Introduction
Acute liver injury represents a stage of sudden
deterioration of liver function that is characterized by cell
necrosis, inflammation and oxidative damage (1). Hepatic fibrosis refers to the
excessive deposition of diffuse extracellular matrix (ECM) in liver
tissue, which is the repair response in organisms to chronic liver
injury (2). The accumulation of ECM
results in hepatic fibrosis and hepatocellular carcinoma, and can
increase the mortality rate of liver diseases (3). Thus, ECM has direct clinical
implications to treat and mitigate acute and chronic liver injury.
Transforming growth factor-β1 (TGF-β1) is the most effective
activator of hepatic stellate cells (HSC) in liver fibrosis, acting
via the Smads pathway (4).
Different measures to block and/or regulate TGF-β/Smad signal
transmission may be an important strategy to prevent hepatic
fibrosis (5). The Smads family have
three functional domains, N-terminal Mad homology 1, C-terminal Mad
homology 2 and intermediate linker region (6). Phosphorylated (p)Smads at the
COOH-terminal or linker region form different complexes with Smad4,
which then translocate to the nucleus (6). In acute liver injury, TGF-β1 and
platelet-derived growth factor synergistically activate pSmad2C/L
and promote collagen synthesis in activated HSC (7). Furthermore, during chronic liver
disease progression, fibrogenic pSmad2C/L signaling was affected by
TGF-β1, which accelerates liver fibrosis (8). Moreover, it was reported that a
crosstalk between the nuclear factor erythroid-2-related factor 2
(Nrf2) and TGF-β1 pathways promotes the development of
hepatocellular carcinoma (9).
Previous studies have shown that Nrf2 negatively acts against
fibrotic TGF-β1 signaling (10),
and that TGF-β1 promotes the generation of reactive oxygen species
(ROS) by inhibiting Nrf2 and its target antioxidant enzymes, such
as heme oxygenase-1 (HO-1) and NAD(P)H dehydrogenase (quinone) 1,
during hepatic fibrosis (11).
Furthermore, the TGF-β/Smad signaling pathway also induces
activating transcription factor 3 (ATF3), which in turn complexes
with Nrf2 to suppress Nrf2 target gene expression (12).
Salvianolic acid B (Sal B) is extracted from the
root and rhizome of Salvia miltiorrhiza, which has
antioxidative and antifibrosis effects (13). In vivo experimentation has
shown that Sal B enhances the ability of antioxidant in liver
injury, eliminates ROS and inhibits the activation of HSC (14). In in vitro experiments, Sal B
treatment could downregulate the expression of α-SMA and collagen
type I in TGF-β1-induced HSC (15).
The authors' previous study found that Sal B exerts antihepatic
fibrosis effects via modulation of pSmad3C/L (16). However, it is not fully understood
how Sal B affects pSmad2C/L in acute and chronic liver injury, nor
its effect on the antioxidant pathway.
The aim of the present study was to investigate the
protective effect of Sal B in CCl4- and
H2O2-induced acute and chronic liver injury,
and to identify the possible underlying mechanisms. The present
study also assessed the effect of pSmad2C, pSmad2L and the
Nrf2/HO-1 pathway on liver injury pathogenesis.
Materials and methods
Chemicals
Sal B (cat. no. PS12091001; purity ≥95%) was
obtained from Chengdu Push Bio-Technology Co., Ltd; Bio-Equiop. Sal
B was initially separated and refined from the dried roots of
Salvia miltiorrhiza. CCl4 (purity ≥99.5%;
concentration, 10.4 mol/l), obtained from Shanghai Puyi Chemical
Reagent Co., Ltd. Commercial kits were used to determine the levels
of malondialdehyde (MDA; cat. no. A003-1-2), glutathione (GSH; cat.
no. A005-1-2) and superoxide dismutase (SOD; cat. no. A001-1-2)
were purchased from Jianchen Institute of Biotechnology.
Immunostaining Streptavidin-Peroxidase (SP) kit (cat. no. SP9001)
was obtained from OriGene Technologies, Inc.
Animals and experimental groups
A total of 60 Kunming male mice (weight 18-22 kg;
age 6-8 weeks) were purchased from Animal Center of Anhui Medical
University (Anhui, China). All mice were maintained in a 12 h
light/dark alternating cycle at a room temperature (23±2˚C) with a
humidity of 50±10%. The experiment was carried out after 1 week of
adaptive feeding. In both the acute and chronic liver injury
experiments, mice were randomized into five groups: Control group
(Control), model group (Model), CCl4 + Sal B 7.5 group
(Sal B 7.5), CCl4 + Sal B 15 group (Sal B 15) and
CCl4 + Sal B 30 group (Sal B 30) (n=6 in each group).
All mice were allowed free access to food and water. In the acute
liver injury experiment, mice in the control and model group were
given 0.9% saline daily, and the three Sal B groups were given Sal
B (7.5, 15 and 30 mg/kg, respectively) daily by intragastric
administration, for a duration of 1 week. Then, 24 h after the
final dose of Sal B, the control group mice were given olive oil
and the other groups received 0.1% CCl4 (dissolved in
olive oil) via single intraperitoneal injection (10 ml/kg)
(17). In the chronic liver injury
experiment, mice in the control and model group were given 0.9%
saline daily, and the three Sal B groups were given Sal B (7.5, 15
and 30 mg/kg, respectively) daily by intragastric administration
for 8 weeks. Control mice received olive oil and Sal B groups
received 20% CCl4 (dissolved in olive oil) by
intraperitoneal injection (1 ml/kg/2 weeks) for 8 weeks (18,19).
Then, 24 h after the last intraperitoneal injection of
CCl4, all mice were anesthetized in a chamber with 1.4%
isoflurane and 100% oxygen for 2 h. After retro-orbital collection
of blood under anesthesia, the mice were transferred to a face mask
providing 5% isoflurane. Mice were placed in sternal recumbency and
a surgical level of anesthesia was assessed by firm bilateral foot
pinch. When each mouse was fully anesthetized, euthanasia was
performed using cervical dislocation. Subsequently, the liver of
the mice was harvested.
All the experiments were conducted in accordance
with the guidelines for Ethical Review of laboratory animal welfare
in the Animal Center of Anhui Medical University. The animal
experiments were approved by the Experimental Animal Ethics
Committee of Anhui Medical University.
Cell culture and Sal B treatment
Rat hepatic stellate cells HSC-T6 were purchased
from Nanjing KeyGen Biotech. Co. Ltd. HSC-T6 cells were incubated
in 6 well plates (2x105 cells/well) for 24 h at 37˚C in
5% CO2 with DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Zhejiang Tianhang Biotechnology Co.,
Ltd.). Cells were treated with various final concentrations of Sal
B (25, 50 and 100 μmol/l). Then, 100 μM
H2O2 (Nanjing KeyGen Biotech Co., Ltd.) was
added to each well and incubated at 37˚C for 6 h. The control group
was treated a same volume of medium (20,21).
Biochemical index assay
Liver function is an index reflecting the liver
physiological status. Moreover serological alanine transaminase
(ALT) and aspartate aminotransferase (AST) are classical indicators
which were used to test liver function. A total of ~1 ml blood
sample were obtained prior to euthanizing the mice, for
determination of serum enzymes levels. ALT (cat. no. C009-2-1) and
AST (cat. no. C010-2-1) levels were determined using commercial
reagent kits (Nanjing Jiancheng Bioengineering Institute) with an
automatic biochemical analyzer following the manufacturer's
protocols.
Hematoxylin and eosin (HE), and Van
Gieson's (VG) staining
Fresh liver tissues were sliced into applicable
sections (5-10 mm), then fixed in 4% buffered paraformaldehyde at
room temperature for 1 week, dehydrated and embedded in paraffin.
These were then sectioned at 5 μm and stained with
hematoxylin (0.2%, 15 min) and eosin (5%, 1 min) for
histopathological analyses. VG staining (1.2% picric acid and 1%
poinsettia, 1 min) method was used to detect collagen fibers at
room temperature. Sections stained with HE (magnification, x400)
and VG (magnification, x100) were photographed under a light
microscope (Nikon 80i; Nikon Corporation). The quantitative
assessment of fibrotic changes in the liver tissues was evaluated
by HPIAS-1000 auto Medical Image Analyzing system.
Assay of oxidative biochemical
parameters
Fresh liver tissue from each group (n=6) was cleaned
with saline solution. Tissues were grinded into 10% tissue
homogenate, then centrifuged for 10 min at 4˚C and 1,509.3 x g, to
obtain the superntant. Subsequently, activities of SOD and the
contents of MDA and GSH in liver tissues were detected using
commercially kits according to the manufacturer's protocol (I). SOD
activity was determined by the hydroxylamine method, in which the
O2- anions oxidize hydroxylamine to the sulfite form,
generating a purple-red color with a color-developing agent. SOD
removes the O2-anions, which reduces sulfite formation
and the absorbance values can be used to quantify SOD activity. In
total, 10 μl liver tissue supernatant was added to
hydroxylamine hydrochloride buffer, xanthine and xanthine oxidase,
mixed and incubated at 37˚C for 40 min. Then, the chromogenic agent
was added. The optical density (OD) value of each group of the
sample was measured at a wavelength of 550 nm after zero correction
with distilled water, a standard curve was drawn and the activity
value of SOD was calculated. (II) GSH reduces
5-dithio-2-nitrobenzoic acid (DTNB) to 2-nitro-5-mercaptobenzoic
acid, which is yellow in an acidic environment and can be
quantified at 412 nm; the measured value is proportional to
glutathione content. In total, 100 μl liver tissue
supernatant was added to phosphate buffer containing 500 mM DTNB.
After 5 min, the color turned yellow and OD values of each group
were measured at 405 nm after zero correction. Standard curves were
established and glutathione levels in the samples were calculated.
(III) MDA concentration was determined by measuring the
concentration of thiobarbituric acid (TBA) reactive substances.
Firstly, 100 μl liver tissue supernatant was added in 4 ml
0.6% TBA solution. The mixture was reacted in boiling water bath
(100˚C) for 15 min, then centrifuged for 10 min at 1,509.3 x g and
25˚C after cooling and the supernatant was collected. The OD values
of each group were measured at a wavelength of 532 nm, zero
correction was performed used distilled water. (IV) Coomassie
Brilliant Blue can be used to determine protein content at 25˚C.
The dye combines with the protein, forming a cyan protein-pigment
conjugate, with maximum light absorption at 595 nm. The light
absorption value is proportional to the protein content. The
supernatant of 0.05 tissue homogenate was mixed with 1 ml Coomassie
brilliant blue solution. After 10 min, the OD value of each tube
was measured at a wavelength of 595 nm with a 1 cm light path.
Immunohistochemical analysis
Paraffin sections were deparaffinized in xylene and
rehydrated in different concentrations of ethanol (100, 90, 80 and
70%). Non enzymatic antigen retrieval was processed by heating the
4 μm sections to 121˚C in 0.01 M sodium citrate buffer (pH
6.0) for 10 min. Then, the sections were cooled, rinsed in TBST
(0.1% Tween-20) and incubated in methanol with 3%
H2O2 for 30 min at 37˚C to restrain
endogenous peroxidase activity. After rinsing three times, the
sections were blocked with 5% BSA at 37˚C for 30 min, then
incubated with rabbit anti Nrf2 (1:3,000; cat. no. ab137550; Abcam)
for 1 h at room temperature in a humid chamber. Liver sections were
then rinsed in TBST and incubated with horseradish
peroxidase-labeled goat anti rabbit IgG polymer (1:3,000; cat. no.
ab137550; Abcam) at 37˚C for 1 h. The results of Nrf2 expression
levels in liver tissues were evaluated using a semi quantitative
technique. At the same light intensity and magnification, x 400,
eight visual fields were randomly taken using a light microscope
and analyzed by Image-Pro Plus (version 4.1; Media Cybernetics,
Inc.), and their mean density was measured. The mean value was
taken to represent the level of protein expression.
Western blot analysis
Total proteins from frozen liver tissue specimens
and/or HSC-T6 cells were extracted with RIPA lysis buffer (Beyotime
Institute of Biotechnology) according to the manufacturer's
protocol. The Bradford assay was used to determine the protein
concentration of samples. Then, equal amounts (8 µg) of each
protein sample was separated on 10% SDS-PAGE, transferred onto PVDF
membranes that were then blocked with 5% skimmed milk at room
temperature for 1 h. After blocking, the membranes were incubated
overnight at 4˚C with primary antibodies, followed by incubation
with horseradish peroxidase-conjugated goat anti-rabbit secondary
antibody (1:5,000; cat. no. 7074; Cell Signaling Technology, Inc.)
at 37˚C for 45 min. Primary antibodies included rabbit anti-GAPDH
(1:1,000; cat. no. 5174; Cell Signaling Technology, Inc.), rabbit
anti-Nrf2 (1:3,000; cat. no. ab137550), rabbit anti-HO-1 (1:2,000;
cat. no. ab13243) both from Abcam, rabbit anti-p-Smad2C (1:1,000;
cat. no. 18338) and rabbit anti-p-Smad2C (1:1,000; cat. no. 3104)
both from Cell Signaling Technology, Inc. Then, the proteins bands
were visualized with the ECL chemiluminescence system (GE
Healthcare). The density of each protein blot was compared with
that of GAPDH using ImageJ software (version 1.46r; National
Institutes of Health) and was shown as a ratio to the endogenous
control. These experiments were repeated three times.
Statistical analysis
Data are presented as the mean ± SD, n=6.
Statistical analysis were performed using SPSS 22 (IBM Corp.).
Differences between groups were compared using ANOVA followed by
Tukey's post hoc test. Data represent ≥3 independent experiments.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effect of Sal B on acute and chronic
liver injury
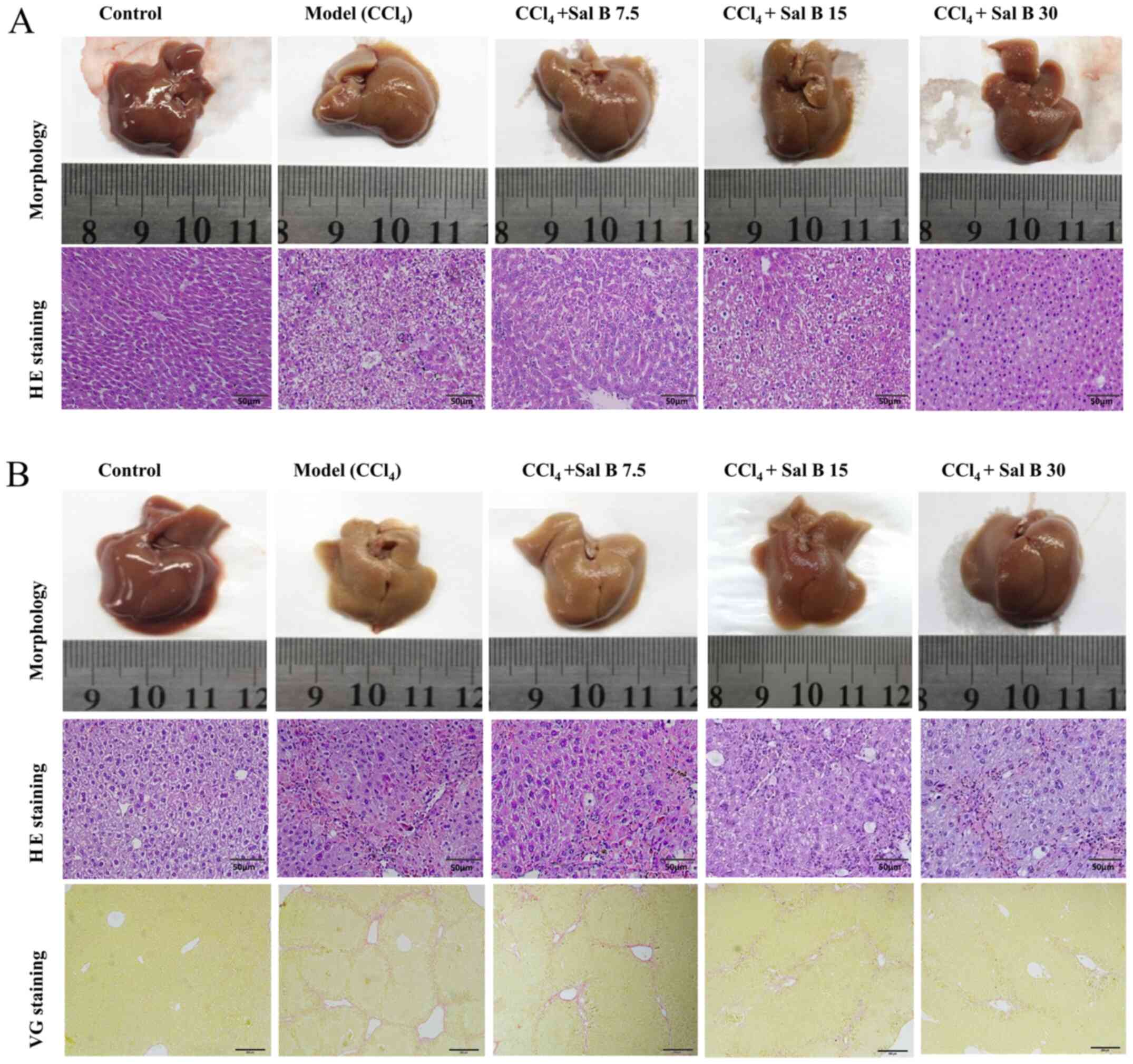
In acute liver injury, it was found that hepatocytes
in the CCl4 model group were irregular with obvious
pathological changes, such as necrosis and inflammatory cell
infiltration (Fig. 1A). When
treated with Sal B, pathological changes in the model group were
significantly improved in a concentration-dependent manner. In
chronic liver injury experiment, hepatocytes in the CCl4
model group were necrotic with liver fibrosis seen in the
intercellular and portal area of hepatocytes (Fig. 1B). In contrast, the degree of liver
fibrosis in the Sal B groups showed a dose-dependent reduction in
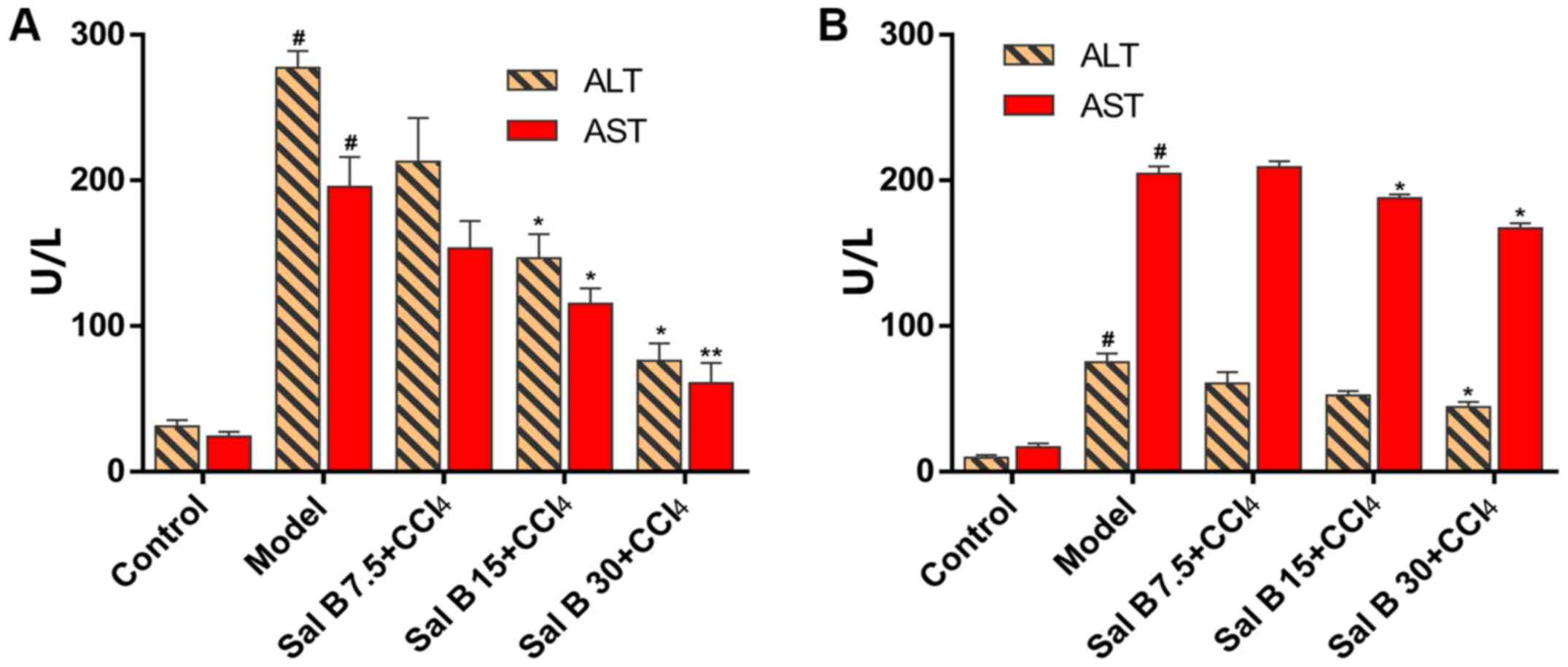
liver injury. The present study also measured ALT and AST levels in
mouse serum. In acute liver injury, CCl4 significantly
increased ALT and AST levels compared with the control group, while
Sal B significantly decreased these factors in
concentration-dependent manner compared with the model group
(Fig. 2A). In the chronic liver
injury model, Sal B significantly decreased ALT and AST levels
compared with the model group (Fig.
2B).
Effects of Sal B on antioxidant
enzymes in acute and chronic liver injury
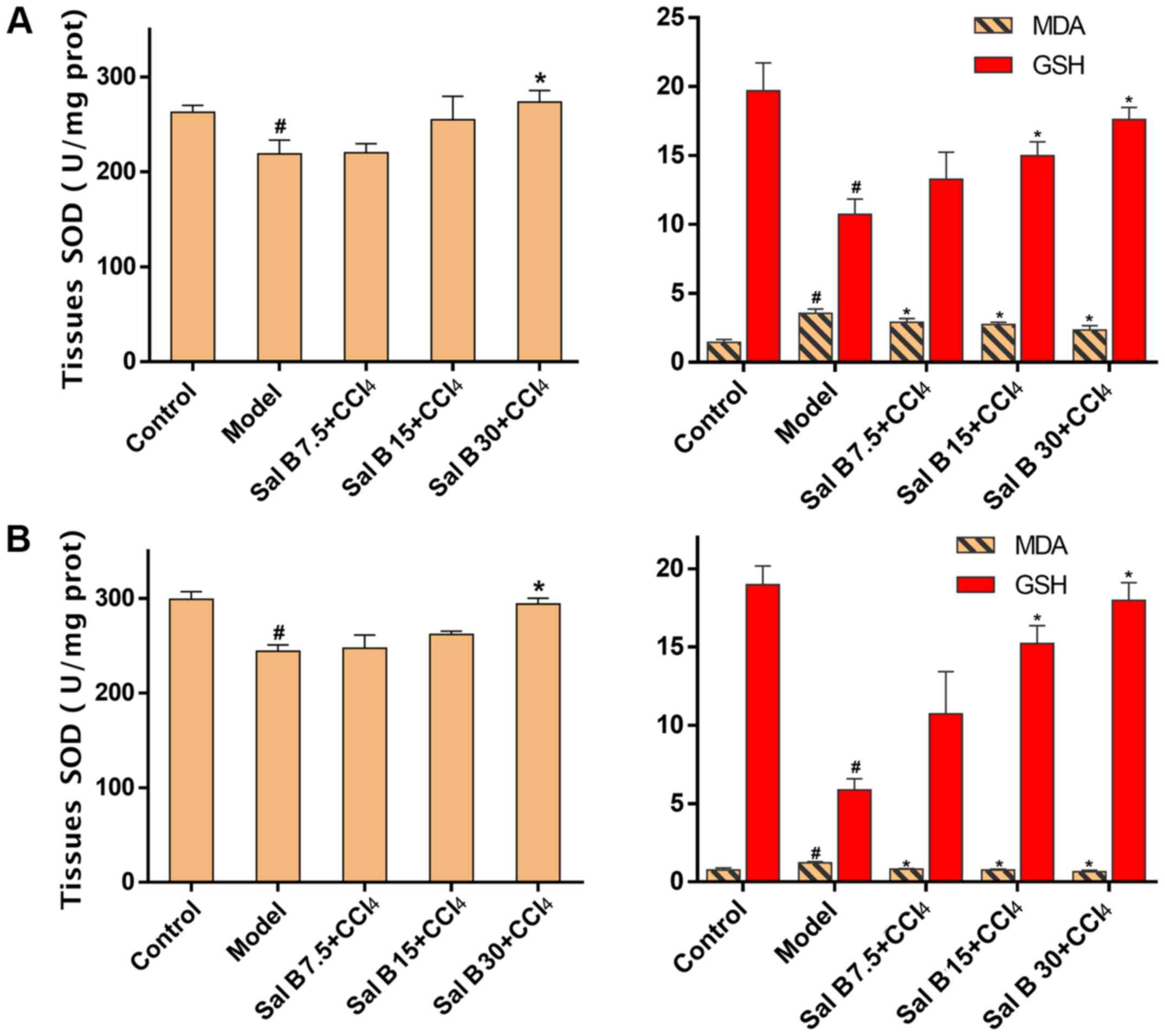
In the acute liver injury experiment, MDA, a
byproduct of lipid peroxidation, was significantly increased after
CCl4 treatment, while the addition of Sal B resulted in
a concentration-dependent decrease in MDA production (Fig. 3A). In contrast, antioxidants such as
SOD and GSH were increased in response to Sal B treatment,
particularly in the Sal B 30 mg/kg group. These results were
consistent with those found in the chronic liver injury model, as
indicated in Fig. 3B.
Effects of Sal B on the Nrf2/HO-1
signaling pathway
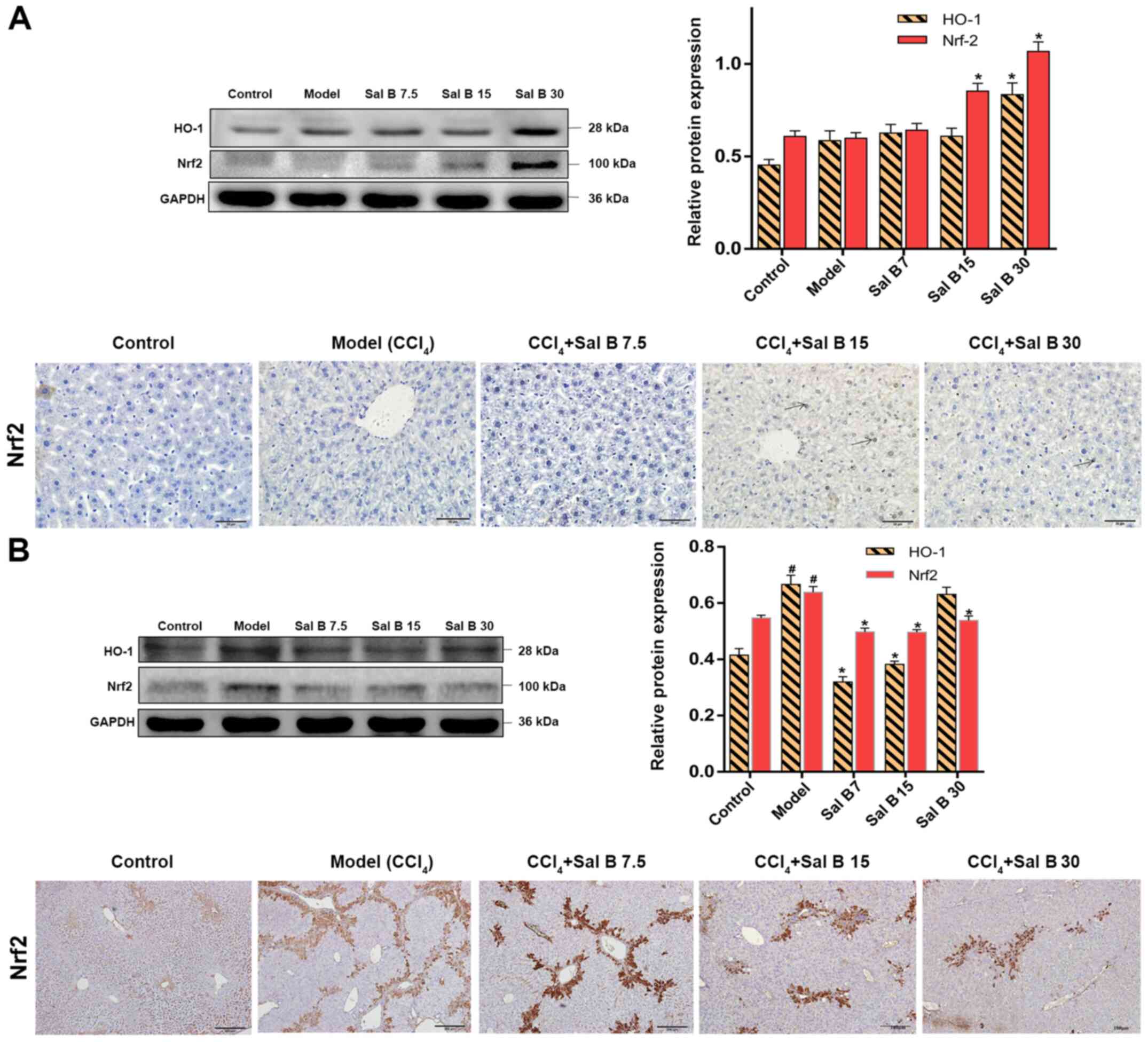
The kelch like ECH associated protein 1/Nrf2/HO-1
signaling pathway plays an important role in the process of
antioxidative damage and inflammatory injury of cells (22). The protein expression levels of Nrf2
and HO-1 were upregulated by CCl4 treatment, and were
further increased after the addition of 30 mg/kg Sal B in acute
liver injury mice (Fig. 4A).
However, in chronic liver injury mice Nrf2 and HO-1 were
downregulated compared with the model group (Fig. 4B). Moreover, immunohistochemistry
showed that activated Nrf2 was present around the central vein in
CCl4, Sal B 7.5, Sal B 15 and Sal B 15 groups. It was
found that liver fibrosis in the Sal B-treated groups was improved
and the area of Nrf2 expression was decreased (Fig. 4B). Additionally, the present results
suggested that Sal B regulates the Nrf2 signal more strongly in
acute liver injury compared with chronic liver injury.
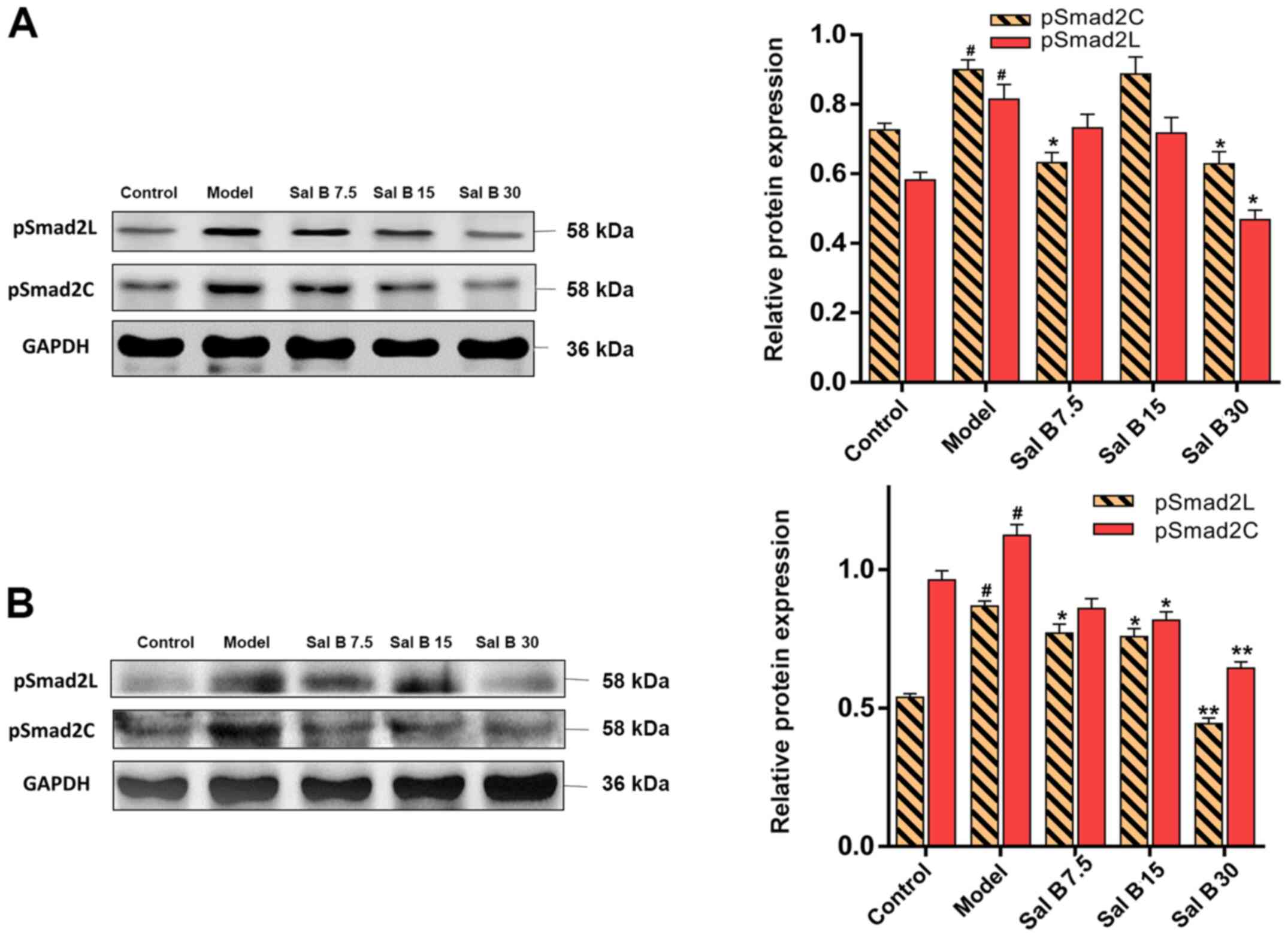
Effects of Sal B on pSmad2C/L in acute
and chronic liver injury mice
To assess whether Sal B could affect the
phosphorylation of Smad2C and Smad2L, acute and chronic liver
injury mice were treated with different doses of Sal B. It was
found that the protein expression levels of pSmad2C and pSmad2L
were decreased after Sal B treatment (Fig. 5A). Furthermore, this downregulation
in pSmad2C and pSmad2L was more distinctive in chronic liver injury
mice (Fig. 5B).
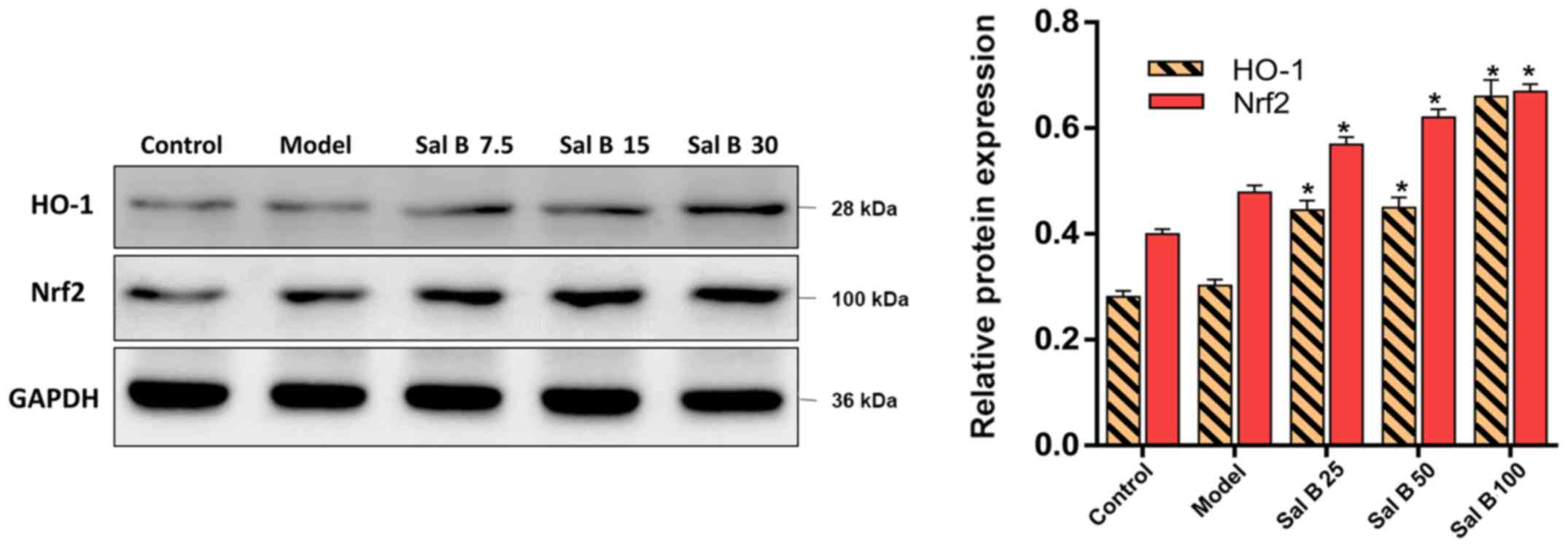
Effects of Sal B on the Nrf2/HO-1
signaling pathway in H2O2 induced HSC-T6
injury
To understand the mechanisms of the attenuation of
oxidative stress by Sal B, the present study hypothesized that the
antioxidant capability of Sal B may result from increased levels of
Nrf2 and HO-1. In comparison with the control group, the protein
expression levels of Nrf2 and HO-1 in the model group were slightly
increased, but this was not significant (Fig. 6). However, pretreatment of HSC-T6
cells stimulated by H2O2 with Sal B (25, 50
or 100 μmol/l) showed a concentration-dependent increase in
the protein expression level of Nrf2. Therefore, the in
vitro data showed similar patterns to the results from the
animal experiments, which suggested that the protective effect of
Sal B are via the activation of the Nrf2/HO-1 signaling pathway in
acute and chronic liver injury mice.
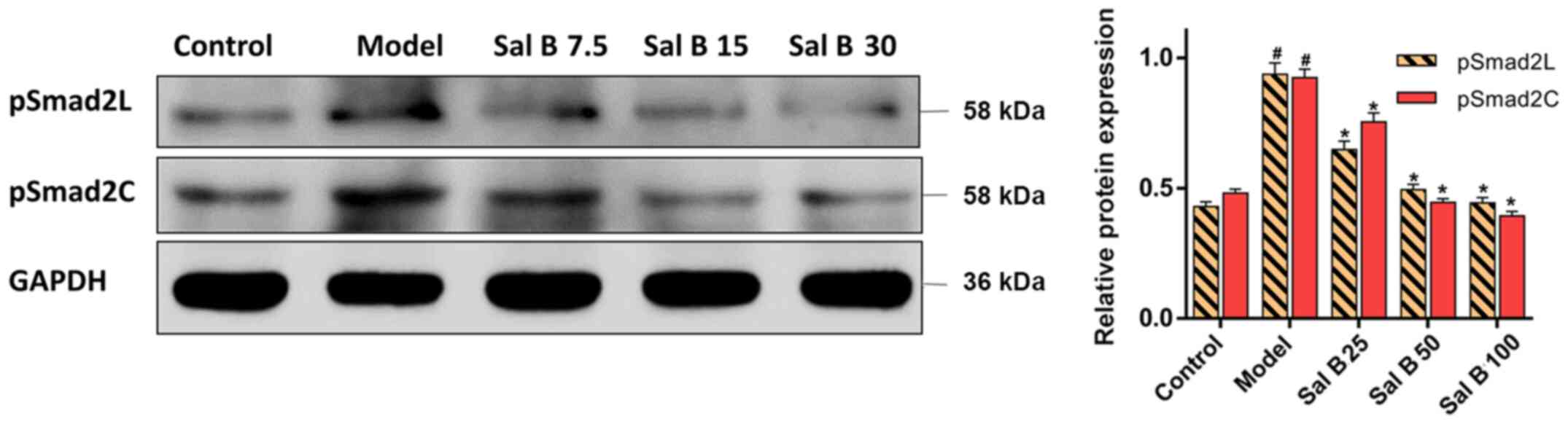
Effects of Sal B on pSmad2C/L in H2O2
induced HSC-T6 injury
It was demonstrated that H2O2
treatment significantly increased the protein expression levels of
pSmad2C and pSmad2L in the model group (Fig. 7). However, the protein expression
levels of pSmad2C and pSmad2L were significantly decreased by Sal B
(25, 50 and 100 μmol/l) treatment. Collectively, the present
results suggested that Sal B might have a protective effect on both
acute and chronic liver injury, which involved the downregulation
of pSmad2C and pSmad2L.
Discussion
Toxic reagents produce excessive electrophilic
products and free radicals, which are important factors of inducing
liver injury, resulting in lipid peroxidation, infiltration of
inflammatory cells and necrosis (23,24).
Thus, these contribute to the progression of persistent liver
injury to liver fibrosis (25).
Currently, clinical antihepatic fibrosis methods mainly include
antiviral therapy and discontinuation of drugs, which can cause
liver injury (26). Therefore,
understanding the underlying mechanisms of anti-acute and -chronic
liver injury drugs may facilitate the clinical treatment of liver
injury. An increasing number of studies have investigated the
anti-hepatic injury activity of natural herbs (27-29).
Clinical and experimental studies have demonstrated that some
herbal extracts have preventive and therapeutic values against
chronic liver diseases. Sal B is considered as one of the drugs
with most potential in this herbal medicine category (30,31).
Another previous study suggested that Sal B exerts an antihepatic
fibrosis-carcinoma effect via the mediation of pSmad3C/pSmad3L
(16). The present study
investigated whether Sal B could protect CCl4- and
H2O2-induced acute and chronic liver injury,
and the possible mechanism involved, with a focus on its effect on
pSmad2C/L in liver injury pathogenesis.
Serum AST and ALT levels are commonly used indexes
to assess the degree of hepatocyte damage. Elevation of serum AST
and ALT levels contributes to liver damage as normally these
factors are localized in the cytoplasm, and are released into the
serum after the increase of membrane permeability (32). In the present animal experiments,
Sal B reduced ALT and AST levels in acute and chronic liver injury
mice, thus suggesting that Sal B may alleviate
CCl4-induced hepatocyte damage in mice. With the
progression of liver injury, necrosis of hepatocytes occurs and the
AST/ALT value can reflect the degree of hepatocyte injury and
necrosis (33). Furthermore, the
mitochondria remain intact and ratio of AST/ALT <1.0 can be seen
in acute liver injury. With the persistence of injury factors,
mitochondria are damaged and AST is released from the cytoplasm of
mitochondria, as a result, the ratio of AST/ALT >1.0 usually
found in chronic liver injury (34). The histological observations in the
present study were consistent with these previous results, which
indicated the protective effect of Sal B on mice against the liver
injury induced by CCl4; 30 mg/kg Sal B had a strong
protective effect on acute and chronic liver injury. Hepatocellular
damage can be triggered by CCl4 and the CYP450 enzyme
system is predominantly responsible for the metabolism of
CCl4 to form free radicals (35). Highly reactive radicals generated
from CCl4, such as trichloromethyl and trichloromethyl
peroxyl, which destroy the respiratory chain on the mitochondrial
membrane (36), then accumulate ROS
in the cells to produce oxidative stress (37). SOD, GSH and MDA are representative
oxidative biochemical parameters (38). To determine oxidant stress, these
parameters were measured in liver tissues in the present study. It
was found that treatment of Sal B increased the production of GSH
and SOD, while it decreased of MDA in liver tissues, which
demonstrates the amelioratory effects of Sal B in oxidative stress
and damage in acute and chronic liver injury. Moreover, TGF-β1
increases intracellular ROS and inhibits antioxidant enzymes
degrading, which leads to redox imbalance; in return, ROS also
activates TGF-β1, mediating a profibrogenic effect; thus, a vicious
cycle forms (39). Sustained
activation of TGF-β/Smads signaling pathways may promote chronic
liver injury progression to liver fibrosis (40,41).
Upon activation by TGF-β1, Smad2 and Smad3 are
activated by C-terminal receptor-mediated phosphorylation and
oligomerized with Smad4 to form a complex, which preferentially
relocates into the nucleus (42).
Previous studies have shown that TGF-β1 and pro-inflammatory
cytokines synergistically promote collagen synthesis and accelerate
liver fibrosis by activating HSCs and promoting the profibrogenic
pSmad2C/L signaling pathway (8,43). To
investigate whether Sal B could affect profibrogenic pSmad2C/L
signaling, phosphorylated Smad2C and Smad2L were detected in liver
injury tissues. The present results suggested that Sal B inhibited
the phosphorylation of Smad2C/L in a dose-dependent manner in
CCl4 induced acute and chronic liver injury, especially
in chronic liver injury. Therefore, the present results indicated
that the anti-injury effect of Sal B inhibited phosphorylation of
Smad2C and Smad2L via the canonical TGF-β/Smad signaling
pathway.
The Nrf2/HO-1 signaling pathway is also of
importance as the activation of this pathway enhances the
expression of antioxidant protective genes, which has a positive
effect in oxidative stress (44).
Previous studies have identified a crosstalk between both pathways
of Nrf2 and TGF-β1 (45,46). TGF-β1 has been demonstrated to
suppress Nrf2-dependent antioxidant enzymes and TGF-β/Smad
signaling induces ATF3, which in turn complexes with Nrf2, and then
the ATF3/Nrf2 complex binds with ARE to suppress Nrf2 target gene
expression. Moreover, Nrf2 inhibits the TGF-β/Smad signaling
pathway by inhibiting TGF-β1 and blocking Smad3 activation
(10,12). The present study found that Sal B
significantly increased the total Nrf2 expression level in acute
liver injury (47,48), which was in line with a previous
study on high-fat diet induced inflammatory function disorder
(49). However, in the chronic
liver injury experiment, the Nrf2 expression levels in Sal
B-treatment groups were decreased. Moreover, it was demonstrated
that Nrf2 expression was mainly localized in the HSCs, which were
around the fibrotic areas. This results is consistent with results
from a previous study that showed that HSCs can be activated by
Nrf2 in liver tissues (50).
Therefore, the decrease in Nrf2 may due to reduction of fibrosis
area. Additionally, the present results suggested that Sal B
significantly decreased Nrf2 expression levels in
H2O2-induced cell injury model, which was in
line with a previous study (50),
and also found in the in vivo experiments where Nrf2
expression levels were not significantly different between the
control and model groups. Collectively, the present results
suggested that Sal B could upregulate the expression levels of Nrf2
and HO-1, and restore the oxidant/antioxidant balance. Thus, the
activation of the Nrf2/HO-1 pathway could inhibit the action of the
TGF-β/Smad signaling pathway.
In conclusion, the present results suggested that
Sal B protected the liver against acute and chronic injury by
inhibiting phosphorylation of Smad2 both at C-terminal and link
region via the TGF-β/Smad signaling pathway. Furthermore, it was
found that the Nrf2/HO-1 pathway might play an auxiliary role in
this process (Fig S1).
Supplementary Material
Sal B improves the oxidative stress
state and inhibits liver fibrosis. Under normal circumstances, the
generation and elimination of ROS are in a state of dynamic
equilibrium. When the body is stimulated by free radicals, the
dynamic balance of ROS production and clearance is destroyed,
resulting in liver damage. The accumulation of ROS in long-term
liver injury can activate the TGF-β/Smad signaling pathway to
promote liver fibrosis and TGF-β1 can promote the
production of ROS, thus forming a vicious cycle. In the present
study, Sal B inhibited liver fibrosis by attenuating the oxidative
stress state via the activation of the Nrf2/HO-1 signaling pathway
and inhibition of pSmad2L/C. ROS, reactive oxygen species; Sal B,
salvianolic acid B; p-, phosphorylated; Smad2C, Smad2 at
COOH-terminal; Smad2L, Smad2 at linker region; HO-1, heme
oxygenase-1; Nrf2, nuclear factor erythroid-2-related factor 2;
TGF-β1, transforming growth factor β 1; ALT, alanine
transaminase; AST, aspartate aminotransferase; SOD, superoxide
dismutase; MDA, malondialdehyde; GSH, glutathione.
Acknowledgements
Not applicable.
Funding
Funding: This study was supported by the National Natural
Science Foundation of China (grant nos. 81573652 and 81874354).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YY designed, initiated and directed this study. XMT,
DL, CZ, GHW, CW, YYX, YK, WPL and HYD performed the experiments. XT
and CW wrote the manuscript. YY revised the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All the experiments were conducted in accordance
with the guidelines for ethical review of laboratory animal welfare
in the Animal Center of Anhui Medical University. The animal
experiments were approved by the Experimental Animal Ethics
Committee of Anhui Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ramadori G, Moriconi F, Malik I and Dudas
J: Physiology and pathophysiology of liver inflammation, damage and
repair. J Physiol Pharmacol. 59:107–117. 2008.PubMed/NCBI
|
|
2
|
Hernandez-Gea V and Friedman SL:
Pathogenesis of liver fibrosis. Annu Rev Pathol. 6:425–456.
2011.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Enomoto M, Morikawa H, Tamori A and Kawada
N: Noninvasive assessment of liver fibrosis in patients with
chronic hepatitis B. World J Gastroenterol. 20:12031–12038.
2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Dewidar B, Meyer C, Dooley S and
Meindl-Beinker AN: TGF-β in hepatic stellate cell activation and
liver fibrogenesis-Updated 2019. Cells. 8(8)2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Xu F, Liu C, Zhou D and Zhang L:
TGF-β/SMAD pathway and its regulation in hepatic fibrosis. J
Histochem Cytochem. 64:157–167. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Yoshida K, Murata M, Yamaguchi T,
Matsuzaki K and Okazaki K: Reversible human TGF-β signal shifting
between tumor suppression and fibro-carcinogenesis: Implications of
Smad phospho-isoforms for hepatic epithelial-mesenchymal
transitions. J Clin Med. 5(5)2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Yoshida K, Murata M, Yamaguchi T and
Matsuzaki K: TGF-β/Smad signaling during hepatic
fibro-carcinogenesis (review). Int J Oncol. 45:1363–1371.
2014.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Matsuzaki K: Smad phosphoisoform signals
in acute and chronic liver injury: Similarities and differences
between epithelial and mesenchymal cells. Cell Tissue Res.
347:225–243. 2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Arfmann-Knübel S, Struck B, Genrich G,
Helm O, Sipos B, Sebens S and Schäfer H: The crosstalk between Nrf2
and TGF-β1 in the epithelial-mesenchymal transition of pancreatic
duct epithelial cells. PLoS One. 10(e0132978)2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Choi HK, Pokharel YR, Lim SC, Han HK, Ryu
CS, Kim SK, Kwak MK and Kang KW: Inhibition of liver fibrosis by
solubilized coenzyme Q10: Role of Nrf2 activation in inhibiting
transforming growth factor-beta1 expression. Toxicol Appl
Pharmacol. 240:377–384. 2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Okita Y, Kamoshida A, Suzuki H, Itoh K,
Motohashi H, Igarashi K, Yamamoto M, Ogami T, Koinuma D and Kato M:
Transforming growth factor-β induces transcription factors MafK and
Bach1 to suppress expression of the heme oxygenase-1 gene. J Biol
Chem. 288:20658–20667. 2013.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Michaeloudes C, Chang PJ, Petrou M and
Chung KF: Transforming growth factor-β and nuclear factor
E2–related factor 2 regulate antioxidant responses in airway smooth
muscle cells: Role in asthma. Am J Respir Crit Care Med.
184:894–903. 2011.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Cao W, Guo XW, Zheng HZ, Li DP, Jia GB and
Wang J: Current progress of research on pharmacologic actions of
salvianolic acid B. Chin J Integr Med. 18:316–320. 2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lin YL, Wu CH, Luo MH, Huang YJ, Wang CN,
Shiao MS and Huang YT: In vitro protective effects of salvianolic
acid B on primary hepatocytes and hepatic stellate cells. J
Ethnopharmacol. 105:215–222. 2006.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Tao YY, Wang QL, Shen L, Fu WW and Liu CH:
Salvianolic acid B inhibits hepatic stellate cell activation
through transforming growth factor beta-1 signal transduction
pathway in vivo and in vitro. Exp Biol Med (Maywood).
238:1284–1296. 2013.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ying M, Meng F, Chao W, et al: Salvianolic
acid B exerts anti-hepatic fibrosis-carcinoma effect via mediation
of pSmad3C/pSmad3L. Zhongguo Yaolixue Tongbao. 1:44–50. 2018.
|
|
17
|
Jiang W, Gao M, Sun S, Bi A, Xin Y, Han X,
Wang L, Yin Z and Luo L: Protective effect of L-theanine on carbon
tetrachloride-induced acute liver injury in mice. Biochem Biophys
Res Commun. 422:344–350. 2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Chu X, Wang H, Jiang YM, Zhang YY, Bao YF,
Zhang X, Zhang JP, Guo H, Yang F, Luan YC, et al: Ameliorative
effects of tannic acid on carbon tetrachloride-induced liver
fibrosis in vivo and in vitro. J Pharmacol Sci. 130:15–23.
2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
El-Wakeel SA, Rahmo RM and El-Abhar HS:
Anti-fibrotic impact of Carvedilol in a CCl-4 model of liver
fibrosis via serum microRNA-200a/SMAD7 enhancement to bridle
TGF-β1/EMT track. Sci Rep. 8(14327)2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Al-Saeedi FJ: Study of the cytotoxicity of
asiaticoside on rats and tumour cells. BMC Cancer.
14(220)2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Hu X, Liang Y, Zhao B and Wang Y:
Oxyresveratrol protects human lens epithelial cells against
hydrogen peroxide-induced oxidative stress and apoptosis by
activation of Akt/HO-1 pathway. J Pharmacol Sci. 139:166–173.
2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Loboda A, Damulewicz M, Pyza E, Jozkowicz
A and Dulak J: Role of Nrf2/HO-1 system in development, oxidative
stress response and diseases: An evolutionarily conserved
mechanism. Cell Mol Life Sci. 73:3221–3247. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Morita M, Ishida N, Uchiyama K, Yamaguchi
K, Itoh Y, Shichiri M, Yoshida Y, Hagihara Y, Naito Y, Yoshikawa T,
et al: Fatty liver induced by free radicals and lipid peroxidation.
Free Radic Res. 46:758–765. 2012.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Patel SJ, Milwid JM, King KR, Bohr S,
Iracheta-Vellve A, Li M, Vitalo A, Parekkadan B, Jindal R and
Yarmush ML: Gap junction inhibition prevents drug-induced liver
toxicity and fulminant hepatic failure. Nat Biotechnol. 30:179–183.
2012.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Tsukamoto H, French SW, Benson N, Delgado
G, Rao GA, Larkin EC and Largman C: Severe and progressive
steatosis and focal necrosis in rat liver induced by continuous
intragastric infusion of ethanol and low fat diet. Hepatology.
5:224–232. 1985.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Schuppan D: Liver fibrosis: Common
mechanisms and antifibrotic therapies. Clin Res Hepatol
Gastroenterol. 39:51–59. 2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Chen SR, Chen XP, Lu JJ, Wang Y and Wang
YT: Potent natural products and herbal medicines for treating liver
fibrosis. Chin Med. 10(7)2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Stickel F and Schuppan D: Herbal medicine
in the treatment of liver diseases. Dig Liver Dis. 39:293–304.
2007.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ma X, Peng JH and Hu YY: Chinese Herbal
Medicine-induced liver injury. J Clin Transl Hepatol. 2:170–175.
2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Rui W, Xie L, Liu X, He S, Wu C, Zhang X,
Zhang L and Yang Y: Compound Astragalus and Salvia
miltiorrhiza extract suppresses hepatocellular carcinoma
progression by inhibiting fibrosis and PAI-1 mRNA transcription. J
Ethnopharmacol. 151:198–209. 2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Liu T, Xia Y, Li J, Li S, Feng J, Wu L,
Zhang R, Xu S, Cheng K, Zhou Y, et al: Shikonin attenuates
Concanavalin A-induced acute liver injury in mice via inhibition of
the JNK pathway. Mediators Inflamm. 2016(2748367)2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Clark JM, Brancati FL and Diehl AM: The
prevalence and etiology of elevated aminotransferase levels in the
United States. Am J Gastroenterol. 98:960–967. 2003.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Anderson FH, Zeng L, Rock NR and Yoshida
EM: An assessment of the clinical utility of serum ALT and AST in
chronic hepatitis C. Hepatol Res. 18:63–71. 2000.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Giannini E, Botta F, Fasoli A, Ceppa P,
Risso D, Lantieri PB, Celle G and Testa R: Progressive liver
functional impairment is associated with an increase in AST/ALT
ratio. Dig Dis Sci. 44:1249–1253. 1999.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Bhadauria M, Nirala SK, Shrivastava S,
Sharma A, Johri S, Chandan BK, Singh B, Saxena AK and Shukla S:
Emodin reverses CCl induced hepatic cytochrome P450 (CYP) enzymatic
and ultrastructural changes: The in vivo evidence. Hepatol Res.
39:290–300. 2009.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Comporti M: Three models of free
radical-induced cell injury. Chem Biol Interact. 72:1–56.
1989.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Sies H: Oxidative stress: A concept in
redox biology and medicine. Redox Biol. 4:180–183. 2015.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Kalpravidh RW, Siritanaratkul N, Insain P,
Charoensakdi R, Panichkul N, Hatairaktham S, Srichairatanakool S,
Phisalaphong C, Rachmilewitz E and Fucharoen S: Improvement in
oxidative stress and antioxidant parameters in beta-thalassemia/Hb
E patients treated with curcuminoids. Clin Biochem. 43:424–429.
2010.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Liu RM and Desai LP: Reciprocal regulation
of TGF-β and reactive oxygen species: A perverse cycle for
fibrosis. Redox Biol. 6:565–577. 2015.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Derynck R and Zhang YE: Smad-dependent and
Smad-inde pendent pathways in TGF-beta family signalling. Nature.
425:577–584. 2003.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Inagaki Y and Okazaki I: Emerging insights
into transforming growth factor beta Smad signal in hepatic
fibrogenesis. Gut. 56:284–292. 2007.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Hu HH, Chen DQ, Wang YN, Feng YL, Cao G,
Vaziri ND and Zhao YY: New insights into TGF-β/Smad signaling in
tissue fibrosis. Chem Biol Interact. 292:76–83. 2018.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Burch ML, Yang SNY, Ballinger ML, Getachew
R, Osman N and Little PJ: TGF-beta stimulates biglycan synthesis
via p38 and ERK phosphorylation of the linker region of Smad2. Cell
Mol Life Sci. 67:2077–2090. 2010.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Zhang H, Liu YY, Jiang Q, Li KR, Zhao YX,
Cao C and Yao J: Salvianolic acid A protects RPE cells against
oxidative stress through activation of Nrf2/HO-1 signaling. Free
Radic Biol Med. 69:219–228. 2014.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Oh CJ, Kim JY, Min AK, Park KG, Harris RA,
Kim HJ and Lee IK: Sulforaphane attenuates hepatic fibrosis via
NF-E2-related factor 2-mediated inhibition of transforming growth
factor-β/Smad signaling. Free Radic Biol Med. 52:671–682.
2012.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Chen Q, Zhang H, Cao Y, Li Y, Sun S, Zhang
J and Zhang G: Schisandrin B attenuates CCl4-induced
liver fibrosis in rats by regulation of Nrf2-ARE and TGF-β/Smad
signaling pathways. Drug Des Devel Ther. 11:2179–2191.
2017.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Zhang X, Wu Q, Lu Y, Wan J, Dai H, Zhou X,
Lv S, Chen X, Zhang X, Hang C, et al: Cerebroprotection by
salvianolic acid B after experimental subarachnoid hemorrhage
occurs via Nrf2- and SIRT1-dependent pathways. Free Radic Biol Med.
124:504–516. 2018.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Liu M, Xu H, Zhang L, Zhang C, Yang L, Ma
E, Liu L and Li Y: Salvianolic acid B inhibits myofibroblast
transdifferentiation in experimental pulmonary fibrosis via the
up-regulation of Nrf2. Biochem Biophys Res Commun. 495:325–331.
2018.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Wang B, Sun J, Shi Y and Le G: Salvianolic
acid B inhibits high-fat diet-induced inflammation by activating
the Nrf2 pathway. J Food Sci. 82:1953–1960. 2017.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Lu C, Xu W and Zheng S: Nrf2 activation is
required for curcumin to induce lipocyte phenotype in hepatic
stellate cells. Biomed Pharmacother. 95:1–10. 2017.PubMed/NCBI View Article : Google Scholar
|