protective
barrier. In the conducting airway, Club cells are commonly
considered progenitors for goblet cells and ciliated cells
(12,13). Club cells are distributed in the
conducting airway epithelium from the proximal to distal axis in
both humans and mice (14,15). Furthermore, in humans, Club cells
substantially contribute to the proliferation of airway epithelium
and represent an important cell population involved in maintaining
the normal epithelium, particularly the bronchiolar epithelium
(15). Therefore, it is essential
to reveal the mechanisms underlying Club cell function.
Gamma-aminobutyric acid (GABA) is known to have a
crucial role in modulating the function of inhibitory synaptic
activity. GABA is also produced by lung cells, including pulmonary
neuroendocrine cells (PNECs) (16).
Song et al (17) established
hereditary tracking mice and demonstrated that PNEC cells are able
to differentiate into Club cells and ciliated cells, but that the
selective killing of PNECs does not affect the regeneration of Club
cells. Therefore, it is possible that PNECs communicate with Club
cells during airway epithelial regeneration. In addition, Barrios
et al (16) discovered that
PNECs secrete GABA and induce goblet cell hyperplasia in primate
models. There are two types of GABA receptors, namely ligand-gated
ion channels GABAA and G protein-coupled receptors GABAB. GABAA
receptor pi (Gabrp) is a member of the GABAA receptor family and
reportedly present on airway and/or alveolar epithelial cells
(16). However, it has remained
elusive whether GABA regulates the differentiation of Club cells
into mucus-secreting goblet cells in the lung.
In the present study, Club cells from mouse lung
tissue were isolated using fluorescence-activated cell sorting
(FACS), transcriptome analysis was performed and an in vivo
naphthalene-induced lung injury model was established to confirm
that Gabrp is highly expressed in Club cells. Furthermore, organoid
culture of Club cells indicated that inhibition of Gabrp activity
with bicuculline methiodide (BMI) blocked Club cell proliferation
and goblet cell differentiation. These results suggested a possible
role for GABA signaling in airway epithelial regeneration and mucus
production.
Materials and methods
Reagents
Biotin-conjugated CD31 antibody (cat. no.
13-0311-85), biotin-conjugated CD34 antibody (cat. no. 13-0341-85),
biotin-conjugated CD45 antibody (cat. no. 13-0451-85),
phycoerythrin (PE)-cyanine 7 (Cy7)-conjugated epithelial cell
adhesion molecule (EpCAM) antibody (cat. no. 118216),
allophycocyanin (APC)-conjugated stem cell antigen (Sca)-1 antibody
(17-5981-81), PE-conjugated CD24 antibody (cat. no. 12-0242-83) and
APC-Cy7-conjugated streptavidin (cat. no. 47-4317-82) were
purchased from eBioscience. BMI was purchased from Sigma-Aldrich
(Merck KGaA). The IQ™ SYBR Green PCR kit was from Bio-Rad
Laboratories, Inc. and SB431542 from Ascent Scientific LLC.
Mice
Wild type mice (purchased from SPF (Beijing)
Biotechnology Co., Ltd.) were maintained under specific
pathogen-free conditions at the animal facility of Tianjin
University Haihe Hospital (Tianjin, China). They were kept under a
12-h light/dark cycle with free access to food and water. A total
of 60 male mice aged between 8 and 16 weeks and weighing 22-24 g
were used for in vitro culture experiments and in
vivo lung injury experiments. Scgb1a1-CreER™; Rosa26-mTmG
(Scgb1a1-mT/mG) mice were established by mating Scgb1a1-CreER™
(Jackson Laboratory) and Rosa26 mTmG
(Gt(ROSA)26Sortm4(ACTB-tdTomato,-EGFP)Luo) mice donated
by Dr Ning of Nankai University. The naphthalene injury experiment
was performed as described previously (18): Age- and sex-matched mice were
injected intraperitoneally with or without 250 mg/kg naphthalene
(Thermo Fisher Scientific, Inc.) dissolved in corn oil
(Sigma-Aldrich; Merck KGaA). At days 0, 3 and 10 post-injection,
mice (day 0, n=10; day 3, n=8; day 10, n=10) were euthanized with
an overdose of pentobarbital sodium (150 mg/kg, intraperitoneally)
and lung tissues were collected for gene expression analysis. All
mice were treated with strict adherence to the protocol approved by
the Haihe Hospital Animal Care and Use Committee (Tianjin, China;
no. 2020HHSQKT-001).
FACS
Preparation of the cell suspension for FACS was
performed as previously described (19). In brief, lung epithelial cells were
isolated by elastase digestion and resuspended in Hank's balanced
salt solution (HBSS) + buffer (HBSS supplemented with 2% fetal
bovine serum, 0.1 mM EDTA, 10 mM HEPES, 100 IU/ml penicillin and
100 µg/ml streptomycin). Primary antibodies, including
biotin-conjugated CD31 antibody (1:40), biotin-conjugated CD34
antibody (1:15), biotin-conjugated CD45 antibody (1:70),
PE-Cy7-conjugated EpCAM antibody (1:50), APC-conjugated Sca-1
antibody (1:100) and PE-conjugated CD24 antibody (1:25), were added
to the cell suspension, and biotin-conjugated antibodies were
identified by incubating cells further with APC-Cy7-conjugated
streptavidin (1:100). Dead cells were removed by using
7-amino-actinomycin D (FACS Aria III; BD Biosciences).
Microarray analysis
The sorted epithelial cells were lysed using an
RNeasy Mini kit (Qiagen GmbH) to isolate RNA. The amount of RNA was
determined using a NanoDrop spectrophotometer (Thermo Fisher
Scientific, Inc.) and 0.2 µg of RNA was used for Affymetrix
microarray analysis using mouse genome 430 2.0 arrays (Affymetrix;
Thermo Fisher Scientific, Inc.). Data were annotated with
Affymetrix Expression Console software (Affymetrix, Inc.).
GeneSpring GX software (http://www.home.agilent.com/agilent/home.jspx?&cc=US&lc=eng;
Agilent Technologies, Inc.) was adopted to analyze the
differentially expressed genes between groups. Raw data were
transformed using a base-2 logarithm (Log2x) function to
calculate the fold change between two groups. Significantly
differently expressed genes were defined as those fulfilling a
Student's t-test P<0.05 and a threshold number of
misclassifications of ≤1. Treeview software (v.1.6; https://treeview.software.informer.com/1.6/) was
adopted to generate heatmaps of gene expression and pathway
analysis was performed using the online Gather Kyoto Encyclopedia
of Genes and Genomes analysis database (KEGG; http://gather.genome.duke.edu/).
Organoid culture
The MLg2908 mouse lung fibroblast cell line CCL-206
(American Type Culture Collection) was cultured in Dulbecco's
modified Eagle's medium (DMEM) (Gibco; Thermo Fisher Scientific,
Inc.)_ with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.). In vitro organoid culture of Club cells
in Matrigel has been described previously (19). In brief, fractionated Club cells
were mixed with MLg cells in Matrigel basic medium (1:1). The
culture medium included DMEM/F12 (Gibco; Thermo Fisher Scientific,
Inc.), insulin/transferrin/selenium (Invitrogen; Thermo Fisher
Scientific, Inc.), 10% fetal bovine serum, 100 IU/ml penicillin,
100 µg/ml streptomycin and 10 µM SB431542. The mixture was then
loaded into Transwell filter inserts (Greiner BioOne), which were
placed in 24-well plates containing the culture medium and cultured
in a humidified atmosphere with 5% CO2 at 37˚C.
Resulting organoids were visualized using a Zeiss Axiovert40
inverted fluorescent microscope (Zeiss AG). To inhibit the
Gabrp-mediated pathway, the cell culture medium was supplemented
with 20 µg/ml of BMI (20-23),
and co-cultured for 10 days.
RNA extraction and reverse
transcription-quantitative (RT-q)PCR
As described previously (24,25),
total RNA was extracted from sorted mouse Club cells or Club
cell-derived organoids using a Qiagen RNeasy Mini kit (Qiagen GmbH)
according to the manufacturer's protocol. The composition of the
reaction mixture for reverse transcription was as follows: 5 µl 10X
PCR buffer, 15 µl MgCl2, 1 µl dNTP, 0.625 µl SSIII, 0.5
µl RRI, 1 µl hexamers, 21.875 µl RNA-free water and 5 µl RNA.
Transcription was conducted using the following conditions: 25˚C
for 10 min, 48˚C for 40 min and 95˚C for 5 min. Quantitative
analysis of transcripts was performed with a SYBR Green kit
according to the manufacturer's protocol. PCR detection was
performed using a Realplex4 real-time PCR System (Eppendorf).
Intron-spanning gene-specific primers are listed in Table I. PCR amplification was conducted
using the following conditions: Initial denaturation at 95˚C for 2
min, followed by 40 cycles of 95˚C for 25 sec for denaturation,
60˚C for 25 sec for primer annealing and 72˚C for 20 sec for
extension. In the isolated the Club cells and clone samples, the
mRNA expression level of the target genes was calculated by the
comparative Ct (threshold cycle) method. β-actin was used as a
housekeeping gene to normalize the amount of RNA in the same
sample. Specific ∆Ct was calculated as follows: ∆Ct =
(Ctβ-actin)-(Cttarget); relative expression
was defined as 2-∆∆Cq (26).
 | Table ISequences of primers for PCR. |
Table I
Sequences of primers for PCR.
| Gene | Forward primer | Reverse primer |
|---|
| β-actin |
5'-GGCCAACCGTGAAAAGATGA-3' |
5'-CAGCCTGGATGGCTACGTACA-3' |
| Scgb1a1 |
5'-ATCAGAGTCTGGTTATGT-3' |
5'-ATCCACCAGTCTCTTCAG-3' |
| FoxJ1 |
5'-AGTGGATCACGGACAACTTCTGCT-3' |
5'-ATCCTTCTCCCGAGGCACTTTGAT-3' |
| Reg3g |
5'-ACACTGGGCTATGAACCCAACAGA-3' |
5'-ACCACAGTGATTGCCTGAGGAAGA-3' |
| Sult1d1 |
5'-TGGAACAACTTGGGTCAGTG-3' |
5'-AAGCTCCATGAATGGTACTCG-3' |
| Iyd |
5'-TGTCATCAAAGCAGCAGGAACAGC-3' |
5'-TTCAGGTCTGTGACCCATCGCTTT-3' |
| Fmo3 |
5'-CAGCATTTACCAATCGGTCTTC-3' |
5'-TGACTTCCCATTTGCCAGTAG-3' |
| Cp |
5'-CCTACAAGGCCTCACAATGCA-3' |
5'-TCATTGCCCATTCCCATCA-3' |
| Casp4 |
5'-AGGCTACGATGTGGTGGTGAAAGA-3' |
5'-TGCCATGAGACATTAGCACCAGGA-3' |
| Gabrp |
5'-AGTCCTGCCTGAGATCTGGA-3' |
5'-AAGGTTGAAGGAGGAATCACC-3' |
| Chn2 |
5'-AGTCCGGTGTTCAGATTGTGGGTT-3' |
5'-TTGTGAGCTTTGACAAGCGTGGTG-3' |
| Igf1R |
5'-GTGGGGGCTCGTGTTTCTC-3' |
5'-GATCACCGTGCAGTTTTCCA-3' |
| Rassf9 |
5'-AGCCAAGGAGTTCAGCTCACTTCA-3' |
5'-AGCCTCAGCTTCCTTTCCTCTGTT-3' |
| Clca3p |
5'-AGAATGAACCCACCACGTCCTGAA-3' |
5'-TCAGATTCACCAGGTTCTGCCCTT-3' |
| Muc5Ac |
5'-AGAATATCTTTCAGGACCCCTGCT-3' |
5'-ACACCAGTGCTGAGCATACTTTT-3' |
| Muc5B |
5'-CATCCATCCCATTTCTACCACAA-3' |
5'-AGGCAACATAGAGTTGCTTTTGG-3' |
Statistical analysis
Data from three or more independent experiments were
collected and analyzed and results were presented as the mean ±
standard error of the mean. The significance of differences was
assessed using Student's t-test or ANOVA. One-way ANOVA was
followed by Tukey's and Bonferroni post hoc tests. P<0.05 was
considered to indicate statistical significance.
Results
Transcriptome analysis identifies Gabrp
expression in mouse Club cells. Transcriptome analysis was
performed to verify Gabrp expression in Club cells and compare it
to the expression in alveolar type 2 (AT2) cells, considered to be
alveolar epithelial progenitor cells in mouse lungs. Mouse Club and
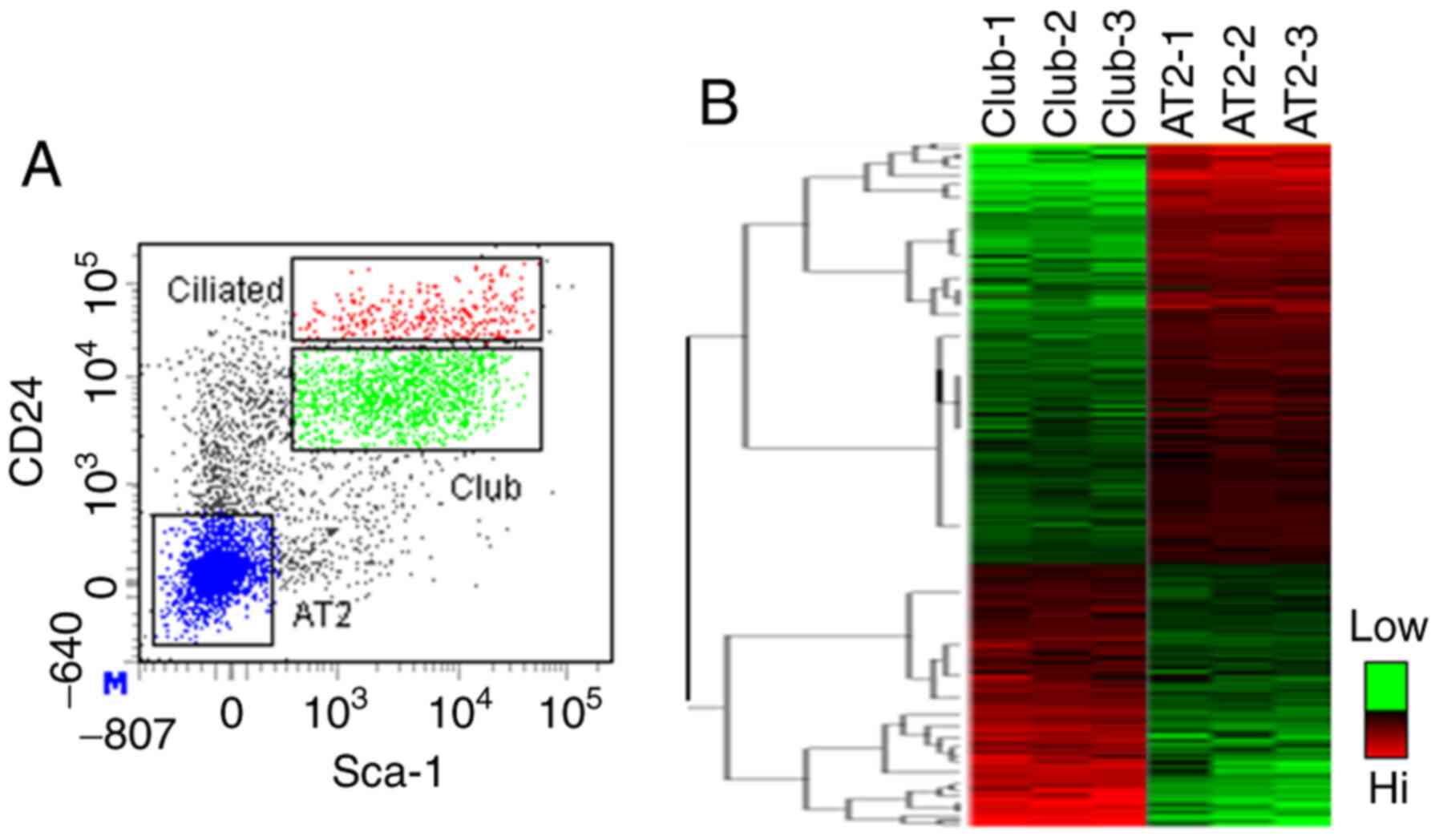
AT2 cells were fractionated by FACS (Fig. 1A) (19) and stromal cells, endothelial cells
and hematopoietic cells were excluded from the analysis by surface
staining with CD34, CD31 and CD45 antibodies, respectively. The
epithelial (EpCAM+) population was further segregated
into Club cells (CD24+Sca-1+), ciliated cells
(CD24hiSca-1+) and AT2 cells
(CD24-Sca-1-). Microarray analysis of Club
and AT2 cells generated unique patterns of gene expression for each
cell fraction (Fig. 1B), with 558
transcripts enriched within Club cells and 1,006 transcripts
enriched within AT2 cells. The distinct gene signatures between the
Club cells and AT2 cells may reflect different signaling pathways
that regulate their functions. Ingenuity analysis indicated that
Wnt/β-catenin signaling and phosphatase and tensin homolog (PTEN)
signaling were active in Club cells (Table II), which is consistent with
published data (27,28). Furthermore, KEGG pathway analysis
indicated that interleukin (IL)-17 signaling and aryl hydrocarbon
receptor (AHR) signaling may also be important in regulating Club
cell function. In addition, studies report metabolic pathways
associated with xenobiotics and glutathione in Club cells, which is
consistent with their preferential metabolism of naphthalene
(29,30). Other metabolic pathways in Club
cells include fatty acid metabolism, glycolysis/gluconeogenesis,
arachidonic acid metabolism, propanoate metabolism, pyruvate
metabolism, tryptophan metabolism and N-glycan biosynthesis
(Table II). By contrast, signaling
pathways in AT2 cells include the JAK/STAT, insulin receptor
signaling, interferon and granulocyte-macrophage colony-stimulating
factor pathways, all of which reportedly regulate AT2 cell
specification, proliferation and differentiation during homeostasis
or following injury (31-34).
Other potential but novel signaling pathways associated with AT2
cells include p21-activated kinase signaling and cell division
cycle 42 signaling, which are closely related to cell cycle
progression and cell transformation (35). Metabolic pathways in AT2 cells
included steroid biosynthesis, liver X receptor-retinoid X receptor
activation, selenoamino acid metabolism, sphingolipid metabolism
and methionine metabolism.
| Table IITop canonical pathways potentially
associated with mouse Club or AT2 cells. |
Table II
Top canonical pathways potentially
associated with mouse Club or AT2 cells.
| A, Club cells |
|---|
| Rank | Pathways |
|---|
| 1 | Glutathione
metabolism |
| 2 | Metabolism of
xenobiotics by Cytochrome P450 |
| 3 | LPS/IL-1-mediated
inhibition of RXR function |
| 4 | NRF2-mediated
oxidative stress response |
| 5 | Acute phase
response signaling |
| 6 | AHR signaling |
| 7 | Fatty acid
metabolism |
| 8 |
Glycolysis/Gluconeogenesis |
| 9 | Xenobiotic
metabolism signaling |
| 10 | Propanoate
metabolism |
| 11 | Arachidonic acid
metabolism |
| 12 | Pyruvate
metabolism |
| 13 | Role of
osteoblasts, osteoclasts, and chondrocytes in rheumatoid
arthritis |
| 14 | Tryptophan
metabolism |
| 15 | Human embryonic
stem cell pluripotency |
| 16 | Wnt/β-catenin
signaling |
| 17 | IL-17A signaling in
airway cells |
| 18 | N-Glycan
biosynthesis |
| 19 | PTEN signaling |
| 20 | IL-17
signaling |
| B, AT2 cells |
| Rank | Pathways |
| 1 | Steroid
biosynthesis |
| 2 | JAK/STAT
signaling |
| 3 | LXR/RXR
activation |
| 4 | IL-4 signaling |
| 5 | Selenoamino acid
metabolism |
| 6 | Breast cancer
regulation by stathmin-1 |
| 7 | Graft-versus-host
disease signaling |
| 8 | Sphingolipid
metabolism |
| 9 | Prolactin
signaling |
| 10 | Insulin receptor
signaling |
| 11 | PAK signaling |
| 12 | Methionine
metabolism |
| 13 | Melanocyte
development and pigmentation signaling |
| 14 | PI3K signaling in B
lymphocytes |
| 15 | Cdc42
signaling |
| 16 | Axonal guidance
signaling |
| 17 | Interferon
signaling |
| 18 | Acute phase
response signaling |
| 19 | Autoimmune thyroid
disease |
| 20 | GM-CSF
signaling |
Transcripts enriched in Club cells included palate,
lung and nasal epithelium clones, flavin-containing monooxygenase 3
(Fmo3), ceruloplasmin (Cp), regenerating islet-derived 3 gamma
(Reg3g) and Muc5B, which have previously been demonstrated to be
expressed in Club cells (Table
III) (4,36-38).
Novel transcripts that may be potential markers for Club cells
included Ras association domain family 9 (Rassf9), caspase 4
(Casp4), chimerin 2 (Chn2), sulfotransferase family 1D, member 1
(Sult1d1), insulin-like growth factor I receptor (Igf1R),
iodotyrosine deiodinase (Iyd) and Gabrp (Tables III and SI). By contrast, transcripts enriched in
AT2 cells included brain-expressed gene 1, EGF-like-domain,
multiple 6 (Egfl6), ets variant gene 5 (Etv5), ectonucleoside
triphosphate diphosphohydrolase 1, tubulointerstitial nephritis
antigen, tetraspanin 8, leucine-rich repeat LGI family member 2,
neuritin 1 and adenylate cyclase 7 (Tables III and SI). Transcriptome analysis suggested that
Gabrp is highly expressed in mouse Club cells rather than AT2
cells.
| Table IIITop (1-20) transcripts enriched in
Club and AT2 progenitor cells. |
Table III
Top (1-20) transcripts enriched in
Club and AT2 progenitor cells.
| A, Club
(Sca-1+AFhi) |
|---|
| Probe ID | Gene ID | Gene symbol | P-value | Fold change |
|---|
| Club vs. AT2 | | | | |
|
1420347_at | 18843 | Plunc | 0.025 | 127.4 |
|
1427942_at | 237504 | Rassf9 | 0.016 | 96.4 |
|
1449525_at | 14262 | Fmo3 | 0.020 | 77.4 |
|
1456404_at | 100048332 ///
23794 | Adamts5 ///
LOC100048332 | 0.001 | 74.9 |
|
1419699_at | 68662 | Scgb3a1 | 0.024 | 74.0 |
|
1434449_at | 11829 | Aqp4 | 0.004 | 72.5 |
|
1417496_at | 12870 | Cp | 0.036 | 71.5 |
|
1448872_at | 19695 | Reg3g | 0.016 | 62.1 |
|
1449591_at | 12363 | Casp4 | 0.018 | 59.2 |
|
1428574_a_at | 69993 | Chn2 | 0.004 | 58.2 |
|
1429514_at | 67916 | Ppap2b | 0.001 | 57.1 |
|
1424647_at | 216643 | Gabrp | 0.021 | 55.3 |
|
1427626_at | 74180 | Muc5b | 0.004 | 49.6 |
|
1459894_at | 544963 | Iqgap2 | 0.007 | 42.1 |
|
1457589_at | 270120 | Fat3 | 0.012 | 42.1 |
|
1426147_s_at | 58187 | Cldn10 | 0.011 | 39.5 |
|
1454764_s_at | 105727 | Slc38a1 | 0.024 | 38.3 |
|
1453782_at | 70678 | 3021401C12Rik | 0.017 | 37.0 |
|
1424842_a_at | 231532 | Arhgap24 | 0.006 | 35.0 |
|
1443832_s_at | 20324 | Sdpr | 0.002 | 34.0 |
| B, AT2
(Sca-1-) |
| Probe ID | Gene ID | Gene symbol | P-value | Fold change |
| Club vs. AT2 | | | | |
|
1448595_a_at | 19716 | Bex1 | 0.011 | -112.5 |
|
1419332_at | 54156 | Egfl6 | 0.010 | -105.0 |
|
1428393_at | 68404 | Nrn1 | 0.038 | -80.8 |
|
1453586_at | 12495 | Entpd1 | 0.002 | -73.2 |
|
1450082_s_at | 104156 | Etv5 | 0.041 | -64.9 |
|
1455995_at | 215821 | D10Bwg1379e | 0.00004 | -61.4 |
|
1460465_at | 68169 | A930038C07Rik | 0.038 | -59.5 |
|
1424649_a_at | 216350 | Tspan8 | 0.007 | -59.4 |
|
1450065_at | 11513 | Adcy7 | 0.002 | -58.7 |
|
1450648_s_at | 14961 | H2-Ab1 | 0.001 | -55.1 |
|
1450213_at | 29863 | Pde7b | 0.003 | -53.4 |
|
1440147_at | 246316 | Lgi2 | 0.002 | -53.0 |
|
1449421_a_at | 246133 | Kcne2 | 0.011 | -50.9 |
|
1418402_at | 100045780 ///
11492 | Adam19 ///
LOC100045780 | 0.002 | -50.8 |
|
1417111_at | 17155 | Man1a | 0.033 | -50.4 |
|
1456307_s_at | 11513 | Adcy7 | 0.024 | -49.8 |
|
1418403_at | 100045780 ///
11492 | Adam19 ///
LOC100045780 | 0.001 | -49.5 |
|
1416200_at | 77125 | Il33 | 0.025 | -47.2 |
|
1452106_at | 114249 | Npnt | 0.044 | -46.3 |
|
1431808_a_at | 16427 | Itih4 | 0.006 | -43.7 |
Gabrp expression is diminished during
naphthalene-induced Club cell injury
Validation of gene expression by RT-qPCR indicated
that in addition to known markers, including Scgb1a1, Fmo3 and Cp,
Gabrp was also highly expressed in sorted mouse Club cells
(Fig. 2). Ciliated cells also
express Gabrp, suggesting its potential role in ciliated cell
differentiation. Similar to Gabrp, Chn2, Igf1R and Reg3g were also
expressed in ciliated cells (Fig.
2). Although significant levels of Iyd, Sult1d1 and Rassf9 were
also observed in Club cells, their expression was not specific to
Club cells (Fig. 2).
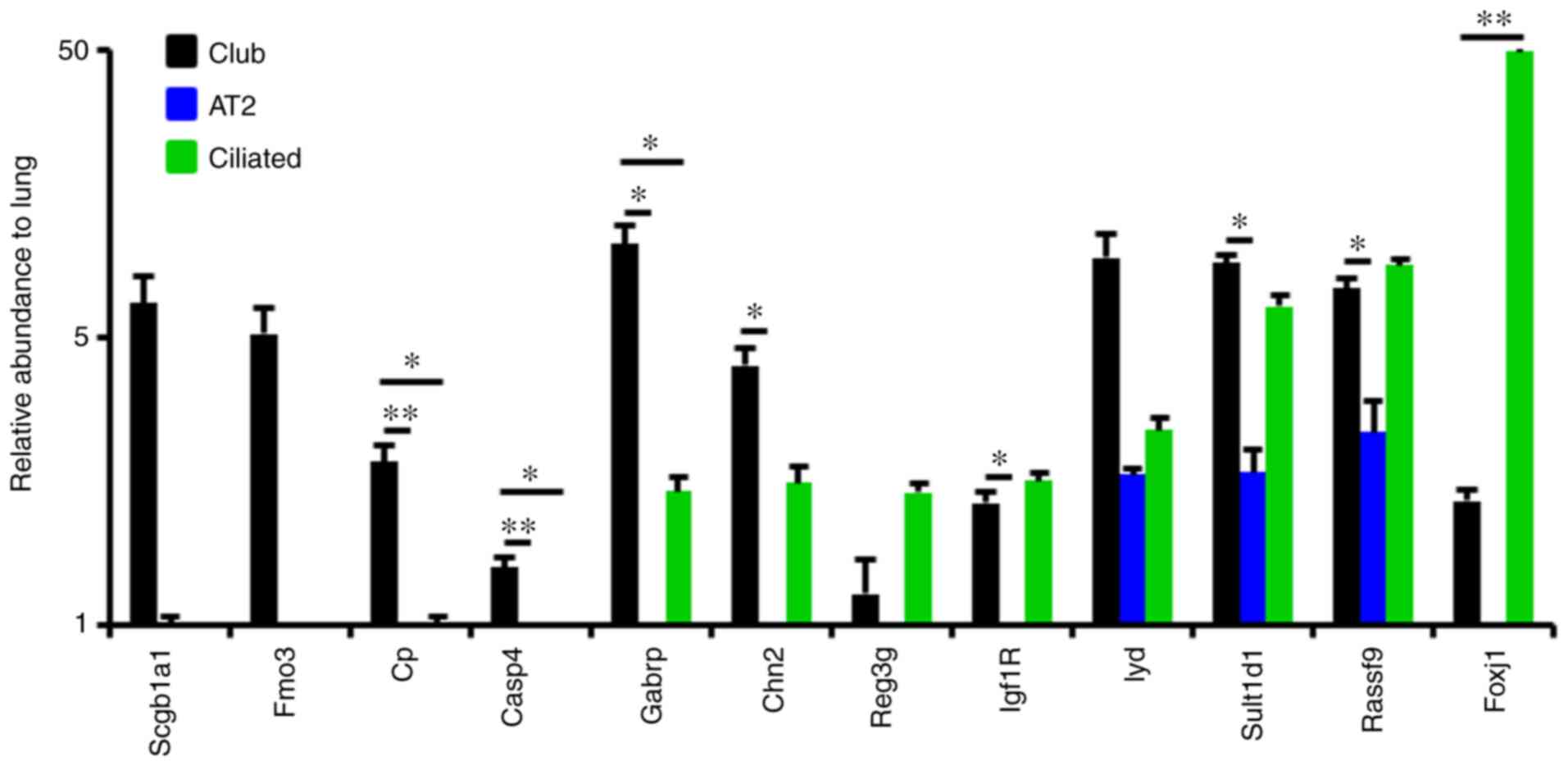 | Figure 2Gabrp is highly expressed in mouse
Club cells. The expression of Gabrp and other markers in Club, AT2
and ciliated cells was determined by reverse
transcription-quantitative PCR analysis. *P<0.05;
**P<0.01. AT2, alveolar type 2; Scgb1a1,
secretoglobin family 1A member 1; Fmo3, Flavin-containing
monooxygenase 3; Cp, ceruloplasmin; caspase 4, Casp4; Gabrp,
gamma-aminobutyric acid A receptor pi; Chn2, chimerin 2; Reg3g,
regenerating islet-derived 3 gamma; Igf1R, insulin-like growth
factor I receptor; Iyd, iodotyrosine deiodinase; Sult1d1,
sulfotransferase family 1D, member 1; Rassf9, Ras association
domain family 9; FoxJ1, forkhead box family member J1. |
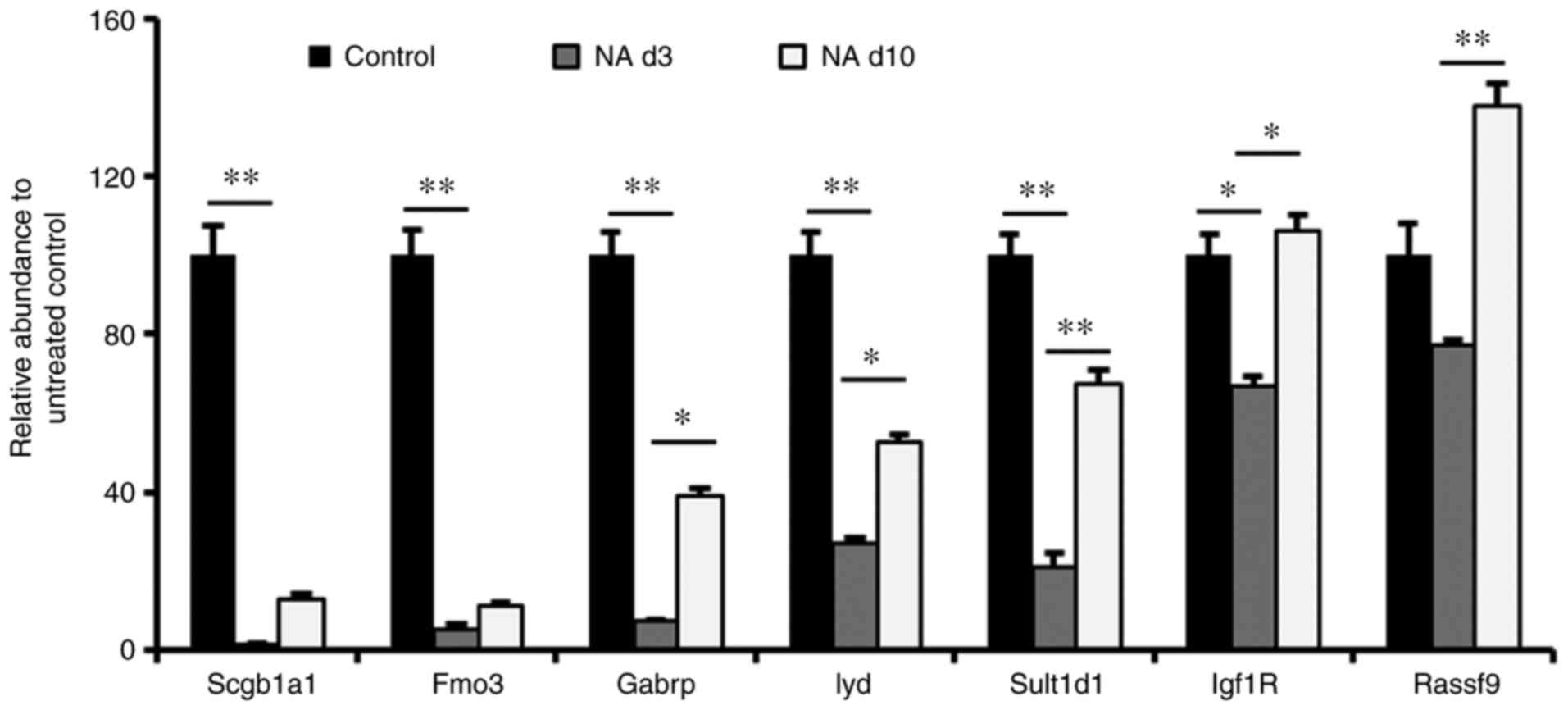
To verify these results, a Club cell-ablated
naphthalene-injury model in mice was established. Based on studies
from our group and others (18,39),
naphthalene causes massive loss of Club cells by day 3; however, by
day 10, significant recovery of Club cells is observed. Therefore,
a window from day 3 to 10 was used to confirm Gabrp expression in
Club cells. Gene-expression analysis of total mouse lungs
demonstrated that the expression of Fmo3 and Gabrp had decreased on
day 3 after naphthalene injury and then began to recover with time,
similar to Scgb1a1 expression kinetics (Fig. 3). Furthermore, Lyd and Sult1d1
expression were reduced to a lesser extent relative to Gabrp
expression (Fig. 3). These results
verified the specific expression of Gabrp in mouse Club cells.
Inhibition of Gabrp signaling blocks
Club cell proliferation and goblet cell differentiation
It was then examined whether Gabrp-mediated signal
transduction regulates the proliferation and differentiation of
Club cells. A previously published organoid culture method was
adopted, which includes a transforming growth factor (TGF)-β
receptor inhibitor (SB431542) that allows for epithelial clonal
expansion. Scgb1a1-tagged Club cells were isolated from
Scgb1a1-mT/mG mice after tamoxifen treatment and maintained in
Matrigel cultures for 2 weeks in either the absence or presence of
the Gabrp-signaling antagonist BMI. A BMI concentration of 20 µg/ml
was previously demonstrated to be sufficient for inhibiting Gabrp
signaling in airway cells (20-23).
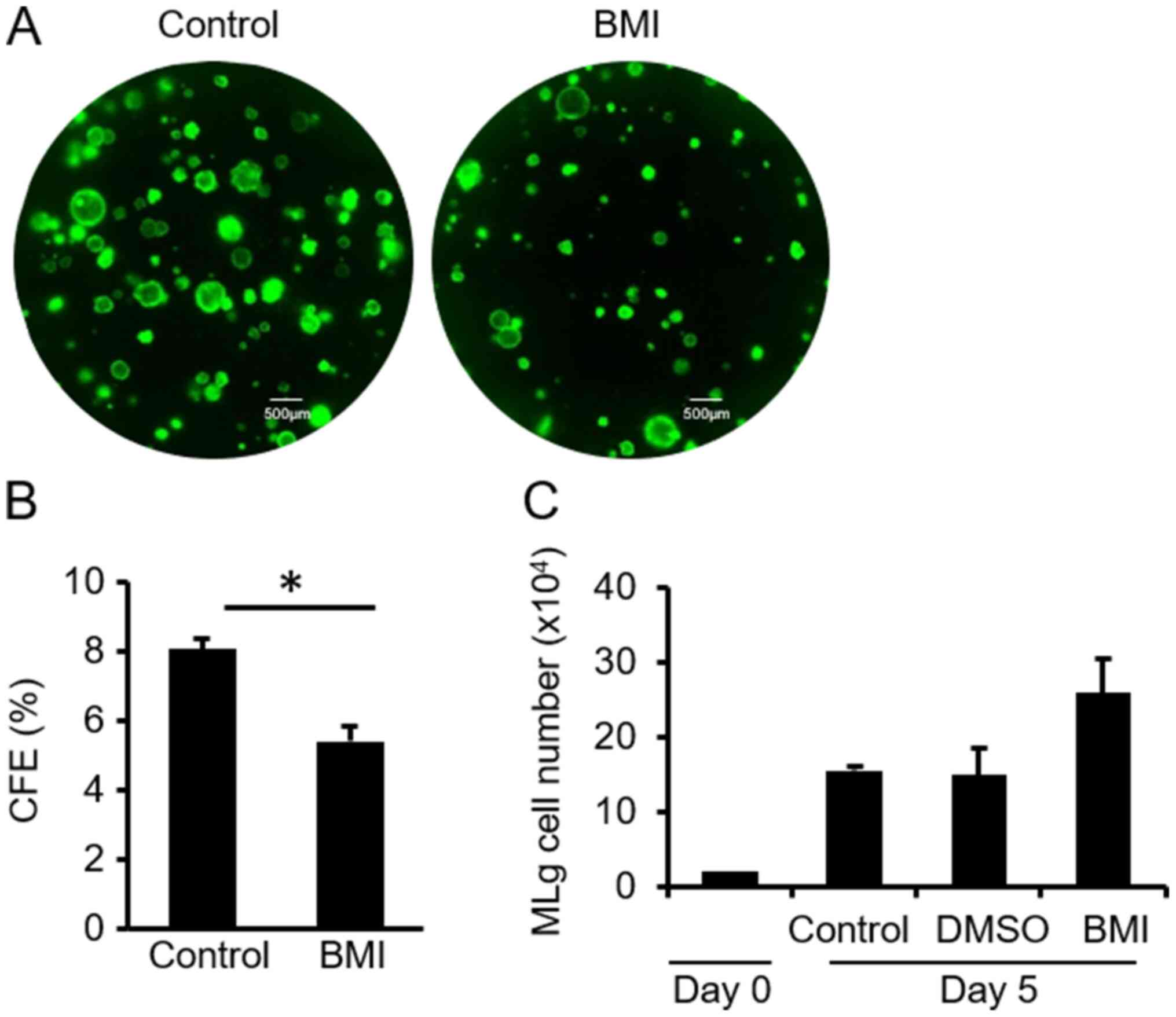
Compared to the control group, the CFE of Club cells was
significantly reduced by ~30% in culture medium supplemented with
BMI (Fig. 4A and B) and BMI did not alter the growth of MLg
cells (Fig. 4C. These results
suggested that BMI may act directly on Club cells.
To further evaluate the role of Gabrp-mediated
signaling in Club cell differentiation, Club cells were maintained
for 10 days in culture medium containing SB431542 and then switched
to culture medium without SB431542. RT-qPCR analysis of total RNA
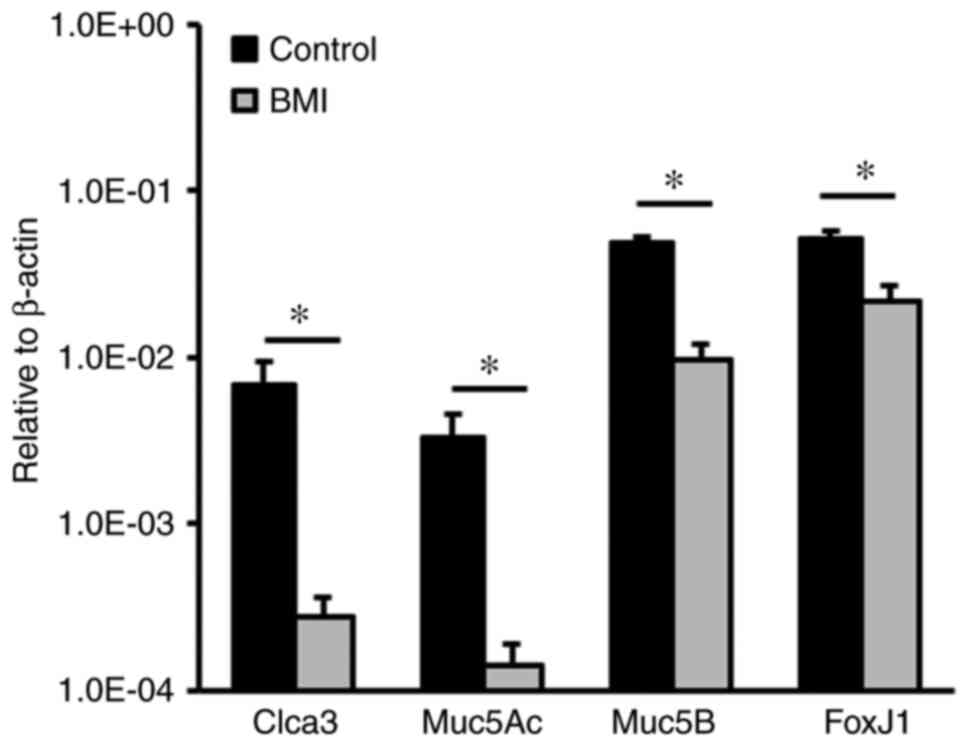
extracted from epithelial organoid cultures indicated that the mRNA
levels of Clca3, Muc5Ac and Muc5B, markers for goblet cells, were
significantly decreased in the presence of BMI as compared with
those in the control culture (Fig.
5). Of note, the mRNA level of forkhead box protein J1 was also
lower in the BMI-treated group than in the control group,
suggesting that blocking Gabrp-mediated signaling inhibits ciliated
cell differentiation. These results indicated that Gabrp-mediated
signal transduction modulates Club cell differentiation into goblet
and ciliated cells.
Discussion
Mucus overproduction is a major characteristic of
bronchial asthma and mucus-producing goblet cells are thought to be
generated by airway Club progenitor cells (40-42).
Club cells are capable of maintaining the pulmonary epithelium at
the steady state and repairing the epithelium after injury
(18). The reparative function of
Club cells is regulated by their transcriptomes; therefore, in the
present study, a transcriptome analysis of Club cells was
performed. The results indicated that Gabrp was highly expressed in
mouse Club cells, which was validated using both RT-qPCR and in
vivo in a mouse model of naphthalene-induced injury.
Furthermore, an organoid culture experiment of mouse Club cells
suggested that BMI inhibition of Gabrp abrogated Club cell
proliferation and goblet cell differentiation. These results
suggested that GABA/Gabrp signaling may have a positive role in
Club cell function.
A recent study indicated that GABA is able to induce
goblet cell hyperplasia in the lungs of primate models and further
contribute to the overproduction of airway mucus, leading to the
worsening of asthma (16). GABA
hypersecretion is necessary for the overproduction of Muc5ac
(43) and a study by Xiang et
al (44) demonstrated that
intravenous injection of the selective GABAA receptor inhibitor BMI
suppressed goblet cell hyperplasia and ovalbumin-induced mucus
overproduction. In the present study, blocking the GABA/Gabrp
signaling pathway with BMI inhibited Club cell differentiation into
goblet cells. This result is in agreement with a previous study
indicating that GABA contributes to mucus production (44). In addition, cilia are reportedly
disrupted in the airways of patients suffering from asthma
(45), and in the present study, it
was observed that ciliated cell differentiation was promoted by
GABA signaling.
Lung epithelial damage is the initial source of
airway inflammation (46). The
present study suggested that GABA/Gabrp signaling positively
regulated Club cell proliferation, which sustains the pool of Club
cells in the airways. In this study, Gabrp enhanced the
proliferation of mouse Club cells. The present results support the
findings of Xiang et al (44) and provide a more specific cellular
mechanism for their observations. As GABA receptors allow the flow
of negatively charged Cl- ions across the cell membrane
(47), future studies may examine
the effects of ion channels on the regulation of Club cell
function. The overproduction of airway mucus and the damage to
airway epithelia contribute to the chronic airway inflammation
occurring in asthma. Therefore, this suggests that GABA/Gabrp
signaling may be involved in asthmatic inflammation through
multiple mechanisms.
As airway epithelial progenitor cells, Club cells
have an important role in host defense by secreting proteins and
regenerating airway epithelium after lung injury (48,49).
Deregulation of Club cells is closely related to the pathogenesis
of diseases involving the conducting airways. Therefore,
understanding the signaling pathways that govern Club cell function
is critical. Transcriptome analysis of mouse Club cells identified
several potential pathways previously demonstrated to be involved
in mouse Club cell function, including PTEN and β-catenin
signaling. PTEN acts as a negative regulator of Club cell function,
with deletion of PTEN using Nkx2.1-Cre causing both epithelial
hyperplasia in the embryonic lung and Club cell expansion in the
adult lung owing to the increased proliferative index of Club cells
(50). In addition, PTEN silencing
contributes to airway remodeling and induces airway smooth muscle
cell proliferation in asthma (51).
These data suggest a relationship between PTEN and β-catenin
signaling in the regulation of ontogeny. The regeneration of Club
cells may be more complex than previously assumed. Consistent with
previous studies (52,53), the present pathway analysis
indicated that IL-17 signaling is involved in Club cell function.
Conditional loss of the TGF-β type 1 receptor using Gata5-Cre in
the embryonic lung epithelium blocked Club cell differentiation
(54). However, previous in
vitro culture data demonstrated that blockade of TGF-β
signaling with the selective inhibitor SB431542 promoted airway
progenitor cell proliferation (19).
Pathway analysis of Club cells suggested several
other novel signaling pathways that may regulate Club cell
proliferation and/or differentiation. Other potential pathways
included AHR signaling, acute phase response signaling and nuclear
factor, erythroid 2 like 2-mediated oxidative stress response
signaling. Of note, transcriptome analysis proposed a number of
metabolic pathways that may affect mouse Club cell function,
including glutathione metabolism, fatty acid metabolism and
pyruvate metabolism. Future studies are required to investigate the
impact of metabolism on Club cell function at the steady state and
in the pathology of asthma.
In conclusion, GABA induces hyperplasia of goblet
cells. The present results revealed that Gabrp is expressed in
mouse airway progenitor cells and that the Gabrp inhibitor BMI
reduced their proliferation and goblet cell differentiation.
Therefore, targeting GABA/Gabrp signaling may represent a promising
strategy for treating goblet cell hyperplasia in bronchial
asthma.
Supplementary Material
Top (21-50) transcripts enriched in
Club and AT2 progenitor cells.
Acknowledgements
Not applicable.
Funding
Funding: This study was supported by the National Natural
Science Foundation of China (grant nos. 81773394, 81970001 and
82070001), the Natural Science Foundation of Tianjin (grant nos.
18ZXDBSY00150, 19JCZDJC33600 and 20JCQNJC01790), Tianjin Health
Science and Technology Project (grant no. 2020XKZ02) and the
Science and Technology Fund of Tianjin Haihe Hospital (grant nos.
HHYY-202008 and HHYY-202012).
Availability of data and materials
The datasets generated and/or analyzed during the
current study are not publicly available due to the transcriptome
data being needed for other research but are available from the
corresponding author on reasonable request.
Authors' contributions
AW, QZ, YW, XL, LL and HC designed the experiments
and wrote the manuscript with input from co-authors. AW and QZ
performed the experiments. XL, KL, YL and JW preformed the animal
studies. All authors discussed the results and commented on the
manuscript. AW and HC confirmed the authenticity of all the raw
data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All mice were treated with strict adherence to the
protocol (no. 2020HHSQKT-001) that was approved by the Haihe
Hospital Animal Care and Use Committee.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gon Y and Hashimoto S: Role of airway
epithelial barrier dysfunction in pathogenesis of asthma. Allergol
Int. 67:12–17. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Lam M, Lamanna E and Bourke JE: Regulation
of airway smooth muscle contraction in health and disease. Adv Exp
Med Biol. 1124:381–422. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Murrison LB, Brandt EB, Myers JB and
Hershey GKK: Environmental exposures and mechanisms in allergy and
asthma development. J Clin Invest. 129:1504–1515. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Evans CM, Williams OW, Tuvim MJ, Nigam R,
Mixides GP, Blackburn MR, DeMayo FJ, Burns AR, Smith C, Reynolds
SD, et al: Mucin is produced by clara cells in the proximal airways
of antigen-challenged mice. Am J Respir Cell Mol Biol. 31:382–394.
2004.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Reynolds SD, Reynolds PR, Pryhuber GS,
Finder JD and Stripp BR: Secretoglobins SCGB3A1 and SCGB3A2 define
secretory cell subsets in mouse and human airways. Am J Respir Crit
Care Med. 166:1498–1509. 2002.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Singh G and Katyal SL: Clara cell
proteins. Ann N Y Acad Sci. 923:43–58. 2000.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Tam A, Wadsworth S, Dorscheid D, Man SF
and Sin DD: The airway epithelium: More than just a structural
barrier. Ther Adv Respir Dis. 5:255–273. 2011.PubMed/NCBI View Article : Google Scholar
|
|
8
|
El-Badrawy MK, Shalabi NM, Mohamed MA,
Ragab A and Abdelwahab HW: Stem cells and lung regeneration. Int J
Stem Cells. 9:31–35. 2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Porotto M, Ferren M, Chen YW, Siu Y,
Makhsous N, Rima B, Briese T, Greninger AL, Snoeck HW and Moscona
A: Authentic modeling of human respiratory virus infection in human
pluripotent stem cell-derived lung organoids. mBio. 10:e00723–19.
2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Sacco O, Silvestri M, Sabatini F, Sale R,
Defilippi AC and Rossi GA: Epithelial cells and fibroblasts:
Structural repair and remodelling in the airways. Paediatr Respir
Rev. 5 (Suppl A):S35–S40. 2004.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Samitas K, Carter A, Kariyawasam HH and
Xanthou G: Upper and lower airway remodelling mechanisms in asthma,
allergic rhinitis and chronic rhinosinusitis: The one airway
concept revisited. Allergy. 73:993–1002. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
McCauley KB, Alysandratos KD, Jacob A,
Hawkins F, Caballero IS, Vedaie M, Yang W, Slovik KJ, Morley M,
Carraro G, et al: Single-cell transcriptomic profiling of
pluripotent stem cell-derived SCGB3A2+ airway epithelium. Stem Cell
Reports. 10:1579–1595. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Parekh KR, Nawroth J, Pai A, Busch SM,
Senger CN and Ryan AL: Stem cells and lung regeneration. Am J
Physiol Cell Physiol. 319:C675–C693. 2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Barkauskas CE, Chung MI, Fioret B, Gao X,
Katsura H and Hogan BL: Lung organoids: Current uses and future
promise. Development. 144:986–997. 2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Boers JE, Ambergen AW and Thunnissen FB:
Number and proliferation of clara cells in normal human airway
epithelium. Am J Respir Crit Care Med. 159:1585–1591.
1999.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Barrios J, Kho AT, Aven L, Mitchel JA,
Park JA, Randell SH, Miller LA, Tantisira KG and Ai X: Pulmonary
neuroendocrine cells secrete gamma-aminobutyric acid to induce
goblet cell hyperplasia in primate models. Am J Respir Cell Mol
Biol. 60:687–694. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Song H, Yao E, Lin C, Gacayan R, Chen MH
and Chuang PT: Functional characterization of pulmonary
neuroendocrine cells in lung development, injury, and
tumorigenesis. Proc Natl Acad Sci USA. 109:17531–17536.
2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Li K, Li M, Li W, Yu H, Sun X, Zhang Q, Li
Y, Li X, Li Y, Abel ED, et al: Airway epithelial regeneration
requires autophagy and glucose metabolism. Cell Death Dis.
10(875)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Teisanu RM, Chen H, Matsumoto K, McQualter
JL, Potts E, Foster WM, Bertoncello I and Stripp BR: Functional
analysis of two distinct bronchiolar progenitors during lung injury
and repair. Am J Respir Cell Mol Biol. 44:794–803. 2011.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Barolet AW, Li A, Liske S and Morris ME:
Antagonist actions of bicuculline methiodide and picrotoxin on
extrasynaptic gamma-aminobutyric acid receptors. Can J Physiol
Pharmacol. 63:1465–1470. 1985.PubMed/NCBI View
Article : Google Scholar
|
|
21
|
Hussin AT, Boychuk JA, Brown AR, Pittman
QJ and Teskey GC: Intracortical microstimulation (ICMS) activates
motor cortex layer 5 pyramidal neurons mainly transsynaptically.
Brain Stimul. 8:742–750. 2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Johnson SW and Seutin V: Bicuculline
methiodide potentiates NMDA-dependent burst firing in rat dopamine
neurons by blocking apamin-sensitive Ca2+-activated K+ currents.
Neurosci Lett. 231:13–16. 1997.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Uva L, Boido D, Avoli M, de Curtis M and
Lévesque M: High-frequency oscillations and seizure-like discharges
in the entorhinal cortex of the in vitro isolated guinea pig brain.
Epilepsy Res. 130:21–26. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Chen H, Matsumoto K, Brockway BL, Rackley
CR, Liang J, Lee JH, Jiang D, Noble PW, Randell SH, Kim CF and
Stripp BR: Airway epithelial progenitors are region specific and
show differential responses to bleomycin-induced lung injury. Stem
Cells. 30:1948–1960. 2012.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Chen H, Sun X, Chi R, Li X, Feng J, Wu J,
Ning W, Liu Z and Wu Q: Glucocorticoid dexamethasone regulates the
differentiation of mouse conducting airway epithelial progenitor
cells. Steroids. 80:44–50. 2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Li X, Wang YM, Wang A, Li Y, Wu Q and Chen
HY: Sex-determining region Y-box 2-positive alveolar cells are
responsive to bleomycin-induced lung injury. Chin Med J (Engl).
134:234–236. 2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Davé V, Wert SE, Tanner T, Thitoff AR,
Loudy DE and Whitsett JA: Conditional deletion of Pten causes
bronchiolar hyperplasia. Am J Respir Cell Mol Biol. 38:337–345.
2008.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Reynolds SD, Zemke AC, Giangreco A,
Brockway BL, Teisanu RM, Drake JA, Mariani T, Di PY, Taketo MM and
Stripp BR: Conditional stabilization of beta-catenin expands the
pool of lung stem cells. Stem Cells. 26:1337–1346. 2008.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Carratt SA, Kovalchuk N, Ding X and Van
Winkle LS: Metabolism and lung toxicity of inhaled naphthalene:
Effects of postnatal age and sex. Toxicol Sci. 170:536–548.
2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Plopper CG, Van Winkle LS, Fanucchi MV,
Malburg SR, Nishio SJ, Chang A and Buckpitt AR: Early events in
naphthalene-induced acute clara cell toxicity. II. Comparison of
glutathione depletion and histopathology by airway location. Am J
Respir Cell Mol Biol. 24:272–281. 2001.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kang BH, Manderschied BD, Huang YC, Crapo
JD and Chang LY: Contrasting response of lung parenchymal cells to
instilled TNF alpha and IFN gamma: The inducibility of specific
cell ICAM-1 in vivo. Am J Respir Cell Mol Biol. 15:540–550.
1996.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Sturrock A, Seedahmed E, Mir-Kasimov M,
Boltax J, McManus ML and Paine R III: GM-CSF provides autocrine
protection for murine alveolar epithelial cells from
oxidant-induced mitochondrial injury. Am J Physiol Lung Cell Mol
Physiol. 302:L343–L351. 2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Sugahara K, Freidenberg GR and Mason RJ:
Insulin binding and effects on glucose and transepithelial
transport by alveolar type II cells. Am J Physiol. 247:C472–C477.
1984.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Xu Y, Ikegami M, Wang Y, Matsuzaki Y and
Whitsett JA: Gene expression and biological processes influenced by
deletion of Stat3 in pulmonary type II epithelial cells. BMC
Genomics. 8(455)2007.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Bokoch GM: Biology of the p21-activated
kinases. Annu Rev Biochem. 72:743–781. 2003.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Gakhar L, Bartlett JA, Penterman J,
Mizrachi D, Singh PK, Mallampalli RK, Ramaswamy S and McCray PB Jr:
PLUNC is a novel airway surfactant protein with anti-biofilm
activity. PLoS One. 5(e9098)2010.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Sountoulidis A, Liontos A, Nguyen HP,
Firsova AB, Fysikopoulos A, Qian X, Seeger W, Sundström E, Nilsson
M and Samakovlis C: SCRINSHOT enables spatial mapping of cell
states in tissue sections with single-cell resolution. PLoS Biol.
18(e3000675)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Zemke AC, Snyder JC, Brockway BL, Drake
JA, Reynolds SD, Kaminski N and Stripp BR: Molecular staging of
epithelial maturation using secretory cell-specific genes as
markers. Am J Respir Cell Mol Biol. 40:340–348. 2009.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Manzo ND, Foster WM and Stripp BR:
Amphiregulin-dependent mucous cell metaplasia in a model of
nonallergic lung injury. Am J Respir Cell Mol Biol. 47:349–357.
2012.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Lambrecht BN and Hammad H: The airway
epithelium in asthma. Nat Med. 18:684–692. 2012.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Reader JR, Tepper JS, Schelegle ES,
Aldrich MC, Putney LF, Pfeiffer JW and Hyde DM: Pathogenesis of
mucous cell metaplasia in a murine asthma model. Am J Pathol.
162:2069–2078. 2003.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Chen G, Korfhagen TR, Xu Y, Kitzmiller J,
Wert SE, Maeda Y, Gregorieff A, Clevers H and Whitsett JA: SPDEF is
required for mouse pulmonary goblet cell differentiation and
regulates a network of genes associated with mucus production. J
Clin Invest. 119:2914–2924. 2009.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Barrios J, Patel KR, Aven L, Achey R,
Minns MS, Lee Y, Trinkaus-Randall VE and Ai X: Early life
allergen-induced mucus overproduction requires augmented neural
stimulation of pulmonary neuroendocrine cell secretion. FASEB J.
31:4117–4128. 2017.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Xiang YY, Wang S, Liu M, Hirota JA, Li J,
Ju W, Fan Y, Kelly MM, Ye B, Orser B, et al: A GABAergic system in
airway epithelium is essential for mucus overproduction in asthma.
Nat Med. 13:862–867. 2007.PubMed/NCBI View
Article : Google Scholar
|
|
45
|
Fireman P: Understanding asthma
pathophysiology. Allergy Asthma Proc. 24:79–83. 2003.PubMed/NCBI
|
|
46
|
Khair OA, Davies RJ and Devalia JL:
Bacterial-induced release of inflammatory mediators by bronchial
epithelial cells. Eur Respir J. 9:1913–1922. 1996.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Jin N, Guo Y, Sun P, Bell A, Chintagari
NR, Bhaskaran M, Rains K, Baviskar P, Chen Z, Weng T and Liu L:
Ionotropic GABA receptor expression in the lung during development.
Gene Expr Patterns. 8:397–403. 2008.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Malvin NP, Kern JT, Liu TC, Brody SL and
Stappenbeck TS: Autophagy proteins are required for club cell
structure and function in airways. Am J Physiol Lung Cell Mol
Physiol. 317:L259–L270. 2019.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Shafiquzzaman M, Biswas S, Li P, Mishina
Y, Li B and Liu H: The noncanonical BMP signaling pathway plays an
important role in club cell regeneration. Stem Cells. 38:437–450.
2020.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Tiozzo C, De Langhe S, Yu M, Londhe VA,
Carraro G, Li M, Li C, Xing Y, Anderson S, Borok Z, et al: Deletion
of Pten expands lung epithelial progenitor pools and confers
resistance to airway injury. Am J Respir Crit Care Med.
180:701–712. 2009.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Wen X, Yan J, Han XR, Zheng GH, Tang R,
Liu LF, Wu DM, Lu J and Zheng YL: PTEN gene silencing contributes
to airway remodeling and induces airway smooth muscle cell
proliferation in mice with allergic asthma. J Thorac Dis.
10:202–211. 2018.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Kreindler JL, Bertrand CA, Lee RJ, Karasic
T, Aujla S, Pilewski JM, Frizzell RA and Kolls JK: Interleukin-17A
induces bicarbonate secretion in normal human bronchial epithelial
cells. Am J Physiol Lung Cell Mol Physiol. 296:L257–L266.
2009.PubMed/NCBI View Article : Google Scholar
|
|
53
|
McAllister F, Henry A, Kreindler JL, Dubin
PJ, Ulrich L, Steele C, Finder JD, Pilewski JM, Carreno BM, Goldman
SJ, et al: Role of IL-17A, IL-17F, and the IL-17 receptor in
regulating growth-related oncogene-alpha and granulocyte
colony-stimulating factor in bronchial epithelium: Implications for
airway inflammation in cystic fibrosis. J Immunol. 175:404–412.
2005.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Xing Y, Li C, Li A, Sridurongrit S, Tiozzo
C, Bellusci S, Borok Z, Kaartinen V and Minoo P: Signaling via Alk5
controls the ontogeny of lung clara cells. Development.
137:825–833. 2010.PubMed/NCBI View Article : Google Scholar
|