Introduction
Fibrosis is defined as an excessive component
deposition of the extracellular matrix (ECM), collagen and
peptidoglycans in organs and tissues as a result of the
proliferation and activation of fibroblasts, stellate cells and
myofibroblasts (1-3).
Inflammatory reactions of both the innate and adaptive immune
system contribute to the development of fibrosis; in the early
stages of fibrosis, neutrophils, macrophages, natural killer (NK)
cells and T lymphocytes promote pro-fibrotic processes, including
hepatic stellate cell activation, increased transforming growth
factor (TGF)-β, platelet-derived growth factor, fibroblast growth
factor, matrix metalloproteinase (MMP) 9 and metalloproteinase
inhibitor-1/-2 expression, and a decrease in MMP13 expression
(1-4).
Activated macrophages produce tumor necrosis factor (TNF)-α and
interleukin (IL)-1, which in turn activate hepatic stellate cells
and fibroblasts to induce ECM overproduction. The signal
transduction triggered by TNF-α leads to the expression of
fibrogenic cytokines, primarily via the NF-κB and SMAD pathways
(1,3). By contrast, interferon (IFN)-γ
produced by activated NK cells (and a subsequent increase in IL-10)
exerts antifibrotic effects (3).
During cirrhosis, collagen types I and III are deposited in the
hepatic stroma, creating fine or wide fibrous septa. Subsequently,
new vascular channels are formed that facilitate communication
between the portal region (hepatic arteries and portal veins) and
the centrilobular veins, establishing an alternative circuit
through which blood can bypass the sinusoids due to the increase of
collagen fibers in the Dissé space (4,5).
Continuous collagen deposition in the Dissé space of
the parenchyma is associated with the loss of sinusoidal
endothelial cell fenestrae, in this process, the sinusoidal space
takes on a capillary-like structure rather than a channel for the
exchange of solutes between hepatocytes and the plasma (4). Collectively, this alters the secretion
of hepatocellular proteins such as albumin, coagulation factors and
lipoproteins (1,5). A number of therapeutic strategies have
been developed to prevent this process and to subsequently decrease
or reverse fibrosis/cirrhosis-associated liver damage (6); for example, the use of antioxidants
(7), adrenoblockers (8), anti-inflammatory cytokines (9,10) and
probiotics (11) has been
suggested, but a complete cure for the disease has yet to be
identified.
The pharmacological basis of a number of fibrosis
treatments is the interaction between the intestinal microbiome and
the host, which helps to maintain homeostasis (12-14).
Haller et al (15)
demonstrated that Lactobacillus (L.) johnsonii of an
intestinal origin did not induce TNF-α or IL-1β release, but
promoted that of TGF-β, presenting a global anti-inflammatory
profile in a colitis model. In addition to in vitro studies,
experimental animal models of colitis have demonstrated the
usefulness of probiotics in the control of intestinal inflammation.
In an acetic acid-induced rat colitis model, administration of
L. reuteri R2LC immediately after induction prevented the
development of colitis (16).
Similarly, L. plantarum administration to rats decreased the
severity of colitis in an intraperitoneal methotrexate-induced
enterocolitis model (17,18).
L. lactis is a gram-positive, spherical,
homolactic, non-sporulant and facultative anaerobic bacterium, with
hundreds of strains and biovariants published to date (19). L. lactis is categorized into
three subspecies: i) L. lactis ssp. Lactis; ii) L.
lactis ssp. Cremoris; and iii) L. lactis ssp.
Hordniae (19-21).
Bajaj et al (11)
demonstrated that L. rhamnosus GG induced a decrease in
endotoxemia and systemic inflammation in patients with cirrhosis.
Similar results were observed following the administration of
lactulose, rifaximin and probiotics containing
Lactobacillus, which partially reversed cirrhosis-associated
enteric dysbiosis, together with improving the severity of
encephalopathy (18). Due to its
immunomodulatory properties (22-24)
and ability to transit through the gastrointestinal tract, L.
lactis does not colonize the intestine in the manner of other
similar organisms, such as Lactobacillus spp. (24).
The primary beneficial effect reported for wild or
recombinant strains of L. lactis is its anti-inflammatory
potential, indicating its potential use as a therapeutic tool for
chronic intestinal diseases. Cellular in vitro models, as
well as mouse models of colitis, have been used to investigate the
anti-inflammatory properties of L. lactis, where an increase
in anti-inflammatory cytokines and a decrease in NF-κB have been
reported (25-28).
In the present study, the protective effects of oral administration
of L. lactis were evaluated after tetrachloromethane
(CCl4)-induced fibrosis in Wistar rats.
Materials and methods
Bacterial strains and culture
conditions
Pure stocks of L. lactis (10 µl) in 1 ml M17
medium (10% glucose and 30% glycerol) were donated by Dr Maria de
Jesus Loera Arias of the Autonomous University of Nuevo León
(Monterrey, Mexico). To reactivate the L. lactis strain, the
cells were incubated overnight in 50 ml M17 (Difco) medium
(supplemented with 10% glucose) at 30˚C without shaking.
Subsequently, 1 ml culture was used to inoculate 50 ml M17 medium
(10% glucose). The optical density (OD) was measured at 600 nm, and
the cells were incubated again until they reached an OD of 0.8. The
final bacterial concentration was 1x109 cells/ml.
Animals
Male Wistar rats (age, 6-8 weeks; weight, 150-250 g)
were obtained from the Laboratory Animal Service of the Autonomous
University of Aguascalientes (Aguascalientes, Mexico). The animals
were maintained on a light/dark cycle (12:12) with ad
libitum access to Purina® Rodent Chow (Cargill,
Inc.) and tap water. All animal experiments were approved by the
Research Ethics Committee of the Autonomous University of
Aguascalientes (approval no. A1-S-21375) and were conducted in
accordance with institutional guidelines for caring for
experimental animals and the national regulatory norm
(NOM-062-Z00-1999). To prepare the intestinal environment, all
animals were previously treated with neomycin sulfate and
sulfadimethylpyrimidine for 7 days. For experimentation, the rats
were divided into the following 4 groups (n=5 rats/group): i)
Control group not treated with L. lactis or CCl4;
ii) L. lactis group administered L. lactis; iii)
L. lactis-CCl4 group orally treated with L.
lactis and induced with CCl4; and iv)
CCl4 group intraperitoneally administered
CCl4 for cirrhosis induction. All animals were
sacrificed by overdose of sodium pentobarbital at 6 weeks, and
different sections of the liver, the terminal region of the ileum
(Peyer's Patches), the ascending portion of the large intestine and
serum samples were obtained. The tissue sections were preserved in
10% formalin in PBS at room temperature and RNAlater (Invitrogen;
Thermo Fisher Scientific, Inc.) at -80˚C for histological and
molecular analysis, respectively, and the serum was used to conduct
liver function tests.
Fibrotic induction
Fibrosis was induced in the L.
lactis-CCl4 and CCl4 groups. Based on a
pilot experiment in our laboratory, CCl4 was diluted in
petrolatum and intraperitoneally administered 3 times per week for
4 weeks as follows (CCl4:petrolatum by volume): Weeks 1,
1:6; 2, 1:5; 3, 1:4; 4, 1:3. These proportions were prepared
according to the number of experimental animals (n=5 for each
group) and the number of applications per week (3 per week).
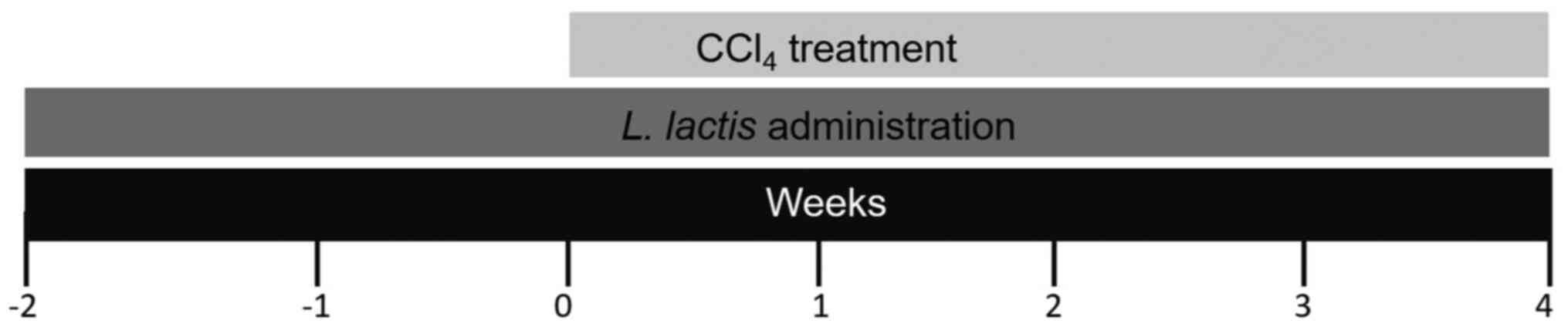
Administration of L. lactis
For 6 weeks, 1 ml L. lactis (1x109
cells), was orally administered to the L. lactis and L.
lactis-CCl4 groups on a daily basis. In the L.
lactis-CCl4 group, the probiotic was administered
two weeks prior to CCl4, and was subsequently continued
for an additional 4 weeks together with CCl4. For the
L. lactis group, the probiotic was administered alone for 6
weeks as a control (Fig. 1).
Histological analysis
Liver damage and Peyer's patches were evaluated
histologically by light microscopy. Sirius red staining (with
polarized light microscopy) was used to identify collagen fiber
deposits (type I, red; type III, green). For histopathological
analysis of Peyer's patches. The tissue sections were preserved in
10% formalin in PBS at room temperature for 24 h, and transverse
cuts of 5-µm thickness were made in the terminal portion of the
ileum to reveal clusters of lymphatic tissue (lymph nodes) that
cover the lamina propria, which were then stained with hematoxylin
and eosin. The histological preparations were visualized under a
Axioskop 40/40 FL light polarized microscope (Carl Zeiss AG) and
analyzed using Image-Pro Plus Software 4.5.1 (Media Cybernetics,
Inc.). The percentage of fibrosis was determined as the ratio of
the fibrotic area to the total tissue area.
Markers of liver damage
To determine the degree of liver damage, serum
levels of glucose (cat. no. BSIS19-P), albumin (cat. no. BSIS02-E),
bilirubin (cat. no. BSIS92-I) total protein (cat. no. BSIS30-E),
urea (cat. no. BSIS35-I), alanine aminotransferase (ALT; cat. no.
BEIS11-E) and aspartate transaminase (AST; cat. no. BEIS09-E) were
quantified. Kits for all biochemical tests were obtained from
Spinreact SAU. Each test was performed according to the
manufacturer's instructions. The samples were read on a BTS-350
semi-automatic spectrophotometric analyzer (BioSystems S.A.).
Total RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was isolated from 100 mg liver tissues
using the SV Total RNA Isolation System (Promega Corporation)
according to the manufacturer's protocol. The RNA was quantified
using NanoDrop-2000 (NanoDrop Technologies; Thermo Fisher
Scientific, Inc.) and stored at -80˚C until required. Reverse
transcription was performed with 1 µg total RNA using the GoScript™
Reverse Transcription System (Promega Corporation) according to the
manufacturer's instructions. Subsequently, qPCR was performed using
the qPCR GreenMaster with UNG-clear (Jena Bioscience GmbH) using
StepOne™ equipment (Applied Biosystems; Thermo Fisher Scientific,
Inc.) with the following thermocycling conditions: 50˚C for 2 min
and 95˚C for 45 sec, followed by 40 cycles of 95˚C for 45 sec and
60˚C for 45 sec. The oligonucleotide primers are displayed in
Table I. Relative expression levels
were normalized to those of β-actin, and the differences were
determined using the 2-ΔΔCq method (29).
 | Table IPrimers used for quantitative
PCR. |
Table I
Primers used for quantitative
PCR.
| Primer | Sequence
(5'→3') |
|---|
| FoxP3 | F:
CGGGAGAGTTTCTCAAGCAC |
| | R:
CACAGGTGGAGCTTTTGTCA |
| IL-1β | F:
CTGTGACTCGTGGGATGATG |
| | R:
GGGATTTTGTCGTTGCTTGT |
| IL-10 | F:
TGGCTCAGCACTGCTATGTT |
| | R:
TTGTCCAGCTGGTCCTTCTT |
| β-actin | F:
GTCGTACCACTGGCATTGTG |
| | R:
GCTGTGGTGGTGAAGCTGTA |
Statistical analysis
GraphPad Prism 5.00 (GraphPad Software, Inc.) was
used for statistical analysis and figures. Data are presented as
the mean ± standard error of the mean for each group. Significant
differences between mean values were assessed using one-way ANOVA
followed by Tukey's test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Macroscopic and histopathological
analysis of the livers of control and treated animals
At a macroscopic level, the livers of the control
and L. lactis groups exhibited a smooth surface and the
classic dark brown color of a healthy liver (Fig. 2A and D). By contrast, the liver tissues of the
CCl4 group were rough and irregular, with a lighter
brown color (Fig. 2G). The livers
of the rats om the L. lactis-CCl4 presented with
a similar coloration and texture to those of the control group
(Fig. 2J). At the microscopic level
(magnification, x10 and x40), the control and L. lactis
groups displayed classic liver lobules, with hepatocytes and normal
hepatic sinusoids (yellow arrows) that did not affect the liver
histology (Figs. 2C and F). In the CCl4 groups,
steatosis (black arrows) and pyknotic nuclei (black asterisk) were
observed in zone I of the liver acini (Fig. 2I). In the L.
lactis-CCl4 group (magnification, x10), a small
number of hepatocytes in zone II presented with
CCl4-induced damage (to a lesser degree compared with
that in the CCl4 group) and a smaller area of steatosis,
described as a microvesicular type (black arrowheads) compared with
the CCl4 group. Additionally, at x40 magnification,
acidophilic cells were observed with a larger cytoplasm; it was
therefore speculated that these cells exhibited a degree of
incipient damage in the CCl4 and L.
lactis-CCl4 groups (yellow asterisk; Fig. 2I and L).
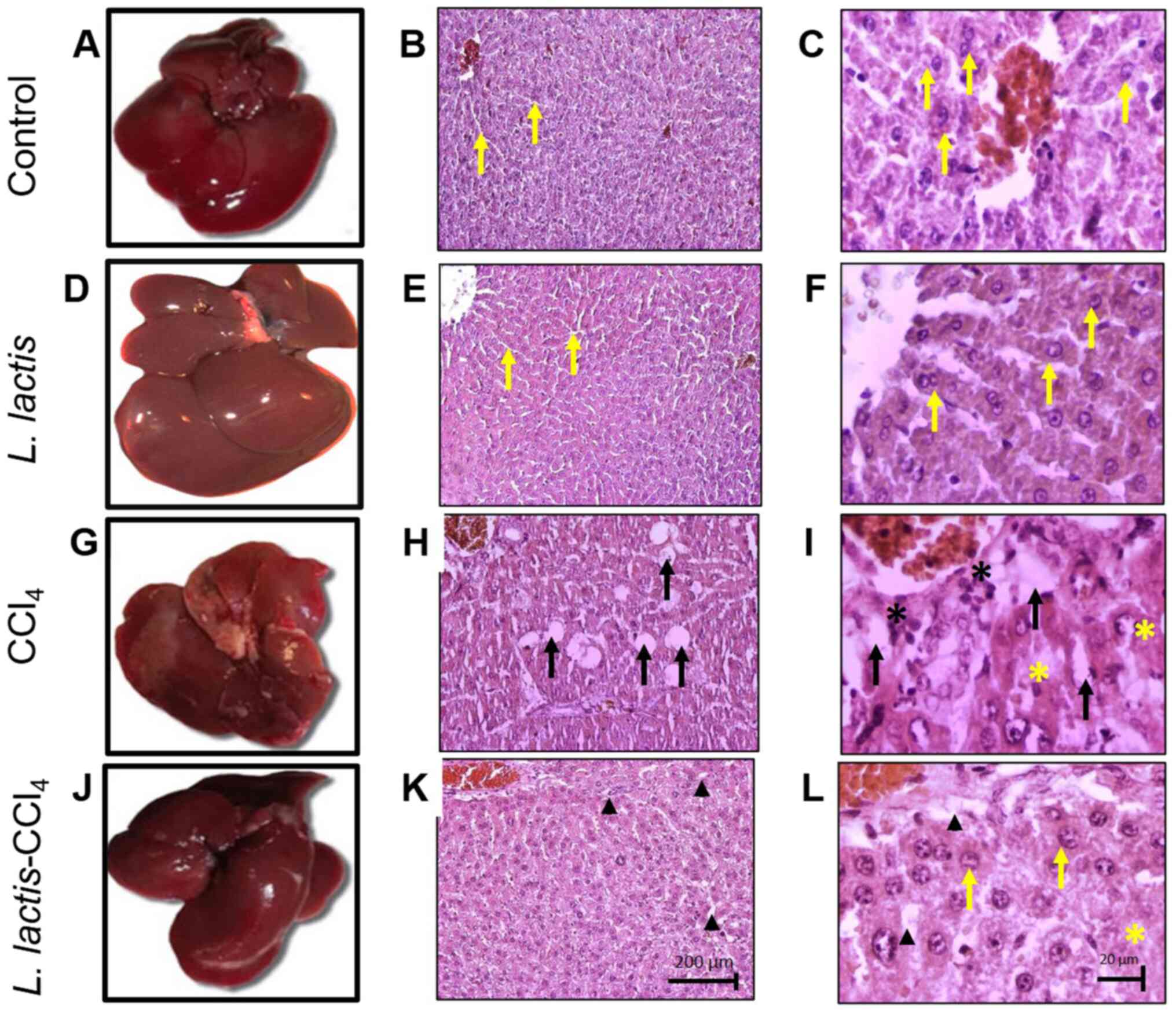 | Figure 2Oral administration of L. lactis
prevents hepatic damage. At the macroscopic level, the livers of
the (A) control and (D) L. lactis groups possessed a smooth surface
and brown color. The livers of the (G) CCl4 group exhibited a rough
and irregular surface, and those of the (J) L. lactis-CCl4 group
presented with normal coloration and texture. At the microscopic
level, in the (B and C) control and (E and F) L. lactis groups
(magnification, x10 and x40), hepatocytes and normal hepatic
sinusoids were observed (yellow arrows); (H) In the CCl4 group
(magnification, x10), wide area with microvesicular steatosis
(black arrows); (I) in the CCl4 group (magnification, x40),
steatotic cells (black arrows) and pyknotic nuclei (black asterisk)
were observed. (K) In the L. lactis-CCl4 group (magnification,
x10), a small number steatotic cells of the incipient
microvesicular type were apparent (black arrowhead); (L) at x40
magnification, cells exhibited a greater proportion of acidophilus
cells with larger cytoplasm (yellow asterisks), suggesting a degree
of incipient damage in the CCl4 andL. lactis-CCl4 groups. |
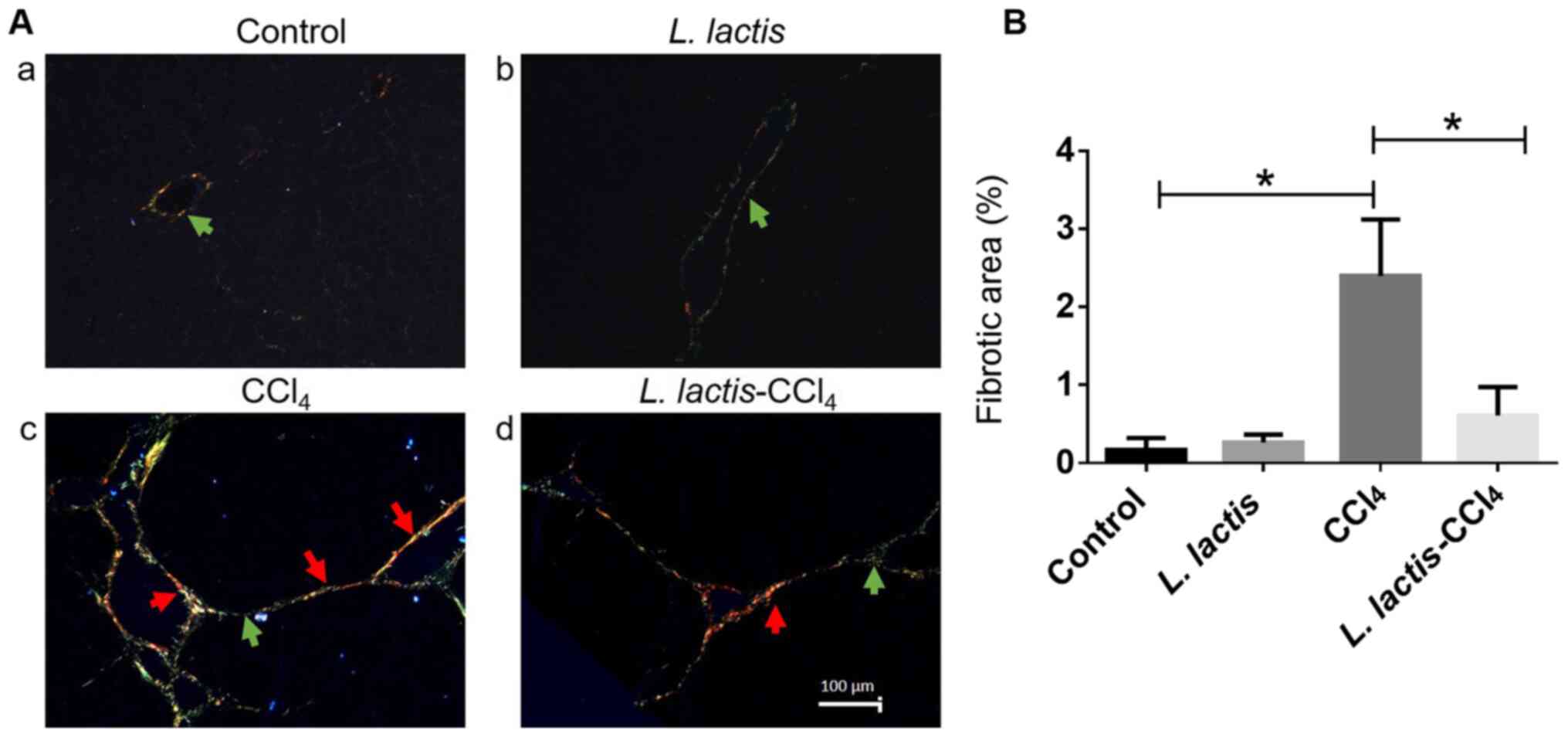
The liver sections stained with Sirius Red and
analyzed under a polarized light microscope exhibited normal
histological architecture in the control and L. lactis
groups, and type III collagen was observed (green arrow; Fig. 3A-a and b). In the CCl4 group, an
increase in type I collagen (red) was evident around blood vessels
(red arrows; Fig. 3A-c) along with
a low level of type III collagen (green arrow), indicating a
fibrotic lesion. By contrast, a significant decrease in type I
collagen fibers was observed in the L.
lactis-CCl4 group (red arrow; Fig. 3A-d). To confirm the degree of
fibrosis, a morphometric analysis of the hepatic parenchyma was
performed; an increase in collagen fibers was evident in the
CCl4 group compared with that in the control group
(P<0.001; Fig. 3B). In addition,
the percentage of total collagen was lower in the L.
lactis-CCl4 group compared with the CCl4
group.
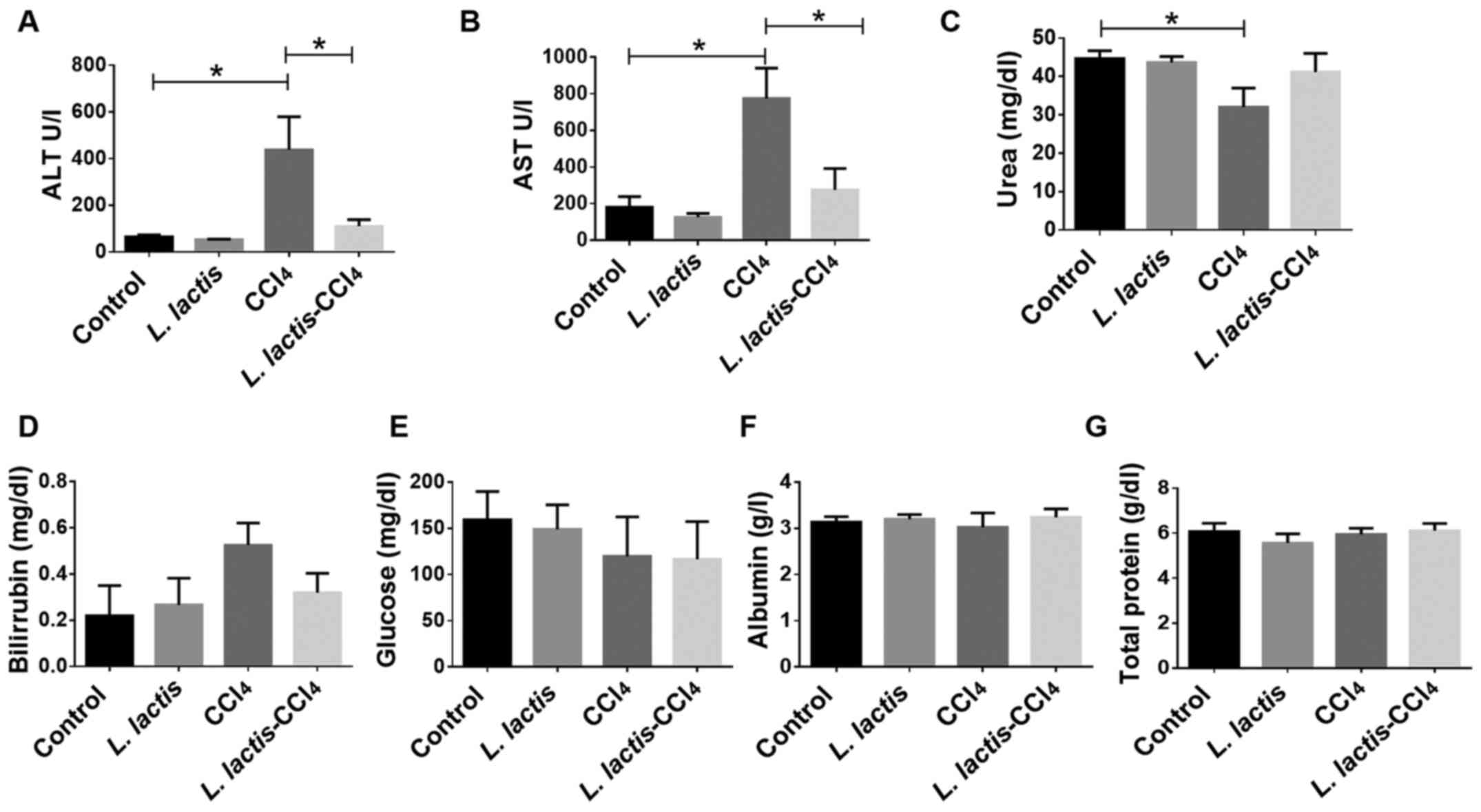
Liver Function
Liver function was evaluated by the quantification
of albumin, glucose, bilirubin and total proteins, and no
significant differences were observed (Fig. 4). However, increased plasma ALT and
AST (indicators of liver damage) levels were observed following
induction with CCl4. The L.
lactis-CCl4 group exhibited a significant decrease
in ALT and AST levels compared with those in the CCl4
group (P<0.001; Fig. 4A and
B), indicating that L.
lactis improved liver function. Urea is primarily formed in the
liver as an end product of protein metabolism (6,11); a
significant decrease in the urea level was observed in the
CCl4 group compared with that in the control group
(P<0.05; Fig. 4C), whereas the
L. lactis-CCl4 group presented with similar
levels to those of the control group, which suggested that the
liver was functionally transforming ammonium to urea for excretion.
The recovery of liver functional enzymes may be associated with the
histological improvement presented in Fig. 2. No changes in hepatic function were
observed in the L. lactis group compared with the control
group.
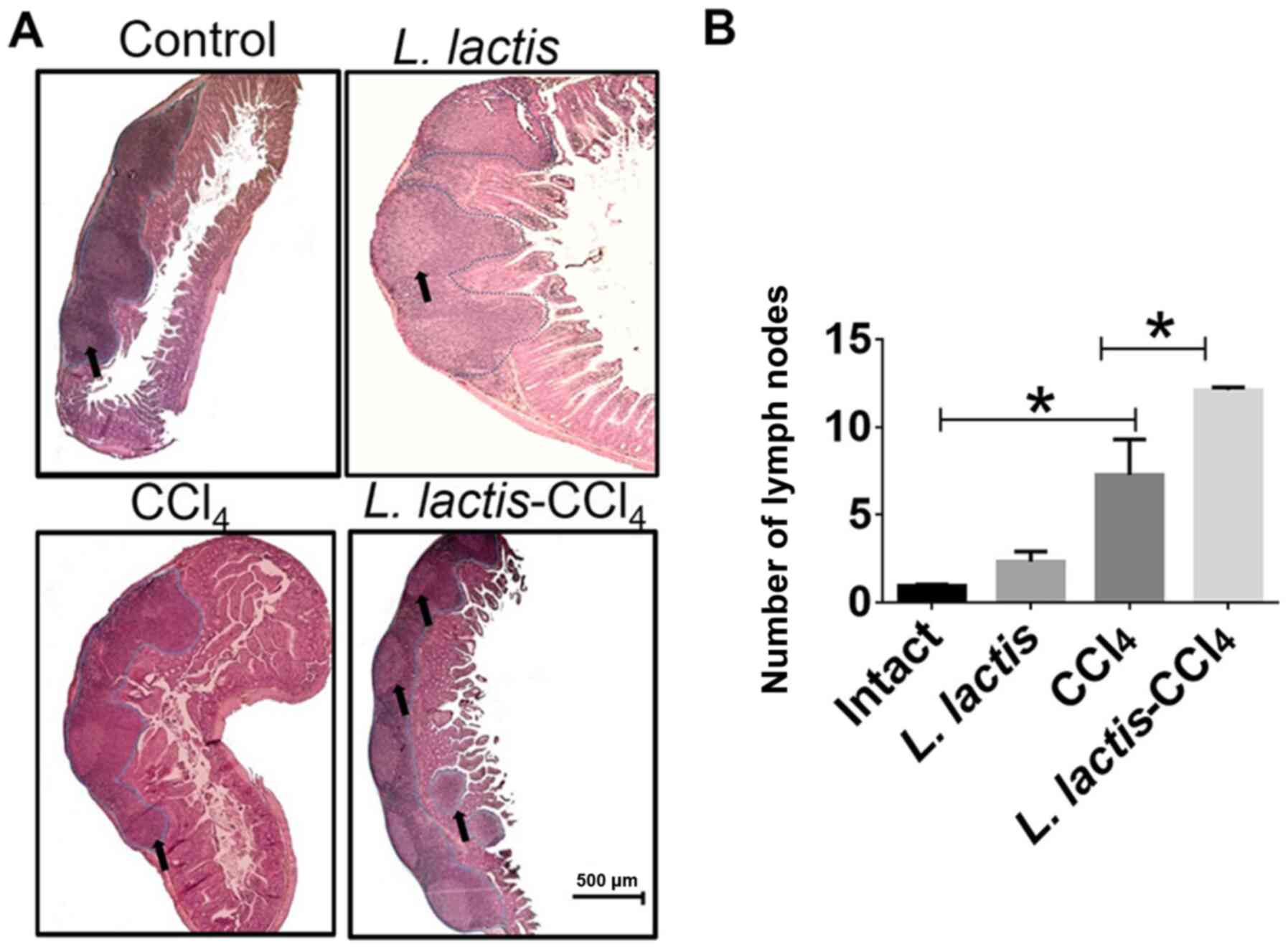
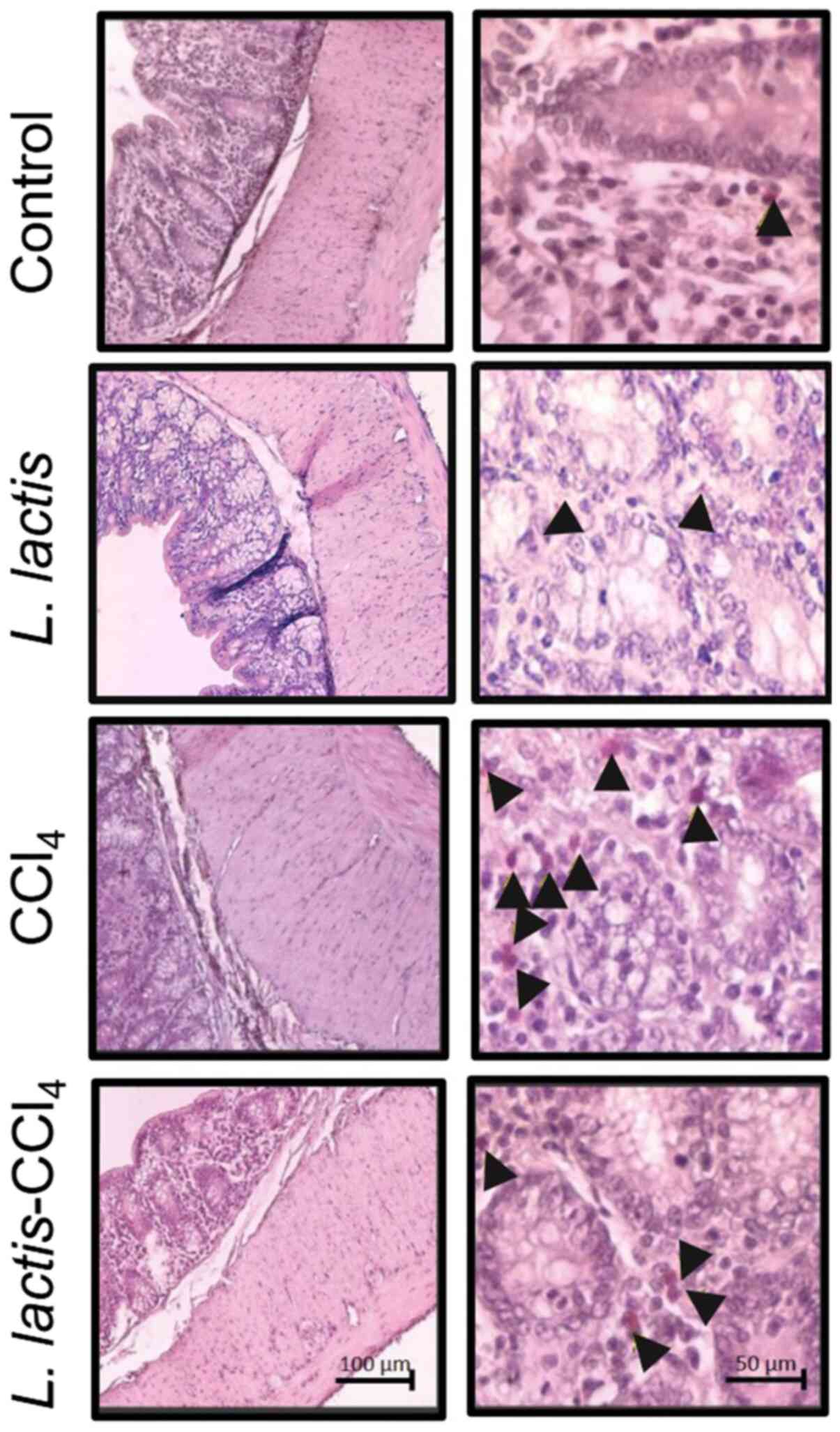
Histopathological analysis of Peyer's
Patches
The L. lactis-CCl4 group induced
~8 well-defined nodules in a single tissue portion (black arrows;
Fig. 5A); however, the control and
CCl4 groups possessed a mean of 3 nodules (P<0.01),
which were smaller in size. Although few lymphoid nodules were
observed in the L. lactis group, these were larger than
those in the control group (Fig. 5A
and B). To corroborate these size
variations, a morphometric analysis was performed, and no
significant differences were apparent between any of the groups.
However, the L. lactis-CCl4 group exhibited the
largest area of these nodules (Fig.
5B).
Histopathological analysis of the
large intestine (cecum)
Transverse cuts of the cecum were made in animals
from each of the study groups (Fig.
6). Normal histology was observed in the control group;
however, the colonic tissue of the CCl4 group presented
with cellular infiltrate (black arrowheads) in the region of the
mucosa layer. This infiltrate was diminished in the CCl4
group treated with L. lactis.
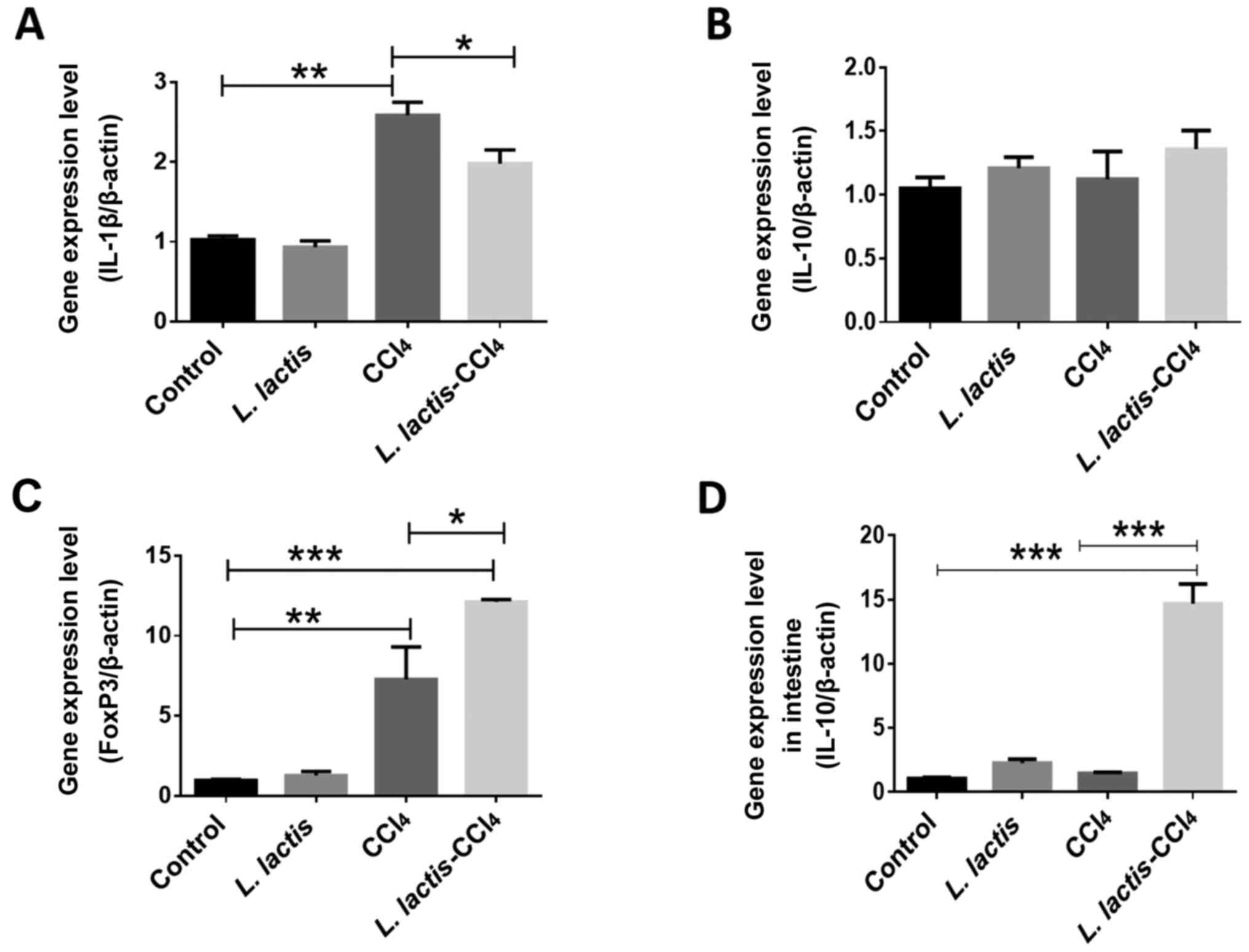
Evaluation of inflammatory markers in
the liver
L. lactis is known to have an
immunomodulatory effect due to its association with IL-10, a potent
anti-inflammatory cytokine that represses the expression of
inflammatory cytokines such as TNF-α, IL-6 and IL-1β produced by
macrophages activated during liver injury (30). To analyze the possible effect of
intestinal L. lactis on CCl4-induced liver
damage, the levels of specific cytokines were assessed in the liver
tissue, such as pro-inflammatory IL-1β, the anti-inflammatory IL-10
and a T-cell regulatory transcription factor forkhead box protein
P3 (FoxP3) (Fig. 7A-C); IL-10
expression was also assessed in intestinal tissues; in the L.
lactis-CCl4 group, intestinal IL-10 expression was
increased compared with the control group (P<0.001), and
CCl4 groups (P<0.001), (Fig. 7D). No significant differences in
IL-10 expression were observed among any of the experimental
groups, although there was a non-significant tendency towards
higher expression in the L. lactis-CCl4 group. IL-1β
expression was decreased in the liver tissues (P<0.05). FoxP3 is
the primary regulator of the development and function of regulatory
T cells, and its expression was increased in the L.
lactis-CCl4 group compared with that in the
CCl4 group (P<0.05). Collectively, these results
demonstrated the immunoregulatory effects of intestinal L.
lactis on hepatic pathology.
Discussion
In the present study, the inhibitory effect of L.
lactis NZ9000 on CCl4-induced hepatic fibrosis was
analyzed. Oral administration of L. lactis induced a
physiological change and modified the development of
CCl4-induced fibrosis in the liver tissue in the
following manners: i) Reducing structural liver damage; ii)
reducing the area of fibrosis; iii) increasing the number of lymph
nodes in the Peyer's patches; iv) decreasing ALT and AST
expression; v) increasing the mRNA expression of IL-10 in small
intestine samples; vi) increasing FoxP3 levels in liver samples;
and vii) decreasing the expression of IL-1β in the liver.
The aim of the present study was to investigate a
protective strategy to reverse liver damage using the physiological
interconnection between the digestive tract and the liver via the
hepatic portal system (31). The
human microbiome is defined as the collective genome of >1,000
different types of microorganisms that exist in association with
the human body, the vast majority of which reside in the distal
intestine (32,33). This ecological system interacts with
internal and external organs, factors that help to maintain the
overall health of the individual (12). Taking advantage of this
physiological association, anatomical-functional communication was
investigated with the aim to induce an immunoregulatory response in
the intestine, with ultimate effects on liver inflammation.
In cases of colitis, the L. reuteri R2LC
strain has been demonstrated to exert an anti-inflammatory effect
in the large intestine (13,14,16,17).
Different experimental animal models (predominantly of colitis)
have demonstrated the benefits of probiotics in controlling
intestinal inflammation. In an acetic acid-induced rat colitis
model, the administration of L. reuteri R2LC immediately
after induction prevented the establishment of colitis (16). Previous reports have demonstrated
the effect of probiotics on the gut microbiota under inflammatory
process. The oral administration of L. plantarum attenuated
inflammatory bowel disease in a mouse model; L. plantarum
also affected the proportion of Firmicutes and
Bacteroides, which may be associated with the inflammation
of the mouse gut (34). The
intestinal microbiota has been reported to serve a fundamental role
in homeostatic maintenance of the systemic immune system; for
example, L. johnsonii of an intestinal origin did not induce
the release of TNF-α or IL-1β following downregulation of the
transcription factor NF-κB, whereas TGF-β expression was increased,
resulting in a global anti-inflammatory profile (15). Specific recognition of commensal
microorganisms occurs in the mesenteric lymph nodes. Most antigens
or infectious agents pass into the venous system or tissues through
mucous membranes, which includes the lining of the
gastrointestinal, respiratory and genitourinary tracts. At these
mucosal surfaces, the mucus represents the first barrier against
the entry of microorganisms, while gut-associated lymphoid tissues
(GALT), which include intestinal Peyer's patches, is critical for
efficient protective immune response, making the GALT an attractive
portion of the small intestine to study (35). In the present study, the probiotic
L. lactis generated an anti-inflammatory environment in the
small intestine by increasing the expression of IL-10, as well as
the number of lymph nodes in the Peyer's patches; these findings
suggested a stimulus that may potentially increase the number of
regulatory, anti-inflammatory lymphoid cells. However, one of the
limitations of the present study was not determining whether L.
lactis may influence the proportions of various taxonomic and
functional groups of the gut microbiota, which may increase our
knowledge about the mechanisms of action of probiotics.
IL-10 decreases and regulates dendritic cell- and
macrophage-associated inflammatory responses by activating
STAT-3(14). IL-10 also suppresses
the adaptive immune response by inhibiting NF-κB secretion by
CD4+ T cells and the production of IL-1 and TNF-α by
macrophages (14). The results of
the present study revealed a notable decrease in hepatic IL-1β
expression in the L. lactis-CCL4 group compared
with the CCl4 group; this was potentially due to an
increase in intestinal IL-10 as a result of CCl4-induced
damage, which was subsequently transported to the liver via the
portal-hepatic system. Additionally, an increase in the expression
of Foxp3 mRNA was observed in the liver, which supports the
increase in intestinal IL-10 in the L.
lactis-CCL4 group, and may been affected by the
downregulation of NF-κB (36,37).
This supports the existence of an immunoregulatory process induced
by L. lactis in the intestine, which has an inhibitory
effect on CCl4-associated fibrosis; thus, L.
lactis may exert an anti-fibrotic effect in the early stages of
inflammation, which potentially modifies the adhesion properties of
epithelial cells, altering the local host immune response (1,38). For
this reason, potential new therapies for liver fibrosis may target
the recovery of the microbiota to reduce the possible adverse
effects associated with pharmacological treatment (18). L. lactis may therefore be an
optional co-treatment for decreasing inflammation in the early
stages of cirrhosis.
In conclusion, L. lactis prevented liver
damage in an animal model of CCl4-induced liver
fibrosis. The results of the present study suggested that oral
administration of L. lactis in its native form may be a
potential means to prevent and protect against
CCl4-induced liver damage.
Acknowledgements
We acknowledge to MVZ Karen Estefany Sánchez
Hernández, Central Bioterium, Autonomous University of
Aguascalientes, for providing and caring for the animals for this
study; and LAQB Cintya Esquivel-Dueñas and LAQB Mariana
Perez-Villalobos, Department of Chemistry, Autonomous University of
Aguascalientes, for their excellent technical assistance.
Funding
Funding: The present study was supported by CONACYT (grant nos.
241312 and A1-S-21375) and the Autonomous University of
Aguascalientes (grant no. PIBB18-8). CSDV gratefully acknowledges
the financial fellowship granted by CONACYT (grant no. 617146) as
part of the master's program in Toxicology Sciences of the
Autonomous University of Aguascalientes.
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
SLMH contributed to the analysis and interpretation
of the histological data. CSDV developed the histological technique
for the intestine and liver tissues. DCG performed reverse
transcription-quantitative PCR analysis. RMDOL and MDJLA donated
the L. lactis cultures and analyzed the quantitative PCR
data. MGMM developed the liver function study. JVJ and MHMO
contributed to study conception and design, and the writing and
revision of the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the
Research Ethics Committee of the Autonomous University of
Aguascalientes (approval no. A1-S-21375) and were conducted in
accordance with institutional guidelines for caring for
experimental animals and the national regulatory norm
(NOM-062-Z00-1999).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Friedman SL: Mechanisms of hepatic
fibrogenesis. Gastroenterology. 134:1655–1669. 2008.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Wick G, Backovic A, Rabensteiner E, Plank
N, Schwentner C and Sgonc R: The immunology of fibrosis: Innate and
adaptive responses. Trends Immunol. 31:110–119. 2010.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Wick G, Grundtman C, Mayerl C,
Wimpissinger TF, Feichtinger J, Zelger B, Sgonc R and Wolfram D:
The immunology of fibrosis. Annu Rev Immunol. 31:107–135.
2013.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Wynn TA: Cellular and molecular mechanisms
of fibrosis. J Pathol. 214:199–210. 2008.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Kershenobich Stalnikowitz D and Weissbrod
AB: Liver fibrosis and inflammation. A review. Ann Hepatol.
2:159–163. 2003.PubMed/NCBI
|
|
6
|
Romanelli RG and Stasi C: Recent
advancements in diagnosis and therapy of liver cirrhosis. Curr Drug
Targets. 17:1804–1817. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Khan H, Ullah H and Nabavi SM: Mechanistic
insights of hepatoprotective effects of curcumin: Therapeutic
updates and future prospects. Food Chem Toxicol. 124:182–191.
2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Serna-Salas SA, Navarro-González YD,
Martínez-Hernández SL, Barba-Gallardo LF, Sánchez-Alemán E,
Aldaba-Muruato LR, Macías-Pérez JR, Ventura-Juárez J and
Muñoz-Ortega MH: Doxazosin and carvedilol treatment improves
hepatic regeneration in a hamster model of cirrhosis. Biomed Res
Int. 2018(4706976)2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Khawar MB, Azam F, Sheikh N and Abdul
Mujeeb K: How does interleukin-22 mediate liver regeneration and
prevent injury and fibrosis? J Immunol Res.
2016(2148129)2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Abd-Elgawad H, Abu-Elsaad N, El-Karef A
and Ibrahim T: Piceatannol increases the expression of hepatocyte
growth factor and IL-10 thereby protecting hepatocytes in
thioacetamide-induced liver fibrosis. Can J Physiol Pharmacol.
94:779–787. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Bajaj JS, Heuman DM, Hylemon PB, Sanyal
AJ, White MB, Monteith P, Noble NA, Unser AB, Daita K, Fisher AR,
et al: Altered profile of human gut microbiome is associated with
cirrhosis and its complications. J Hepatol. 60:940–947.
2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Moraes-Filho JP and Quigley EM: The
intestinal microbiota and the role of probiotics in irritable bowel
syndrome: A review. Arq Gastroenterol. 52:331–338. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Festen EAM, Szperl AM, Weersma RK,
Wikmenga C and Wapenaar MC: Inflammatory bowel disease and celiac
disease: Overlaps in the pathology and genetics, and their
potential drug targets. Endocr Metab Immune Disord Drug Targets.
9:199–218. 2009.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lee NK, Kim SY, Han KJ, Eom SJ and Paik
HD: Probiotic potential of Lactobacillus strains with
anti-allergic effects from kimchi for yogurt starters. LWT Food Sci
Technol. 58:130–134. 2014.
|
|
15
|
Haller D, Bode C, Hammes WP, Pfeifer AM,
Schiffrin EJ and Blum S: Non-pathogenic bacteria elicit a
differential cytokine response by intestinal epithelial
cell/leucocyte co-cultures. Gut. 47:79–87. 2000.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Fabia R, Ar'Rajab A, Johansssib ML, Willén
R, Andersson R, Molin G and Bengmark S: The effect of exogenous
administration of Lactobacillus reuteri R2LC and oat fiber
on acetic acid-induced colitis in the rat. Scand J Gastroenterol.
28:155–162. 1993.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Mao Y, Nobaeck S, Kasravi B, Adawi D,
Stenram U, Molin G and Jeppsson B: The effects of Lactobacillus
strains and oat fiber on methotrexate-induced enterocolitis in
rats. Gastroenterology. 111:334–344. 1996.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Bhat M, Arendt BM, Bhat V, Renner EL,
Humar A and Allard JP: Implication of the intestinal microbiome in
complications of cirrhosis. World J Hepatol. 8:1128–1136.
2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Parapouli M, Delbès-Paus C, Kakouri A,
Koukkou AI, Montel MC and Samelis J: Characterization of a wild,
novel nisin a-producing Lactococcus strain with an L.
lactis subsp. cremoris genotype and an L. lactis
subsp. lactis phenotype, isolated from Greek raw milk. Appl Environ
Microbiol. 79:3476–3484. 2013.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Duwat P, Sourice S, Cesselin B, Lamberet
G, Vido K, Gaudu P, Le Loir Y, Violet F, Loubière P and Gruss A:
Respiration capacity of the fermenting bacterium Lactococcus
lactis and its positive effects on growth and survival. J
Bacteriol. 183:4509–4516. 2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Garrigues C, Loubiere P, Lindley ND and
Cocaign-Bousquet M: Control of the shift from homolactic acid to
mixed-acid fermentation in Lactococcus lactis: Predominant
role of the NADH/NAD+ ratio. J Bacteriol. 179:5282–5287.
1997.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Daniel C, Repa A, Wild C, Pollak A, Pot B,
Breiteneder H, Wiedermann U and Mercenier A: Modulation of allergic
immune responses by mucosal application of recombinant lactic acid
bacteria producing the major birch pollen allergen Bet v 1.
Allergy. 61:812–819. 2006.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Alvarenga DM, Perez DA, Gomes-Santos AC,
Miyoshi A, Azevedo V, Coelho-Dos-Reis JG, Martins-Filho OA, Faria
AM, Cara DC and Andrade MC: Previous ingestion of Lactococcus
lactis by ethanol-treated mice preserves antigen presentation
hierarchy in the gut and oral tolerance susceptibility. Alcohol
Clin Exp Res. 39:1453–1464. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Marinho FA, Pacífico LG, Miyoshi A,
Azevedo V, Le Loir Y, Guimarães VD, Langella P, Cassali GD, Fonseca
CT and Oliveira SC: An intranasal administration of Lactococcus
lactis strains expressing recombinant interleukin-10 modulates
acute allergic airway inflammation in a murine model. Clin Exp
Allergy. 40:1541–1551. 2010.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Nishitani Y, Tanoue T, Yamada K, Ishida T,
Yoshida M, Azuma T and Mizuno M: Lactococcus lactis subsp.
cremoris FC alleviates symptoms of colitis induced by
dextran sulfate sodium in mice. Int Immunopharmacol. 9:1444–1451.
2009.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Luerce TD, Gomes-Santos AC, Rocha CS,
Moreira TG, Cruz DN, Lemos L, Sousa AL, Pereira VB, de Azevedo M,
Moraes K, et al: Anti-inflammatory effects of Lactococcus
lactis NCDO 2118 during the remission period of chemically
induced colitis. Gut Pathog. 6(33)2014.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Ballal SA, Veiga P, Fenn K, Michaud M, Kim
JH, Gallini CA, Glickman JN, Quéré G, Garault P, Béal C, et al:
Host lysozyme-mediated lysis of Lactococcus lactis
facilitates delivery of colitis-attenuating superoxide dismutase to
inflamed colons. Proc Natl Acad Sci USA. 112:7803–7808.
2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Steidler L, Hans W, Schotte L, Neirynck S,
Obermeier F, Falk W, Fiers W and Remaut E: Treatment of murine
colitis by Lactococcus lactis secreting interleukin-10.
Science. 289:1352–1355. 2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Williams LM, Ricchetti G, Sarma U, Smallie
T and Foxwell BM: Interleukin-10 suppression of myeloid cell
activation-a continuing puzzle. Immunology. 113:281–292.
2004.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Aller MA, Vara E, Garcia C, Palma MD,
Arias JL, Nava MP and Arias J: Proinflammatory liver and
antiinflammatory intestinal mediators involved in portal
hypertensive rats. Mediators Inflamm. 2005:101–111. 2005.PubMed/NCBI View Article : Google Scholar
|
|
32
|
US National Institutes of Health: NIH
human microbiome project. https://www.hmpdacc.org/overview/.
|
|
33
|
Kibe R, Sakamoto M, Yokota H, Ishikawa H,
Aiba Y, Koga Y and Benno Y: Movement and fixation of intestinal
microbiota after administration of human feces to germfree mice.
Appl Environ Microbiol. 71:3171–3178. 2005.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Chen H, Xia Y, Zhu S, Yang J, Yao J, Di J,
Liang Y, Gao R, Wu W, Yang Y, et al: Lactobacillus plantarum
LP-nlly alters the gut flora and attenuates colitis by inducing
microbiome alteration in interleukin-10 knockout mice. Mol Med Rep.
16:5979–5985. 2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Macpherson AJ and Uhr T: Induction of
protective IgA by intestinal dendritic cells carrying commensal
bacteria. Science. 303:1662–1665. 2004.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Liu X, Lou J, Chen Y and Duan Z: Changes
of regulatory T cells related to CCl4-induced liver
fibrosis in mice. Zhonghua Gan Zang Bing Za Zhi. 22:277–280.
2014.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
37
|
Rios DA, Valva P, Casciato PC, Frias S,
Soledad Caldirola M, Gaillard MI, Bezrodnik L, Bandi J, Galdame O,
et al: Chronic hepatitis C liver microenvironment: Role of the
Th17/Treg interplay related to fibrogenesis. Sci Rep.
7(13283)2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Schiffin EJ, Brassart D, Servin AL, Rochat
F and Donnet-Hughes A: Immune modulation of blood leukocytes in
humans by lactic acid bacteria: Criteria for strain selection. Am J
Clin Nutr. 66:515S–520S. 1997.PubMed/NCBI View Article : Google Scholar
|