Introduction
Atherosclerosis is well recognized as a chronic
inflammatory disease (1) that can
cause various cardiovascular pathologies, including myocardial
infarction, sudden cardiac death and stroke (2). During the development of
atherosclerosis, infiltrated macrophages triggered by endothelial
dysfunction play an important role (3). Macrophage polarization refers to an
estimate of macrophage activation at a given point in space and
time. Currently, the macrophage field has arrived at a partial
consensus to describe the broad grouping of macrophage activation
phenotypes and the underlying mechanisms (4). The classical inflammatory macrophage
phenotype has been termed the M1 phenotype and is usually induced
by a combination of interferon-γ and the Toll-like receptor 4
ligand lipopolysaccharide. By contrast, the alternatively activated
macrophage populations are termed M2 populations and can be induced
by incubation of macrophages with interleukin (IL)-4 and IL-13.
Upon exposure to multiple environmental stimuli, macrophages
exhibit considerable plasticity, which is responsible for the
inflammatory response underlying atherogenesis through a switch
between classically activated M1 macrophages, which produce a wide
variety of proinflammatory mediators, and alternatively activated
M2 macrophages that alleviate inflammation (5-7).
Additionally, lesion macrophages can give rise to lipid-laden
foam-cell formation following excess engulfment of oxidized
low-density lipoprotein (Ox-LDL) (8,9).
Importantly, previous studies have also suggested that M1 or M2
macrophages may be the main foam cell precursors through the
regulation of cholesterol transport by a liver-X-receptor-dependent
mechanism (10,11). Macrophage activation constituted by
the aforementioned macrophage polarization shift and foam-cell
formation leads to the enlargement of necrotic cores and formation
of vulnerable plaques (12). Thus,
exploring the key regulators and potential mechanisms of
atherosclerosis-associated macrophage activation may provide
clinicians with an effective strategy for preventing
atherosclerosis.
MicroRNAs (miRNAs/miRs) are an evolutionarily
conserved class of small non-coding RNAs with 20-23 nucleotides
(13) and exert important
regulatory effects on various biological processes, including cell
development, cell differentiation, cell proliferation, apoptosis
and immune inflammation, through regulation of gene expression by
directly binding to the 3'-untranslated regions (3'-UTRs) of their
target mRNAs (14-16).
Accumulating evidence has demonstrated that miR-183 is strongly
implicated and functions in various aspects of tumorigenesis.
miR-183 is reported to be a potent prognostic marker for lung
cancer through negative regulation of the metastatic potential of
lung cancer cells but acts as a tumor suppressor in human non-small
cell lung cancer by downregulating MTA1(17). miR-183 inhibits osteosarcoma cell
growth and invasion by regulating the LRP6-Wnt/β-catenin signaling
pathway (18), and promotes
proliferation and invasion in esophageal squamous cell carcinoma by
targeting programmed cell death 4(19). Recently, miR-183 has emerged as an
important player in cardiovascular disease and showed downregulated
expression in hypertrophic cardiac tissues subjected to transverse
aortic constriction surgery (20).
Additionally, inhibition of miR-183 expression in HUVECs with
Ox-LDL-induced injury can enhance cell activity, decrease
inflammation level, and thus prevent cell injury by increasing
insulin receptor substrate 1 expression (21). miR-183 alleviates early injury,
including decreasing brain edema, neurobehavioral defects,
inflammation, oxidative stress and iron deposition, and by
inhibiting heme oxygenase-1 expression after intracerebral
hemorrhage. A miR-183 inhibitor was also found to suppress
anaphylaxis and atopic dermatitis by decreasing NF-κB activation
(22). However, the potential role
and precise underlying mechanism of miR-183 in macrophage
activation have not been elucidated.
The present study demonstrated that significantly
upregulated miR-183 expression was found in atherosclerotic plaques
of ApoE-/- mice fed a high-fat diet (HFD) and in BMDMs
upon Ox-LDL stimulation. miR-183 knockdown dramatically accelerated
the activation of resolving M2-polarized macrophages but attenuated
the activation of classical M1-polarized macrophages, as well as
inhibited NF-κB activation. Additionally, miR-183 silencing
attenuated foam-cell formation accompanied by increased
upregulation of genes involved in cholesterol efflux, and
downregulation of genes implicated in cholesterol influx.
Mechanistically, it was verified that NR4A2 was the direct target
for the attenuating effect on macrophage activation regulated by
miR-183 knockdown. Overall, the present study results demonstrated
that blockade of the miR-183-NR4A2 axis might be a promising
strategy for the regulation of macrophage activation implicated in
atherogenesis.
Materials and methods
Animals and morphological
analysis
The animal study procedures were performed in
accordance with the National Institutes of Health Guide for the
Care and Use of Laboratory Animals and were approved by the Animal
Care and Use Committee of the Tongren municipal people's Hospital.
A total of 25 male ApoE deficient mice (ApoE-/-; age,
8-12 weeks; weight, 21-28 g; Beijing Zhongke Biotechnology Co.,
Ltd.) were fed a high-fat diet (HFD; 15.8% fat and 1.25%
cholesterol) or normal chow (NC) purchased from Beijing Huafukang
Bioscience Co Inc., respectively, for up to 12 weeks. Animals were
maintained under a 12 h light/dark cycle with free access to food
and water at a controlled temperature of 22-26˚C and relative
humidity (65±10%). Hearts from mice subjected to NC or HFD
treatment for 12 weeks were excised after anesthetization via the
intraperitoneal injection of pentobarbital sodium (50 mg/kg). The
tissues were subsequently dehydrated at room temperature in 85%
alcohol for 15 min, 95% alcohol twice for 10 min each and in 100%
alcohol twice for 15 min and embedded in paraffin for histological
analysis. Consecutive 5-µm sections of the atrioventricular valve
region of each heart were collected and stained with hematoxylin
and eosin (H&E) for 5 min for morphological analysis at a
temperature of 26˚C. Samples were observed under a light microscope
at a x40 magnification.
Cell culture and small interfering
(si)RNA transfection
Approximately 5x107 nucleated BMDMs
isolated form femurs and tibias of ApoE-mice were cultured in 10 ml
of RPMI with 10% fetal bovine serum and Macrophage
Colony-Stimulating Factor (MCSF; 50 ng/ml; cat. no. M9170,
Sigma-Aldrich; Merck KGaA) (23-25).
Cells were cultured in RPMI-1640 (Sigma-Aldrich; Merck KGaA)
containing 10% fetal bovine serum (Sigma-Aldrich; Merck KGaA) and
1% penicillin-streptomycin. The synthetic miR-183 antagomir was
synthesized by GenePharma. BMDMs were transfected with miR-183
antagomir and the corresponding controls (NC) at a concentration of
50 nM using a riboFEC CP Transfection kit according to the
manufacturer's protocol. The infected BMDMs stimulated with 15
ng/ml Ox-LDL, for 24 h after serum starvation for one day, were
utilized for the in vitro functional study.
Reverse transcription-quantitative
(RT-q)PCR and western blotting
Total mRNA was extracted by trichloromethane,
dissolved in DEPC-water, and then reverse transcribed into cDNA
with a Transcriptor First Strand cDNA Synthesis kit (F. Hoffmann-La
Roche AG) as previously described (26). The expression levels of the target
genes were quantified by real-time PCR using LightCycler 480 SYBR
Green 1 Master mix and a LightCycler 480 QPCR System (F.
Hoffmann-La Roche AG). The thermocycling conditions were as
follows: 95˚C for 10 min followed by 50 cycles of 95˚C for 10 sec,
55˚C for 10 sec, 72˚C for 5 sec; 99˚C for 1 sec; 59˚C for 15 sec;
95˚C for 1 sec; and cooling at 40˚C. The relative transcription
levels of the target genes were normalized against GAPDH, while the
level of miR-375 was normalized to U6 level. The 2-ΔΔCq
method was used (27). The primers
are listed in the Table I.
 | Table ISequences of primers used for reverse
transcription-quantitative PCR. |
Table I
Sequences of primers used for reverse
transcription-quantitative PCR.
| Primer | Sequence, 5' to
3' |
|---|
| TNF-α-F |
ATGGCCTCCCTCTCATCAGT |
| TNF-α-R |
ATAGCAAATCGGCTGACGGT |
| Inos-F |
AGGGCCACCTCTACATTTGC |
| Inos-R |
TGCCCCATAGGAAAAGACTGC |
| COX-2-F |
ATTGCCCTCCCCTCTCTACG |
| COX-2-R |
CGGCTCATGAGTGGAGAACG |
| IL-6-F |
CTGGAGTACCATAGCTACCTGG |
| IL-6-R |
ACTCCTTCTGTGACTCCAGC |
| Arg-1-F |
AAAGGCCGATTCACCTGAGC |
| Arg-1-R |
AGGTAGTCAGTCCCTGGCTT |
| TGF-β-F |
AGAGCCCTGGATACCAACTATTG |
| TGF-β-R |
TGCGACCCACGTAGTAGACG |
| Mrc-1-F |
CCTGTAACTACACACTCATCCA |
| Mrc-1-R |
CATTCTGCTCGATGTTGCCC |
| PPARγ-F |
GCTTGTGAAGGATGCAAGGG |
| PPARγ-R |
GATATCACTGGAGATCTCCGCC |
| SR-A-F |
TGGAGGAGAGAATCGAAAGCA |
| SR-A-R |
CTGGACTGACGAAATCAAGGAA |
| CD36-F |
GACTGGGACCATTGGTGATGA |
| CD36-R |
AAGGCCATCTCTACCATGCC |
| ABCA1-F |
AGGCACTCAAGCCACTGCTTGT |
| ABCA1-R |
TGCCTCTGCTGTCTAACAGCGT |
| ABCG1-F |
GGTTGCGACATTTGTGGGTC |
| ABCG1-R |
TTCTCGGTCCAAGCCGTAGA |
| NR4A2-F |
GCCATGCCTTGTGTTCAGGCGCAG |
| NR4A2-R |
GGAGGTCTTAGAAAGGTAAAGTGTCC |
| GAPDH-F |
GGTGGACCTCATGGCCTACA |
| GAPDH-R |
CTCTCTTGCTCTCAGTATCCTTGCT |
| U6-F |
CTCGCTTCGGCAGCACA |
| U6-R |
AACGCTTCACGAATTTGCGT |
Protein concentrations were determined by
Bicinchoninic Acid Kit assay (cat. no. BCA1; Sigma-Aldrich; Merck
KGaA). The proteins (5 µg) were separated by 7.5% SDS-PAGE
(Invitrogen; Thermo Fisher Scientific, Inc.) and then transferred
to polyvinylidene fluoride membranes, which were blocked in
Tris-buffered saline (TBS) and Tween-20 containing 5% non-fat milk
1 h at room temperate. Subsequently, the membranes were incubated
with the appropriate primary antibodies overnight at 4˚C.
The membranes were then incubated with a secondary
IRDye® 800CW-conjugated antibody (1:5,000; cat. no.
ab216773; Abcam) for 1 h at 37˚C, and treated with enhanced
chemiluminescence reagent (Thermo Fisher Scientific, Inc.) before
being visualized with Molecular Imager ChemiDoc™ XRS (Bio-Rad
Laboratories, Inc.) using Image Lab™ Software 5.1 (Bio-Legend
Scientific Co., Ltd). The expression levels of specific proteins
were normalized against GAPDH expression. The antibodies used are
listed in the Table II.
| Table IIAntibodies for immunoblot
analyses. |
Table II
Antibodies for immunoblot
analyses.
| Antibody | Catalogue
number | Dilution | Supplier | Sources of
species |
|---|
| NR4A2 | 10975-2-AP | 1:200 | ProteinTech Group,
Inc. | Rabbit |
| IL-6 | AF-406-NA | 1:200 | R&D Systems,
Inc. | Goat |
| Arg-1 | 610708 | 1:200 | BD Biosciences | Mouse |
| CD36 | sc-7309 | 1:100 | Santa Cruz
Biotechnology, Inc. | Mouse |
| ABCA1 | ab7360 | 1:200 | Abcam | Rabbit |
| P-p65 | ab76302 | 1:200 | Abcam | Rabbit |
| P65 | ab16502 | 1:100 | Abcam | Rabbit |
| GAPDH | 2118 | 1:1,000 | Cell Signaling
Technology, Inc. | Rabbit |
Foam-cell formation
BMDMs serum-starved for 24 h and stimulated with 15
ng/ml Ox-LDL for 24 h were collected. To visualize lipid droplets
by oil red O staining, the cells were washed twice with PBS and
fixed with 4% paraformaldehyde for 10 min at temperature of 26˚C.
The slides were rinsed quickly in 60% isopropyl alcohol and stained
in 0.3% oil red O solution for 10 min, respectively. Subsequently,
the slides were washed briefly in 60% isopropyl alcohol for 3-10
sec and mounted using 50% glycerol, which were observed under a
light microscope (magnification, x200).
Luciferase activity
Luciferase reporters were generated based on
Pezx-MT01 vector purchased from Shanghai GenePharma Co., Ltd.
HEK293 cells were purchased from the Type Culture Collection of the
Chinese Academy of Sciences and cultured at 37˚C in a humidified 5%
CO2 incubator. Cells were seeded into a 24-well plate
and infected with miR-183 antagomir (50 nm) or NC (50 nm) and then
transfected with luciferase reporter containing the wild-type or
mutant 3'-UTR of NR4A2 using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) for 48 h according to
the manufacturer's instructions. Then, dual luciferase assays
(Promega Corporation) was performed to analysis the luciferase
activity according to the manufacturer's instructions.
TargetScan
TargetScan predicts the biological targets of miRNAs
by searching for the presence of conserved 8mer, 7mer and 6mer
sites that match the seed region of each miRNA. The URL of the
database was http://www.targetscan.org/mmu_71/.
Statistical analysis
All statistical data were analyzed using SPSS
software version 22.0 (IBM Corp.) and are presented as the mean ±
SD. Differences between two groups were analyzed by
independent-samples t-tests, while differences among multiple
groups were analyzed by one-way ANOVA followed by a Bonferroni post
hoc test or with Tamhane's T2 analysis. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-183 expression is upregulated in
atheromatous plaques and macrophages
To investigate the involvement of miR-183 in
atherogenesis, atherosclerotic plaques from ApoE-/- mice
induced by HFD feeding were first examined to determine whether
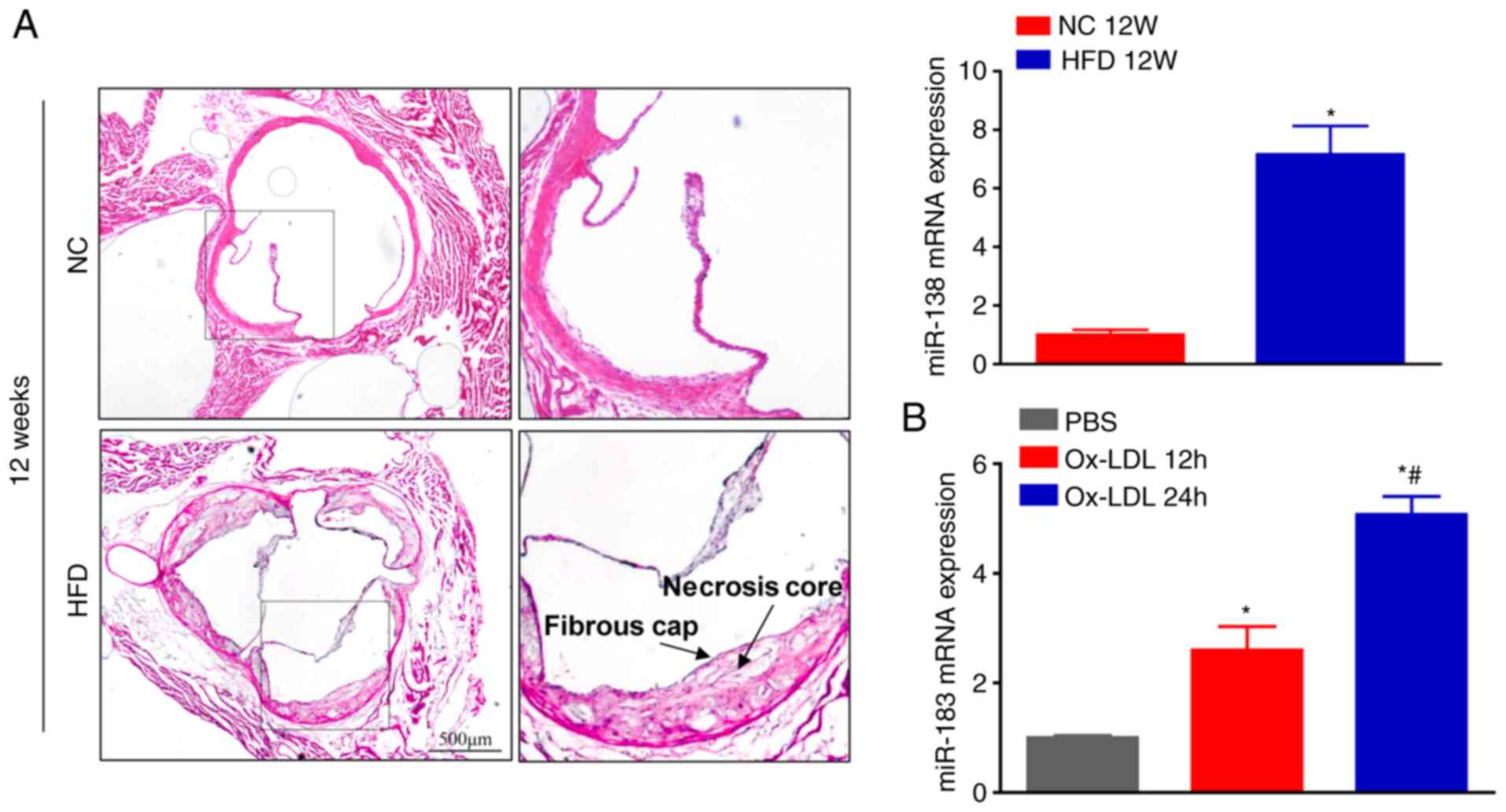
miR-183 expression was altered. Hematoxylin and eosin staining
demonstrated the typical pathological changes of atherosclerotic
plaques in ApoE-/- mice fed a HFD for 12 weeks,
characterized by the thin fibrous cap and large necrotic core
(Fig. 1A). RT-Qpcr analysis
revealed that miR-183 Mrna expression levels were markedly
upregulated in arteries of HFD-fed ApoE-/- mice compared
with those of NC-treated mice (Fig.
1A). Furthermore, the expression of miR-183 in BMDMs subjected
to Ox-LDL stimulation was also evaluated. It was found that miR-183
expression was gradually upregulated in BMDMs following Ox-LDL
stimulation (Fig. 1B). These
findings suggest that miR-183 is involved in the development of
atherosclerosis, partially through functioning in macrophages.
miR-183 silencing ameliorates the
inflammatory response
Given the significant change in miR-183 expression
in atheromatous plaques and macrophages treated with Ox-LDL,
miR-183 loss-of-function experiments were performed in macrophages.
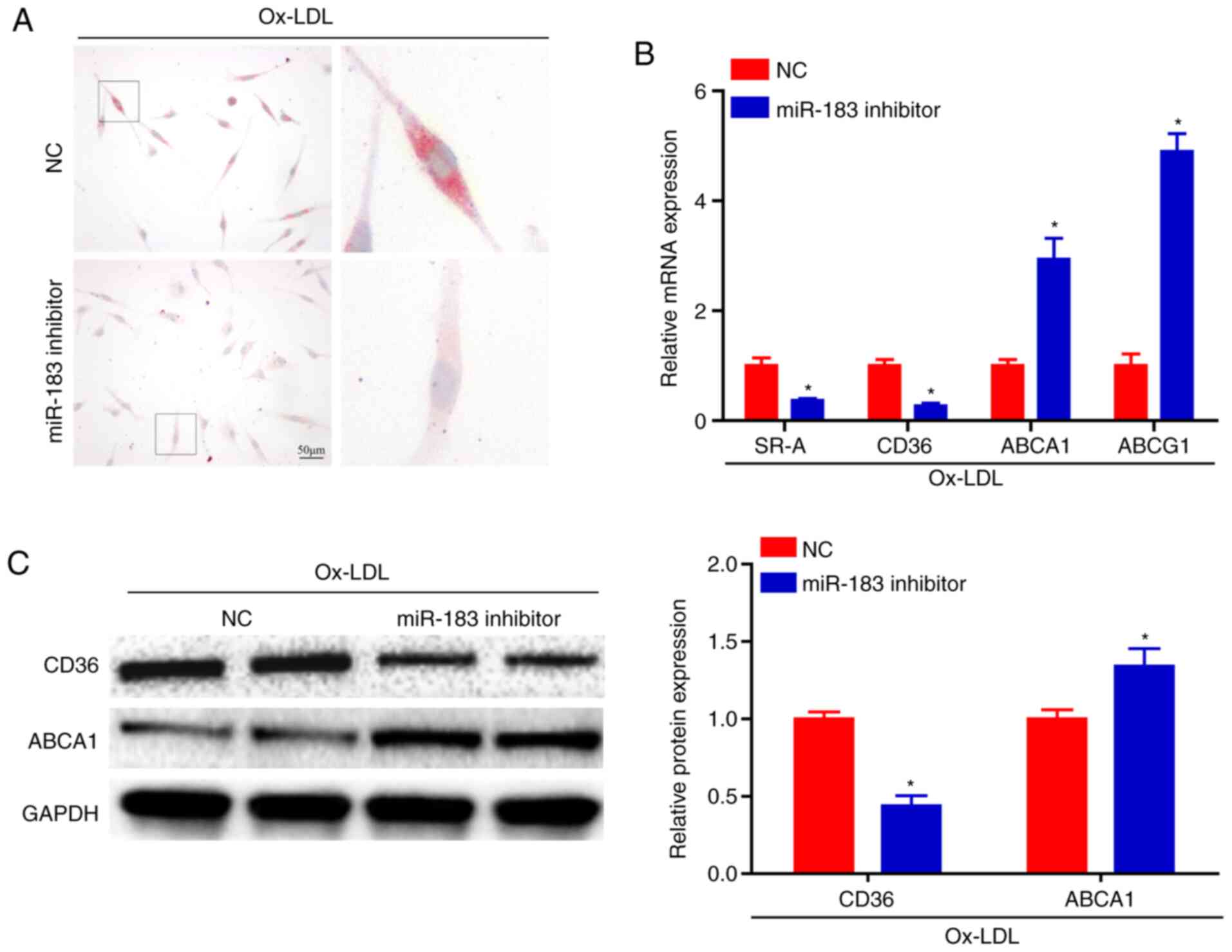
It was observed that miR-183 expression was notably decreased in
BMDMs following miR-183 inhibitor transfection (Fig. 2A). Furthermore, the Mrna levels of
genes associated with proinflammatory M1-polarized macrophages,
including tumor necrosis factor-α, inducible nitric oxide synthase,
cyclooxygenase 2 and IL-6, were dramatically downregulated in BMDMs
transfected with the miR-183 inhibitor upon Ox-LDL treatment
compared with the control group (Fig.
2B). In contrast, the Mrna expression levels of genes
associated with resolving M2-polarized macrophages, including
Arginase-1 (Arg-1), transforming growth factor-β, mediator of
replication checkpoint protein 1 and peroxisome
proliferator-activated receptors γ were significantly upregulated
in miR-183 inhibitor-transfected BMDMs with Ox-LDL treatment
compared with control cells (Fig.
2C). Moreover, IL-6 and Arg-1 protein levels determined by
western blotting analysis were consistent with the Mrna expression
levels (Fig. 2D). Furthermore,
phosphorylation of p65, an important event in the activation of the
inflammatory mediator nuclear factor-kappa B (NF-κB), was decreased
in BMDMs transfected with the miR-183 inhibitor (Fig. 2E).
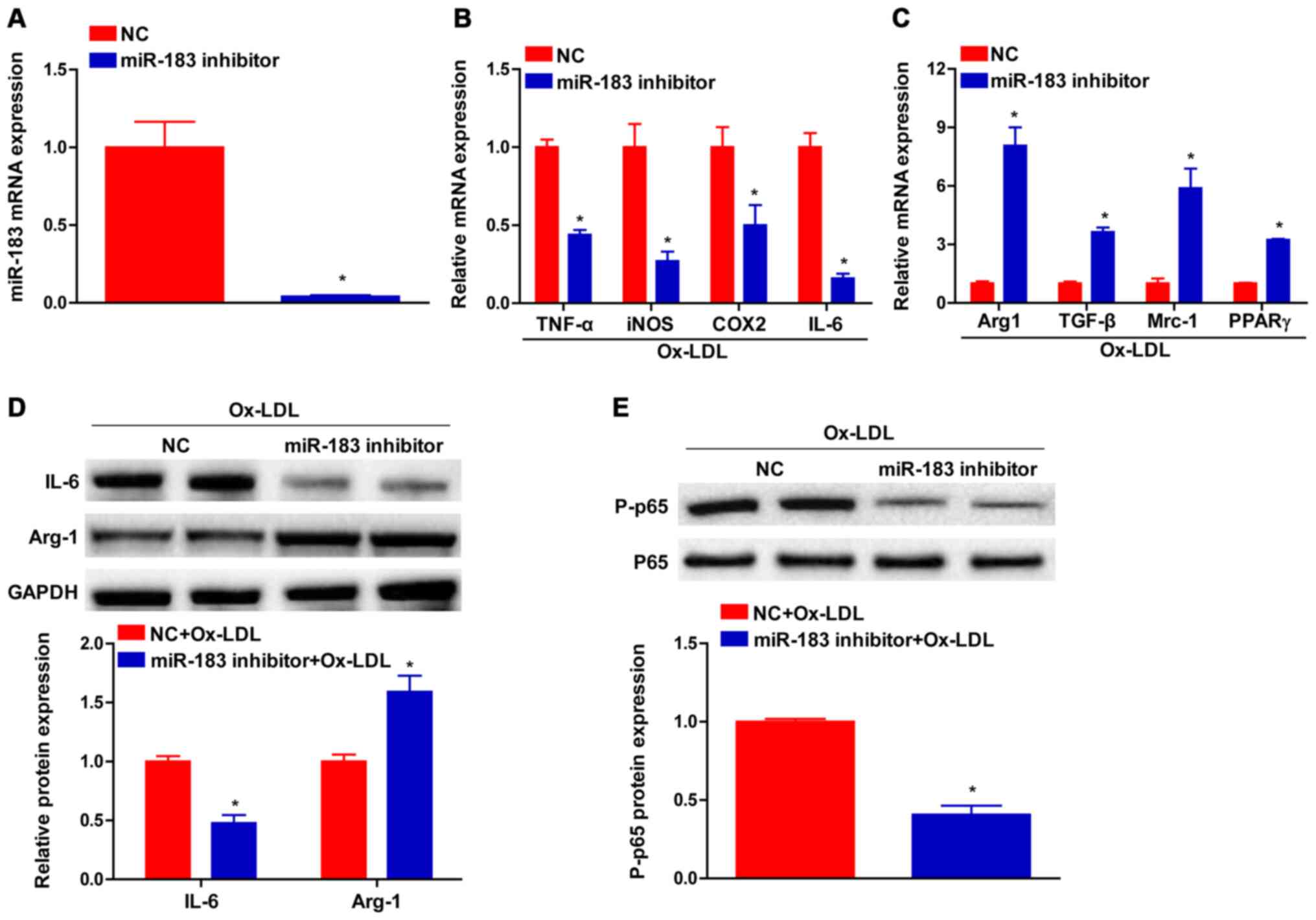 | Figure 2Pro-inflammatory macrophage response
regulated by miR-183 knockdown. (A) miR-183 expression in BMDMs
infected with miR-183 inhibitor or NC. Mrna expression of M1 marker
genes (B) and M2 marker genes (C) in BMDMs transfected with miR-183
inhibitor or NC examined by reverse transcription-quantitative PCR
analysis. (D and E) The protein level of IL-6 and Arg-1, as well as
phosphorylated and total p65 in BMDMs infected with miR-183
inhibitor or NC tested by western blot analysis.
*P<0.05 vs. control group. miR, microRNA; BMDMs, bone
marrow-derived macrophages; NC, negative control; TNFα, tumor
necrosis factor α; Inos, inducible nitric oxide synthase; COX,
cyclooxygenase; IL, interleukin; Ox-LDL, oxidized low-density
lipoprotein; Arg, arginase; TGF, transforming growth factor; Mrc,
mediator of replication checkpoint protein 1; PPAR, peroxisome
proliferator-activated receptors. |
miR-183 knockdown attenuates foam-cell
formation
After excess engulfment of Ox-LDL, macrophages form
foam cells, which play an important role in atherogenesis. A
significant decrease in foam-cell formation in BMDMs transfected
with the miR-183 inhibitor was observed upon Ox-LDL stimulation, as
evaluated by neutral lipid staining with oil red O, compared with
the control group (Fig. 3A).
Subsequently, the markers associated with cholesterol uptake and
efflux that contribute to foam-cell formation were evaluated. BMDMs
transfected with miR-183 inhibitor presented with decreased Mrna
levels of genes (SR-A and CD36) involved in cholesterol influx,
whereas the expression levels of markers implicated in cholesterol
efflux (ABCA1 and ABCG1) were increased (Fig. 3B). Moreover, the CD36 and ABCA1
protein levels determined by western blot analysis were consistent
with the Mrna expression levels (Fig.
3C).
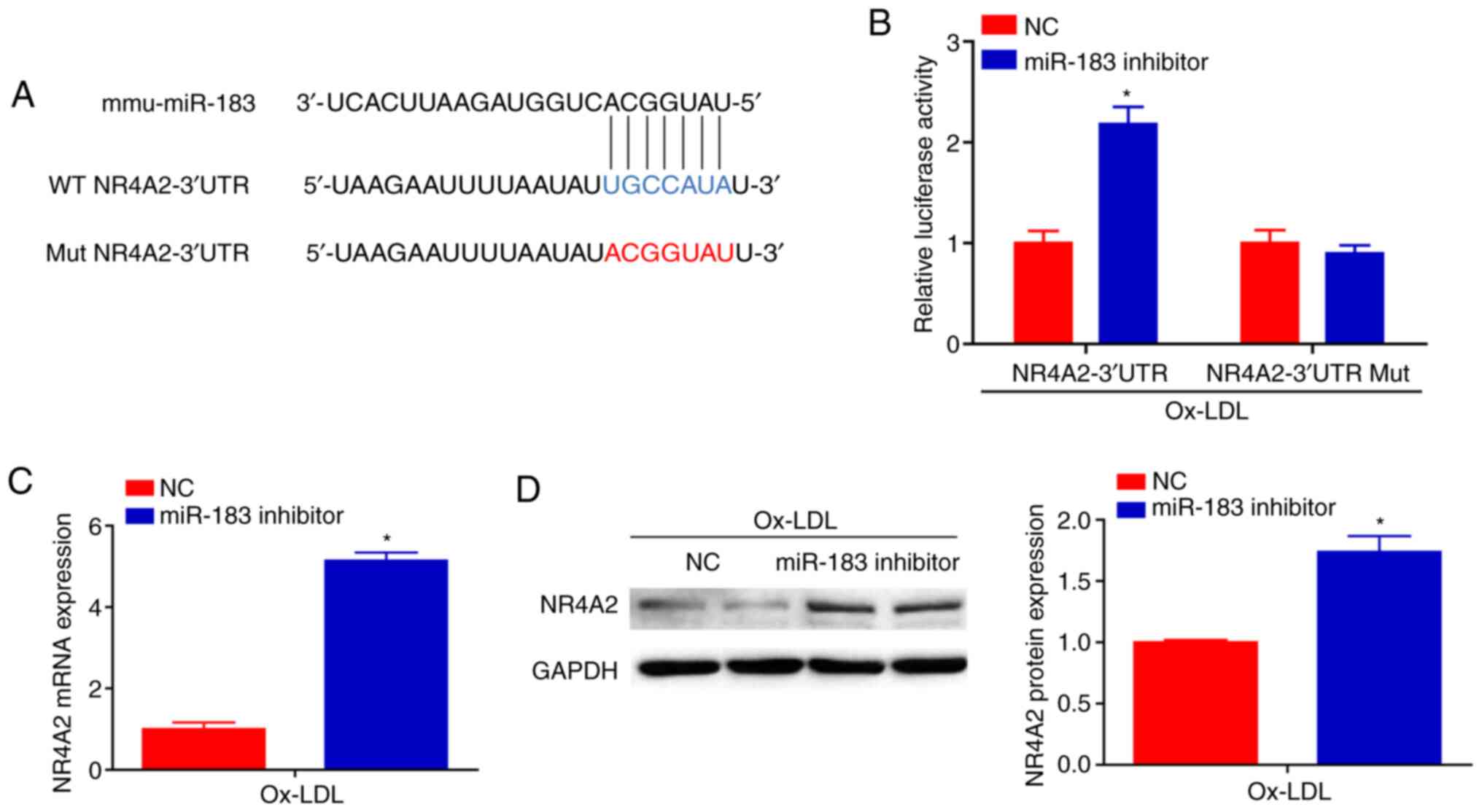
Knockdown of miR-183 promotes NR4A2
expression
Next, bioinformatic analysis using the program
TargetScan was performed to elucidate the mechanism by which
miR-183 affects macrophage activation. The results indicated that
NR4A2 contained a putative binding site for miR-183 in its 3'UTR
(Fig. 4A). Moreover, luciferase
reporter assays demonstrated that miR-183 inhibitor transfection
promoted NR4A2 luciferase activity, whereas the above effect was
abolished when the predicted binding sites within the NR4A2 3'UTR
were mutated (Fig. 4B). As
expected, NR4A2 showed significantly upregulated Mrna and protein
expression in BMDMs transfected with the miR-183 inhibitor
(Fig. 4C and D). Collectively, these findings suggested
that the effects of miR-183 knockdown on macrophage activation were
partially mediated by the upregulated expression of its molecular
target, NR4A2.
Discussion
The present study first demonstrated that miR-183 is
a novel and independent indicator of macrophage activation
implicated in atherogenesis, which exhibited an upregulated
expression in atherosclerotic plaques of ApoE-/- mice
fed with HFD and BMDMs with Ox-LDL stimulation. The
loss-of-function strategy demonstrated that miR-183 knockdown
promoted M2 phenotypic macrophage but attenuated M1 macrophage
switching. Meanwhile, miR-183 knockdown significantly ameliorated
foam cell forming by increased cholesterol efflux but decreased
cholesterol uptake. Mechanistically, NR4A2 was the direct target of
miR-183 and the silencing of miR-183 dramatically promoted NR4A2
expression. On the basis of the results from the present study, an
unexplored role of miR-183 in regulation of macrophage activation
was identified, which is at least partially through regulation of
NR4A2 expression.
Over the past decade, multiple studies have
demonstrated that various miRNAs are widely involved in numerous
human disorders and are especially implicated in multiple
cardiometabolic diseases, such as obesity, diabetes, heart failure
and atherosclerosis; the collective evidence implies their
potential as therapeutic targets and provides a new direction for
the study of these diseases (15,28-34).
Atherosclerosis is recognized as a complex multifactorial
pathological process (35), and
microRNAs have emerged as important regulators of atherosclerosis
development through the regulation of the inflammatory response via
the switch between classical proinflammatory M1 macrophage
activation and alternative inflammation-resolving M2 macrophage
activation (36-39).
In the present study, it was observed that the expression of
miR-183 was gradually upregulated in atherosclerotic plaques of
ApoE-/- mice and in BMDMs following Ox-LDL stimulation.
miR-183 silencing attenuated the Mrna expression of prototypical
genes associated with proinflammatory M1-polarized macrophages but
increased the Mrna expression levels of genes associated with
anti-inflammatory M2-polarized macrophages. The predominance of
M2-polarized macrophages in bladder cancer affects angiogenesis,
the tumor grade and invasiveness. Macrophage polarization has been
well recognized to exert an important role in the regulation of
tumorigenesis and angiogenesis (40), and has emerged as the main precursor
of foam-cell formation mediated by macrophages, which together play
important roles in the development of atherosclerosis. The present
study demonstrated that a miR-183 inhibitor attenuated the
formation of foam cell contaminants, with decreased Mrna levels of
genes involved in cholesterol influx but increased expression
levels of markers implicated in cholesterol efflux.
miRNAs are capable of regulating target gene
expression by binding to the 3'-UTRs of their target mRNAs, and
miRNAs are involved in negatively regulating gene expression at the
posttranscriptional level (13).
Bioinformatic analysis was performed using the program TargetScan,
which found that NR4A2 contained a putative binding site for
miR-183 in its 3'UTR. Moreover, it was demonstrated that miR-183
inhibitor transfection increased NR4A2 luciferase activity, as
evaluated by a luciferase reporter assay, whereas the
aforementioned effect was abolished when the predicted binding
sites within the NR4A2 3'UTR were mutated. Importantly, NR4A2 Mrna
and protein expression were also significantly increased in BMDMs
transfected with the miR-183 inhibitor. A previous study
demonstrated that members of the NR4A subfamily (NR4A1/Nur77,
NR4A2/Nurr1 and NR4A3/Nor1) of orphan NRs (nuclear receptors) act
as ligand-independent and constitutively active receptors, which
are tightly controlled at the levels of expression,
posttranslational modification and subcellular localization
(41). NR4A subfamily members have
emerged as key transcriptional regulators of cytokine and growth
factor action in various diseases. Recently, accumulating evidence
has suggested that NR4A nuclear receptors play pivotal roles in
immunity and atherosclerosis (41),
while all three NR4A family members are expressed within
atherosclerotic lesions. The orphan nuclear receptor NR4A1
attenuates atherosclerosis by downregulating inflammatory gene
expression in macrophages (42,43).
NR4A3 deficiency has been shown to inhibit vascular injury and
atherosclerosis in mice (44,45).
NR4A2 has been shown to act as a repressor of NF-Κb activation in
microglia and astrocytes and to attenuate atherosclerosis in mice
(46). Exogenous expression of
NR4A2 in macrophages leads to polarization toward their alternative
phenotype, with induction of genes that are prototypical M2 markers
(47). Moreover, NR4A2
transcriptionally activates Arg1 expression by directly binding to
its promoter (7). Thus, it is
speculated that miR-183 knockdown attenuated macrophage activation
partially by upregulating NR4A2 expression, which may play an
important protective role against atherogenesis.
In conclusion, the present study demonstrated that
miR-183 attenuated macrophage activation contributed by decreasing
the classic pro-inflammatory M1 macrophage but increasing
alternative inflammation resolving M2 macrophage activation,
accompanied by attenuation of foam-cell formation, along with
enhanced genes associated with cholesterol efflux compared to
uptake. This study highlights the important role of neutralizing
miR-183 as a promising therapeutic strategy for atherosclerosis
management.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the Science
and Technology Fund of Health commission of Guizhou province (grant
no. gzwjkj2019-1-038).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FHG and LL designed and conducted in vitro
experiments, analyzed the data and drafted the paper. YSY and DHS
performed the biomolecule experiment. YSZ, XSW and XPZ performed
the pathological study and analyzed the data. XQX designed all of
the experiments, supervised and funded the study, and contributed
to the data analysis and to the writing of the paper. All authors
read and approved the final manuscript and FHG and XQX confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
The animal study procedures were performed in
accordance with the National Institutes of Health Guide for the
Care and Use of Laboratory Animals and were approved by the Animal
Care and Use Committee of the Tongren municipal people's Hospital
(approval no. 20190609).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Libby P: Inflammation in atherosclerosis.
Nature. 420:868–874. 2002.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Virani SS, Alonso A, Benjamin EJ,
Bittencourt MS, Callaway CW, Carson AP, Chamberlain AM, Chang AR,
Cheng S, Delling FN, et al: Heart disease and stroke
statistics-2020 update: A report from the American heart
association. Circulation. 141:e139–e596. 2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Tabas I and Bornfeldt KE: Macrophage
phenotype and function in different stages of atherosclerosis. Circ
Res. 118:653–667. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Murray PJ: Macrophage polarization. Annu
Rev Physiol. 79:541–566. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Mantovani A, Garlanda C and Locati M:
Macrophage diversity and polarization in atherosclerosis: A
question of balance. Arterioscler Thromb Vasc Biol. 29:1419–1423.
2009.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Sica A and Mantovani A: Macrophage
plasticity and polarization: In vivo veritas. J Clin Invest.
122:787–795. 2012.PubMed/NCBI View
Article : Google Scholar
|
|
7
|
Leitinger N and Schulman IG: Phenotypic
polarization of macrophages in atherosclerosis. Arterioscler Thromb
Vasc Biol. 33:1120–1126. 2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Tall AR, Yvan-Charvet L, Terasaka N,
Pagler T and Wang N: HDL, ABC transporters, and cholesterol efflux:
Implications for the treatment of atherosclerosis. Cell Metab.
7:365–375. 2008.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Tall AR and Yvan-Charvet L: Cholesterol,
inflammation and innate immunity. Nat Rev Immunol. 15:104–116.
2015.PubMed/NCBI View
Article : Google Scholar
|
|
10
|
Gleissner CA, Shaked I, Little KM and Ley
K: CXC chemokine ligand 4 induces a unique transcriptome in
monocyte-derived macrophages. J Immunol. 184:4810–4818.
2010.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Finn AV, Nakano M, Polavarapu R, Karmali
V, Saeed O, Zhao X, Yazdani S, Otsuka F, Davis T, Habib A, et al:
Hemoglobin directs macrophage differentiation and prevents foam
cell formation in human atherosclerotic plaques. J Am Coll Cardiol.
59:166–177. 2012.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Tabas I: Macrophage death and defective
inflammation resolution in atherosclerosis. Nat Rev Immunol.
10:36–46. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
13
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Xu P, Guo M and Hay BA: MicroRNAs and the
regulation of cell death. Trends Genet. 20:617–624. 2004.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Divakaran V and Mann DL: The emerging role
of microRNAs in cardiac remodeling and heart failure. Circ Res.
103:1072–1083. 2008.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Yang CL, Zheng XL, Ye K, Ge H, Sun YN, Lu
YF and Fan QX: MicroRNA-183 Acts as a tumor suppressor in human
non-small cell lung cancer by down-regulating MTA1. Cell Physiol
Biochem. 46:93–106. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yang X, Wang L, Wang Q, Li L, Fu Y and Sun
J: miR-183 inhibits osteosarcoma cell growth and invasion by
regulating LRP6-Wnt/β-catenin signaling pathway. Biochem Biophys
Res Commun. 496:1197–1203. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Wei C, Song H, Sun X, Li D, Song J, Hua K
and Fang L: miR-183 regulates biological behavior in papillary
thyroid carcinoma by targeting the programmed cell death 4. Oncol
Rep. 34:211–220. 2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Chen C, Ponnusamy M, Liu C, Gao J, Wang K
and Li P: MicroRNA as a therapeutic target in cardiac remodeling.
Biomed Res Int. 2017(1278436)2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhang Y, Zhan Y, Liu D and Yu B:
Inhibition of microRNA-183 expression resists human umbilical
vascular endothelial cells injury by upregulating expression of
IRS1. Drug Deliv. 26:612–621. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kim M, Jo H, Kwon Y, Kim Y, Jung HS and
Jeoung D: Homoharringtonine inhibits allergic inflammations by
regulating NF-κB-miR-183-5p-BTG1 axis. Front Pharmacol.
11(1032)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Krausgruber T, Blazek K, Smallie T,
Alzabin S, Lockstone H, Sahgal N, Hussell T, Feldmann M and Udalova
IA: IRF5 promotes inflammatory macrophage polarization and TH1-TH17
responses. Nat Immunol. 12:231–238. 2011.PubMed/NCBI View
Article : Google Scholar
|
|
24
|
Liao X, Sharma N, Kapadia F, Zhou G, Lu Y,
Hong H, Paruchuri K, Mahabeleshwar GH, Dalmas E, Venteclef N, et
al: Kruppel-like factor 4 regulates macrophage polarization. J Clin
Invest. 121:2736–2749. 2011.PubMed/NCBI View
Article : Google Scholar
|
|
25
|
Liu H, Cheng WL, Jiang X, Wang PX, Fang C,
Zhu XY, Huang Z, She ZG and Li H: Ablation of interferon regulatory
factor 3 protects against atherosclerosis in apolipoprotein
E-deficient mice. Hypertension. 69:510–520. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wang PX, Ji YX, Zhang XJ, Zhao LP, Yan ZZ,
Zhang P, Shen LJ, Yang X, Fang J, Tian S, et al: Targeting CASP8
and FADD-like apoptosis regulator ameliorates nonalcoholic
steatohepatitis in mice and nonhuman primates. Nat Med. 23:439–449.
2017.PubMed/NCBI View
Article : Google Scholar
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Arner P and Kulyte A: MicroRNA regulatory
networks in human adipose tissue and obesity. Nat Rev Endocrinol.
11:276–288. 2015.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Nazari-Jahantigh M, Egea V, Schober A and
Weber C: MicroRNA-specific regulatory mechanisms in
atherosclerosis. J Mol Cell Cardiol. 89:35–41. 2015.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Schober A, Nazari-Jahantigh M and Weber C:
MicroRNA-mediated mechanisms of the cellular stress response in
atherosclerosis. Nat Rev Cardiol. 12:361–374. 2015.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Tana C, Giamberardino MA and Cipollone F:
microRNA profiling in atherosclerosis, diabetes, and migraine. Ann
Med. 49:93–105. 2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wang H and Cai J: The role of microRNAs in
heart failure. Biochim Biophys Acta Mol Basis Dis. 1863:2019–2030.
2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Zhang L, Miller D, Yang Q and Wu B:
MicroRNA regulatory networks as biomarkers in obesity: The emerging
role. Methods Mol Biol. 1617:241–260. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Li C, Qu L, Farragher C, Vella A and Zhou
B: MicroRNA regulated macrophage activation in obesity. J Transl
Int Med. 7:46–52. 2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Lusis AJ: Atherosclerosis. Nature.
407:233–241. 2000.PubMed/NCBI View
Article : Google Scholar
|
|
36
|
Banerjee S, Xie N, Cui H, Tan Z, Yang S,
Icyuz M, Abraham E and Liu G: MicroRNA let-7c regulates macrophage
polarization. J Immunol. 190:6542–6549. 2013.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Ouimet M, Ediriweera HN, Gundra UM, Sheedy
FJ, Ramkhelawon B, Hutchison SB, Rinehold K, van Solingen C,
Fullerton MD, Cecchini K, et al: MicroRNA-33-dependent regulation
of macrophage metabolism directs immune cell polarization in
atherosclerosis. J Clin Invest. 125:4334–4348. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
38
|
Ying W, Tseng A, Chang RC, Morin A, Brehm
T, Triff K, Nair V, Zhuang G, Song H, Kanameni S, et al:
MicroRNA-223 is a crucial mediator of PPARγ-regulated alternative
macrophage activation. J Clin Invest. 125:4149–4159.
2015.PubMed/NCBI View
Article : Google Scholar
|
|
39
|
Qiu M, Ma J, Zhang J, Guo X, Liu Q and
Yang Z: MicroRNA-150 deficiency accelerates intimal hyperplasia by
acting as a novel regulator of macrophage polarization. Life Sci.
240(116985)2020.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Takeuchi H, Tanaka M, Tanaka A, Tsunemi A
and Yamamoto H: Predominance of M2-polarized macrophages in bladder
cancer affects angiogenesis, tumor grade and invasiveness. Oncol
Lett. 11:3403–3408. 2016.PubMed/NCBI View Article : Google Scholar
|
|
41
|
McMorrow JP and Murphy EP: Inflammation: A
role for NR4A orphan nuclear receptors? Biochem Soc Trans.
39:688–693. 2011.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Pei L, Castrillo A and Tontonoz P:
Regulation of macrophage inflammatory gene expression by the orphan
nuclear receptor Nur77. Mol Endocrinol. 20:786–794. 2006.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Hanna RN, Shaked I, Hubbeling HG, Punt JA,
Wu R, Herrley E, Zaugg C, Pei H, Geissmann F, Ley K and Hedrick CC:
NR4A1 (Nur77) deletion polarizes macrophages toward an inflammatory
phenotype and increases atherosclerosis. Circ Res. 110:416–427.
2012.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Qing H, Liu Y, Zhao Y, Aono J, Jones KL,
Heywood EB, Howatt D, Binkley CM, Daugherty A, Liang Y and Bruemmer
D: Deficiency of the NR4A orphan nuclear receptor NOR1 in
hematopoietic stem cells accelerates atherosclerosis. Stem Cells.
32:2419–2429. 2014.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Qing H, Jones KL, Heywood EB, Lu H,
Daugherty A and Bruemmer D: Deletion of the NR4A nuclear receptor
NOR1 in hematopoietic stem cells reduces inflammation but not
abdominal aortic aneurysm formation. BMC Cardiovasc Disord.
17(271)2017.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Hamers AA, Hanna RN, Nowyhed H, Hedrick CC
and de Vries CJ: NR4A nuclear receptors in immunity and
atherosclerosis. Curr Opin Lipidol. 24:381–385. 2013.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Mahajan S, Saini A, Chandra V, Nanduri R,
Kalra R, Bhagyaraj E, Khatri N and Gupta P: Nuclear Receptor Nr4a2
promotes alternative polarization of macrophages and confers
protection in sepsis. J Biol Chem. 290:18304–18314. 2015.PubMed/NCBI View Article : Google Scholar
|