Introduction
Ovarian cancer (OC) is one of the most common
malignant tumors in gynecology (1).
Compared with endometrial cancer, cervical cancer and vulvar
cancer, the mortality rate of OC ranks first among the female
genital tract malignant tumors, and its incidence is increasing
year by year, which seriously threatens the life and health of
women worldwide (2). For instance,
the American Cancer Society estimated ~22,530 new OC cases and
13,980 mortalities in the United States in 2019(3). Anatomically, as OC is located in the
deep structure of the pelvic cavity, there are no obvious signs of
early pathological changes, which are difficult to detected
(1). In addition, there is no
specific diagnostic method, and thus ~59% of patients with
abdominal distension, dropsy or pain are in advanced stage at
diagnosis. Therefore, insidious onset, late onset, high degree of
malignancy, easy metastasis and poor prognosis are the main
clinical features of OC (1). It has
been shown that the 5-year survival rate of patients with early OC
is >75%, while that of patients with advanced OC is only 29%
(4). Cytoreductive surgery and
cisplatin-based chemotherapy are standardized therapies for
patients with advanced OC (5), and
with the improvement of diagnosis and treatment, the 5-year
survival rate of patients with OC increased from 36% in 1975 to 47%
in 2014 (6,7). However, >50% of patients with
advanced OC relapse within the first 5 years after treatment,
resulting in cross-resistance to standardized therapy and other
functional and structural unrelated chemotherapeutic drugs
(8). Thus, the development of novel
treatment strategies is urgently required to improve the treatment
of patients with OC.
Differential embryo-chondrocyte expressed gene 1
(DEC1) is a helix-loop-helix transcription factor that directly
binds to the class B type E-box in target genes, and is expressed
to different degrees in most organs and tissues of the human body
(9-11).
DEC1 serves an essential role in various physiological functions of
the body, including cell differentiation, cell cycle and circadian
rhythm regulation, hypoxia, stress response and other processes
(12,13). It has been reported that DEC1 is
abnormally expressed in several tumors and is related to the growth
and apoptosis of tumors. For example, DEC1 has a positive
anti-apoptotic effect on the proliferation of gastric cancer cells
induced with hypoxia (14).
Moreover, DEC1 negatively affects the expression of E-cadherin in
HepG2 cells induced with hypoxia (15), while silencing DEC1 inhibits the
proliferation of breast cancer by inducing cell cycle arrest in S
phase (16). However, the possible
effects and underlying mechanisms of DEC1 on OC are yet to be
elucidated.
It has been revealed that DEC1 overexpression and
knockdown inversely regulate the expression levels of β-catenin and
phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit α
in glucocorticoid-induced osteoporotic changes in SaoS-2 cells and
mice (17). Furthermore,
downregulation of DEC1 contributes to
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced neurotoxicity
by suppressing the PI3K/Akt/glycogen synthase kinase 3 β (GSK-3β)
pathway (18). The Wnt/β-catenin
signaling pathway is one of the most important signal transduction
pathways in cells, and serves an essential role in regulating
normal cell proliferation, movement, differentiation and
tumorigenesis (19,20). β-catenin, as a key molecule of the
Wnt/β-catenin pathway, accumulates in the cytoplasm and then
translocates to the nucleus to activate downstream signal molecules
in the nucleus (21,22). Previous studies have shown that
abnormal activation of the Wnt/β-catenin pathway is closely
corrected with the occurrence and development of numerous tumors.
For instance, Wang et al (23) reported that after downregulation of
odd-skipped related transcription factor 1 gene expression in H1299
cells, the Wnt/β-catenin signaling pathway was inhibited and GSK-3β
expression was significantly increased, while phosphorylated
(p)-GSK-3β, nuclear-β-catenin, cyclin D1, c-Myc and matrix
metalloproteinase (MMP)-7 expression levels were significantly
decreased, resulting in the reduction of proliferative and invasive
abilities of H1299 cells (23).
Furthermore, the Wnt pathway is activated in OC cells to promote
β-catenin nuclear translocation, which subsequently acts on T-cell
specific transcription factor (TCF) and lymphoid enhancer binding
factor (LEF) transcription factors to induce the proliferation,
differentiation and metastasis of OC stem cells (24). It has also been shown that in
healthy colorectal tissues, the Wnt signaling pathway regulates the
balance, proliferation and differentiation of intestinal stem cells
(25).
The aim of the present study was to investigate the
possible role and underlying mechanisms of DEC1 in OC. It was
demonstrated that inhibition of DEC1 significantly inhibited
proliferation, migration and invasion and induced apoptosis, which
may be regulated by the Wnt/β-catenin pathway. The results of the
present study provide novel insights into the biological functions
and underlying mechanisms of DEC1 in OC, and may facilitate the
development of a novel therapeutic target for diagnosing or
treating OC.
Materials and methods
Tissue samples
The present study involved 106 patients (aged 30-75
years; median age, 60 years) who were diagnosed with OC in the
Gynecology Department of Jiangxi Cancer Hospital Affiliated to
Nanchang University (Nanchang, China) between September 2015 and
September 2018. No chemoradiotherapy was performed before the
operation. OC tissues and corresponding adjacent healthy tissue
were resected from each patient and immediately placed in a liquid
nitrogen flash freezer at -80˚C. None of the patients had received
any chemotherapy or radiotherapy. All tumors were histologically
classified as well (grade 1), moderately (grade 2), poorly (grade
3) and others (grade 4) according to the Edmondson-Steiner grading
system (26). This study has
obtained approval from the Jiangxi Cancer Hospital Medical Ethics
Committee and oral informed consent was obtained from all
patients.
Cell lines
In total, two OC cell lines (SKOV3 and OVCAR3) and a
normal human ovarian epithelial cell line (IOSE80) were purchased
from Stem Cell Bank, Chinese Academy of Sciences. RPMI-1640 (Gibco;
Thermo Fisher Scientific, Inc.) medium containing 1%
penicillin/streptomycin (Sigma-Aldrich; Merck KGaA) and 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) was used for cell culture
in a 5% CO2 incubator maintained at 37˚C.
Cell transfection
Short hairpin (sh)RNA targeting DEC1 (shEDC1) and
its negative control (NC shRNA) were synthesized by Shanghai
GenePharma Co., Ltd., and subsequently inserted into pSuper plasmid
(Shanghai GenePharma Co., Ltd.) using DNA recombination technology.
shEDC1 or NC (100 ng) was transfected in SKOV3 and OVCAR3 cells
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Transfected cells were cultured for 48 h, then the subsequent
functional experiments were performed.
Cell Counting Kit (CCK)-8 assay
Viability of transfected OC cells was evaluated
using CCK-8 assay (Beyotime Institute of Biotechnology), according
to the manufacturer's instructions. SKOV3 and OVCAR3 cells
(1x104/well) were seeded in 96-well plates and incubated
for 0, 24, 48 and 72 h at 37˚C. Then, cells were incubated with 10
µl CCK-8 solution for 4 h at 37˚C. The optical density (OD) was
measured using a microplate at 490 nm (Multiskan MK3; Thermo Fisher
Scientific, Inc.). Each experiment was performed in triplicate.
EdU assay
The proliferation of OC cells after transfection was
determined using a EdU assay. Transfected SKOV3 and OVCAR3 cells
(1x104/well) were seeded in 96-well plates and cultured
for 48 h. Then, cells were fixed with 4% paraformaldehyde at room
temperature for 30 min and 0.5% Triton X-100 was used to
permeabilize the nuclear membrane at room temperature for 10 min.
Cells were blocked with goat serum (Gibco; Thermo Fisher
Scientific, Inc.) at room temperature for 1 h, and then The nuclei
were counterstained for 15 min at room temperature with 100 ng/ml
DAPI (Thermo Fisher Scientific, Inc.). EdU-positive cells were
observed using a fluorescence microscope (magnification, x200;
Olympus Corporation). Each experiment was performed in
triplicate.
TUNEL assay
The OC tissue slices (4 µm) were washed with PBS and
then fixed at room temperature for 30 min with 4% paraformaldehyde.
After washing once with PBS, the slices were incubated with 0.1%
Triton X-100 at room temperature for 2 min and then washed once
with PBS. Subsequently, 3% H2O2 was used for
incubation (5 min) at room temperature. Slides was rinsed and
incubated with 50 µl TUNEL at 37˚C overnight. After rinsing with
PBS, the TUNEL reaction was visualized using chromogenic staining
with 3,3'-diaminobenzidine (DAB; Sigma-Aldrich; Merck KGaA). The
nuclei were counterstained for 15 min at room temperature with 100
ng/ml DAPI (Thermo Fisher Scientific, Inc.). Neutral gum was used
to seal the slides. A fluorescent microscope (magnification x100;
Leica Microsystems, Inc.) was used to observe the slides and the
TUNEL-positive SKOV3 and OVCAR3 cells were calculated in ImageJ
software (version 1.48; National Institutes of Health). Three
visual fields were observed. All measurements were performed three
times to ensure accuracy. Each experiment was performed in
triplicate.
Wound healing assay
Transfected SKOV3 and OVCAR3 cells were treated as
aforementioned before monolayer inoculation into a 6-well culture
plate at the density of 5x104 cells/well. The
supernatant fluid was removed when SKOV3 and OVCAR3 cells were
highly confluent (>90%). Scratches were made using a sterile
pipette tip, with the scratch width remaining the same. Then, the
culture solution was replaced with FBS-free RPMI-1640 solution.
After continuous culture for 48 h, Images were obtained using an
optical microscope (magnification, x100; BX-51; Olympus
Corporation). The scratch width of SKOV3 and OVCAR3 cells was
measured to calculate the wound healing rate in ImageJ software
(version 1.48; National Institutes of Health). Each experiment was
performed in triplicate.
Transwell invasion assay
Matrigel stored at -20˚C was thawed overnight at
2-8˚C. Transfected SKOV3 and OVCAR3 cells (1x103
cells/well) were transferred to 24-well Transwell inserts (5 µm;
Corning, Inc.) for monolayer inoculation or inoculated into the
upper chamber loaded with Matrigel using serum-free medium.
RPMI-1640 solution containing 10% FBS (v/v) (Gibco; Thermo Fisher
Scientific, Inc.) was added in the lower chamber, and the cells
were cultured at 37˚C for 48 h. Then, the Matrigel and the cells
that had not invaded the lower chamber were removed using test
paper. Cells passing through the Transwell membrane were counted
after fixing at room temperature for 30 min with 4%
paraformaldehyde, and then stained with 1% crystal violet at room
temperature for 10 min. Images were obtained using an light
microscope (magnification, x100; BX-51; Olympus Corporation). The
experiment was replicated three times independently under the same
conditions.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR) analysis
Total RNA was extracted from OC tissues and cell
lines using TRIzol® reagent (Thermo Fisher Scientific,
Inc.), and then reverse-transcribed into cDNA at 55˚C for 10 min
using PrimeScript RT-PCR kit (Takara Biotechnology Co., Ltd.). The
mRNA levels were quantified by RT-qPCR with the StepOne™
Real-Time PCR System (Thermo Fisher Scientific, Inc.) using
SYBR® Premix Ex Taq™ (Takara Biotechnology
Co., Ltd.). The reaction conditions were as follows: 94˚C 3 min; 30
cycles of 94˚C for 45 sec, 57˚C for 45 sec, 72˚C for 45 sec; and
final extension at 72˚C for 10 min. Primer sequences used were
designed as follows: DEC1 forward, 5'-TTGCTTTCCTTCCTCG-3' and
reverse, 5'-CACACACACCCTGCCTCTGC-3'; and GAPDH forward,
5'-GAAGGTGAAGGTCGGAGTCA-3' and reverse, 5'-AGGGGCCATCCACAGTCTTC-3'.
All target gene transcripts were normalized to GAPDH using the
2-ΔΔCq method (27).
Each experiment was performed in triplicate.
Western blotting
Proteins from transfected OC tissues and cell lines
were lysed with RIPA lysis buffer (Bio-Rad Laboratories, Inc.), and
total protein concentrations were determined using a bicinchoninic
acid protein assay kit (Beyotime Institute of Biotechnology).
Equivalent samples (30 µg) were separated using 10% SDS-PAGE and
then transferred onto a PVDF membrane for 2 h. Membranes were
blocked with 5% non-fat skim milk in TBS-0.1% Tween-20 buffer at
25˚C for 1 h and incubated with the primary antibodies at 4˚C
overnight: DEC1 (cat. no. ab97525; 1:1,000), caspase-3 (cat. no.
ab197202; 1:1,000), cleaved caspase-3 (cat. no. ab2302; 1:1,000),
poly(ADP-ribose) polymerase 1 (PARP; cat. no. ab74290; 1:1,000),
cleaved PARP (cat. no. ab32064; 1:1,000), E-cadherin (cat. no.
ab1416; 1:1,000), vimentin (cat. no. ab92547; 1:1,000), α-smooth
muscle actin (α-SMA; cat. no. ab32575; 1:1,000), glycogen synthase
kinase 3 β (GSK-3β; cat. no. ab93926; 1:1,000), p-GSK-3β (cat. no.
ab75814; 1:1,000), β-catenin (cat. no. ab32572; 1:1,000) and GAPDH
(cat. no. ab181602; 1:1,000). Then, the HRP-conjugated rabbit
anti-mouse IgG H&L antibodies (1:2,000; cat. no. ab6728) or
HRP-conjugated goat anti-rabbit IgG H&L antibodies (1:2,000;
cat. no. ab6721) was used at room temperature for 2 h. All
antibodies were purchased from Abcam. The relative band density was
determined using the Bio-Rad Imaging System (Bio-Rad Laboratories,
Inc.) with an ECL western blotting substrate kit (Tanon Science and
Technology Co., Ltd.). GAPDH was used as internal reference.
Protein bands were visualized using an ECL kit (Beyotime Institute
of Biotechnology) and quantified using ImageJ software (version
4.3; National Institutes of Health). Each experiment was performed
in triplicate.
Immunohistochemical analysis
OC tissues and corresponding adjacent healthy
tissues samples were fixed at room temperature for 30 min in 10%
neutral buffered formalin and embedded in paraffin wax. Slides with
a thickness of 4 µm were deparaffinized with 100% xylene,
rehydrated with anhydrous, 95, 75% ethanol and incubated with 30%
H2O2 at 37˚C for 10 min. Antigen repair was
performed by using 10 mM citric acid at 95˚C for 15 min. After
cooling, the sections were blocked at room temperature for 30 min
using 10% FBS. Then, slides were incubated at 4˚C overnight with
DEC1 antibody (1:1,000; cat. no. ab97525; Abcam). Subsequently, the
sections were incubated with biotin-conjugated goat anti-rabbit IgG
secondary antibody (1:1,000; cat. no. ab205718; Abcam) at 37˚C for
30 min. DAB was used as chromogenic agent. After washing with PBS,
the sections were counterstained with hematoxylin, dehydrated with
ethanol (70, 80, 90, 95 and 100%) and xylene, and then sealed with
neutral resin. Images were obtained using a fluorescence microscope
(magnification, x100; FSX100; Olympus Corporation). In total, five
representative visual fields were randomly selected to calculate
the percentage of positive cells. The results of DEC1 protein
positive staining were indicated by cells with brown granules. The
adjacent healthy tissues were used as the control.
Statistical analysis
SPSS 20.0 (IBM Corp.) was used for statistical
analysis. The survival curve was constructed using the Kaplan-Meier
method and compared using the log-rank test. Data are presented as
the mean ± standard deviation. Categorical data were analyzed using
a χ2 test. Intergroup comparison was examined using a
paired t-test (two groups) or one-way ANOVA (≥3 groups), followed
by Bonferroni test. P<0.05 was considered to indicate a
statistically significant difference.
Results
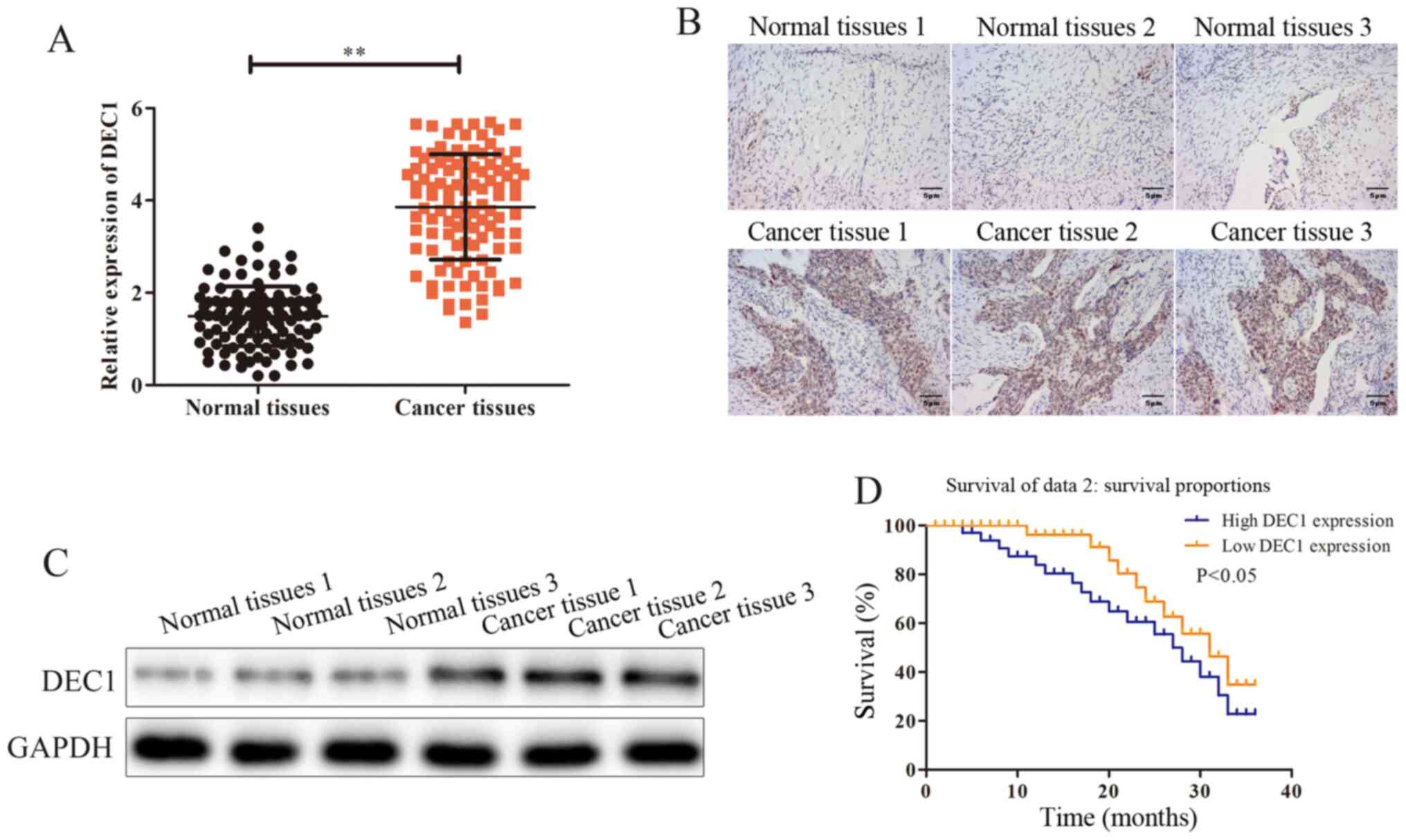
DEC1 is upregulated in OC tissues
To investigate the possible effects and mechanisms
of DEC1 on the occurrence of OC, RT-qPCR was performed to assess
DEC1 mRNA expression in OC tissues (n=106) and adjacent healthy
tissues (n=106). The results demonstrated that DEC1 was
significantly upregulated in OC tissues compared with adjacent
healthy tissues (Fig. 1A).
Subsequently, the association between DEC1 level and the
progression of OC was assessed. The general features of 106
patients with OC were presented and the association between the
expression of DEC1 and those features was analyzed. It was found
that the expression of DEC1 was positively associated with tumor
size and metastasis (Table I).
 | Table IAssociation between DEC1 expression
and clinicopathologic of patients with ovarian cancer. |
Table I
Association between DEC1 expression
and clinicopathologic of patients with ovarian cancer.
| | DEC1
expression | |
|---|
|
Characteristics | Case number | High (n=50) | Low (n=50) | P-value |
|---|
| Age, years | | | | 0.594 |
|
≤60 | 41 | 23 | 1 | |
|
>60 | 59 | 27 | 32 | |
| Tumor size, cm | | | | 0.026a |
|
≤5 | 35 | 17 | 36 | |
|
>5 | 65 | 33 | 14 | |
| Distant
metastasis | | | | 0.037a |
|
Yes | 54 | 38 | 18 | |
|
No | 46 | 12 | 32 | |
| Differentiation
grade | | | | 0.381 |
|
I + II | 69 | 26 | 31 | |
|
III +
IV | 31 | 24 | 19 | |
| Histological
grade | | | | 0.219 |
|
Well and
moderately | 56 | 29 | 27 |
|
Poorly and
others | 44 | 21 | 23 | |
| Clinical stage | | | | 0.321 |
|
I + II | 49 | 16 | 20 | |
|
III +
IV | 51 | 34 | 30 | |
Immunohistochemical and western blotting assays were
conducted to evaluate the protein expression of DEC1 in OC tissues
(n=3) and adjacent healthy tissues (n=3), and it was identified
that the expression of DEC1 was markedly higher in OC tissues
compared with adjacent healthy tissues (Fig. 1B and C). Furthermore, the Kaplan-Meier survival
analysis indicated that patients with high DEC1 expression had a
significantly poorer prognosis compared with those with a low
expression (Fig 1D). Thus, these
data suggested that DEC1 was upregulated in OC and that high
expression of DEC1 predicted poor prognosis of patients with
OC.
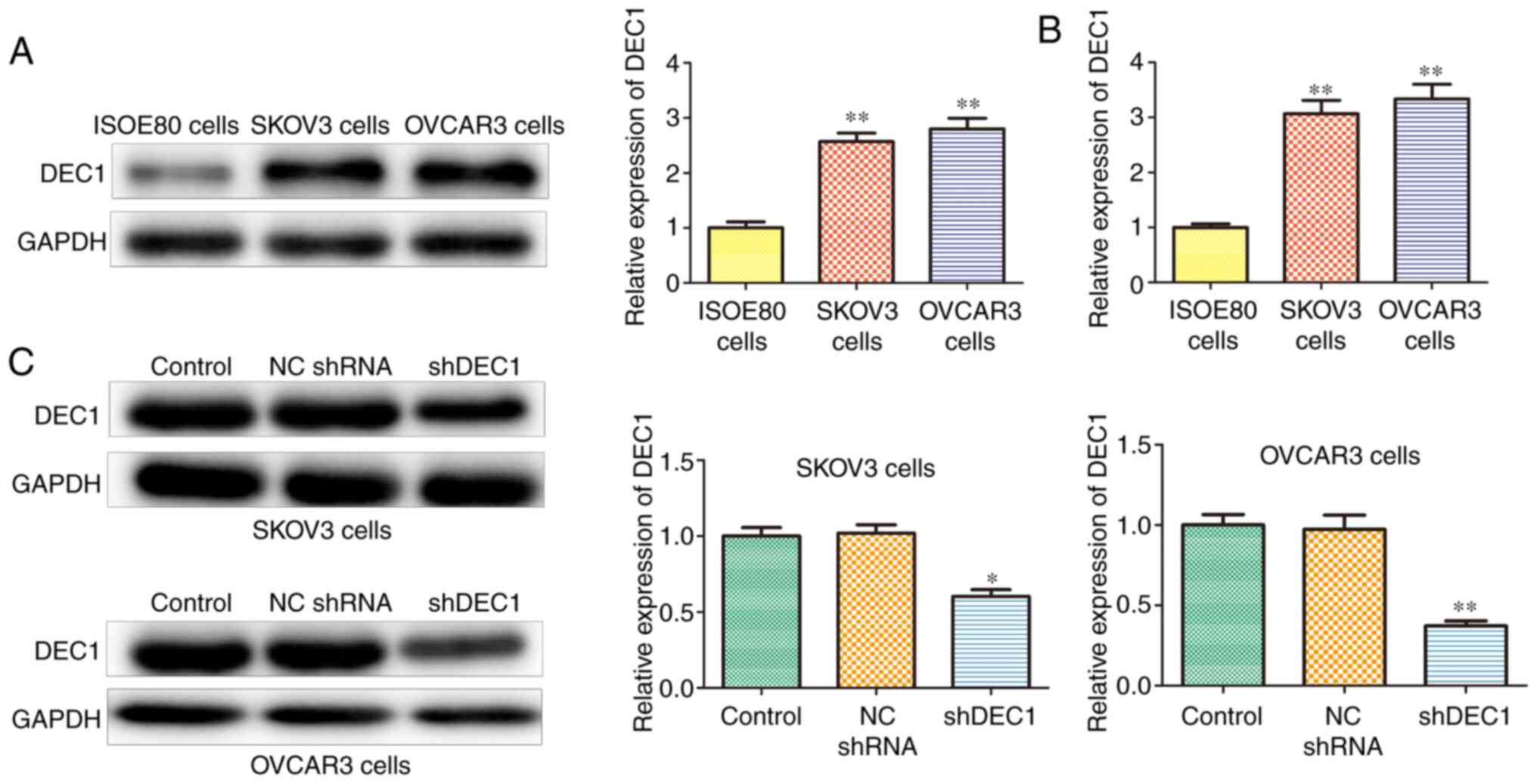
DEC1 is upregulated in OC cell
lines
Next, DEC1 expression was assessed in OC cell lines,
including SKOV3 and OVCAR3 using western blotting and RT-qPCR
assays. The protein and mRNA expression levels of DEC1 were
upregulated in SKOV3 and OVCAR3 cells compared with IOSE80 cells
(Fig. 2A and B). In addition, shRNA was used to
establish stable DEC1 knockdown SKOV3 and OVCAR3 cells, and
transfection efficiency was determined using western blotting. It
was found that shDEC1 significantly inhibited the expression of
DEC1 in both SKOV3 and OVCAR3 cells compared with ISOE80 cells
(Fig. 2C).
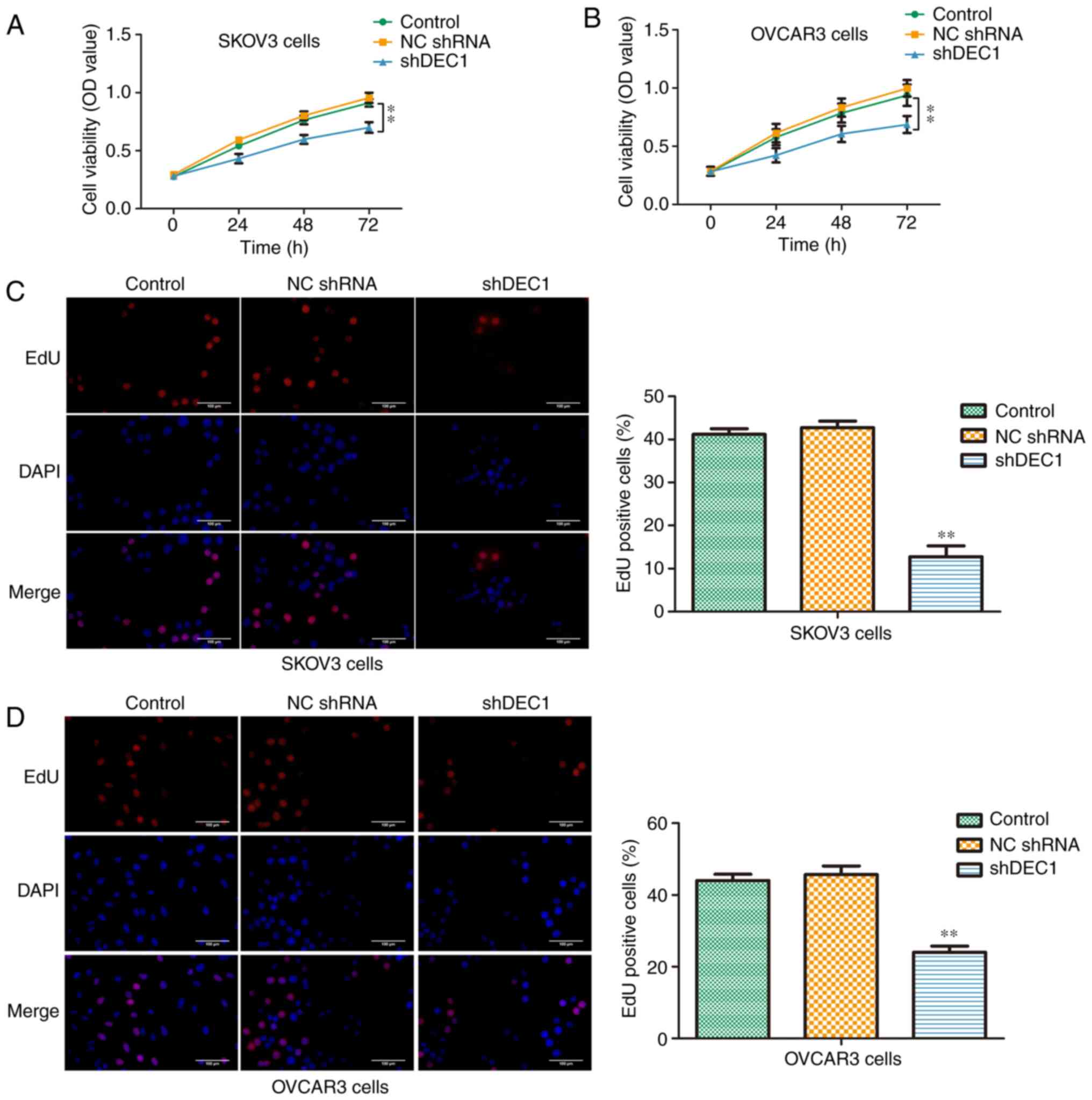
Knockdown of DEC1 inhibits
proliferation of OC cells
Firstly, a CCK-8 assay was performed to evaluate the
viabilities of SKOV3 and OVCAR3 cells transfected with shDEC1 for
0, 24, 48 and 72 h. Knockdown of DEC1 significantly inhibited the
viabilities of both cell lines compared with negative control
(Fig. 3A and B). Moreover, an EdU assay was used to
assess the suppressive effects of shDEC1 on the proliferation of
SKOV3 and OVCAR3 cells. It was demonstrated that knockdown of DEC1
could significantly decrease the number of EdU positive cells
(Fig. 3C and D). These data indicated that DEC1
knockdown inhibited the proliferation of SKOV3 and OVCAR3
cells.
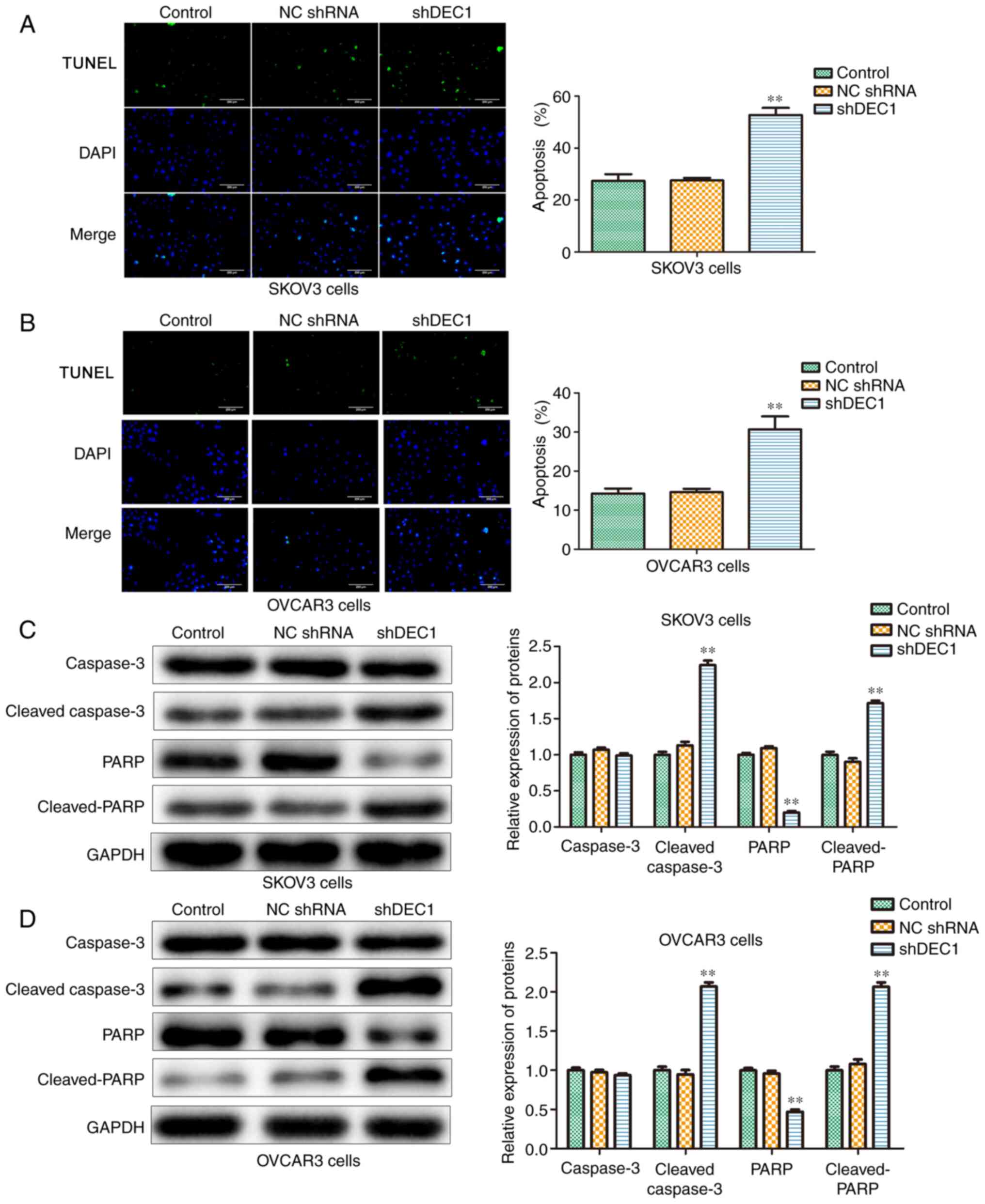
Knockdown of DEC1 induces apoptosis of
OC cells
The effects of DEC1 on apoptosis of SKOV3 and OVCAR3
cells were evaluated using TUNEL assay, and the results
demonstrated that inhibition of DEC1 significantly increased
apoptosis in these cells compared with the control group (Fig. 4A and B).
Furthermore, western blotting was conducted to
examine the role of shDEC1 in apoptotic-related proteins including
caspase-3, cleaved-caspase-3, PARP and cleaved-PARP. Knockdown of
DEC1 significantly increased the expression levels of
cleaved-caspase-3 and cleaved-PARP in both SKOV3 and OVCAR3 cells
(Fig. 4C and D). Therefore, it was identified that DEC1
knockdown promoted apoptosis of SKOV3 and OVCAR3 cells.
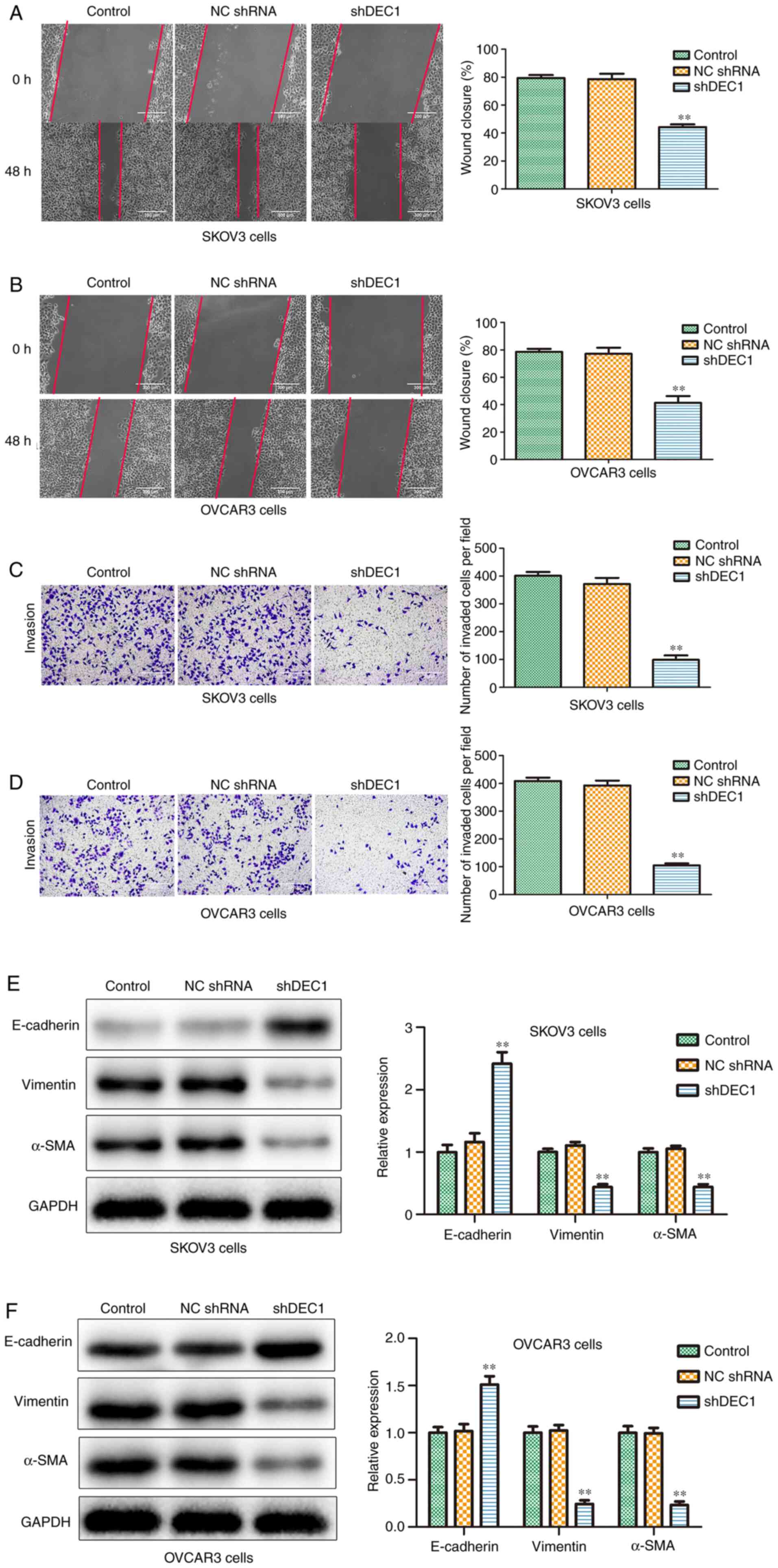
Knockdown of DEC1 inhibits migration
and invasion of OC cells
To investigate the potential role of shDEC1 in
migration and invasion of OC cells, wound healing and Transwell
invasion analysis were performed. The wounding healing rates in
shDEC1 group were reduced compared with the control group in both
SKOV3 and OVCAR3 cells (Fig. 5A and
B). Moreover, the results of the
Transwell invasion assay (Fig. 5C
and D) indicated that after
transfection with shDEC1, the invaded cell numbers were
significantly suppressed compared with the control group.
Epithelial-mesenchymal transition (EMT) provides
invasive and motile abilities for OC cells, and is considered to be
an important factor in invasion and metastasis (28). Thus, western blotting was conducted
to evaluate the effects of shDEC1 on the expression levels of
E-cadherin, vimentin and α-SMA. It was identified that knockdown of
DEC1 significantly increased the expression of E-cadherin, but
significantly decreased the expression levels of vimentin and α-SMA
in both cell types (Fig. 5E and
F). Therefore, the results
suggested that DEC1 knockdown inhibited the migration and invasion
of SKOV3 and OVCAR3 cells.
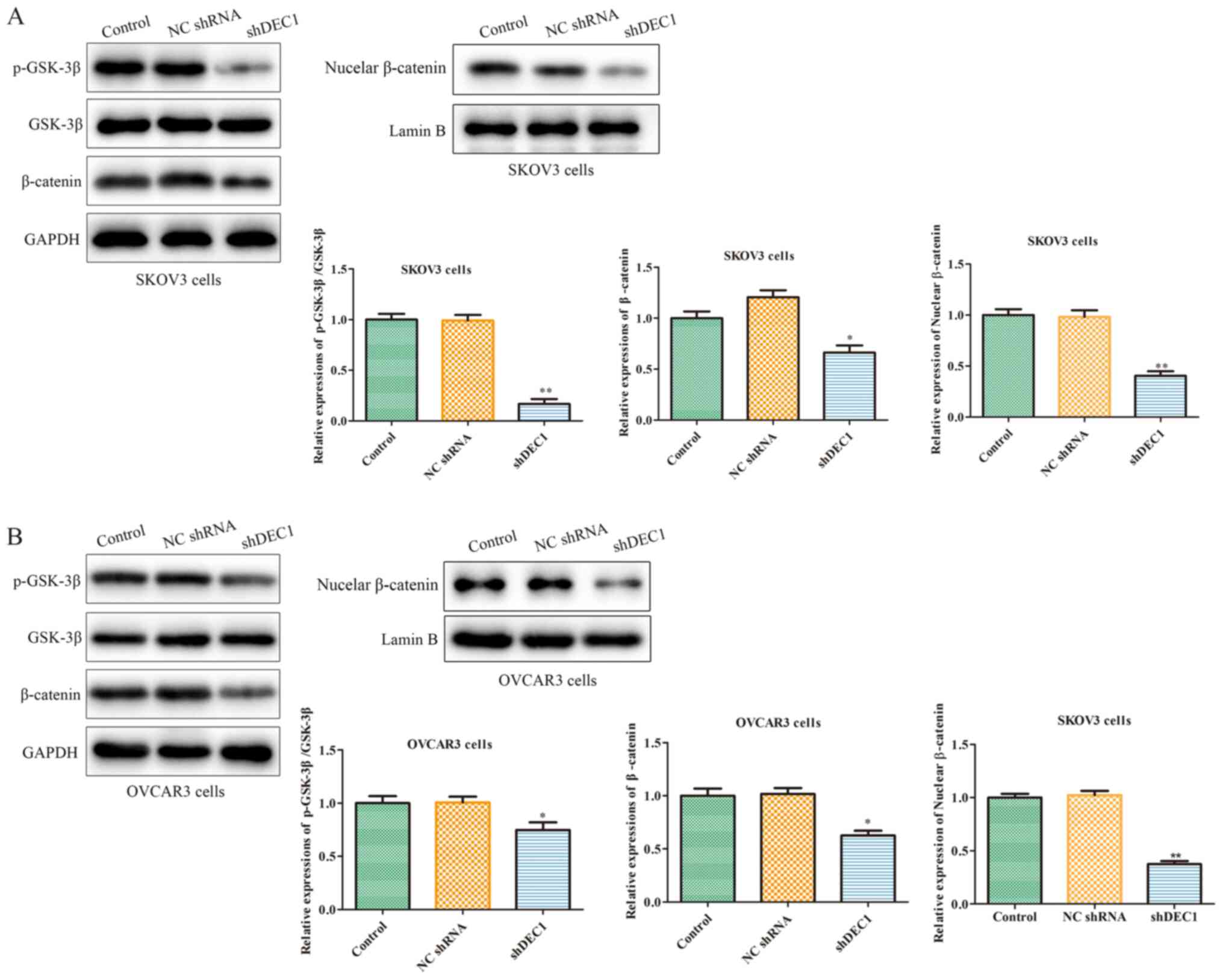
Knockdown of DEC1 inhibits the
Wnt/β-catenin signaling pathway of OC cells
The occurrence and development of tumors are
regulated by numerous factors, among which abnormally activated
cell signaling pathways serve a crucial role (29). For example, the Wnt/β-catenin
signaling pathway participates in the occurrence of various human
tumors, including OC (30).
Therefore, western blotting was used to assess the effects of
shDEC1 on the expression of the Wnt/β-catenin pathway, and the
results demonstrated that inhibition of DEC1 significantly
inhibited the expression levels of p-GSK-3β and β-catenin in SKOV3
and OVCAR3 cells (Fig. 6A and
B).
Discussion
OC is common form of malignant tumors worldwide, and
the molecular mechanisms of OC occurrence and development have been
a hot topic in life science research (31,32).
Changes in the expression levels of tumor-related genes or specific
genes, and activation of multiple signal transduction proteins and
transcription factors can lead to the occurrence and development of
OC (33). Among these,
transcription factors exhibit key effects on regulating the growth,
evolution and metastasis of OC by modulating oncogenes and tumor
suppressor genes (34). Previous
studies have reported that DEC1 is highly expressed in several
tumors, and regulates the expression of genes closely associated
with cancer via its complex transcriptional regulatory network. For
instance, DEC1 expression is corrected with the malignancy and
invasiveness of breast cancer, and high expression of DEC1 is
positively associated with the grade of breast cancer (35). Moreover, the expression of DEC1 in
oral squamous cell carcinoma is higher compared with healthy oral
mucosa (36). It has also been
shown that DEC1 expression is highest in recurrent oral squamous
cell carcinoma within 1 year and lowest in non-recurrent oral
squamous cell carcinoma within 3 years, indicating that DEC1 is
negatively correlated with the prognosis of oral squamous cell
carcinoma (36). DEC1 affects the
progression of gastric cancer by upregulating hypoxia inducible
factor 1 subunit α (37), and
overexpression of DEC1 can enhance reactive oxygen species to
activate the PI3K/Akt signaling pathway (10). The results of the present study
indicated that DEC1 was upregulated in OC tissues and cell lines,
and the expression of DEC1 was negatively associated to the
prognosis of patients with OC.
Malignant proliferation is one of the important
biological characteristics of cancer cells (38). OC has a malignant phenotype
involving infinite proliferation and anti-apoptosis (39). In the malignant development of OC,
abnormal expression levels of multiple genes lead to cell
proliferation, apoptosis reduction and cell dedifferentiation,
which eventually lead to uncontrolled cell proliferation (40,41). A
previous study has shown that silencing DEC1 inhibited the
proliferation of breast cancer by inducing cell cycle arrest in S
phase (17). Similarly, the current
findings demonstrated that knockdown of DEC1 inhibited OC cell
proliferation, using CCK-8 and EdU assay.
Apoptosis is an autonomous and orderly cell death
involving the activation, expression and regulation of a series of
genes under physiological or pathological conditions (42). In the present study, knockdown of
DEC1 induced apoptosis of OC cells, as well as the expression of
cleaved-caspase-3 and cleaved-PARP proteins, which was consistent
with the results of Li et al (43), who reported that downregulation of
DEC1 can inhibit the apoptosis induced by serum starvation in colon
cancer cells and selectively inhibit the activities of caspases-3,
7 and 9.
Cell metastasis has positive role in embryonic
development, maintaining homeostasis of intracellular environment,
wound healing and other physiological functions, and is also
associated with pathophysiological processes such as cancer
metastasis (44). The process of
metastasis is involved in changes of cell structure, cytoskeleton
dynamics, expression levels of adhesion molecules and activation of
signals necessary for cell migration (45,46).
Therefore, the migration and invasion of cancer cells are a complex
process involving multiple genes, steps and factors. A previous
study revealed that transforming growth factor-β induced PANC-1
cells to gain a spindle-shaped morphology, while treatment with
shDEC1 reversed the effects and inhibited the migration and
invasion of PANC-1 cells (47),
which was in line with the present results. In the present study,
it was found that knockdown of DEC1 suppressed the migration and
invasion of OC cells.
EMT is a reversible biological process involving
changes in cell morphology and features (48). In the process of EMT, the expression
levels of epithelial markers, such as E-cadherin and keratin, are
upregulated, while the expression levels of interstitial markers,
such as vimentin and N-cadherin, are increased. These effects
subsequently lead to increased secretion of MMPs and extracellular
matrix proteins, as well as the destruction of basement membrane
and extracellular matrix (49).
Moreover, EMT in epithelial cell tumors provides invasive and
motile abilities for tumor cells, and is considered to be an
essential factor in invasion and metastasis (50). Therefore, the effects of EMT in the
invasion and metastasis of cancer, including OC, and its regulation
mechanisms have gained increased attention in recent years. In the
present study, it was demonstrated that knockdown of DEC1
significantly decreased the migratory and invasive abilities of OC
cells.
The Wnt/β-catenin pathway is highly conserved in the
evolutionary process and participates in the regulation of cell
proliferation and differentiation during embryonic development
(51). β-catenin, as an oncogene
and a component of the Wnt signaling transduction pathway,
accumulates in the nucleus and forms a complex with TCF/LEF of
DNA-binding transcription factor family to enhance the expression
of various genes, which not only promotes the proliferation of
cancer cells, but also activates the transcriptional expressions of
several genes, such as endoplasmic reticulum membrane protein
complex proteins, fibronectin and MMPs, so that tumor cells are
loose and prone to invasion and metastasis (52,53).
Furthermore, there is no Wnt signal in healthy mature cells, and
only a small amount of β-catenin is free in cells; some of which is
phosphorylated by GSK-3β (54).
Rask et al (55) detected
and localized β-catenin and other members of the Wnt family using
immunohistochemistry and western blotting, and revealed that
members of the Wnt family, including β-catenin, were expressed in
healthy human ovarian epithelial cells and the epithelial ovarian
cancer cell line OVCAR3, while the nuclear staining of β-catenin
was only expressed in OVCAR3 cells (55). These results indicated that
β-catenin was located in the cell membrane in healthy cells. When
the Wnt pathway is activated, the activity of GSK-3β is inhibited,
and thus β-catenin cannot be phosphorylated and degraded.
Therefore, β-catenin aggregates and moves into the nucleus,
stimulating the expression levels of a series of oncogenes, which
affects the progression of tumors (56). In the present study, it was
demonstrated that knockdown of DEC1 inhibited the Wnt/β-catenin
pathway.
In conclusion, the present study identified that
DEC1 was highly expressed in OC tissues and cell lines, and high
expression of DEC1 was positively associated with the prognosis of
patients with OC. Moreover, knockdown of DEC1 inhibited the
proliferation, migration and invasion, and induced apoptosis of OC
cells, which may be achieved by modulating the Wnt/β-catenin
pathway. However, in the experimental design of the present study
only SKOV3 and OVCAR3 cell lines were included, and additional
ovarian cancer cells should be investigated in future studies.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YYi, BT and XinY conceived and designed the study.
YYi, BL, ZZ, XiaY, YZ and YYa performed the literature search and
acquisition of data. YYi, BL, ZZ and XiaY contributed to the
analysis and interpretation of data. YYi and BT drafted and revised
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
This study has obtained approval from the Jiangxi
Cancer Hospital Medical Ethics Committee and oral informed consent
from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Norouzi-Barough L, Sarookhani MR, Sharifi
M, Moghbelinejad S, Jangjoo S and Salehi R: Molecular mechanisms of
drug resistance in ovarian cancer. J Cell Physiol. 233:4546–4562.
2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Wang H, Xu T, Zheng L and Li G:
Angiogenesis inhibitors for the treatment of ovarian cancer: An
updated systematic review and meta-analysis of randomized
controlled trials. Int J Gynecol Cancer. 28:903–914.
2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Hirte HW: Profile of erlotinib and its
potential in the treatment of advanced ovarian carcinoma. Onco
Targets Ther. 6:427–435. 2013.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Du Bois A and Pfisterer J: Future options
for first-line therapy of advanced ovarian cancer. Int J Gynecol
Cancer. (Suppl 1):S42–S50. 2005.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Miller KD, Siegel RL, Lin CC, Mariotto AB,
Kramer JL, Rowland JH, Stein KD, Alteri R and Jemal A: Cancer
treatment and survivorship statistics, 2016. CA Cancer J Clin.
66:271–289. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Torre LA, Trabert B, DeSantis CE, Miller
KD, Samimi G, Runowicz CD, Gaudet MM, Jemal A and Siegel RL:
Ovarian cancer statistics, 2018. CA Cancer J Clin. 68:284–296.
2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Sato S and Itamochi H: Neoadjuvant
chemotherapy in advanced ovarian cancer: Latest results and place
in therapy. Ther Adv Med Oncol. 6:293–304. 2014.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Bhawal UK, Sato F, Arakawa Y, Fujimoto K,
Kawamoto T, Tanimoto K, Ito Y, Sasahira T, Sakurai T, Kobayashi M,
et al: Basic Helix-loop-helix transcription factor DEC1 negatively
regulates cyclin D1. J Pathol. 224:420–429. 2011.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kato Y, Kawamoto T, Fujimoto K and Noshiro
M: DEC1/STRA13/SHARP2 and DEC2/SHARP1 coordinate physiological
processes, including circadian rhythms in response to environmental
stimuli. Curr Top Dev Biol. 110:339–372. 2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Miyazaki K, Miyazaki M, Guo Y, Yamasaki N,
Kanno M, Honda Z, Oda H, Kawamoto H and Honda H: The role of the
basic Helix-loop-helix transcription factor Dec1 in the regulatory
T cells. J Immunol. 185:7330–7339. 2010.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Liu Y, Wang L, Lin XY, Wang J, Yu JH, Miao
Y and Wang EH: The transcription factor DEC1
(BHLHE40/STRA13/SHARP-2) is negatively associated with TNM stage in
non-small-cell lung cancer and inhibits the proliferation through
cyclin D1 in A549 and BE1 cells. Tumour Biol. 34:1641–1650.
2013.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Xu Q, Ma P, Hu C, Chen L, Xue L, Wang Z,
Liu M, Zhu H, Xu N and Lu N: Overexpression of the DEC1 protein
induces senescence in vitro and is related to better survival in
esophageal squamous cell carcinoma. PLoS One.
7(e41862)2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Jia Y, Hu R, Li P, Zheng Y, Wang Y and Ma
X: DEC1 is required for anti-apoptotic activity of gastric cancer
cells under hypoxia by promoting Survivin expression. Gastric
Cancer. 21:632–642. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Murakami K, Wu Y, Imaizumi T, Aoki Y, Liu
Q, Yan X, Seino H, Yoshizawa T, Morohashi S, Kato Y and Kijima H:
DEC1 promotes hypoxia-induced epithelial-mesenchymal transition
(EMT) in human hepatocellular carcinoma cells. Biomed Res.
38:221–227. 2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Bi H, Li S, Qu X, Wang M, Bai X, Xu Z, Ao
X, Jia Z, Jiang X, Yang Y and Wu H: DEC1 regulates breast cancer
cell proliferation by stabilizing cyclin E protein and delays the
progression of cell cycle S phase. Cell Death Dis.
6(e1891)2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Hu J, Mao Z, He S, Zhan Y, Ning R, Liu W,
Yan B and Yang J: Icariin protects against glucocorticoid induced
osteoporosis, increases the expression of the bone enhancer DEC1
and modulates the PI3K/Akt/GSK3β/β-catenin integrated signaling
pathway. Biochem Pharmacol. 136:109–121. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhu Z, Wang YW, Ge DH, Lu M, Liu W, Xiong
J, Hu G, Li XP and Yang J: Downregulation of DEC1 contributes to
the neurotoxicity induced by MPP(+) by suppressing PI3K/Akt/GSK3β
pathway. CNS Neurosci Ther. 23:736–747. 2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Shi J, Zhu Q, Wu J and Zhu P: FAM46C
suppresses gastric cancer by inhibition of Wnt/beta-catenin. Front
Biosci (Landmark Ed). 25:549–563. 2020.PubMed/NCBI
|
|
20
|
Zhang C, Zhang Z, Zhang S, Wang W and Hu
P: Targeting of Wnt/β-catenin by anthelmintic drug pyrvinium
enhances sensitivity of ovarian cancer cells to chemotherapy. Med
Sci Monit. 23:266–275. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Hu Z, Wang P, Lin J, Zheng X, Yang F,
Zhang G, Chen D, Xie J, Gao Z, Peng L and Xie C: MicroRNA-197
Promotes metastasis of hepatocellular carcinoma by activating
wnt/β-catenin signaling. Cell Physiol Biochem. 51:470–486.
2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kumar R, Kotapalli V, Naz A, Gowrishankar
S, Rao S, Pollack JR and Bashyam MD: XPNPEP3 is a novel
transcriptional target of canonical Wnt/β-catenin signaling. Genes
Chromosomes Cancer. 57:304–310. 2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wang Y, Lei L, Zheng YW, Zhang L, Li ZH,
Shen HY, Jiang GY, Zhang XP, Wang EH and Xu HT: Odd-skipped related
1 inhibits lung cancer proliferation and invasion by reducing Wnt
signaling through the suppression of SOX9 and β-catenin. Cancer
Sci. 109:1799–1810. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Raghavan S, Mehta P, Xie Y, Lei YL and
Mehta G: Ovarian cancer stem cells and macrophages reciprocally
interact through the WNT pathway to promote pro-tumoral and
malignant phenotypes in 3D engineered microenvironments. J
Immunother Cancer. 7(190)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Kim MS, Cho HI, Yoon HJ, Ahn YH, Park EJ,
Jin YH and Jang YK: JIB-04, A Small molecule histone demethylase
inhibitor, selectively targets colorectal cancer stem cells by
inhibiting the wnt/β-catenin signaling pathway. Sci Rep.
8(6611)2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhou L, Rui JA, Ye DX, Wang SB, Chen SG
and Qu Q: Edmondson-Steiner grading increases the predictive
efficiency of TNM staging for long-term survival of patients with
hepatocellular carcinoma after curative resection. World J Surg.
32:1748–1756. 2008.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zhou J, Du Y, Lu Y, Luan B, Xu C, Yu Y and
Zhao H: CD44 expression predicts prognosis of ovarian cancer
patients through promoting Epithelial-mesenchymal transition (EMT)
by regulating snail, ZEB1, and caveolin-1. Front Oncol.
9(802)2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Gonçalves V, Pereira JFS and Jordan P:
Signaling pathways driving aberrant splicing in cancer cells. Genes
(Basel). 9(9)2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Arend RC, Londoño-Joshi AI, Straughn JM Jr
and Buchsbaum DJ: The Wnt/β-catenin pathway in ovarian cancer: A
review. Gynecol Oncol. 131:772–779. 2013.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Yeung TL, Leung CS, Yip KP, Au Yeung CL,
Wong ST and Mok SC: Cellular and molecular processes in ovarian
cancer metastasis. A review in the theme: Cell and molecular
processes in cancer metastasis. Am J Physiol Cell Physiol.
309:C444–C456. 2015.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Grunewald T and Ledermann JA: Targeted
therapies for ovarian cancer. Best Pract Res Clin Obstet Gynaecol.
41:139–152. 2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Kroeger PT Jr and Drapkin R: Pathogenesis
and heterogeneity of ovarian cancer. Curr Opin Obstet Gynecol.
29:26–34. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Kaldawy A, Segev Y, Lavie O, Auslender R,
Sopik V and Narod SA: Low-grade serous ovarian cancer: A review.
Gynecol Oncol. 143:433–438. 2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Liu Y, Miao Y, Wang J, Lin X, Wang L, Xu
HT and Wang EH: DEC1 is positively associated with the malignant
phenotype of invasive breast cancers and negatively correlated with
the expression of claudin-1. Int J Mol Med. 31:855–860.
2013.PubMed/NCBI View Article : Google Scholar
|
|
36
|
You J, Lin L, Liu Q, Zhu T, Xia K and Su
T: The correlation between the expression of differentiated
embryo-chondrocyte expressed gene l and oral squamous cell
carcinoma. Eur J Med Res. 19(21)2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Yan Z, Shi X, Min W, Jia Y, Li B, Zhang Y,
Liu Q and Wang Y: The increased expression of DEC1 gene is related
to HIF-1α protein in gastric cancer cell lines. Mol Biol Rep.
39:4229–4236. 2012.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Czekierdowska S, Stachowicz N, Chróściel M
and Czekierdowski A: Proliferation and maturation of intratumoral
blood vessels in women with malignant ovarian tumors assessed with
cancer stem cells marker nestin and platelet derived growth factor
PDGF-B. Ginekol Pol. 88:120–128. 2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Wei ZT, Zhang X, Wang XY, Gao F, Zhou CJ,
Zhu FL, Wang Q, Gao Q, Ma CH, Sun WS, et al: PDCD4 inhibits the
malignant phenotype of ovarian cancer cells. Cancer Sci.
100:1408–1413. 2009.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Zuo K, Zhao Y, Zheng Y, Chen D, Liu X, Du
S and Liu Q: Long non-coding RNA XIST promotes malignant behavior
of epithelial ovarian cancer. Onco Targets Ther. 12:7261–7267.
2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Yu HL, Ma XD, Tong JF, Li JQ, Guan XJ and
Yang JH: WTAP is a prognostic marker of high-grade serous ovarian
cancer and regulates the progression of ovarian cancer cells. Onco
Targets Ther. 12:6191–6201. 2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Twomey C and Mccarthy JV: Pathways of
apoptosis and importance in development. J Cell Mol Med. 9:345–359.
2010.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Li Y, Zhang H, Xie M, Hu M, Ge S, Yang D,
Wan Y and Yan B: Abundant expression of Dec1/stra13/sharp2 in colon
carcinoma: Its antagonizing role in serum deprivation-induced
apoptosis and selective inhibition of procaspase activation.
Biochem J. 367:413–422. 2002.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Liu Y, Ren CC, Yang L, Xu YM and Chen YN:
Role of CXCL12-CXCR4 axis in ovarian cancer metastasis and
CXCL12-CXCR4 blockade with AMD3100 suppresses tumor cell migration
and invasion in vitro. J Cell Physiol. 234:3897–3909.
2019.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Al Ameri W, Ahmed I, Al-Dasim FM, Ali
Mohamoud Y, Al-Azwani IK, Malek JA and Karedath T: Cell
type-specific TGF-β mediated EMT in 3D and 2D models and its
reversal by TGF-β receptor kinase inhibitor in ovarian cancer cell
lines. Int J Mol Sci. 20(3568)2019.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Zhang H, Wang Y, Chen T, Zhang Y, Xu R,
Wang W, Cheng M and Chen Q: Aberrant activation of hedgehog
signalling promotes cell migration and invasion via matrix
metalloproteinase-7 in ovarian cancer cells. J Cancer. 10:990–1003.
2019.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Wu Y, Sato F, Yamada T, Bhawal UK,
Kawamoto T, Fujimoto K, Noshiro M, Seino H, Morohashi S, Hakamada
K, et al: The BHLH transcription factor DEC1 plays an important
role in the epithelial-mesenchymal transition of pancreatic cancer.
Int J Oncol. 41:1337–1346. 2012.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Duan H, Yan Z, Chen W, Wu Y, Han J, Guo H
and Qiao J: TET1 inhibits EMT of ovarian cancer cells through
activating Wnt/β-catenin signaling inhibitors DKK1 and SFRP2.
Gynecol Oncol. 147:408–417. 2017.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Felipe Lima J, Nofech-Mozes S, Bayani J
and Bartlett JM: EMT in breast carcinoma-a review. J Clin Med.
5(65)2016.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Chen Y, Wang DD, Wu YP, Su D, Zhou TY, Gai
RH, Fu YY, Zheng L, He QJ, Zhu H and Yang B: MDM2 promotes
epithelial-mesenchymal transition and metastasis of ovarian cancer
SKOV3 cells. Br J Cancer. 117:1192–1201. 2017.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Walston T, Tuskey C, Edgar L, Hawkins N,
Ellis G, Bowerman B, Wood W and Hardin J: Multiple Wnt signaling
pathways converge to orient the mitotic spindle in early C.
elegans embryos. Dev Cell. 7:831–841. 2004.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Yang S, Liu Y, Li MY, Ng CSH, Yang SL,
Wang S, Zou C, Dong Y, Du J, Long X, et al: FOXP3 promotes tumor
growth and metastasis by activating Wnt/β-catenin signaling pathway
and EMT in non-small cell lung cancer. Mol Cancer.
16(124)2017.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Wang H, Wu M, Lu Y, He K, Cai X, Yu X, Lu
J and Teng L: LncRNA MIR4435-2HG targets desmoplakin and promotes
growth and metastasis of gastric cancer by activating Wnt/β-catenin
signaling. Aging (Albany NY). 11:6657–6673. 2019.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Smolders LA, Meij BP, Riemers FM, Licht R,
Wubbolts R, Heuvel D, Grinwis GC, Vernooij HC, Hazewinkel HA,
Penning LC and Tryfonidou MA: Canonical Wnt signaling in the
notochordal cell is upregulated in early intervertebral disk
degeneration. J Orthop Res. 30:950–957. 2012.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Rask K, Nilsson A, Brännström M, Carlsson
P, Hellberg P, Janson PO, Hedin L and Sundfeldt K: Wnt-signalling
pathway in ovarian epithelial tumours: Increased expression of
beta-catenin and GSK3beta. Br J Cancer. 89:1298–1304.
2003.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Li J, Yin J, Shen W, Gao R, Liu Y, Chen Y,
Li X, Liu C, Xiang R and Luo N: TLR4 promotes breast cancer
metastasis via Akt/GSK3β/β-catenin pathway upon LPS stimulation.
Anat Rec (Hoboken). 300:1219–1229. 2017.PubMed/NCBI View Article : Google Scholar
|