Introduction
Hypertrophic scars (HS) caused by various traumas
are a product of the healing process and seriously affect the
patient's physical health (1-3).
HS is characterized by hyperproliferation of scar fibroblasts and
excessive accumulation of extracellular matrix (ECM) (4,5). At
present, the main treatment strategy for HS is surgical excision
with superficial X-ray radiation and dot-array laser (6); however, surgical treatment may cause
secondary injuries (7), and
radiation therapy can cause radiation dermatitis, delayed incision
healing and skin canceration at the irradiation site (6,7).
Therefore, identifying the molecular mechanisms underlying HS
formation to aid with the development of novel therapeutic
strategies for HS is important.
MicroRNAs (miRNAs/miRs) are endogenous, non-coding,
single-stranded small RNAs, which regulate gene and protein
expression at the post-transcriptional level by binding to target
genes in a complementary or incompletely complementary manner
(8). Evidence has indicated that
miRNAs serve key roles in pathophysiological processes, such as
cell proliferation, apoptosis, organ development and immune
regulation (8-12).
Increasing evidence has also suggested that miRNAs serve a critical
role in HS formation (13-16).
miR-497-5p has been reported to serve critical roles in various
types of cancer via regulating cell proliferation and apoptosis,
and modulating the expression of several genes and proteins
(17-20).
In addition, the important roles of miR-497-5p in myofibroblast
differentiation and pulmonary fibrogenesis have been reported
(21). The aforementioned studies
indicated that miR-497-5p may serve an important role in the
occurrence and development of HS. However, the role of miR-497-5p
in HS formation is not completely understood.
Smad7 serves as a negative regulator of transforming
growth factor (TGF)-β signaling via multiple mechanisms, such as
TGF-β type I receptor degradation induction, Smad2/3
phosphorylation and recruitment blocking (22,23).
Smad7 can protect against TGF-β-induced fibrosis in various organs,
including the lungs, kidneys and liver (24,25).
Several studies have reported an important role of Smad7 in the
regulation of fibroblast proliferation and ECM accumulation
(26-28),
indicating an important role for Smad7 in HS formation.
The aim of the present study was to investigate the
expression and role of miR-497-5p in HS formation, and to further
explore the underlying molecular mechanism.
Materials and methods
Clinical samples
A total of 22 HS tissues (age 27-59 years; 11 male
patients and 11 female patients) were collected during scar
excision at Shanghai Meizhen Medical Cosmetology Clinic.
Additionally, 22 healthy control skin tissues (age, 24-63 years; 11
male patients and 11 female patients) were collected during
auto-skin grafting at Shanghai Meizhen Medical Cosmetology Clinic
between May 23, 2017 and June 4, 2019. Written informed consent was
obtained from the patients and all patients approved the use of
their samples in the present study. The present study was approved
by the Ethics Committee of Shanghai Meizhizhen Medical Cosmetology
Clinic.
Cell culture and cell
transfection
Human embryonic skin fibroblasts CCC-ESF-1 (cat. no.
YB-ATCC-3084; Shanghai Zibo Biological Technology Co., Ltd.) and
human HS fibroblasts (hHSFs; cat. no. C0618; Shanghai Guandao
Biological engineering Co., Ltd.) were cultured in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) at 37˚C with 5% CO2.
Control-plasmid (1 µg; cat. no. sc-437275; Santa
Cruz Biotechnology, Inc.), Smad7-plasmid (1 µg; cat. no.
sc-400251-ACT; Santa Cruz Biotechnology, Inc.), miR-497-5p
inhibitor (100 nM; 5'-ACAAACCACAGTGTGCTGCTG-3'; Sangon Biotech Co.,
Ltd.), inhibitor control (100 nM; 5'-CAGTACTTTTGTGTAGTACAA-3';
Sangon Biotech Co., Ltd.), Smad7-short hairpin (sh)RNA (1 µg; cat.
no. sc-36508-SH; Santa Cruz Biotechnology, Inc.), control-shRNA (1
µg; cat. no. sc-108060; Santa Cruz Biotechnology, Inc.) or 100 nM
miR-497-5p inhibitor + 1 µg Smad7-shRNA were transfected into hHSFs
(5x104 per well) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. At 48 h post-transfection,
transfection efficiency was assessed via reverse
transcription-quantitative PCR (RT-qPCR).
Luciferase reporter analysis
The binding sites between miR-497-5p and Smad7 were
predicted by TargetScan version 7.2 (www.targetscan.org/vert_72). To confirm the binding
sites between miR-497-5p and Smad7, a dual-luciferase reporter
assay was conducted. Briefly, the wild-type (WT-Smad7) and mutant
(MUT-Smad7) 3'-untranslated region (UTR) of Smad7 were cloned into
a pmiR-RB-Report™ dual luciferase reporter gene plasmid
vector (Guangzhou RiboBio Co., Ltd.) according to the
manufacturer's instructions. hHSFs (5x104 per well) were
co-transfected with 1 ng WT-Smad7 or 1 ng MUT-Smad7 and 50 nM
miR-497-5p mimic (5'-CAGCAGCACACUGUGGUUUGUAAACCACAGUGUGCUGCUGUU-3';
Guangzhou RiboBio Co., Ltd.) or 50 nM mimic control
(5'-UUCUCCGAACGUGUCACGUTT-3'; Guangzhou RiboBio Co., Ltd.) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Renilla luciferase pRL-TK vector (Promega
Corporation) was used as the internal control. At 48 h
post-transfection, relative luciferase activities were assessed
using the Dual-Luciferase Reporter Assay system (Promega
Corporation). Firefly luciferase activity was normalized to
Renilla luciferase activity.
Cell Counting Kit-8 (CCK-8) assay
The CCK-8 assay (Beyotime Institute of
Biotechnology) was performed to assess hHSF cell viability
according to the manufacturer's protocol. hHSFs seeded
(1x104 cells/well) into 96-well plates were transfected
with 1 µg control-plasmid, 1 µg Smad7-plasmid, 100 nM miR-497-5p
inhibitor, 100 nM inhibitor control or 100 nM miR-497-5p inhibitor
+ 1 µg Smad7-shRNA. At 48 h post-transfection, 10 µl CCK-8 solution
was added to each well and incubated at 37˚C with 5% CO2
for 4 h. Subsequently, hHSF cell viability was assessed by
measuring the absorbance at a wavelength of 490 nm using a
microplate reader.
Flow cytometry assay
At 48 h post-transfection, hHSF apoptosis was
assessed using the Annexin V-FITC/PI apoptosis detection kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's instructions. FACScan flow cytometer (Beckman
Coulter, Inc.) was used to detect early and late hHSF apoptosis.
Data were analyzed using FlowJo software version 7.6.1 (FlowJo
LLC).
RT-qPCR
Total RNA was extracted from tissues or cells using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Total RNA was reverse
transcribed into cDNA using the miScript Reverse Transcription kit
(Qiagen GmbH) according to the manufacturer's instructions. The
conditions for reverse transcription were as follows: 70˚C for 5
min, 37˚C for 5 min and 42˚C for 60 min. Subsequently, qPCR was
performed using the QuantiFast SYBR Green PCR kit (Qiagen GmbH).
The following thermocycling conditions were used for qPCR: Initial
denaturation at 95˚C for 10 min; followed by 35 cycles of 15 sec at
95˚C, 40 sec at 55˚C and a final extension step at 72˚C for 30 sec.
miRNA and mRNA expression levels were quantified using the
2-ΔΔCq method (29) and
normalized to the internal reference genes U6 and GAPDH,
respectively. The primer sequences used for PCR were as follows:
GAPDH forward, 5'-TGTTGCCATCAATGACCCCTT-3' and reverse,
5'-CTCCACGACGTACTCAGCG-3'; Smad7 forward,
5'-GCTCCCATCCTGTGTGTTAA-3' and reverse,
5'-TAGGTGTCAGCCTAGGATGGT-3'; U6 forward,
5'-GCTTCGGCAGCACATATACTAAAAT-3' and reverse,
5'-CGCTTCACGAATTTGCGTGTCAT-3'; miR-497-5p forward,
5'-ATCCAGTGCGTGTCGTG-3' and reverse, 5'-TGCTCAGCAGCACACTGT-3'.
Western blotting
Total protein was extracted from tissues or cells
using RIPA lysis buffer (Beyotime Institute of Biotechnology). A
bicinchoninic acid assay kit (Thermo Fisher Scientific, Inc.) was
used to measure protein concentrations. Proteins (40 µg/lane) were
separated via 12% SDS-PAGE gel and electrically transferred onto
PVDF membranes. After blocking with 5% skim milk at room
temperature for 1.5 h, the membranes were incubated at 4˚C
overnight with primary antibodies targeted against: Smad7 (1:1,000;
cat. no. ab227309; Abcam), α-smooth muscle actin (α-SMA; 1:1,000;
cat. no. ab32575; Abcam), type I collagen (Col I; 1:1,000; cat. no.
ab34710; Abcam), type III collagen (Col III; 1:1,000; cat. no.
ab7778; Abcam) and GAPDH (1:1,000; cat. no. ab9485; Abcam).
Subsequently, the membranes were incubated with an anti-rabbit
horseradish peroxidase-conjugated IgG secondary antibody (cat. no.
7074; 1:2,000; Cell Signaling Technology, Inc.) at room temperature
for 2 h. Protein bands were visualized using an enhanced
chemiluminescence detection system (Thermo Fisher Scientific,
Inc.). ImageJ version 2.0 software (National Institutes of Health)
was used to quantify band intensities.
Statistical analysis
Data are presented as the mean ± SD of three
independent experiments. Statistical analyses were performed using
SPSS software (version 18.0; SPSS, Inc.). Comparisons between two
groups were analyzed using the Student's t-test. Comparisons among
multiple groups were analyzed using one-way ANOVA followed by
Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-497-5p is upregulated in HS
tissues and hHSFs
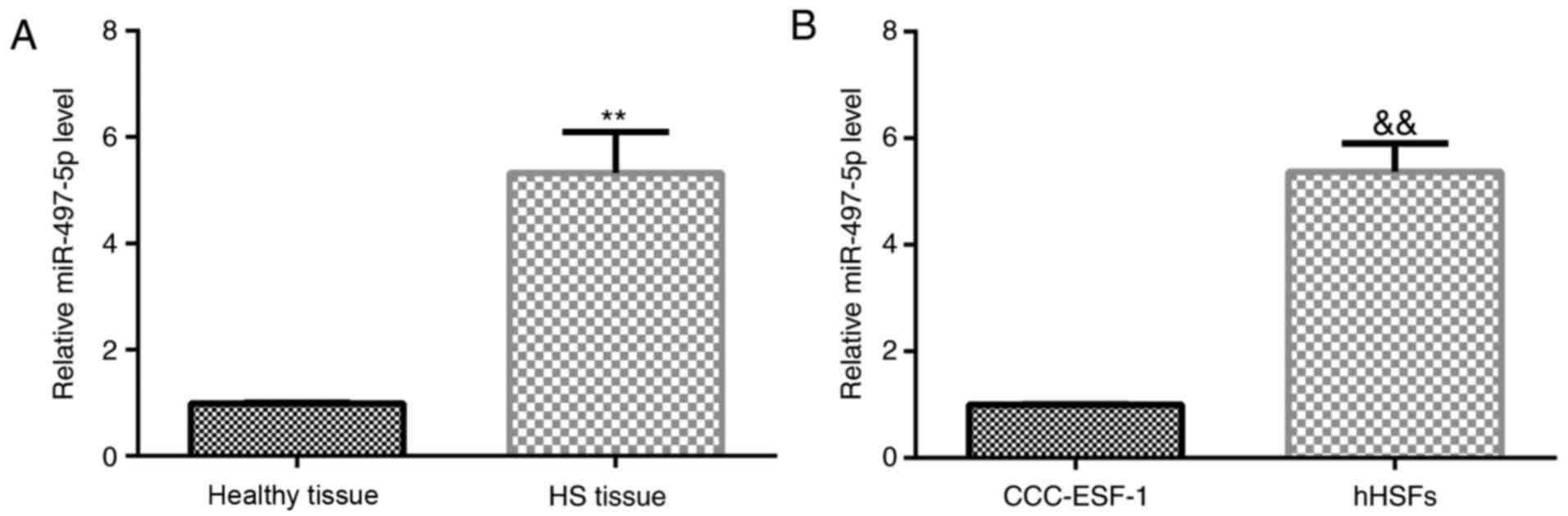
miR-497-5p expression levels in 22 HS tissues, 22
healthy control skin tissues, human embryonic skin fibroblasts
CCC-ESF-1 and hHSFs were determined via RT-qPCR. The results
indicated that the expression levels of miR-497-5p were
significantly upregulated in HS tissues compared with healthy
control skin tissues (Fig. 1A).
Similarly, the expression level of miR-497-5p in hHSFs was
significantly higher compared with CCC-ESF-1 cells (Fig. 1B). The results indicated that
miR-497-5p may be involved in the development of HS.
Smad7 is a target gene of
miR-497-5p
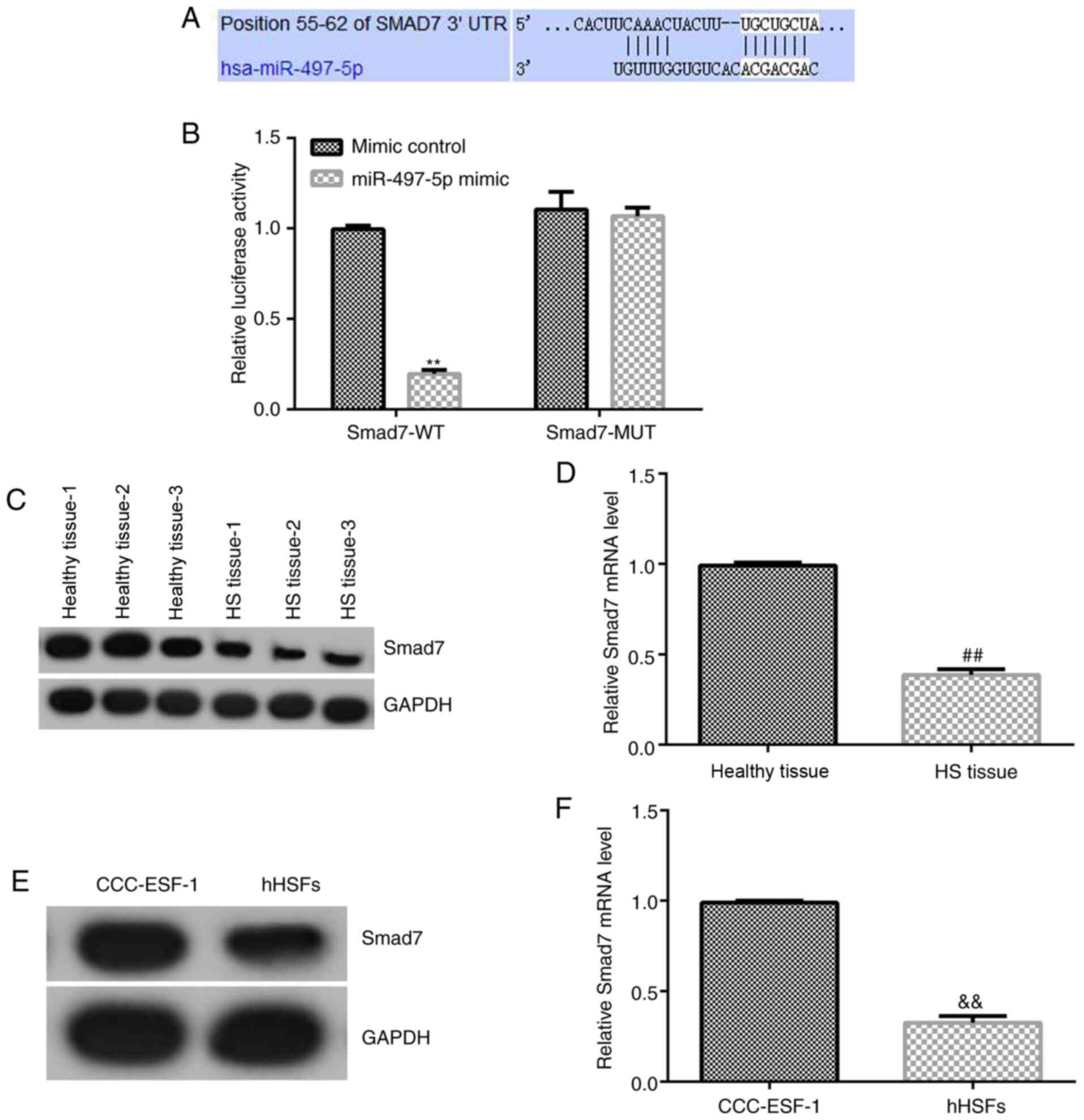
TargetScan was used to predict the potential targets
of miR-497-5p, which indicated a binding site between miR-497-5p
and the 3'UTR of Smad7 mRNA (Fig.
2A). Subsequently, a dual-luciferase reporter assay was
conducted to assess the binding site between miR-497-5p and the
3'UTR of Smad7 (Fig. 2B). Compared
with cells co-transfected with mimic control and Smad7-WT, the
luciferase activity of cells co-transfected with miR-497-5p mimic
and Smad7-WT was significantly reduced. By contrast, there was no
significant difference between the luciferase activity of cells
co-transfected with mimic control and Smad7-MUT, and cells
co-transfected with miR-497-5p mimic and Smad7-MUT. The results
indicated that Smad7 may be a direct target gene of miR-497-5p.
Furthermore, compared with healthy control skin
tissues and CCC-ESF-1 cells, the mRNA and protein expression levels
of Smad7 were significantly downregulated in HS tissues and hHSFs,
respectively (Fig. 2C-F).
Effect of Smad7 on hHSFs
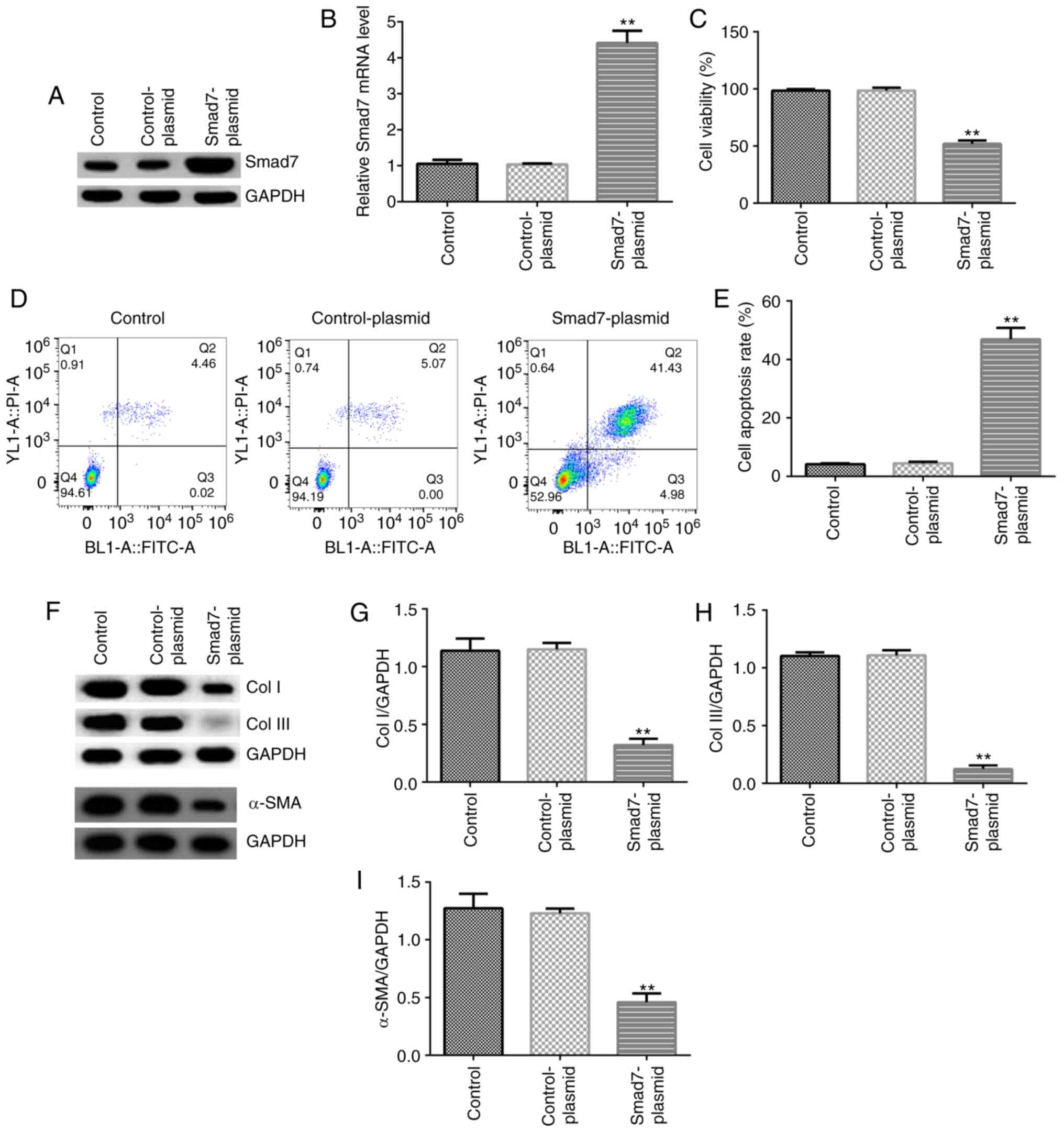
The effect of Smad7 was investigated in hHSFs. hHSFs
were transfected with control-plasmid or Smad7-plasmid for 48 h.
Compared with the control-plasmid group, Smad7-plasmid
significantly increased Smad7 mRNA and protein expression levels in
hHSFs (Fig. 3A and B). In addition, compared with the
control-plasmid group, Smad7-plasmid significantly decreased cell
viability, enhanced apoptosis, and inhibited the protein expression
of Col I, Col III and α-SMA in hHSFs (Fig. 3C-I).
Effect of miR-497-5p on hHSF viability
and apoptosis
The effect of miR-497-5p on hHSF viability and
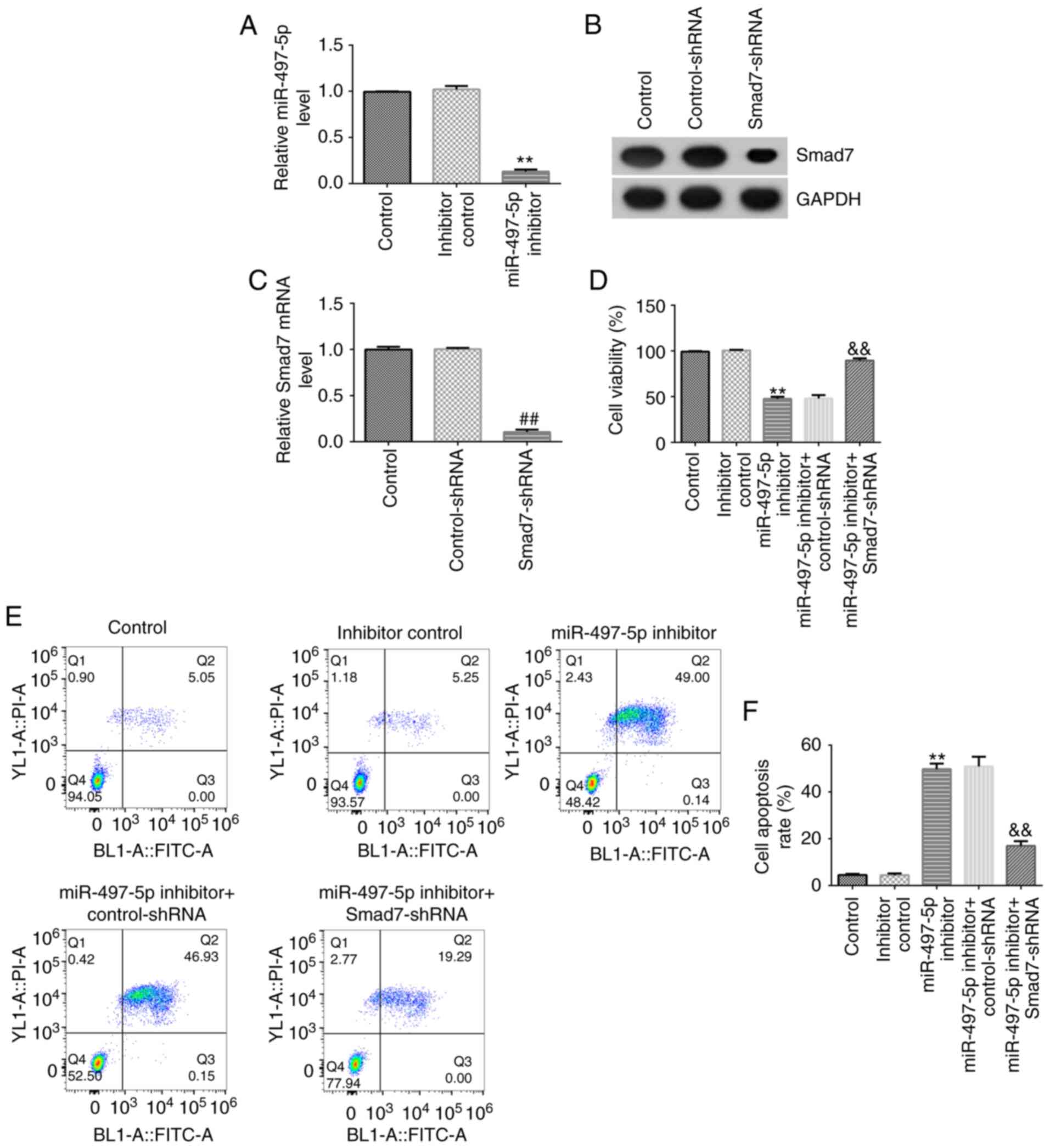
apoptosis was analyzed. Compared with the inhibitor control group,
miR-497-5p inhibitor significantly inhibited miR-497-5p expression
in hHSFs (Fig. 4A). Smad7-shRNA
notably decreased the mRNA and protein expression levels of Smad7
in hHSFs compared with the control-shRNA group (Fig. 4B and C). miR-497-5p inhibitor-mediated reduction
in hHSF viability and induction of apoptosis were reversed by
co-transfection with Smad7-shRNA (Fig.
4D-F).
Effect of miR-497-5p on the protein
expression of Smad7, Col I, Col III and α-SMA in hHSFs
The effect of miR-497-5p on extracellular matrix
deposition in hHSFs was investigated. At 48 h post-transfection,
the expression of Smad7, Col I, Col III and α-SMA in hHSFs was
assessed. The results indicated that miR-497-5p inhibitor markedly
increased the mRNA and protein expression levels of Smad7 in hHSFs
compared with the inhibitor control group (Fig. 5A and B), which was reversed by co-transfection
with Smad7-shRNA. Moreover, the results indicated that miR-497-5p
inhibitor significantly decreased the protein expression levels of
Col I, Col III and α-SMA in hHSFs compared with the inhibitor
control group, which was reversed by co-transfection with
Smad7-shRNA (Fig. 5C-F).
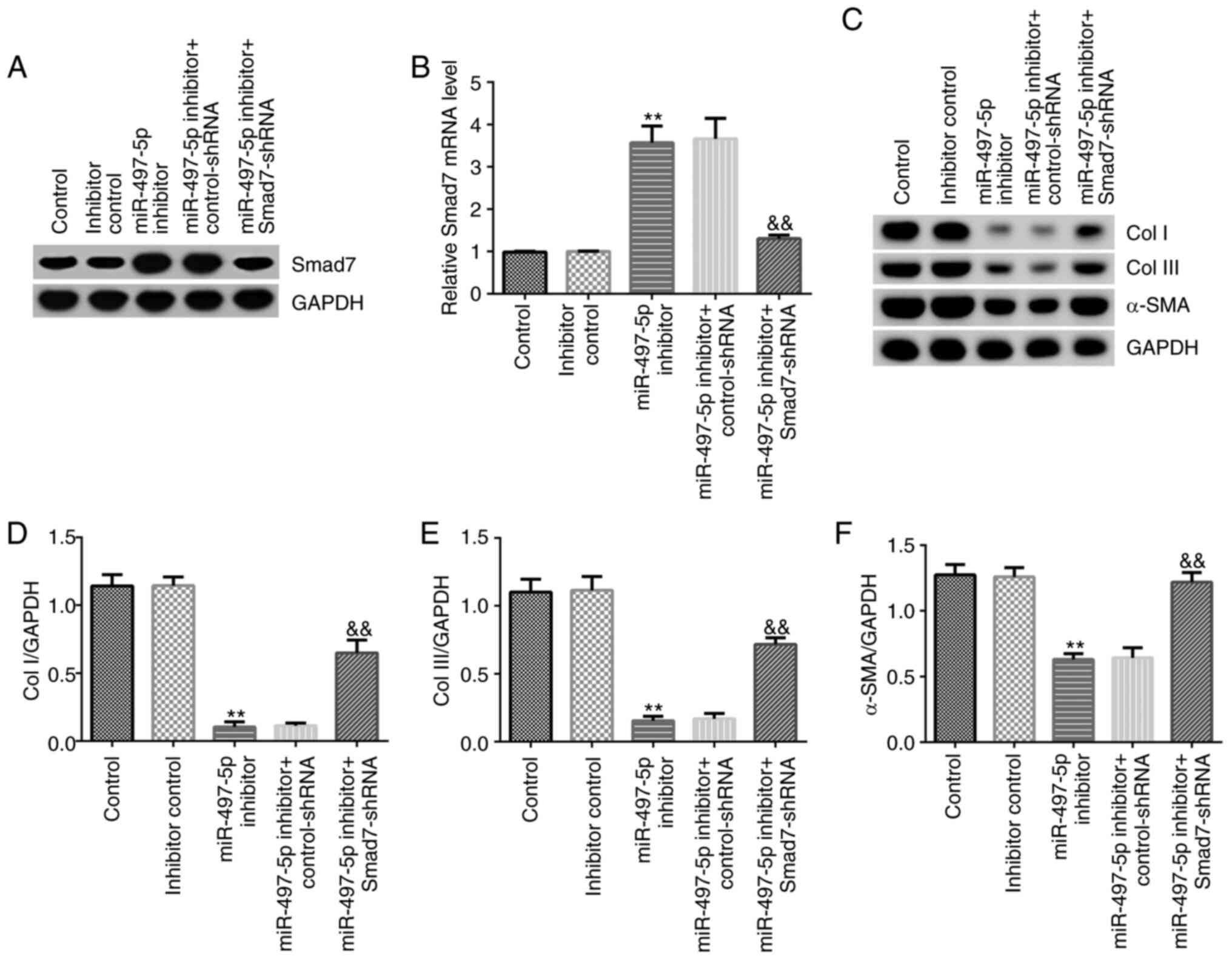 | Figure 5Effect of miR-497-5p inhibitor on
Smad7, Col I, Col III and α-SMA expression in hHSFs. hHSFs were
transfected with control-shRNA, Smad7-shRNA, inhibitor control,
miR-497-5p inhibitor, miR-497-5p inhibitor + control-shRNA or
miR-497-5p inhibitor + Smad7-shRNA for 48 h. (A) Protein and (B)
mRNA expression levels of Smad7. Protein expression levels were (C)
determined by western blotting and semi-quantified for (D) Col I,
(E) Col III and (F) α-SMA. **P<0.01 vs. inhibitor
control &&P<0.01 vs. miR-497-5p inhibitor +
control-shRNA. miR, microRNA; Col I, type I collagen; Col III, type
III collagen; α-SMA, α-smooth muscle actin; hHSFs, human
hyperplastic scar fibroblasts; shRNA, short hairpin RNA. |
Discussion
The present study demonstrated that miR-497-5p was
upregulated in HS tissues and hHSFs compared with healthy control
skin tissues and CCC-ESF-1 cells, respectively. Smad7 was a direct
target of miR-497-5p, and was downregulated in HS tissues and hHSFs
compared with healthy control skin tissues and CCC-ESF-1 cells,
respectively. miR-497-5p downregulation significantly inhibited ECM
deposition, inhibited hHSF proliferation and induced cell apoptosis
compared with the inhibitor-control group, which were all reversed
by Smad7 downregulation. Collectively, the results of the present
study may aid with the development of novel therapeutic strategies
for HS.
To date, HS has been the focus of research on trauma
and plastic surgery (30). miRNAs
have been identified to serve critical roles in the progression of
skin wound repair and skin diseases (31,32).
Research on miRNAs in HS has attracted increasing attention
(33-38).
miR-497-5p, which has been reported to serve critical roles in the
regulation of cell proliferation and apoptosis (17-21),
was investigated in the present study.
The level of miR-497-5p was detected in HS tissues,
healthy control skin tissues, human embryonic skin fibroblasts
CCC-ESF-1 and hHSFs via RT-qPCR. The results suggested that
miR-497-5p was highly expressed in HS tissues and hHSFs compared
with healthy control skin tissues and CCC-ESF-1 cells,
respectively, and Smad7 was identified as a direct target of
miR-497-5p. Consistent with a previous study (39), the present study indicated that
Smad7 was downregulated in HS tissues and hHSFs compared with
healthy control skin tissues and CCC-ESF-1 cells, respectively. The
results suggested that miR-497-5p might participate in HS formation
by regulating the expression of Smad7. HS is characterized by
abnormal fibroblast proliferation, and collagen and ECM deposition
(4,5). Zhang et al (35) reported that miR-137 inhibits HS
fibroblast proliferation and metastasis via targeting pleiotrophin.
Wu et al (36) suggested
that miR-155 inhibits the formation of HSs by inhibiting HS
fibroblast proliferation and collagen expression by targeting
hypoxia inducible factor-1α via the PI3K/AKT signaling pathway.
Zhou et al (37) indicated
that miR-519d inhibits proliferation and induces apoptosis of human
HS fibroblasts, serving a critical role in HS formation. Moreover,
previous studies have reported that Smad7 serves a key role in the
regulation of fibroblast growth and ECM accumulation (26-28).
Therefore, the present study investigated the effect of Smad7 on
ECM secretion, and hHSF proliferation and apoptosis. The results
indicated that Smad7 upregulation significantly inhibited cell
viability, induced cell apoptosis, and decreased the protein
expression levels of Col I, Col III and α-SMA in hHSFs compared
with the control-plasmid group. miRNAs work by negatively
regulating the expression of target genes (40,41).
Using bioinformatics software, Smad7 was identified as a potential
target gene of miR-497-5p in the present study. Therefore, it was
hypothesized that miR-497-5p may regulate scar fibroblast
proliferation and ECM production by regulating the expression of
Smad7, thereby serving a role in HS formation. As expected, the
results of the present study indicated that miR-497-5p inhibition
decreased cell viability, induced apoptosis, and decreased the
protein expression levels of Col I, Col III and α-SMA in hHSFs
compared with the inhibitor control group. Notably, miR-497-5p
knockdown-mediated effects on hHSFs were eliminated by Smad7
downregulation. However, in the present study, only the CCK-8 assay
was performed to assess the effect of miR-497-5p/Smad7 on hHSF
viability, which was a limitation. Future studies should perform
EdU staining to investigate the effect of miR-497-5p/Smad7 on hHSF
proliferation.
In summary, the present study indicated that
miR-497-5p was upregulated in HS. miR-497-5p downregulation
inhibited HS formation by inhibiting ECM deposition, preventing
hHSF proliferation at least partly by targeting Smad7. Therefore,
the miR-497-5p/Smad7 axis may serve as a promising therapeutic
target for the treatment of HS. However, the present study was only
a preliminary study of the effect of miR-497-5p on hHSFs and
further investigation is required. For example, the effects of
miR-497-5p/Smad7 on hHSF migration and the role of the
miR-497-5p/Smad7 axis in HSs in vivo require further
investigation. Moreover, whether there is a correlation between the
expression of miR-497-5p/Smad7 and the clinicopathological
parameters of patients with HS should be investigated in the
future.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZL designed the study, collected, analyzed and
interpreted the data and prepared the manuscript. PW, JZ and DZ
collected the data and performed statistical analysis. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Shanghai Meizhizhen Medical Cosmetology Clinic.
Written informed consent was obtained from the patients and all
patients approved the use of their samples in the present
study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gabriel V: Hypertrophic scar. Phys Med
Rehabil Clin N Am. 22:301–310. 2011.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Fan SQ, Cai JL, Qin LY, Wang ZH, Liu ZZ
and Sun ML: Effect of heparin on production of transforming growth
factor (TGF)-betal and TGF-betal mRNA expression by human normal
skin and hyperplastic scar fibroblasts. Ann Plast Surg. 60:299–305.
2008.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Gauglitz GG, Korting HC, Pavicic T,
Ruzicka T and Jeschke MG: Hypertrophic scarring and keloids:
Pathomechanisms and current and emerging treatment strategies. Mol
Med. 17:113–125. 2011.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Sidgwick GP, Iqbal SA and Bayat A: Altered
expression of hyaluronan synthase and hyaluronidase mRNA may affect
hyaluronic acid distribution in keloid disease compared with normal
skin. Exp Dermatol. 22:377–379. 2013.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Syed F, Ahmadi E, Iqbal SA, Singh S,
McGrouther DA and Bayat A: Fibroblasts from the growing margin of
keloid scars produce higher levels of collagen I and III compared
with intralesional and extralesional sites: Clinical implications
for lesional site-directed therapy. Br J Dermatol. 164:83–96.
2011.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zuccaro J, Ziolkowski N and Fish J: A
systematic review of the effectiveness of laser therapy for
hypertrophic burn scars. Clin Plast Surg. 44:767–779.
2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Xu X, Lai L, Zhang X, Chen J, Chen J, Wang
F, Zheng J and Chen M: Autologous chyle fat grafting for the
treatment of hypertrophic scars and scar-related conditions. Stem
Cell Res Ther. 9(64)2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Ebert MS and Sharp PA: Roles for microRNAs
in conferring robustness to biological processes. Cell.
149:515–524. 2012.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Rogers K and Chen X: Biogenesis, turnover,
and mode of action of plant microRNAs. Plant Cell. 25:2383–2399.
2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Hayes J, Peruzzi PP and Lawler S:
MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol
Med. 20:460–469. 2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wojtas B, Ferraz C, Stokowy T, Hauptmann
S, Lange D, Dralle H, Musholt T, Jarzab B, Paschke R and Eszlinger
M: Differential miRNA expression defines migration and reduced
apoptosis in follicular thyroid carcinomas. Mol Cell Endocrinol.
388:1–9. 2014.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Mu S, Kang B, Zeng W, Sun Y and Yang F:
MicroRNA-143-3p inhibits hyperplastic scar formation by targeting
connective tissue growth factor CTGF/CCN2 via the Akt/mTOR pathway.
Mol Cell Biochem. 416:99–108. 2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Hao XZ and Fan HM: Identification of
miRNAs as atherosclerosis biomarkers and functional role of miR-126
in atherosclerosis progression through MAPK signalling pathway. Eur
Rev Med Pharmacol Sci. 21:2725–2733. 2017.PubMed/NCBI
|
|
15
|
Bi S, Chai L, Yuan X, Cao C and Li S:
MicroRNA-98 inhibits the cell proliferation of human hypertrophic
scar fibroblasts via targeting Col1A1. Biol Res.
50(22)2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zhu HY, Li C, Bai WD, Su LL, Liu JQ, Li Y,
Shi JH, Cai WX, Bai XZ, Jia YH, et al: MicroRNA-21 regulates hTERT
via PTEN in hypertrophic scar fibroblasts. PLoS One.
9(e97114)2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Qu F, Ye J, Pan X, Wang J, Gan S, Chu C,
Chu J, Zhang X, Liu M, He H and Cui X: MicroRNA-497-5p
down-regulation increases PD-L1 expression in clear cell renal cell
carcinoma. J Drug Target. 27:67–74. 2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Chai L, Kang XJ, Sun ZZ, Zeng MF, Yu SR,
Ding Y, Liang JQ, Li TT and Zhao J: MiR-497-5p, miR-195-5p and
miR-455-3p function as tumor suppressors by targeting hTERT in
melanoma A375 cells. Cancer Manag Res. 10:989–1003. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Sun Z, Li A, Yu Z, Li X, Guo X and Chen R:
MicroRNA-497-5p suppresses tumor cell growth of osteosarcoma by
targeting ADP ribosylation factor-like protein 2. Cancer Biother
Radiopharm. 32:371–378. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Chen Y, Kuang D, Zhao X, Chen D, Wang X,
Yang Q, Wan J, Zhu Y, Wang Y, Zhang S, et al: miR-497-5p inhibits
cell proliferation and invasion by targeting KCa3.1 in
angiosarcoma. Oncotarget. 7:58148–58161. 2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Chen X, Shi C, Wang C, Liu W, Chu Y, Xiang
Z, Hu K, Dong P and Han X: The role of miR-497-5p in myofibroblast
differentiation of LR-MSCs and pulmonary fibrogenesis. Sci Rep.
7(40958)2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Nakao A, Afrakhte M, Morén A, Nakayama T,
Christian JL, Heuchel R, Itoh S, Kawabata M, Heldin NE, Heldin CH
and ten Dijke P: Identification of Smad7, a TGFbeta-inducible
antagonist of TGF-beta signalling. Nature. 389:631–635.
1997.PubMed/NCBI View
Article : Google Scholar
|
|
23
|
Kavsak P, Rasmussen RK, Causing CG, Bonni
S, Zhu H, Thomsen GH and Wrana JL: Smad7 binds to Smurf2 to form an
E3 ubiquitin ligase that targets the TGF beta receptor for
degradation. Mol Cell. 6:1365–1375. 2000.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Yeung ML, Yao Y, Jia L, Chan JFW, Chan KH,
Cheung KF, Chen H, Poon VKM, Tsang AKL, To KKW, et al: MERS
coronavirus induces apoptosis in kidney and lung by upregulating
Smad7 and FGF2. Nat Microbiol. 1(16004)2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Liu J, Kong D, Qiu J, Xie Y, Lu Z, Zhou C,
Liu X, Zhang R and Wang Y: Praziquantel ameliorates
CCl4-induced liver fibrosis in mice by inhibiting
TGF-β/Smad signalling via up-regulating Smad7 in hepatic stellate
cells. Br J Pharmacol. 176:4666–4680. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhou R, Wang C, Wen C and Wang D: miR-21
promotes collagen production in keloid via Smad7. Burns.
43:555–561. 2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Chen Z, Liu Y, Xiao M, Xiao H, Qiu W and
Zhao Y: Effects of over-expressing smad7 gene on keloid
fibroblasts. Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi. 30:871–875.
2016.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
28
|
Tao J, Wang J, Li C, Wang W, Yu H, Liu J,
Kong X and Chen Y: MiR-216a accelerates proliferation and
fibrogenesis via targeting PTEN and SMAD7 in human cardiac
fibroblasts. Cardiovasc Diagn Ther. 9:535–544. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Lee HJ and Jang YJ: Recent understandings
of biology, prophylaxis and treatment strategies for hypertrophic
scars and keloids. Int J Mol Sci. 19(711)2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Aberdam D, Candi E, Knight RK and Melino
G: miRNAs, ‘sternness’ and skin. Trends Biochem Sci. 33:583–591.
2008.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Botchkareva NV: MicroRNA/mRNA regulatory
networks in the control of skin development and regeneration. Cell
Cycle. 11:468–474. 2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Chen L and Li J, Li Q, Yan H, Zhou B, Gao
Y and Li J: Non-coding RNAs: The new insight on hypertrophic scar.
J Cell Biochem. 118:1965–1968. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Tsai CH and Ogawa R: Keloid research:
Current status and future directions. Scars Burn Heal.
5(2059513119868659)2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zhang Q, Guo B, Hui Q, Chang P and Tao K:
miR-137 inhibits proliferation and metastasis of hypertrophic scar
fibroblasts via targeting pleiotrophin. Cell Physiol Biochem.
49:985–995. 2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Wu X, Li J, Yang X, Bai X, Shi J, Gao J,
Li Y, Han S, Zhang Y, Han F, et al: miR-155 inhibits the formation
of hypertrophic scar fibroblasts by targeting HIF-1α via PI3K/AKT
pathway. J Mol Histol. 49:377–387. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zhou X, Xie Y, Xiao H, Deng X, Wang Y,
Jiang L, Liu C and Zhou R: MicroRNA-519d inhibits proliferation and
induces apoptosis of human hypertrophic scar fibroblasts through
targeting Sirtuin 7. Biomed Pharmacother. 100:184–190.
2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Wang X, Zhang Y, Jiang BH, Zhang Q, Zhou
RP, Zhang L and Wang C: Study on the role of Hsa-miR-31-5p in
hypertrophic scar formation and the mechanism. Exp Cell Res.
361:201–209. 2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zunwen L, Shizhen Z, Dewu L, Yungui M and
Pu N: Effect of tetrandrine on the TGF-β-induced smad signal
transduction pathway in human hypertrophic scar fibroblasts in
vitro. Burns. 38:404–413. 2012.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Liu B, Li J and Cairns MJ: Identifying
miRNAs, targets and functions. Brief Bioinform. 15:1–19.
2014.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Lu TX and Rothenberg ME: MicroRNA. J
Allergy Clin Immunol. 141:1202–1207. 2018.PubMed/NCBI View Article : Google Scholar
|