Introduction
Diabetic retinopathy (DR) is an ophthalmic
complication caused by diabetes mellitus that may lead to severe
damage of vision and even blindness. In industrialized countries,
DR has become the leading cause of blindness in the working
population (1,2). In particular, when DR develops to the
proliferative diabetic retinopathy (PDR) stage, it may cause severe
complications, such as retinal neovascularization (RNV), vitreous
hemorrhage, traction retinal detachment and neovascularization
glaucoma (3), leading to severe
vision impairment. RNV is one of the major pathological changes of
PDR and its formation is a complex process, including the
regulation of inflammatory factors, oxidative damage and various
cytokines (4-7).
Research has indicated that adipose tissue is not only a simple
energy storage organ but also an endocrine organ. Adipose tissue
synthesizes and releases various adipocytokines, such as leptin and
resistin, which are widely involved in numerous pathophysiological
states, such as obesity, insulin resistance and cardiovascular
disease (8).
Visfatin is a novel adipocytokine that is
synthesized and released primarily from visceral fat (9). Visfatin is able to bind to and
activate the insulin receptor on specific loci to simulate an
insulin-like effect to decrease blood sugar levels. It also
participates in the inflammatory response, regulates lipid
metabolism, promotes differentiation, participates in the formation
of atherosclerosis and adjusts the maturity of vascular smooth
muscle. A previous study has indicated that visfatin levels are
elevated in serum and vitreous fluids in patients with PDR and that
the plasma levels of visfatin are associated with vascular
endothelial function in patients with diabetes (10). Shen et al (11) studied the effect of various doses of
glucose on the proliferation of RF/6A cells in a time-course study
performed for 24-72 h. The results from the MTT assay suggested
that glucose promoted cell proliferation at all concentrations
tested (10-50 mM) without any effect on their viability and at all
time-periods tested (24-72 h). Another study indicated that the
expression level of visfatin was increased in retinal tissues of
diabetic rats (12). These results
suggest that visfatin may be involved in the development of DR.
However, whether visfatin promotes the formation of RNV in diabetic
patients and its underlying mechanisms have remained elusive. Thus,
in the present study, the effect of visfatin on angiogenesis in
RF/6A monkey chorioretinal retinal endothelial cells under
high-glucose (HG) conditions was investigated in vitro.
Materials and methods
Materials
RF/6A cells were purchased from The Cell Bank of the
Type Culture Collection of the Chinese Academy of Sciences. Human
recombinant visfatin was purchased from Peprotech, Inc. The
anti-VEGF (cat. no. BS91432) antibody and antibody-HRP (cat. no.
BS6007MH) was obtained from Bioworld Technology, Inc. The
anti-VEGFR-2 (cat. no. SC-393163) antibody, VEGFR2 inhibitor
(SU1498) and GAPDH (cat. no. SC-166574) were purchased from Santa
Cruz Biotechnology, Inc. RPMI-1640 was from Gibco (Thermo Fisher
Scientific, Inc.). The other cell culture reagents were obtained
from Beyotime Institute of Biotechnology.
Cell culture and treatment
RF/6A cells were incubated in RPMI-1640 medium
containing 10% fetal bovine serum and 1% penicillin-streptomycin
solution at 37˚C with 5% CO2 (all from Beyotime
Institute of Biotechnology). The culture medium was replaced 24 h
after seeding and different concentrations of visfatin and/or
D-glucose were added to the medium. RF/6A cells were divided into 5
groups: Control group, HG group, visfatin group 1 (10 nM visfatin +
25 mM D-glucose), visfatin group 2 (20 nM visfatin + 25 mM
D-glucose), visfatin group 3 (30 nM visfatin + 25 mM D-glucose) and
the visfatin group 4 (HG + SU1498).
Cell counting kit-8 (CCK-8) cell
proliferation assay
RF/6A cells were inoculated into 96-well plates at
5x103 cells/well in 100 µl culture medium. Following
incubation overnight in a 5% CO2 incubator at 37˚C, the
cells were treated according to their assigned group for 24 h.
Subsequently, 10 µl CCK-8 solution was added to each well, followed
by incubation for 4 h at 37˚C. This was repeated three times
independently of each other. An ELISA plate reader at the
wavelength of 450 nM was used to determine the absorbance and the
optical density was recorded in order to calculate the cell
proliferation rate.
Cell migration determined by cell
scratch assay
A marker was used to draw three evenly spaced
straight lines behind the 6-well plate as positioning lines. RF/6A
cells were inoculated in 6-well plates in at 5x105 cells
per well. After 24 h of culture in vitro in a dish, a
monolayer had formed and a 100-µl pipette tip was used to scratch
the cells, perpendicular to the positioning line. The intersection
point of the scratch and the positioning line were taken as the
monitoring point. PBS was used to gently rinse the bottom of the
culture well three times to remove any detached cells. Medium
containing 10% fetal bovine serum (13,14)
and different concentrations of visfatin and D-glucose were added
according to the different group assignments. Under an inverted
microscope, an image of the scratch width at the monitoring point
was captured, which was considered as the 0-h time point. The cell
culture was continued for 24 and 48 h and the width of the
scratches at the monitoring points was recorded. ImageJ software
version 1.7.0 [National Institutes of Health (NIH)] was used to
calculate the width of the scratches. A total of five fields of
view were selected for observation in each group. The independent
experiments were repeated three times.
Cell tube formation detected by
Matrigel assay
Matrigel was dissolved according to the
manufacturer's instructions. Subsequently, 100 µl Matrigel was
slowly added to each well of a 96-well plate. RF/6A cells were
digested and diluted to 2x105/ml with cell culture
medium containing 10% fetal bovine serum. Cell suspension (50 µl)
was added to each well and 50 µl visfatin and D-glucose were added
according to the different group assignments. After incubation for
24 h, the cells were observed under a phase-contrast microscope,
and images of 5 randomly selected fields were captured at a
magnification of x100. ImageJ software version 1.7.0 was used to
calculate the number of complete tubes formed. Each group was set
up in triplicate wells. The independent experiments were repeated
three times.
Expression of VEGF and VEGFR-2
detected by western blot analysis
RF/6A cells in each group were lysed using RIPA
buffer (Beyotime Institute of Biotechnology). Nucleus proteins of
RF/6A cells of each group were also obtained through using nucleus
protein extraction kit (Beyotime Institute of Biotechnology). Cell
lysate (30 µg protein per lane) was then separated by 12% SDS-PAGE.
The separated proteins were transferred onto nitrocellulose
membranes (Beyotime Institute of Biotechnology), which were then
blocked with Tris-buffered saline containing Tween-20 with 5%
non-fat milk. The membrane was incubated overnight with primary
antibodies to GAPDH (1:1,000), VEGF (1:1,000) and VEGFR-2
(1:1,000), and then incubated with a horseradish
peroxidase-conjugated secondary antibody (Beyotime Institute of
Biotechnology; cat. no. P0239; 1:1,000) for 1 h at room
temperature. The labeled bands were visualized and quantified using
a chemiluminescence imaging system (Tanon 5000; CliNX). CliNX
analysis software (version 1.7.0) was used to scan the gels and
determine the gray value. The ratio of the target protein gray
value to that of β-actin represented the relative expression levels
of the target protein.
mRNA expression levels of VEGF and
VEGFR-2 detected by reverse transcription-quantitative
(RT-q)PCR
After total RNA was extracted with the TRIzol
method, the RT reaction was performed according to the
manufacturer's protoco. The complementary (c)DNA obtained was
amplified on the fluorescence quantitative real-time PCR System
(7300 Real-Time PCR System; Applied Biosystems; Thermo Fisher
Scientific, Inc.). Using 1 µl cDNA as a template and GAPDH as the
internal reference, the Power SYBR-Green PCR MasterMix kit
(Beyotime Institute of Biotechnology) was used for PCR
amplification of gene fragments according to the manufacturer's
protocol. Primer sequences are listed in Table I. The PCR reaction parameters were
as follows: Pre-denaturation at 95˚C for 10 min, followed by 40
cycles of denaturation for 15 sec at 95˚C and 60˚C for 1 min for
annealing and extension, during which fluorescence signals were
collected. The data were analyzed with 7300 System SDS software and
the statistical 2-ΔΔCq method was used to compare the
mRNA expression in each group (15).
 | Table IPrimer sequences. |
Table I
Primer sequences.
| mRNA/primer
direction | Sequence |
|---|
| VEGF mRNA |
|
F |
5'-GAGCCTTGCCTTGCTGCTCTAG-3' |
|
R |
5'-CACCAGGGTCTCGATTGGATG-3' |
| VEGFR-2 mRNA |
|
F |
5'-GCGTGATTCTGAGGAAAGG-3' |
|
R |
5'-ATAAACAGTGGAGGCTATGTCG-3' |
| GAPDH |
|
F |
5'-CGACAGTCAGCCGCATCTT-3' |
|
R |
5'-TCACCTTCCCCATGGTGTCT-3' |
Statistical analysis
SPSS 22.0 software (IBM Corp.) was used for
statistical analysis. Values are expressed as the mean ± standard
deviation. The results obtained for all the tests were submitted to
a normality test and groups were compared using parametric ANOVA
and Tukey's multiple-comparisons test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Effect of visfatin on RF/6A cell
proliferation
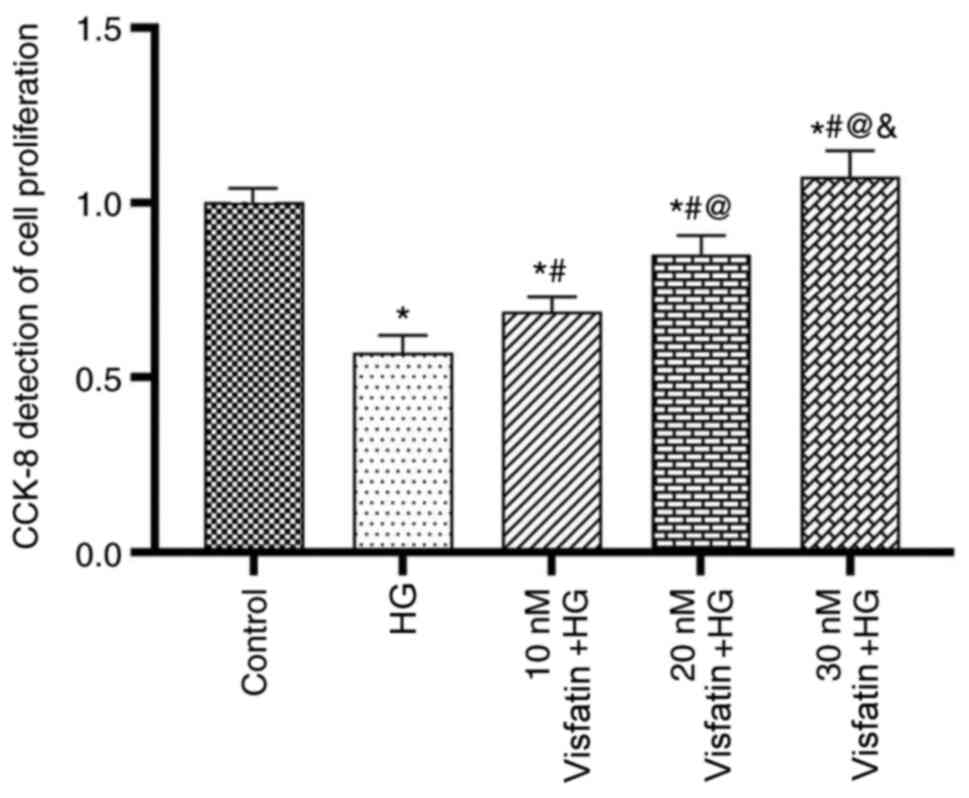
The results of the CCK-8 assay indicated that
various concentrations of visfatin influenced the proliferation of
RF/6A cells (Fig. 1). At 24 and 48
h, the number of viable cells decreased in the HG group compared
with that in the control group (P<0.05). Compared with that in
the HG group, the cell proliferation in the three visfatin groups
was significantly increased in a dose-dependent manner (F=47.13;
P<0.05), with the most obvious increase in visfatin group 3.
Effect of visfatin on RF/6A cell
migration
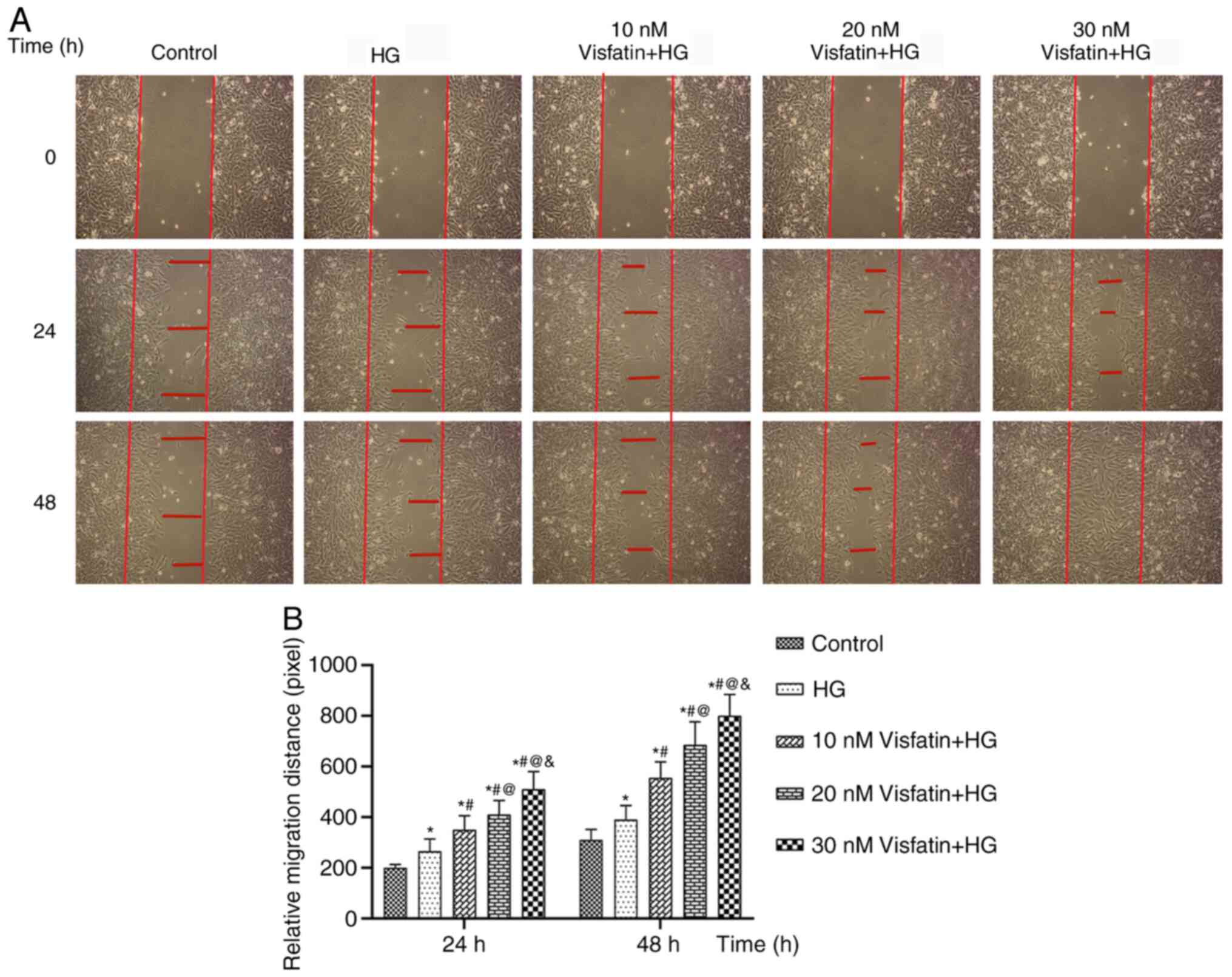
In a wound-healing assay, RF/6A cells were treated
with visfatin at concentrations of 10, 20 and 30 nM under HG
conditions and images of cell migration were captured and analyzed
(Fig. 2). At 24 and 48 h, the cell
migration distance in the HG group was increased compared with that
in the control group (P<0.05). Compared with that in the HG
group, the migration distance in the three visfatin groups
increased significantly in a dose-dependent manner (F=571.67;
P<0.05) and the migration distance in the visfatin group 3 was
the largest. The results indicated that visfatin significantly
promoted the migration of RF/6A cells.
Effect of visfatin on tube formation
of RF/6A cells
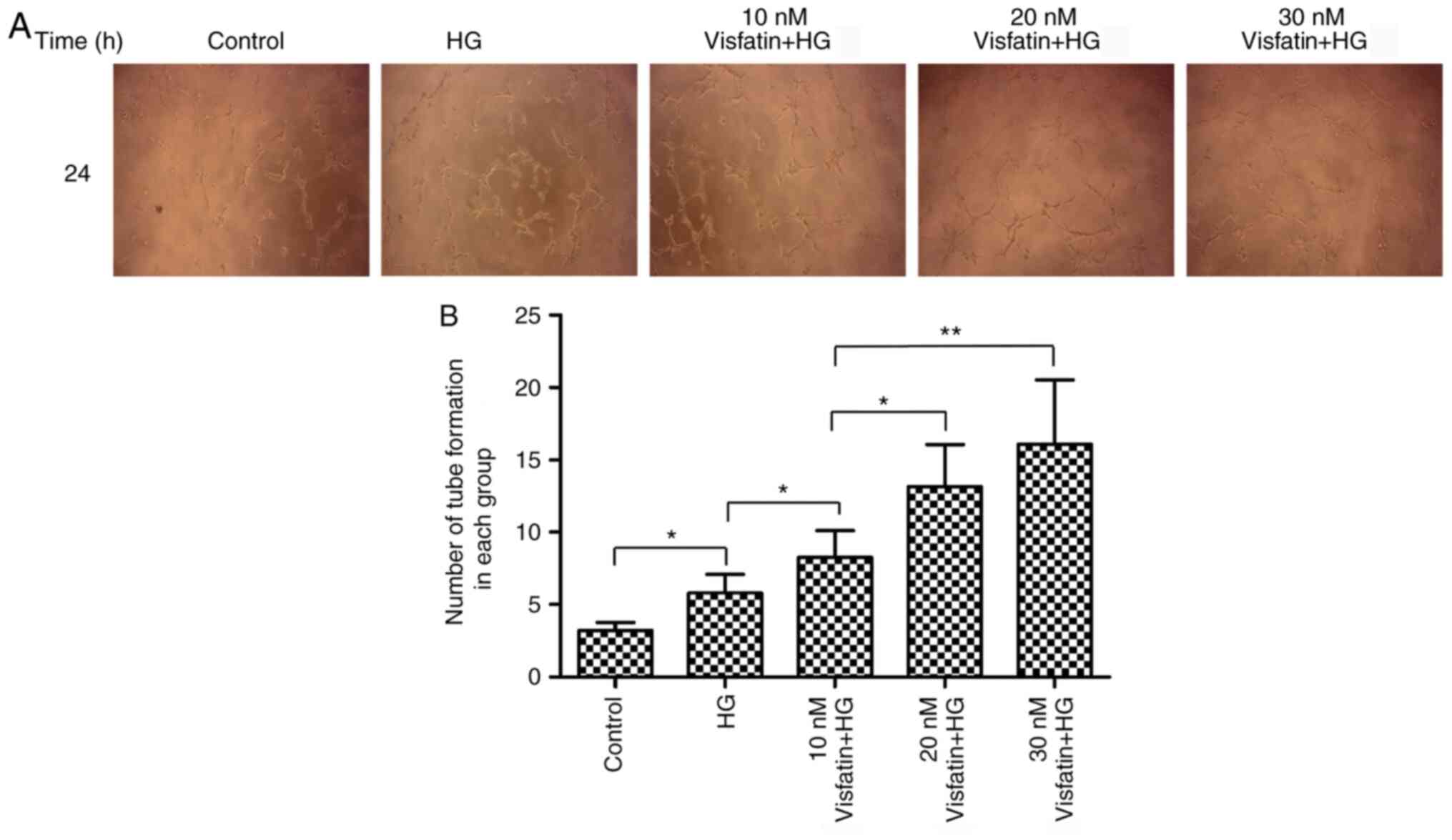
On Matrigel, RF/6A cells were treated with visfatin
at the three aforementioned different concentrations and the number
of tube-like structures formed by RF/6A cells was analyzed after 24
h of cell culture (Fig. 3). The
results suggested that, compared with that in the control group,
the number of tubes formed by RF/6A cells was increased in the HG
group (P<0.05). Compared with that in the HG group, the number
of tubes formed in the three visfatin groups was significantly
increased in a dose-dependent manner (F=253.78, P<0.05) and the
amount of tube formation in the visfatin group 3 was highest. These
results suggested that visfatin was able to significantly promote
the formation of tube-like structures by RF/6A cells, especially at
the concentration of 30 nM.
Expression of VEGF and VEGFR-2 in
RF/6A cells
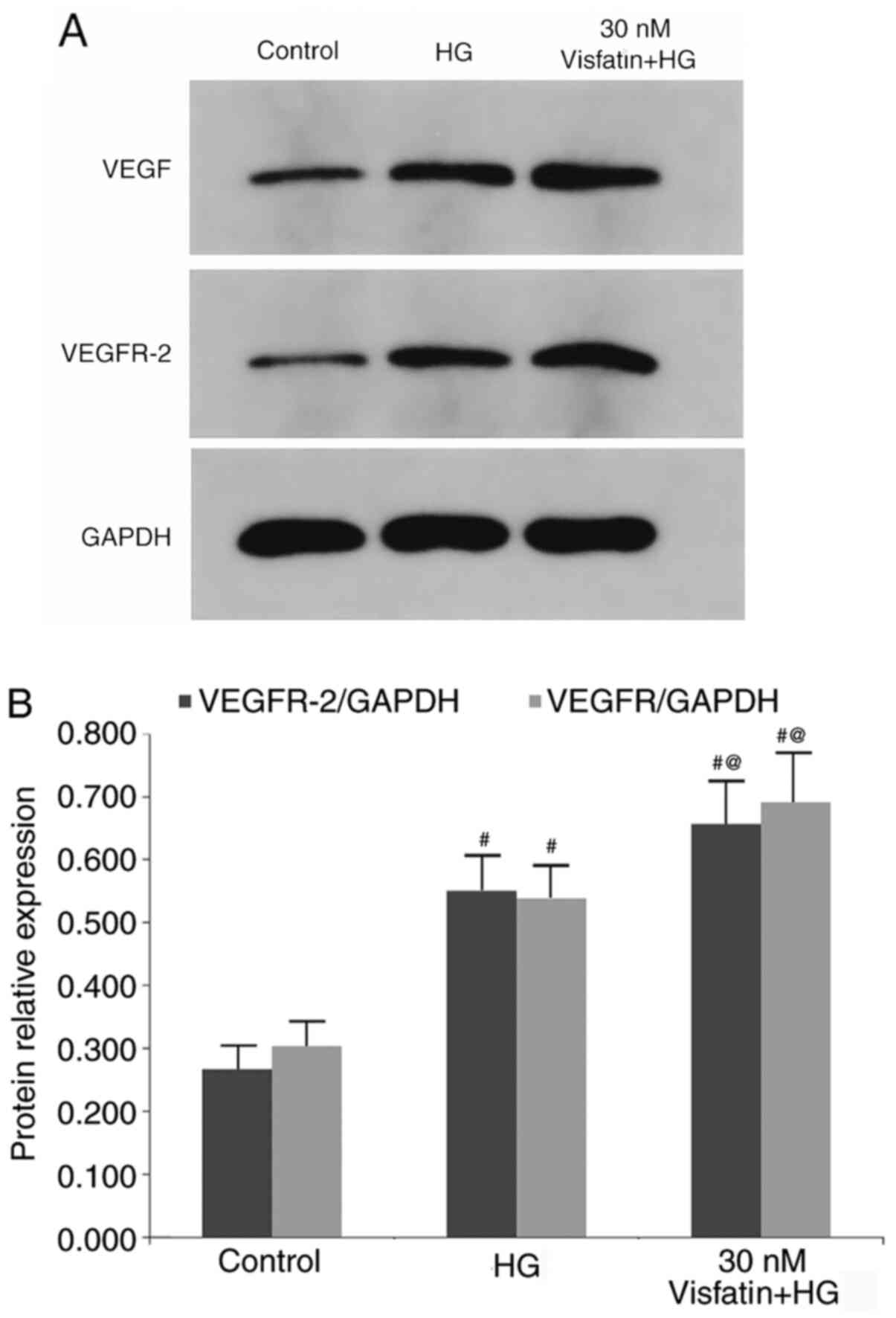
The protein expression levels of VEGF and VEGFR-2
were detected by western blot analysis (Fig. 4). The results suggested that the
protein expression levels of VEGF and VEGFR-2 were significantly
increased in the HG group as compared with those in the control
group (P<0.05). In comparison with those in the HG group, the
protein expression levels of VEGF and VEGFR-2 were increased in the
30 nM visfatin + HG group (P<0.05).
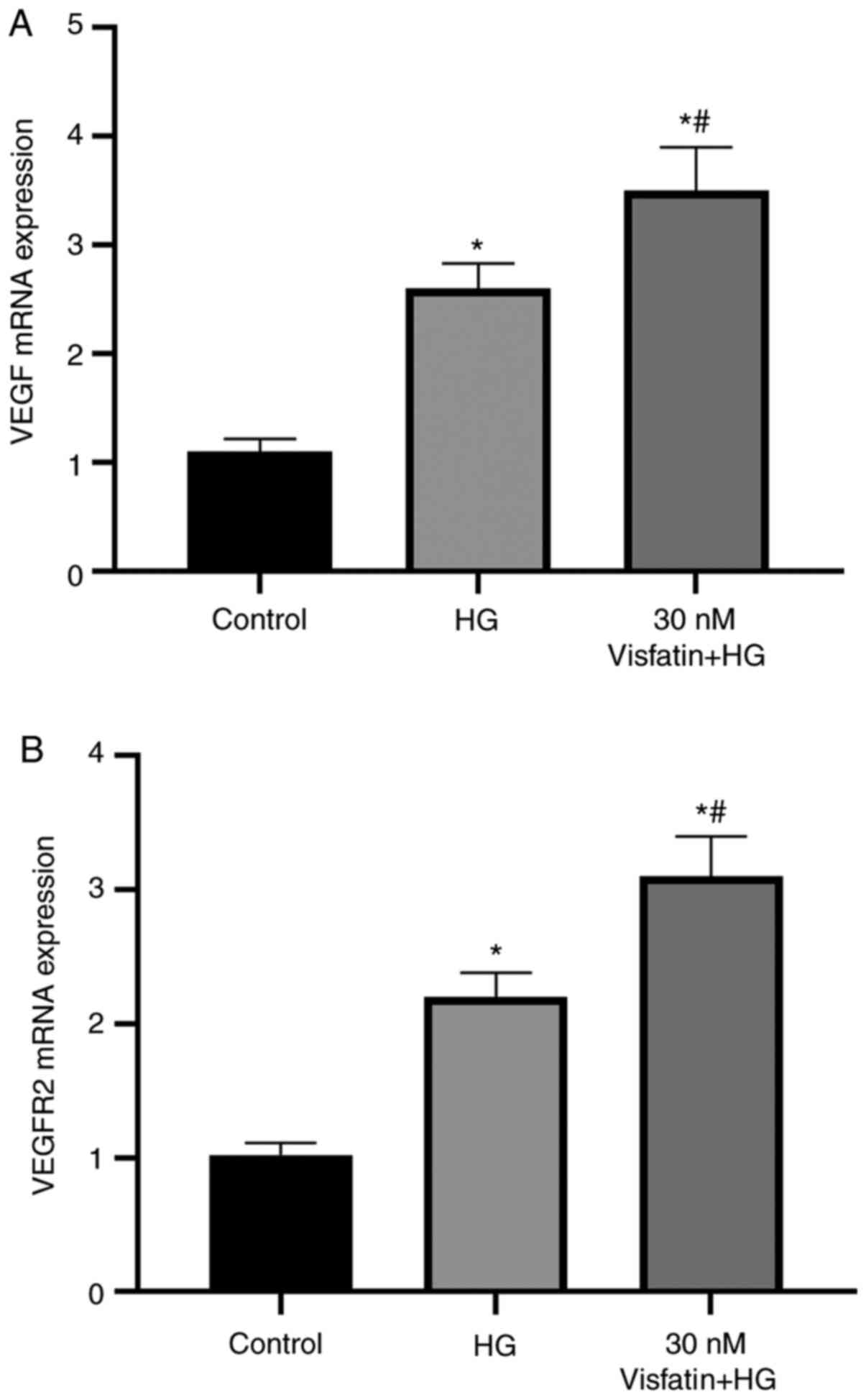
RT-qPCR was used to detect the mRNA expression
levels of VEGF and VEGFR-2 in the cells (Fig. 5). The results suggested that the
mRNA expression levels of VEGF and VEGFR-2 were significantly
increased in the HG group as compared with those in the control
group (P<0.05). Furthermore, as compared with those in the HG
group, the mRNA expression levels of VEGF and VEGFR-2 were
significantly increased in the 30 nM visfatin + HG group
(P<0.05).
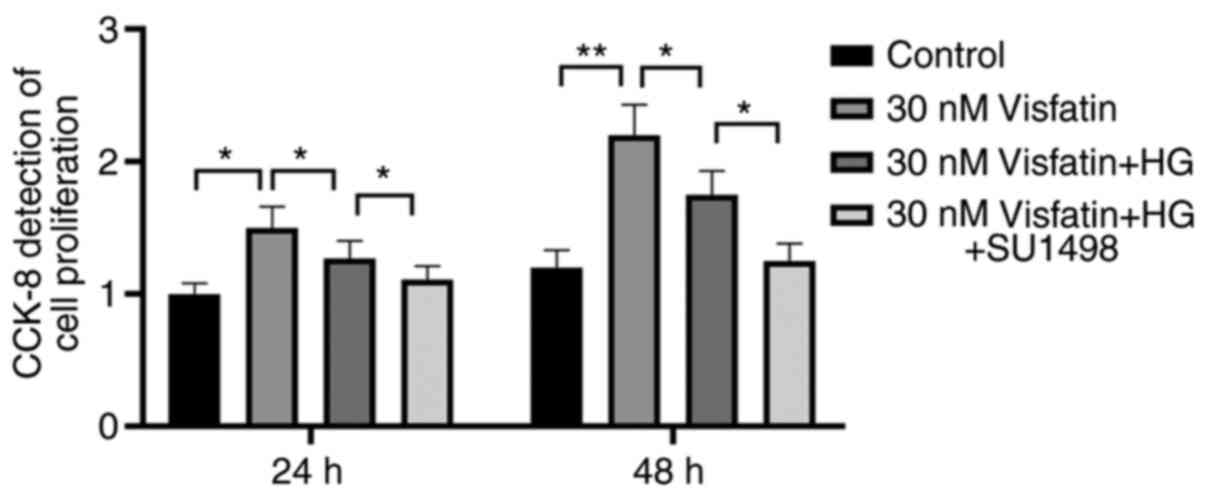
Functional experiments
The results of the CCK8 assay suggested that
visfatin at different concentrations influenced the proliferation
of RF/6A cells. At 24 and 48 h, cell proliferation in the control
group was lower compared with that in the 30 nM visfatin group +
HG. Compared with that in the HG + SU1498 (VEGF block) group, the
cell proliferation in the 30 nM visfatin + HG group was
significantly increased (P<0.05). Furthermore, compared with
that in the 30 nM visfatin + HG group, the cell proliferation in
the 30 nM visfatin group was significantly increased (F=15.14,
P<0.05; Fig. 6).
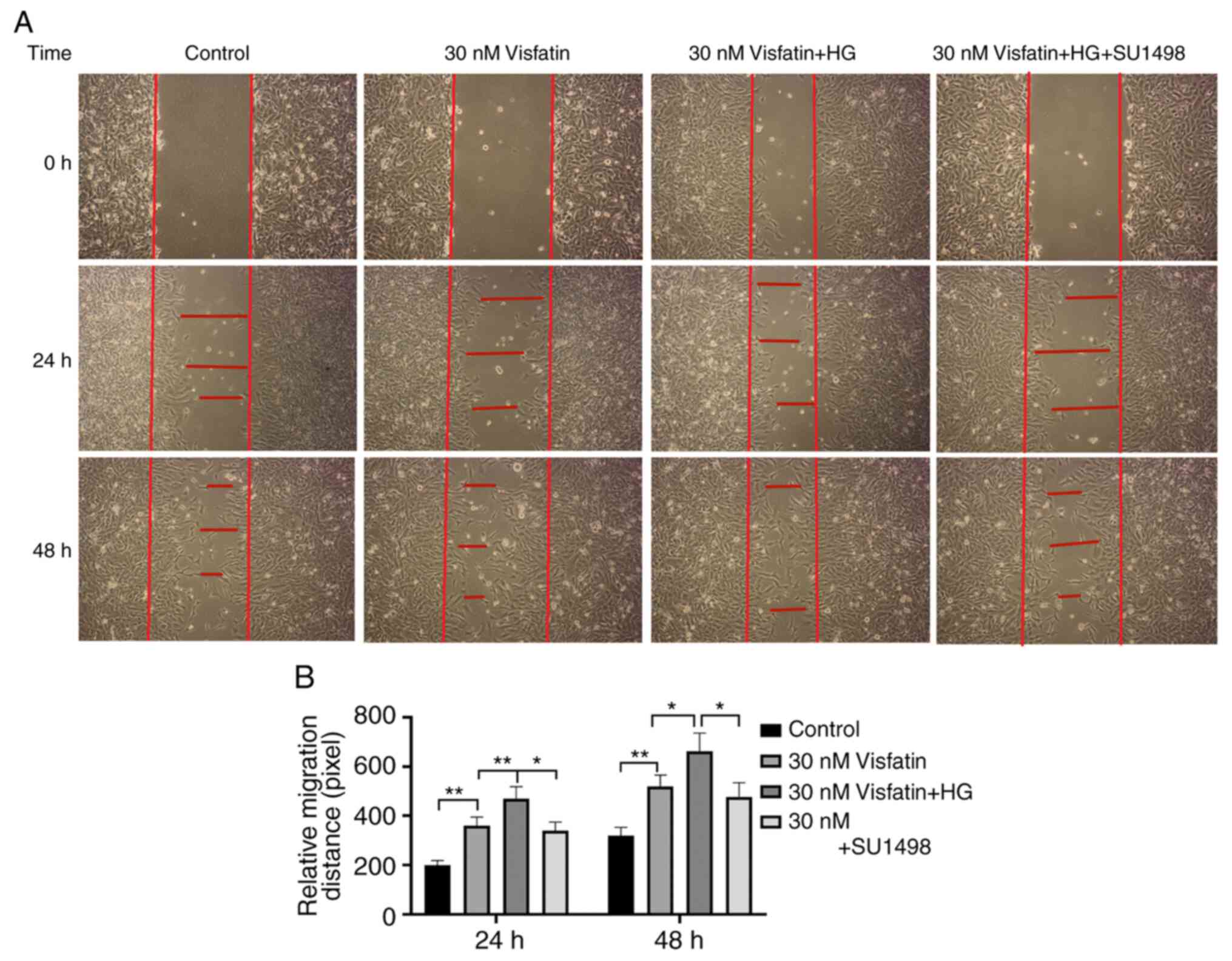
The cell migration distance in the visfatin groups
(30 nM visfatin and 30 nM visfatin + HG) at 24 and 48 h was
increased compared with that in the control group (P<0.05) and
the migration distance in the 30 nM visfatin + HG group was the
largest (P<0.05). Compared with that in the visfatin + HG +
SU1498 group, the migration distance in the 30 nM visfatin + HG
group was significantly greater (F=177.89, P<0.05; Fig. 7).
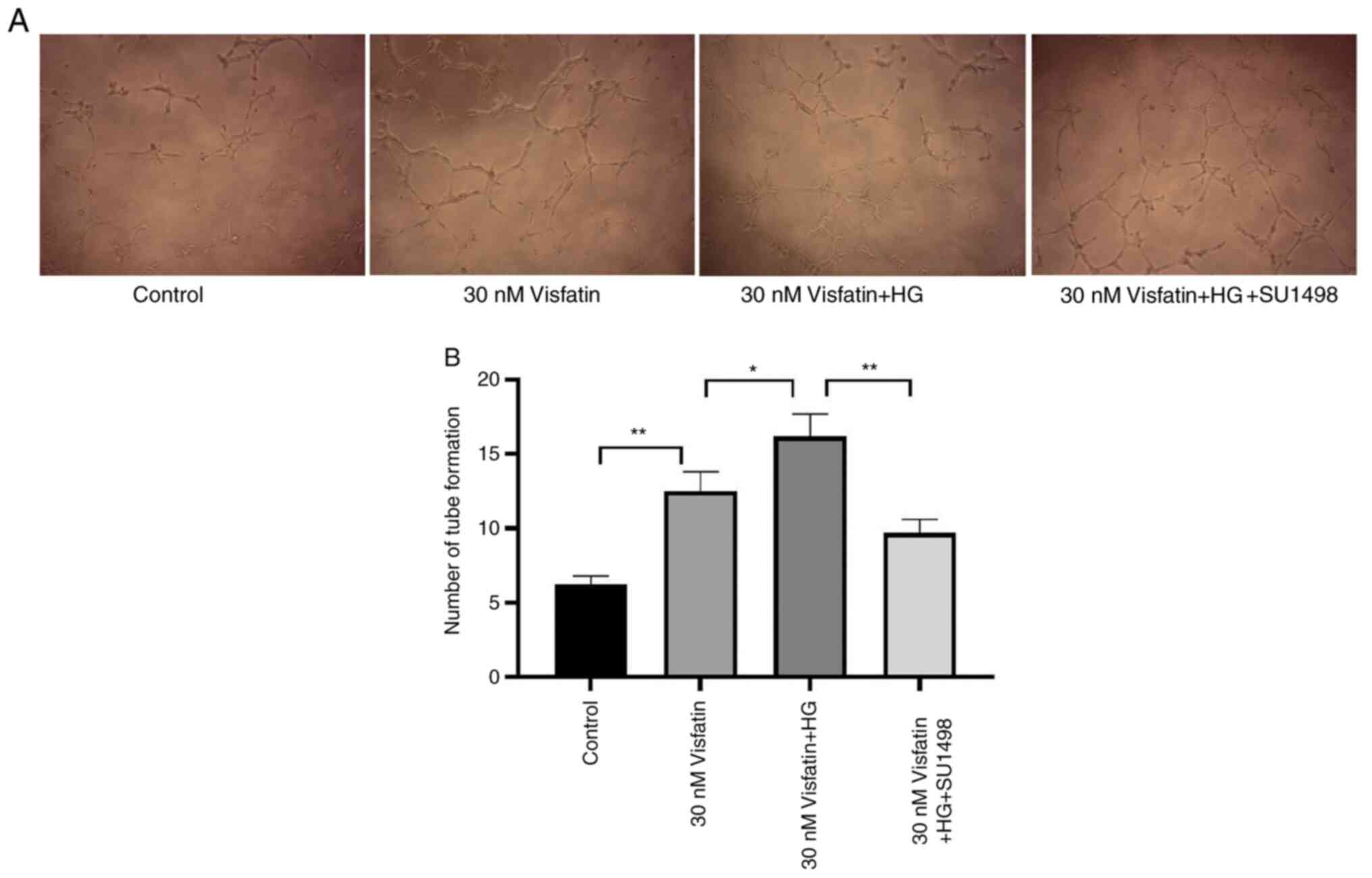
The number of tubes formed by RF/6A cells in the 30
nM visfatin group was increased compared with that in the control
group (P<0.05). Compared with that in the 30 nM visfatin + HG
group, the number of tubes formed in the 30 nM visfatin and
visfatin + HG + SU1498 groups was significantly decreased (F=13.50,
P<0.05; Fig. 8).
Discussion
DR is one of the most common and serious
microvascular complications in patients with diabetes and seriously
affects their quality of life (16,17).
While its pathogenesis remains to be fully elucidated, it is widely
accepted that diabetes leads to damage to retinal capillaries,
tissue edema, bleeding and capillary blockage, resulting in retinal
ischemia and hypoxia, further leading to microaneurysms and other
pathological changes, including the formation of new blood vessels
(18). It has been indicated that
cytokines secreted by adipose tissue may participate in the
pathological changes in the eyes of patients with diabetes, such as
the formation of new blood vessels and cell proliferation, which
are closely associated with the onset of DR (19). The results of the present study
suggested that visfatin is able to significantly promote the
proliferation and migration of RF/6A cells under HG conditions.
Among the concentrations tested, visfatin at 30 nM had the most
obvious effect. Quantitative analysis of RF/6A cell tube-structure
formation in a Matrigel assay indicated that after 24 h of culture,
visfatin significantly promoted the tube-structure formation of
RF/6A cells, particularly at the concentration of 30 nM. Visfatin
and angiotensin II have been reported to influence each other:
Angiotensin II is able to promote retinal angiogenesis by
upregulating VEGF and higher angiotensin II levels increase insulin
resistance, which in turn increases the production of visfatin
(20). Similar to VEGF, visfatin
may promote the proliferation of retinal capillary endothelial
cells and directly induce the occurrence of DR. Visfatin is able to
interact with inflammatory factors and oxidative stress. Serum
inflammatory factors are able to upregulate the expression and
secretion of visfatin. However, increases in visfatin were
determined to further influence the protein kinase C signal
transduction pathway, promote the synthesis of reactive oxygen
species, aggravate oxidative stress and promote the occurrence of
DR (21).
Chen (9) reported
that the level of visfatin increased in the plasma of patients with
DR; however, the precise mechanism of the effect of visfatin on
angiogenesis in DR had remained to be fully elucidated. In the
present study, RF/6A cells were treated with different
concentrations of visfatin to observe its effect on VEGF and its
receptor VEGFR-2. The results indicated that the protein and mRNA
expression levels of VEGF and VEGFR-2 were significantly increased
in the HG group as compared with those in the control group.
Different concentrations of visfatin were able to significantly
increase the expression of VEGF and VEGFR-2 in RF/6A cells cultured
with HG. VEGF is a classical vascular growth factor, which may be
produced by a variety of tumor cell and certain normal cell types.
The main function of VEGF is to promote endothelial cell
proliferation and induce the formation of new blood vessels
(22,23). There are many factors promoting
angiogenesis, such as VEGF, fibroblast growth factor, angiogenic
nutrients, IL-1, IL-8, as well as some small molecules of lipids,
nucleotides and vitamins, among which VEGF plays an important role
in the occurrence of blood vessels (24-26).
Under normal conditions, the expression level of VEGF is relatively
low, while under pathological conditions such as hypoxia, both the
gene expression and protein levels of VEGF are upregulated
(27). VEGF is able to bind to its
receptor VEGFR-2, which promotes endothelial cell proliferation and
angiogenesis.
These results suggested that high levels of visfatin
may help promote the process of RNV. Previous studies have
indicated that visfatin is able to upregulate the mRNA expression
and release of VEGF in endothelial cells (28). As a key regulator of angiogenesis,
VEGF is able to significantly promote endothelial cell division,
cell proliferation, migration and tube formation, which may be the
mechanism of visfatin promoting neovascularization. The results of
the present study suggested that visfatin may be an endogenous
angiogenic molecule that promotes RNV. The effective concentration
of visfatin used in the present study was similar to that reported
in two previous studies (29,30).
In addition, the current study indicated that the migration
distance in the 30 nM visfatin + HG group was significantly greater
compared with that in the 30 nM visfatin + HG + SU1498 group.
SU1498 is an inhibitor of VEGF. Thus, in an HG environment,
visfatin plays an important role in the angiogenesis process. FBS
was used for the cell-scratch experiment, which contains components
that promote cell proliferation and maintenance, this represents a
limitation of the current study.
In conclusion, the present results suggested that
visfatin is able to promote the proliferation, migration and tube
formation of RF/6A cells under HG conditions, suggesting that
visfatin has an important role in RNV and its mechanism may be
related to the promotion of VEGF and VEGFR-2 expression under HG
conditions.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by the Foundation of Xi'an
Health Committee (grant no. 2020ms07), the Fundamental Research
Funds for the Central Universities (grant no.1191329116), the
Natural Science Foundation of Shaanxi province (grant
no.2020JM-685), the Foundation of Xi'an Science and Technology
Project (grant no.2019114613YX001SF0414).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DC: Study concepts and design, literature research,
manuscript preparation, manuscript editing, statistical analysis;
YW: Study concepts and design, literature research; ML:
Contributions to analysis and interpretation of data, drafting the
manuscript; JC: Participate in experimental design and literature
research; ZL: Data analysis; YS: Manuscript preparation; JD,
Guarantor of integrity of the entire study, study concepts and
design. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Klein R, Klein BE and Moss SE: The
wisconsin epidemiologic study of diabetic retinopathy: An update.
Arch Ophthalmol. 113:702–703. 2012.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Vilsbøll T, Bain SC, Leiter LA, Lingvay I,
Matthews D, Simó R, Helmark IC, Wijayasinghe N and Larsen M:
Semaglutide, reduction in glycated haemoglobin and the risk of
diabetic retinopathy. Diabetes Obes Metab. 20:889–897.
2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bressler SB, Beaulieu WT, Glassman AR,
Gross JG, Jampol LM, Melia M, Peters MA and Rauser ME: Diabetic
Retinopathy Clinical Research Network: Factors associated with
worsening proliferative diabetic retinopathy in eyes treated with
panretinal photocoagulation or ranibizumab. Ophthalmology.
124:431–439. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Kwon JW, Jee D and La TY: Neovascular
glaucoma after vitrectomy in patients with proliferative diabetic
retinopathy. Medicine (Baltimore). 96(e6263)2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
de Barros Garcia JMB, Isaac DLC and Avila
M: Diabetic retinopathy and OCT angiography: Clinical findings and
future perspectives. Int J Retina Vitreous. 3(14)2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zhu L, Xu J, Liu Y, Gong T, Liu J, Huang
Q, Fischbach S, Zou W and Xiao X: Prion protein is essential for
diabetic retinopathy-associated neovascularization. Angiogenesis.
21:767–775. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Lai X, Lili LV and Huang Z: Computerized
diabetic retinopathy diagnosis with optimized random forest. J Med
Imaging Health Inf. 9:274–283. 2019. View Article : Google Scholar
|
|
8
|
Reilly SM and Saltiel AR: Adapting to
obesity with adipose tissue inflammation. Nat Rev Endocrinol.
13:633–643. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Chen YS: Clinical investigation about the
relationship between visfatin, SAA and type 2 diabetic retinopathy.
Int J Ophthalmol. 12:39–42. 2012. View Article : Google Scholar
|
|
10
|
Mayer-Davis EJ, Davis C, Saadine J,
D'Agostino RB Jr, Dabelea D, Dolan L, Garg S, Lawrence JM, Pihoker
C, Rodriguez BL, et al: Diabetic retinopathy in the SEARCH for
diabetes in youth cohort: A pilot study. Diabet Med. 29:1148–1152.
2012.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Shen J, Shen S, Das UN and Xu G: Effect of
essential fatty acids on glucose-induced cytotoxicity to retinal
vascular endothelial cells. Lipids Health Dis. 11(90)2012.
View Article : Google Scholar
|
|
12
|
Qu S, Mo L, Niu Y, Sun X, Li H, Wang Z, Xu
W and Rong A: Expression of visfatin in the diabetic rat retina.
Clin Exp Ophthalmol. 44:251–259. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zheng P, Yin Z, Wu Y, Xu Y, Luo Y and
Zhang TC: LncRNA HOTAIR promotes cell migration and invasion by
regulating MKL1 via inhibition miR206 expression in HeLa cells.
Cell Commun Signal. 16(5)2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Rong LQ, Mei LJ, Yong C, Xie XQ, Xiong XX,
Qiu XY, Pan F, Liu D, Yu SB and Chen XQ: Piperlongumine inhibits
migration of glioblastoma cells via activation of ROS-dependent p38
and JNK signaling pathways. Oxid Med Cell Longev.
2014(653732)2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Joseph EW, Pratilas CA, Poulikakos PI,
Tadi M, Wang W, Taylor BS, Halilovic E, Persaud Y, Xing F, Viale A,
et al: The RAF inhibitor PLX4032 inhibits ERK signaling and tumor
cell proliferation in a V600E BRAF-selective manner. Proc Natl Acad
Sci USA. 107:14903–14908. 2010.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Uslu S, Kebapçi N, Kara M and Bal C:
Relationship between adipocytokines and cardiovascular risk factors
in patients with type 2 diabetes mellitus. ExpTher Med. 4:113–120.
2012.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Scanlon PH: Diabetic retinopathy.
Medicine. 38:656–660. 2010.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Azeze TK, Sisay MM and Zeleke EG:
Incidence of diabetes retinopathy and determinants of time to
diabetes retinopathy among diabetes patients at tikur anbessa
hospital, ethiopia: A retrospective follow up study. BMC Res Notes.
11(542)2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Maimaiti NG and Tuhuti A: Changes of serum
inflammatory factors, adipokines and oxidative stress in patients
with diabetic retinopathy. J Hainan Med Univ. 23:2048–2051. 2017.
View Article : Google Scholar
|
|
20
|
Ludwig PE, Freeman SC and Janot AC: Novel
stem cell and gene therapy in diabetic retinopathy, age related
macular degeneration, and retinitis pigmentosa. Int J Retina
Vitreous. 5(7)2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Ahmed MB, Ismail MI and Meki AR: Relation
of osteoprotegerin, visfatin and ghrelin to metabolic syndrome in
type 2 diabetic patients. Int J Health Sci (Qassim). 9:127–139.
2015.PubMed/NCBI
|
|
22
|
Yoshida S, Murata M, Noda K, Matsuda T,
Saito M, Saito W, Kanda A and Ishida S: Proteolytic cleavage of
vascular adhesion protein-1 induced by vascular endothelial growth
factor in retinal capillary endothelial cells. Jpn J Ophthalmol.
62:256–264. 2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Kim BS, Yang SS, You HK, Shin HI and Lee
J: Fucoidan-induced osteogenic differentiation promotes
angiogenesis by inducing vascular endothelial growth factor
secretion and accelerates bone repair. J Tissue Eng Regen Med.
12(e1311-e1324)2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Bhattacharya R, Fan F, Wang R, Ye X, Xia
L, Boulbes D and Ellis LM: Intracrine VEGF signalling mediates
colorectal cancer cell migration and invasion. Br J Cancer.
117:848–855. 2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Tzeng HE, Chen PC, Lin KW, Lin CY, Tsai
CH, Han SM, Teng CL, Hwang WL, Wang SW and Tang CH: Basic
fibroblast growth factor induces VEGF expression in chondrosarcoma
cells and subsequently promotes endothelial progenitor cell-primed
angiogenesis. Clin Sci (Lond). 129:147–158. 2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Fahey E and Doyle SL: IL-1 family cytokine
regulation of vascular permeability and angiogenesis. Front
Immunol. 10(1426)2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Kurihara T: Roles of hypoxia response in
retinal development and pathophysiology. Keio J Med. 67:1–9.
2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Xiao J, Xiao ZJ, Liu ZG, Gong HY, Yuan Q,
Wang S, Li YJ and Jiang DJ: Involvement of dimethylarginine
dimethylaminohydrolase-2 in visfatin-enhanced angiogenic function
of endothelial cells. Diabetes Metab Res Rev. 25:242–249.
2009.PubMed/NCBI View
Article : Google Scholar
|
|
29
|
Adya R, Tan BK Punn A, Chen J and Randeva
HS: Visfatin induces human endothelial VEGF and MMP-2/9 production
via MAPK and PI3K/Akt signalling pathways: Novel insights into
visfatin-induced angiogenesis. Cardiovasc Res. 78:356–365.
2008.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Kim SR, Bae SK, Choi KS, Park SY, Jun HO,
Lee JY, Jang HO, Yun I, Yoon KH, Kim YJ, et al: Visfatin promotes
angiogenesis by activation of extracellular signal-regulated kinase
1/2. Biochem Biophys Res Commun. 357:150–156. 2007.PubMed/NCBI View Article : Google Scholar
|