Introduction
Debilitating eye diseases, such as age-related
macular degeneration, retinal detachment (RD) and rhegmatogenous RD
associated with choroidal detachment (RRDCD), represent a series of
visual health issues that impact the quality of life of affected
patients (1-3).
While the etiology and pathogenesis of these disorders remain to be
fully elucidated, retinal pigment epithelium (RPE) loss and
dysfunction appear to be significant contributors (4-6).
Understanding the underlying mechanisms of phenotypic alterations,
including cell viability and inflammation in RPE, should greatly
assist the therapy of eye diseases (6-8).
Growing evidence has revealed the vital role of
complement signaling in retinal physiopathology (9,10).
Complement components, such as C2, C4b, C5/C5a and C9, have been
indicated to be elevated in the vitreous humor of patients with
RRDCD (11). Of note, RPE cells
become more susceptible to complement-mediated damage (10). Complement and its receptor
signaling may activate oxidative stress, contribute to
transcriptional and metabolic homeostasis and promote inflammation
in RPE (12,13). C3a and C5a, the bioactive fragments
of C3 and C5, may upregulate the secretion of VEGF in RPE (14). In addition, RPE expresses the
complement receptors C3aR and C5aR, and exhibits an increase in
free cytosolic Ca2+, PI3K/Akt activation and forkhead
box (FOX)P3, and FOXO1 phosphorylation and cytokine/chemokine
secretion correspond to C3a and C5a (15). In another study, mice with
complement factor H (CFH) mutation exhibited retinal degeneration,
edema and detachment, which was rescued by pharmacologic blockade
of C5(16). Furthermore, the
deficit of C3a-C3aR and C5a-C5aR signaling was confirmed to be
linked to abnormal retinal structure and function (9).
However, it has been confirmed that the
participation of C3a-C3aR and C5a-C5aR pathways in various cellular
processes is mediated by the regulation of distinct signaling
pathways, such as MAPK, NF-κB, PI3K-Akt and nuclear factor of
activated T cells (NFAT) (17-20).
Of note, both C3a-C3aR and C5a-C5aR pathways may trigger the
activation of the NF-κB signaling pathway (9). Accumulating evidence has indicated
that phenotypic alterations of RPE, including the junctions between
RPE, the expression of inflammatory cytokines within RPE and its
antioxidant ability, are frequently related to the NF-κB signaling
pathway (21-23).
Furthermore, a recent study has established a close association
among NF-κB signaling, CFH loss-induced abnormality of the
complement system and the occurrence of inflammation in RPE cells
(24). However, it remains
undetermined whether the C3a-C3aR and C5a-C5aR pathways are able to
regulate RPE and what is the underlying mechanism, in particular in
the context of RD or RRDCD.
To this end, the present study intended to explore
the role of the C3a-C3aR and C5a-C5aR pathways in regulating RPE
viability and inflammation during RD or RRDCD progression and
investigate the underlying molecular mechanisms, with an emphasis
on the NF-κB signaling pathway.
Materials and methods
Data collection
The design of the present study was in compliance
with the tenets of the 1975 Declaration of Helsinki and the
protocol was approved by the Ethics Committee of the Nanjing
Medical University Affiliated Wuxi Second Hospital (Wuxi, China;
no. 2019Y-30). Written informed consent was provided by all 40
patients recruited for the purposes of this study. A total of 20
patients with RRDCD and 20 patients with idiopathic epimacular
membrane (IEM), used as the control, enrolled at Nanjing Medical
University Affiliated Wuxi Second Hospital between January 2020 and
July 2021, were included in the cohort. Individuals with recurrent
or secondary IEM or RRDCD, or a previous endophthalmitis
complication, or a history of eye surgery during the last six
months and vitreous hemorrhage, were excluded from this study. The
age and sex were matched between the two groups. For all recruited
participants, two senior experienced ophthalmologists performed a
systematic and comprehensive eye examination. The numbers of
detached quadrants and the PVR grades were scored according to the
1983 International Classification of PVR (25) in patients with RRDCD and were then
evaluated.
Collection of vitreous samples
Prior to primary pars plana vitrectomy, the vitreous
samples were obtained using a three-port 25-gauge transconjunctival
suture-less vitrectomy system (TSV25G; Alcon Constellation; Alcon
Laboratories) with the aid of a non-contact wide-angle viewing
system (Resight; Carl Zeiss Meditec AG) for visualization. The
samples were then suctioned directly into a 5-ml syringe, were
immediately transferred into microcentrifuge tubes and kept on ice.
Following centrifugation at 1,360 x g for 10 min at 4˚C, the
supernatants were collected and stored at -80˚C prior to
analysis.
Cell culture
Human RPE (HRPE) cells (ARPE-19; CRL-2302) purchased
from the American Type Culture Collection were maintained in DMEM
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) and penicillin/streptomycin
(Gibco; Thermo Fisher Scientific, Inc.) in a humidified incubator
with 5% CO2 at 37˚C.
Cell treatment
HRPE cells were divided into six groups and treated
as follows. In the first group, HRPE cells (3x103 or
3x105 per well) were subjected to 0.5, 1, 2, 4 and 8
µg/ml of recombinant endotoxin-free human complement component C3a
protein (R&D Systems, Inc) for 24 or 48 h. In the second group,
HRPE cells (3x103 or 3x105 per well) were
exposed to 0.5, 1, 2, 4 and 8 µg/ml of recombinant endotoxin-free
human complement component C5a protein (R&D Systems, Inc.) for
24 or 48 h (26). In the third
group, HRPE cells (3x105 per well) were treated with
recombinant human complement component C3a protein (2 µg/ml) and
NF-κB inhibitor pyrrolidine dithiocarbamate (PDTC; 10 µM;
MilliporeSigma) for 24 h. In the fourth group, HRPE cells
(3x105 per well) received treatment of recombinant human
complement component C5a protein (1 µg/ml) and PDTC (10 µM) for 24
h. In the fifth group, HRPE cells (3x105 per well) were
treated with recombinant human complement component C3a protein (2
µg/ml) and C3aR antagonist SB290157 (20 µM; MilliporeSigma) for 24
h. In the sixth group, HRPE cells (3x105 per well) were
treated with recombinant human complement component C5a protein (1
µg/ml) and C5aR antagonist CCX168 (2 µM; Abmole Bioscience, Inc.)
for 24 h.
Measurement of cell viability
Approximately 3x103 HRPE cells were
seeded in each well of a 96-well/plate and incubated overnight.
Following 0, 12, 24 and 48 h of treatment, 10 µl Cell Counting
Kit-8 solution (Signalway Antibody LLC) was applied to the cells
for 1 h. The viability of the HRPE cells was evaluated by measuring
the absorbance at 450 nm on a microplate reader (Hua Dong).
ELISA
Approximately 3x105 HRPE cells were
seeded in each well of a 6-well/plate and incubated overnight.
Following 24 h of treatment, the contents of various cytokines and
other compounds were analyzed in HRPE cell supernatants and/or
human vitreous humor were determined using the following ELISA kits
in accordance with the manufacturers' protocols: Tumor Necrosis
Factor-α Assay Kit (cat. no. H052-1), Interleukin-1β Assay Kit
(cat. no. H002), Interleukin-6 Assay Kit (cat. no. H007-1-1) and
Interleukin-10 Assay Kit (cat. no. H009-1; all from Nanjing
Jiancheng Bioengineering Institute); prostaglandin E2 (PGE2) ELISA
Kit (cat. no. E-EL-0034c; Elabscience Biotechnology Co., Ltd.);
Human Complement Fragment 3a ELISA Kit (cat no. CSB-E08509h) and
Human Complement Fragment 5a ELISA Kit (cat. no. CSB-E08512h; both
from Cusabio).
Reverse transcription-quantitative
(RT-q)PCR
Approximately 3x105 HRPE cells were
seeded in each well of a 6-well/plate and incubated overnight.
Following 24 h of treatment, TRIzol® reagent (Thermo
Fisher Scientific, Inc.) was used for the extraction of total RNA
from the HRPE cells, while the PrimeScript RT reagent kit (Takara
Bio, Inc.) was employed according to the manufacturer's protocol
for cDNA synthesis. qPCR was performed with the SYBR Green PCR
master mix on an ABI 9700 PCR machine (both from Applied
Biosystems; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The following thermocycling conditions
were used for qPCR: 95˚C for 10 min, followed by 40 cycles at 95˚C
for 15 sec and 60˚C for 45 sec; final extension at 95˚C for 15 sec,
60˚C for 1 min, 95˚C for 15 sec and 60˚C for 15 sec. The PCR primer
pairs were as follows: C3aR forward, 5'-TCCTTTATGCCCTCTTGG-3' and
reverse, 5'-GAACCGCTGGATTGATTC-3'; C5aR forward,
5'-TAGGGAGAGCAAGTCATTC-3' and reverse, 5'-TGGAGGAGAGTTAGTTAGC-3';
and GAPDH forward, 5'-AATCCCATCACCATCTTC-3' and reverse,
5'-AGGCTGTTGTCATACTTC-3'. The 2-ΔΔCq method was used to
calculate the expression levels (13).
Western blot analysis
Approximately 3x105 HRPE cells were
seeded in each well of a 6-well/plate and incubated overnight.
Following 24 h of treatment, RIPA buffer (Beyotime Institute of
Biotechnology) was employed to prepare the cell lysates. The
protein concentration was measured using a bicinchoninic acid
protein assay kit (Pierce; Thermo Fisher Scientific, Inc.), and
absorbance was measured using a microplate reader (SM600 Labsystem;
Shanghai Utrao Medical Instrument Co., Ltd.). Proteins (20 µg) were
fractioned by 10 or 15% SDS-PAGE and were subsequently transferred
onto polyvinylidene fluoride membranes (EMD Millipore), which were
subsequently incubated with 5% fat-free milk (Solarbio) for 1 h.
The membranes were then exposed to primary antibodies against C3aR
(cat. no. ab126250), C5aR (cat. no. ab234757; both from Abcam),
phospho (p)-NF-κB (cat. no. 3033), NF-κB (cat. no. 8242) and GAPDH
(cat. no. 5174; all from Cell Signaling Technology, Inc.) at 4˚C
overnight, washed three times with Tris-buffered saline containing
Tween-20 and then incubated with the secondary antibody (cat. no.
A0208; Beyotime Institute of Biotechnology) for 1 h at 37˚C. The
chemiluminescent signals of the targets were generated with an
Enhanced Chemiluminescence Detection kit (Pierce; Thermo Fisher
Scientific, Inc.). GAPDH was used as an internal control.
Statistical analysis
Prism v.8.0.2 (GraphPad Software, Inc.) was used to
perform all statistical analyses. The mean ± standard deviation was
used to denote data from triplicates. The Shapiro-Wilk normality
test was used to evaluate the normal distribution of the collected
data. When data followed a normal distribution, a two-sided
Student's t-test or one-way ANOVA with Tukey's post-hoc test were
adopted. P<0.05 was considered to indicate a statistically
significant difference.
Results
Complement C3a and C5a promote cell
viability and inflammation of HRPE cells
Table I presents
the demographic and clinical data of the patients enrolled in the
study (20 patients with RRDCD and 20 patients with IEM). Patients
with RRDCD or IEM included 10 males and 10 females, aged 63.1±5.22
and 64.1±6.69, respectively. To determine the possible role of
complement C3a and C5a in the progression of RD or RRDCD, their
contents were examined in patients with RRDCD (n=20) and in
individuals with IEM as controls (n=20) by ELISA. The results
revealed that the levels of both C3a and C5a in patients with RRDCD
were markedly higher than those in patients with IEM (Fig. 1A and B). To investigate the relevance of this
C3a and C5a alteration to visual disorders, HRPE cells were treated
with various doses of purified recombinant human complement C3a and
C5a protein. The application of C3a or C5a triggered a considerable
increase in HRPE cell viability (Fig.
1C and D), suggesting that
these factors influence cell survival. Considering the relevance of
these factors to the occurrence of inflammation, the secretion of
inflammatory cytokines, such as TNF-α, IL-1β, IL-6, IL-10 and PGE2,
in C3a- or C5a-treated cells, was evaluated by ELISA. As indicated
in Fig. 1E-H, the secretion of
TNF-α, IL-1β, IL-6 and PGE2 by HRPE cells was markedly enhanced by
C3a or C5a stimulation and a dose-dependent effect was observed.
However, C3a or C5a stimulation significantly decreased IL-10
secretion (Fig. 1I). These results
indicated that elevated C3a or C5a levels may be associated with
increased cell viability and inflammatory response in HRPE
cells.
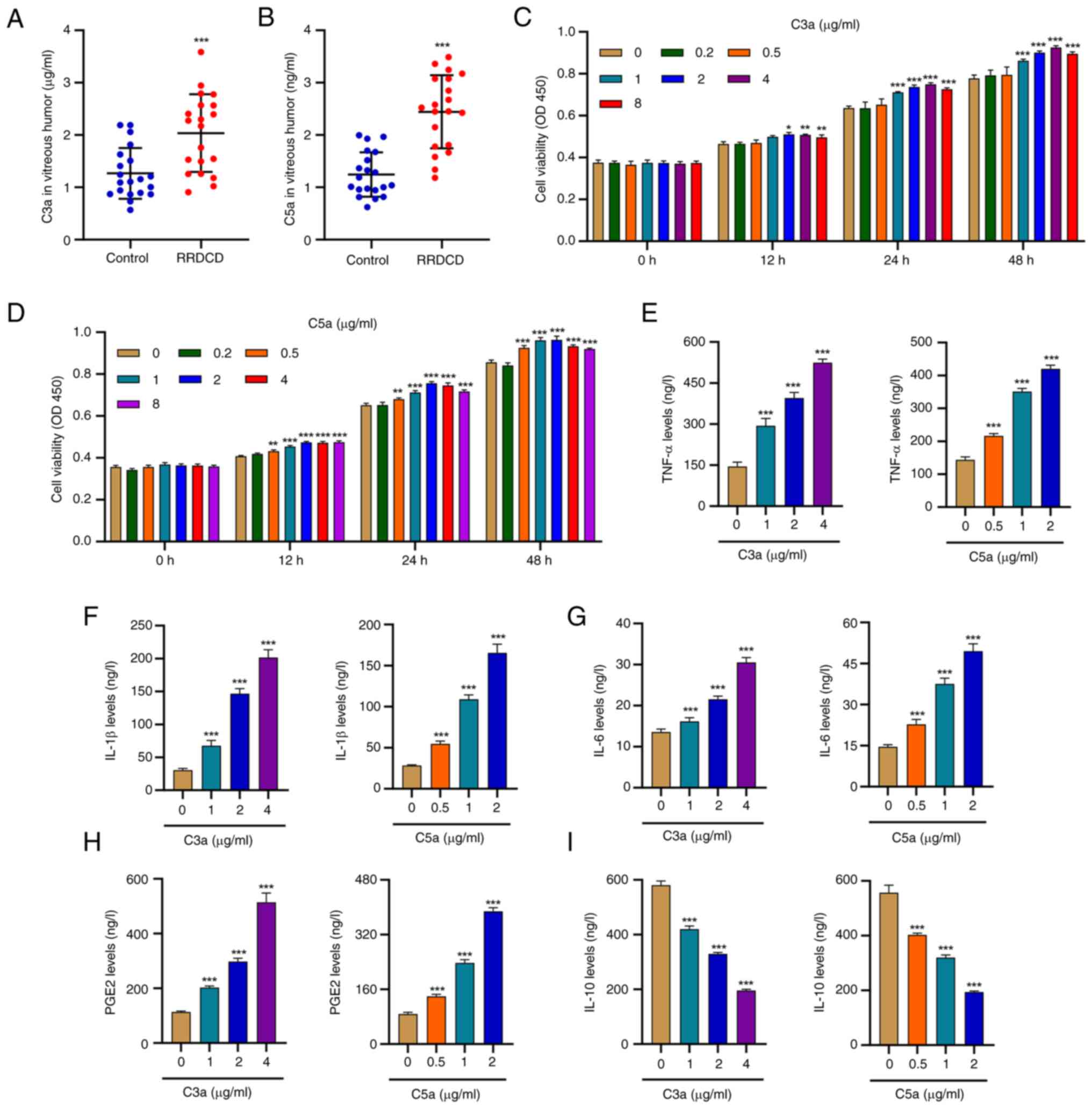 | Figure 1Complement C3a and C5a promote cell
viability and inflammation of HRPE cells. (A) Complement C3a and
(B) C5a protein concentrations in patients with RRDCD (n=20) and
idiopathic epimacular membrane as a control (n=20). HRPE cells were
treated with different concentrations of recombinant human
complement component C3a or C5a, and the cell viability following
(C) C3a or (D) C5a treatment, and the release of (E) TNF-α, (F)
IL-1β, (G) IL-6, (H) PGE2 and (I) IL-10 were measured.
*P<0.05, **P<0.01,
***P<0.001 compared with control or 0 µg/ml. OD450,
optical density at 450 nm; PGE2, prostaglandin E2; RRDCD,
rhegmatogenous retinal detachment associated with choroidal
detachment; HRPE, human retinal pigment epithelium. |
 | Table IClinical characteristics of the study
population. |
Table I
Clinical characteristics of the study
population.
| Item | RRDCD (n=20) | IEM (n=20) |
|---|
| Sex | | |
|
Male | 10 | 10 |
|
Female | 10 | 10 |
| Age, years | 63.1±5.22 | 64.1±6.69 |
| Duration of
detachments, days | 9 (7-13) | - |
| PVR grade | | |
|
Mild, A,
B | 5 | - |
|
Heavy, C,
D | 15 | - |
| IOP, mmHg | 7.5 (6.9-7.9) | - |
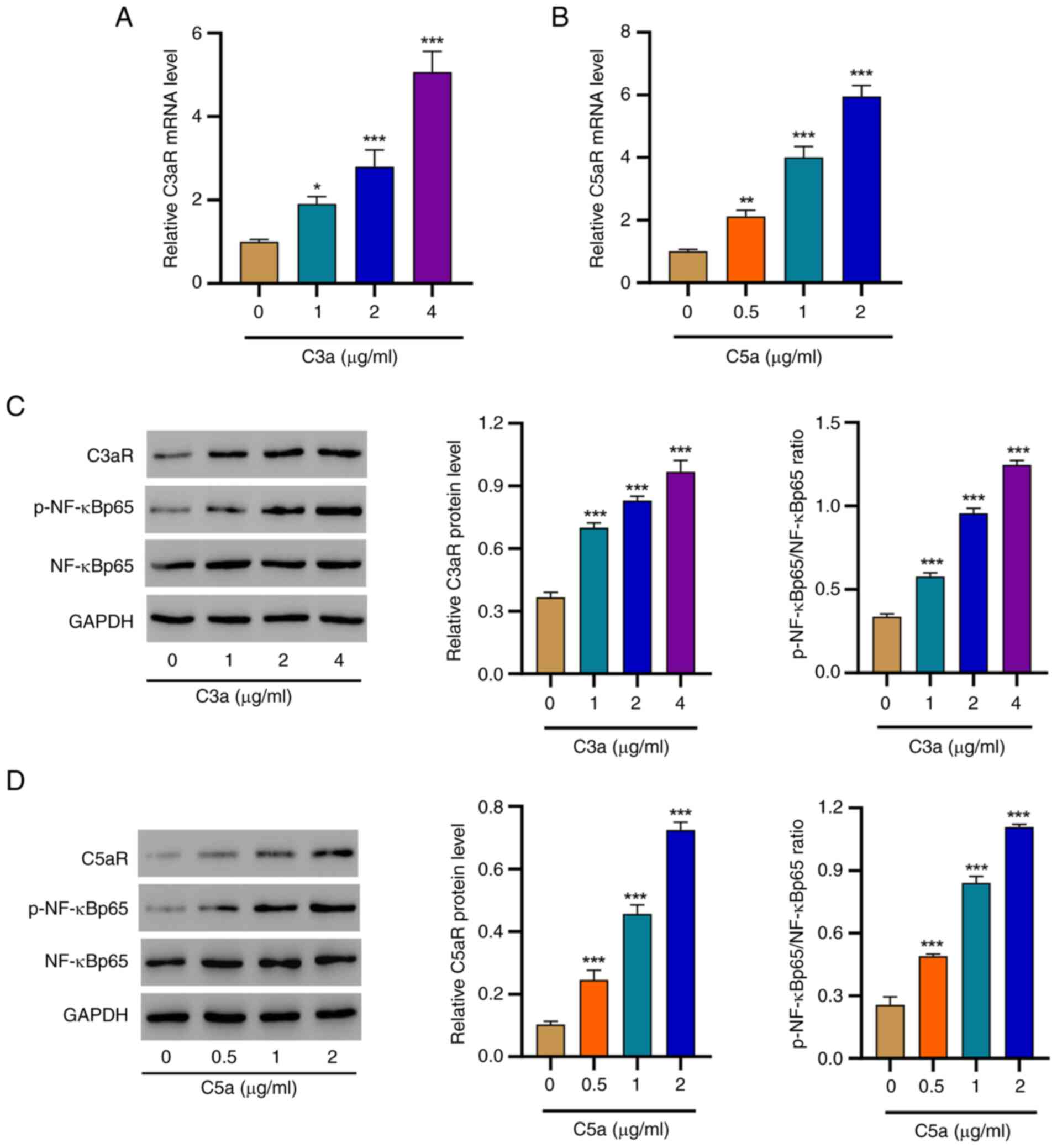
Complement C3a and C5a enhance the
expression of C3aR and C5aR and activate NF-κB signaling
It is established that C3a and C5a act through
binding to their corresponding receptors. Indeed, markedly
increased mRNA levels of C3aR or C5aR were observed in the presence
of the C3a or C5a ligands, respectively (Fig. 2A and B). Considering the vital role of the
NF-κB signaling pathway in mediating complement-triggered
responses, the activation of this pathway was investigated by
western blot analysis. Of note, the protein levels of C3aR and the
p-NF-κBp65/NF-κBp65 ratio gradually increased with increasing C3a
concentrations (Fig. 2C), and a
similar trend was observed in the presence of various C5a
concentrations (Fig. 2D). These
results demonstrated the upregulation of C3aR and C5aR and
activation of the NF-κB pathway in response to C3a and C5a
challenge.
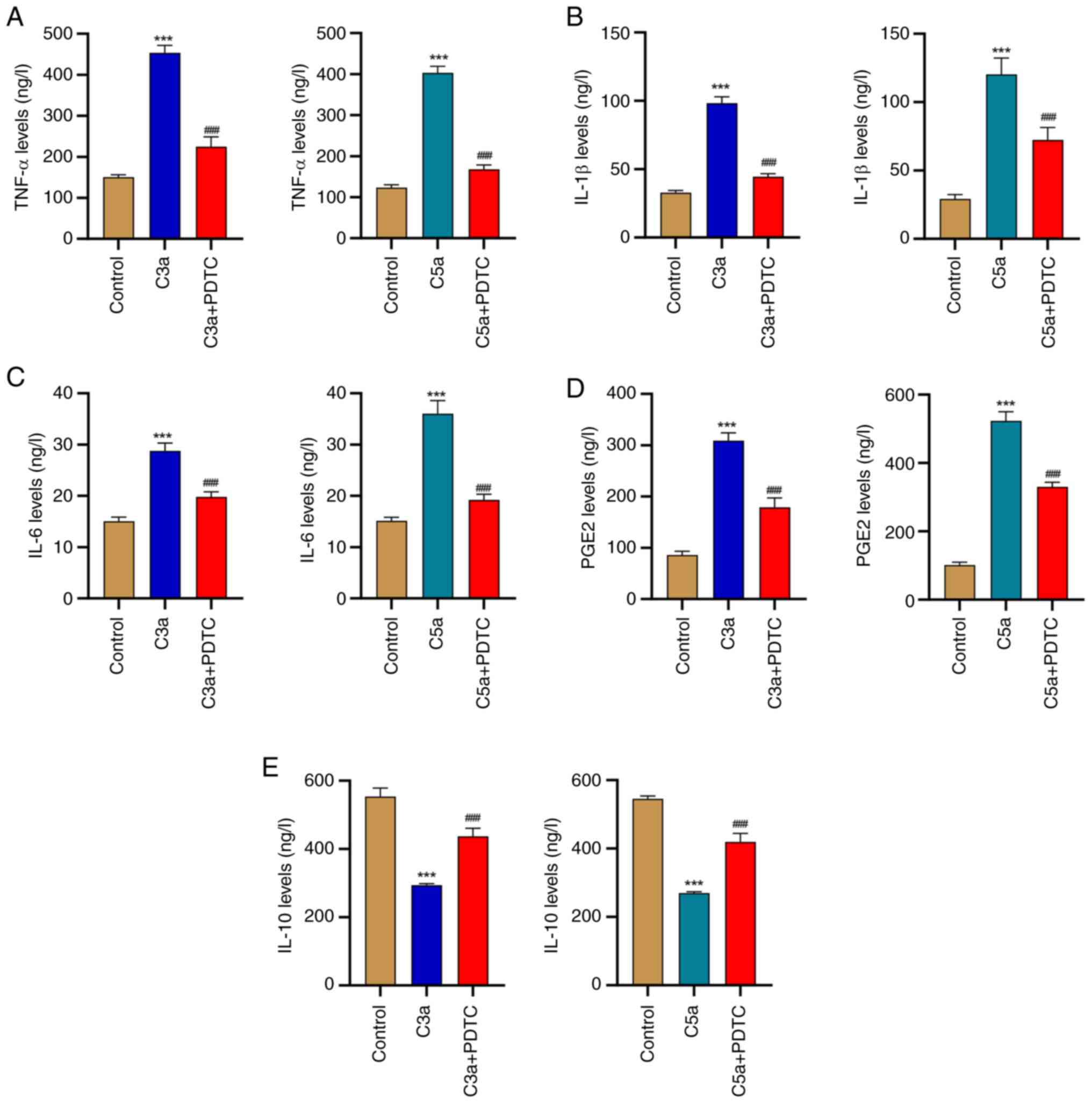
Complement C3a and C5a promote
inflammation of HRPE cells via the NF-κB signaling pathway
To confirm that C3a and C5a triggered inflammation
in HRPE cells is indeed mediated by NF-κB signaling, the cytokine
levels were analyzed following incubation with or without the NF-κB
inhibitor PDTC. Administration of C3a or C5a induced the release of
TNF-α, IL-1β, IL-6 and PGE2, while this promoting effect was
inhibited by the application of PDTC in HRPE cells (Fig. 3A-D). However, C3a or C5a
administration inhibited the release of IL-10, while this effect
was inhibited by the application of PDTC in HRPE cells (Fig. 3E). These findings demonstrate that
C3a and C5a triggered inflammatory responses in a process that is
largely mediated by the NF-κB pathway.
Complement C3aR and C5aR antagonists
repress inflammation and NF-κB signaling in HRPE cells challenged
with complement C3a and C5a
To further confirm the role of the C3a-C3aR and
C5a-C5aR pathways in the occurrence of inflammation and the
activation of NF-κB signaling in HRPE cells, antagonists of these
receptors were also applied. C3a-mediated secretion of TNF-α,
IL-1β, IL-6, IL-10 and PGE2 was inhibited by the presence of the
C3aR antagonist SB290157 (Fig.
4A-E). Likewise, the C5a-mediated release of these cytokines
was repressed by the application of the C5aR antagonist CCX168
(Fig. 4F-J). In addition,
activation of NF-κB induced by C3a or C5a was robustly blocked by
their corresponding receptor antagonists (Fig. 4K and L). These findings validate that the
C3a-C3aR and C5a-C5aR pathways participate in the subsequent
inflammatory responses but also in NF-κB activation in HRPE
cells.
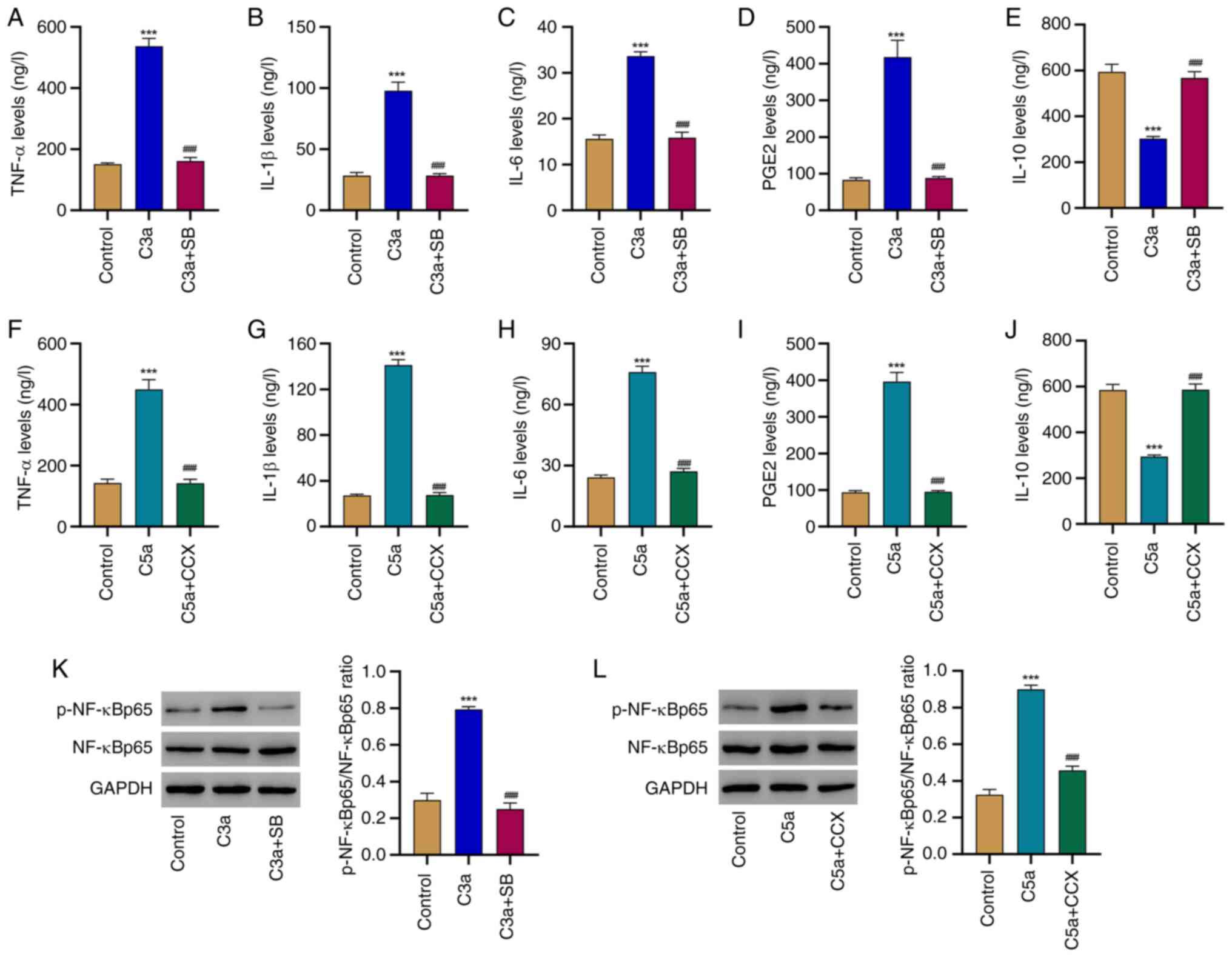 | Figure 4Complement C3aR and C5aR antagonist
inhibit inflammation and NF-κB signaling in HRPE cells challenged
with complement C3a and C5a. HRPE cells were treated with
recombinant human complement component C3a (2 µg/ml) and with or
without C3aR antagonist SB290157 (20 µM), and the release of (A)
TNF-α, (B) IL-1β, (C) IL-6, (D) PGE2 and (E) IL-10 was determined
by ELISA. HRPE cells were treated with recombinant human complement
component C5a (1 µg/ml) and with or without C5aR antagonist CCX168
(2 µM), and the release of (F) TNF-α, (G) IL-1β, (H) IL-6, (I) PGE2
and (J) IL-10 was determined by ELISA. (K) The phosphorylation of
NF-κB and expression of NF-κB in HRPE cells treated with
recombinant human complement component C3a (2 µg/ml) and with or
without C3aR antagonist SB290157 (20 µM) were determined by western
blot. (L) The phosphorylation of NF-κB and expression of NF-κB in
HRPE cells treated with recombinant human complement component C5a
(1 µg/ml) and with or without C5aR antagonist CCX168 (2 µM) were
determined by western blot. ***P<0.001 relative to
control; ###P<0.001 relative to C3a or C5a treatment.
p-NF-κB, phosphorylated NF-κB; C5aR, C5a receptor; HRPE, human
retinal pigment epithelium; CCX, CCX168; SB, SB290157. |
Complement C3a and C5a are positively
correlated with the contents of TNF-α, IL-1β, IL-6 and PGE2 in
patients with RRDCD
Finally, the cytokine levels in patients with RRDCD
and the controls were analyzed and the relationship of C3a and C5a
with these inflammatory factors was assessed. As presented in
Fig. 5A-E, the patients with RRDCD
had significantly higher contents of TNF-α, IL-1β, IL-6 and PGE2
and a lower content of IL-10 compared to the controls, indicating a
stronger inflammatory response in these individuals. Furthermore,
the levels of TNF-α, IL-1β, IL-6 and PGE2 exhibited a positive
correlation, as opposed to IL-10, which had a negative correlation
with C3a (Fig. 5E-J) and C5a
(Fig. 5K-O). These data suggested
that progression of RD or RRDCD disorders may be accompanied by
higher levels of inflammation. In addition, it may be inferred that
this condition is associated with elevated C3a and C5a
contents.
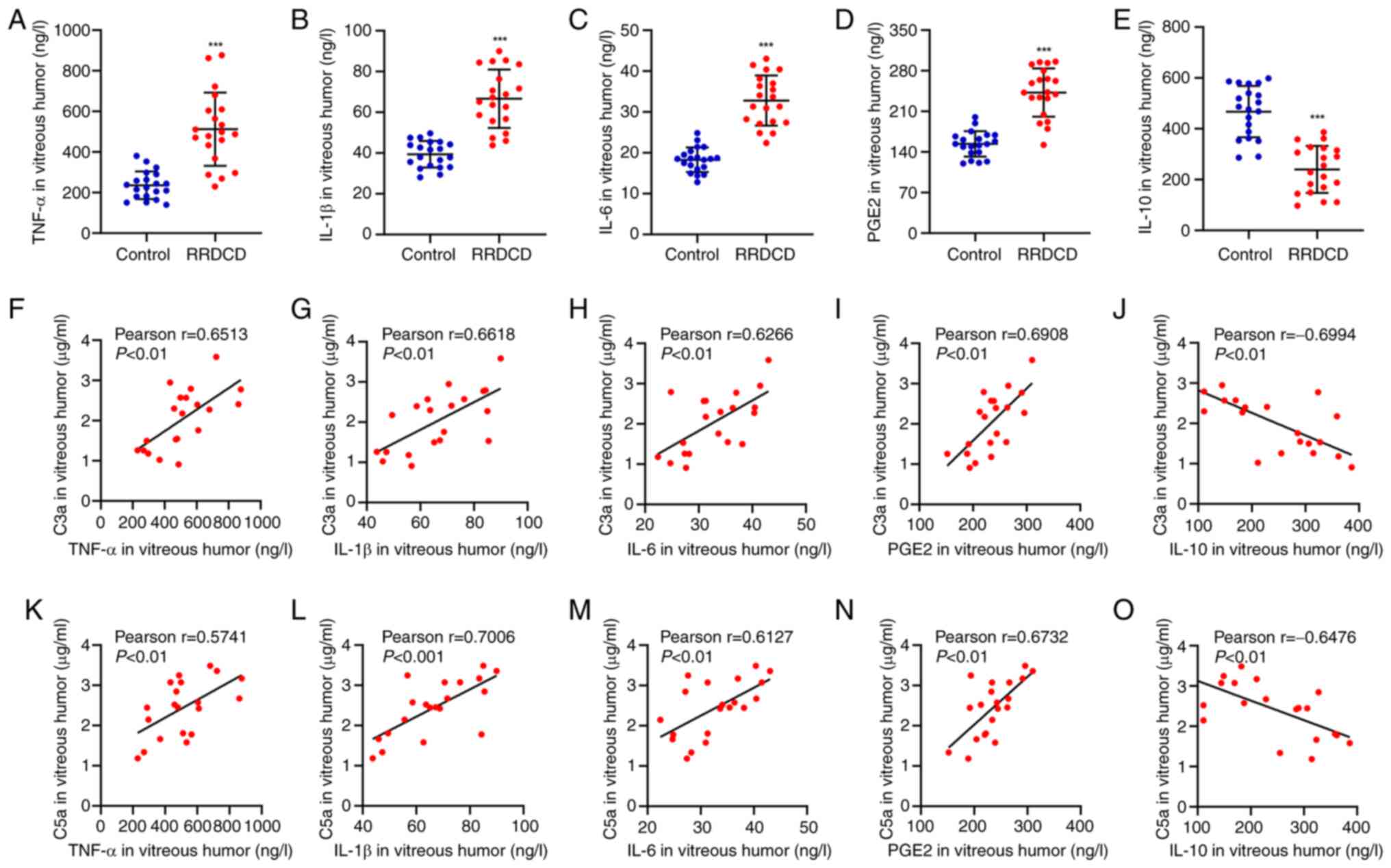 | Figure 5Complement C3a and C5a are positively
correlated with the contents of TNF-α, IL-1β, IL-6 and PGE2 in
patients with RRDCD. (A) TNF-α, (B) IL-1β, (C) IL-6, (D) PGE2 and
(E) IL-10 concentrations in patients with RRDCD (n=20) and
idiopathic epimacular membrane as control (n=20). Pearson
correlation scatter plots between C3a and (F) TNF-α, (G) IL-1β, (H)
IL-6, (I) PGE2 or (J) IL-10, or between C5a and (K) TNF-α, (L)
IL-1β, (M) IL-6, (N) PGE2 or (O) IL-10 in patients with RRDCD
(n=20). ***P<0.001 compared with control. PGE2,
prostaglandin E2; RRDCD, rhegmatogenous retinal detachment
associated with choroidal detachment. |
Discussion
Loss or dysfunction of photoreceptors or the
adjacent RPE is strongly associated with the progression of retinal
diseases, which affect millions of individuals globally (24,27),
highlighting the necessity to investigate the pathogenesis and
develop novel therapeutic strategies. In the present study, C3a and
C5a contents were higher in patients with RRDCD than in control
subjects. Consistently, elevated C3a and C5a levels led to
increased cell viability and aggravated inflammation in HRPE cells.
The results of the present mechanistic study indicated that higher
C3a and C5a contents were able to enhance the expression levels of
their corresponding receptors C3aR and C5aR, which in turn resulted
in NF-κB signaling activation in vitro. This was further
supported by the fact that the patients with RRDCD had higher
levels of inflammatory factors, such as TNF-α, IL-1β, IL-6 and
PGE2. Furthermore, the C3a and C5a levels were positively
correlated with these cytokines.
It has been demonstrated that inflammatory responses
are present in common pathological alterations of photoreceptor and
accessory cells in RD and other related disorders (28). Previous studies have indicated that
the C3a-C3aR and C5a-C5aR signaling axes are crucial pathways for
triggering and modulating inflammatory response; accordingly, they
are linked to numerous inflammation-related disorders (29-31).
Of note, these pathways have also been validated to be essential
for preserving normal retinal structure and function (9). The present study revealed
significantly enhanced levels of both C3a and C5a under the
pathological conditions of a retinal disorder, i.e. RRDCD.
Furthermore, the elevation of these complement components was
accompanied by an increase in a series of inflammatory factors. A
previous study reported increased inflammatory response in RRDCD
(7). Although altered inflammatory
mediators were inconsistent with the observations of the present
study, this finding may be due to the different scope of the
examined targets. The present results highlight the role of
C3a-C3aR and C5a-C5aR signaling in cell viability and inflammation
in HRPE cells.
It is well-known that the NF-κB signaling pathway is
a vital modulator of cell survival and inflammation (24,32-34).
When the NF-κB signaling pathway is activated, the NF-κB protein
undergoes phosphorylation modification (35). C3a-C3aR signaling significantly
enhances T-cell proliferation and IL-17A expression, a process that
is mediated by the activation of NF-κB in T cells (36). C5a-C5aR signaling enhances the IL-8
promoter and activates NF-κB in the human leukemia cell line HL-60,
peripheral blood mononuclear cells and RAW264.7 macrophage cells
(37). Consistently, it was
observed in the present study that increases of C3a or C5a were
accompanied by an increase in the level of p-NF-κBp65, suggesting
that the NF-κB signaling pathway mediates the phenotypic
alterations of HRPE cells induced by these complement components.
These findings were further confirmed by the application of C3aR
and C5aR antagonists or NF-κB inhibitor. C3a was previously
reported to promote the production of chemokine through NFAT
activation in mast cells (19),
and it was indicated that its regulation on its receptor gene was
mediated by activator protein-1 in glial cells (19,20).
Furthermore, the activation of the PI3K/Akt pathway by both
C3a-C3aR and C5a-C5aR in T cells has been previously described in
the literature (29). The present
results robustly demonstrated that both the C3a-C3aR and C5a-C5aR
pathways were able to activate NF-κB signaling, thereby regulating
the phenotypes of HRPE cells, which is, to a certain degree,
consistent with their important role in retinal structure and
function (9). Furthermore, NF-κB
regulation by the C3a-C3aR pathway in tubular epithelial cells has
been described previously (18);
therefore, whether C3a-C3aR has the same role in other epithelial
cells and whether the C5a-C5aR pathway has a similar function
require to be investigated. In addition, the RPE is a physical
barrier and provides immune-suppressive molecules, which contribute
to the privileged immune status of the eye (38). Damage to the RPE layer and the
underlying choroid may promote infiltration of immune cells and
induce retinal microglia and choroidal inflammatory cells to
release pro-inflammatory cytokines. A previous study suggested that
the anti-inflammatory cytokine IL-10 inhibits RPE cell
proliferation and inflammation through the regulation of VEGF in
RRD rats (39), a finding that is
consistent with the role of IL-10 in C3a-C3aR- or C5a-C5aR-induced
RPE cells. However, the role of C3a-C3aR or C5a-C5aR in the
regulation of RPE and immune cells at the retina-choroidal
interface remains to be investigated. In the present study, C3a,
C5a, TNF-α, IL-1β, IL-6 and PGE2 contents were higher, while the
IL-10 content was lower in patients with RRDCD than in control
subjects. Although no public datasets are currently available for
patients with RRDCD, the complement anaphylatoxins and inflammatory
factors detected in the present study may be further supplemented
with data from our institution including additional patients.
Meanwhile, selection bias may be another potential limitation.
Future studies will aim to determine the correlation between
complement anaphylatoxins and inflammatory factors in patients with
recurrent or secondary RRDCD.
In conclusion, the present results revealed the
significance of the complement components C3a and C5a, as well as
their pathways, in the pathogenesis of retinal disorders such as
RRDCD. These molecules and their pathways influenced the viability
of HRPE cells and enhanced the cellular inflammatory level through
regulation of the NF-κB signaling pathway. This finding suggests
that C3a and C5a are both involved in the progression of retinal
diseases and that the C3a-C3aR and C5a-C5aR pathways, as well as
their downstream NF-κB signaling, may be considered novel targets
for the development of new drugs to treat RD and related
disorders.
Acknowledgements
Not applicable.
Funding
Funding: This study was supported by the Social Development
Project of Jiangsu Provincial Science and Technology Department
(grant no. BE2017627), the youth project of Wuxi Health Committee
(grant no. Q202038) and the reserve top talent project of Wuxi
Municipal Health Commission (grant no. HB2020032).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SL and HX designed this study. HX, XG and JS
performed the experiments. SL, XC and ZW analyzed and interpreted
the data. XG and JS checked and approved the authenticity of the
raw data. SL and ZW wrote the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
This study is in compliance with the tenets of the
1975 Declaration of Helsinki and was approved by the Ethics
Committee of Nanjing Medical University Affiliated Wuxi Second
Hospital (Wuxi, China; no. 2019Y-30). Written informed consent was
obtained from each patient.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Leach LL and Clegg DO: Concise review:
Making stem cells retinal: Methods for deriving retinal pigment
epithelium and implications for patients with ocular disease. Stem
Cells. 33:2363–2373. 2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Xu H, Lutrin D and Wu Z: Outcomes of
23-gauge pars plana vitrectomy combined with phacoemulsification
and capsulotomy without intraocular lens implantation in
rhegmatogenous retinal detachment associated with choroidal
detachment. Medicine (Baltimore). 96(e7869)2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Xu C, Wu J and Feng C: Changes in the
postoperative foveal avascular zone in patients with rhegmatogenous
retinal detachment associated with choroidal detachment. Int
Ophthalmol. 40:2535–2543. 2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Wu Z, Ding N, Yu M, Wang K, Luo S, Zou W,
Zhou Y, Yan B and Jiang Q: Identification of potential biomarkers
for rhegmatogenous retinal detachment associated with choroidal
detachment by vitreous iTRAQ-based proteomic profiling. Int J Mol
Sci. 17(2052)2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Klettner A and Roider J: Retinal pigment
epithelium expressed toll-like receptors and their potential role
in age-related macular degeneration. Int J Mol Sci.
22(8387)2021.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Al-Nawaiseh S, Thieltges F, Liu Z, Strack
C, Brinken R, Braun N, Wolschendorf M, Maminishkis A, Eter N and
Stanzel BV: A step by step protocol for subretinal surgery in
rabbits. J Vis Exp. (53927)2016.PubMed/NCBI View
Article : Google Scholar
|
|
7
|
Dai Y, Wu Z, Sheng H, Zhang Z, Yu M and
Zhang Q: Identification of inflammatory mediators in patients with
rhegmatogenous retinal detachment associated with choroidal
detachment. Mol Vis. 21:417–427. 2015.PubMed/NCBI
|
|
8
|
Strauss O: The retinal pigment epithelium
in visual function. Physiol Rev. 85:845–881. 2005.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Yu M, Zou W, Peachey NS, McIntyre TM and
Liu J: A novel role of complement in retinal degeneration. Invest
Ophthalmol Vis Sci. 53:7684–7692. 2012.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Ao J, Wood JP, Chidlow G, Gillies MC and
Casson RJ: Retinal pigment epithelium in the pathogenesis of
age-related macular degeneration and photobiomodulation as a
potential therapy? Clin Exp Ophthalmol. 46:670–686. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Luo S, Chen Y, Yang L, Gong X and Wu Z:
The complement system in retinal detachment with choroidal
detachment. Curr Eye Res. 1–14. 2022.PubMed/NCBI View Article : Google Scholar : (Epub ahead of
print).
|
|
12
|
Wang J, Wang Y, Yu D, Liu Q, Lin S, Tian
R, Li J and Luo Y: Protective effect of a bispecific Fc-fusion
protein on the barrier of human retinal pigment epithelial cells.
Ophthalmic Res. 64:656–663. 2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Hollborn M, Ackmann C, Kuhrt H, Doktor F,
Kohen L, Wiedemann P and Bringmann A: Osmotic and hypoxic induction
of the complement factor C9 in cultured human retinal pigment
epithelial cells: Regulation of VEGF and NLRP3 expression. Mol Vis.
24:518–535. 2018.PubMed/NCBI
|
|
15
|
Busch C, Annamalai B, Abdusalamova K,
Reichhart N, Huber C, Lin Y, Jo EAH, Zipfel PF, Skerka C, Wildner
G, et al: Anaphylatoxins activate Ca2+, Akt/PI3-kinase,
and FOXO1/FoxP3 in the retinal pigment epithelium. Front Immunol.
8(703)2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Song D, Ueda Y, Bhuyan R, Mohammed I, Miwa
T, Gullipali D, Kim H, Zhou L, Song Y, Schultz H, et al: Complement
factor H mutation W1206R causes retinal thrombosis and ischemic
retinopathy in mice. Am J Pathol. 189:826–838. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Bamberg CE, Mackay CR, Lee H, Zahra D,
Jackson J, Lim YS, Whitfeld PL, Craig S, Corsini E, Lu B, et al:
The C5a receptor (C5aR) C5L2 is a modulator of C5aR-mediated signal
transduction. J Biol Chem. 285:7633–7644. 2010.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Thurman JM, Lenderink AM, Royer PA,
Coleman KE, Zhou J, Lambris JD, Nemenoff RA, Quigg RJ and Holers
VM: C3a is required for the production of CXC chemokines by tubular
epithelial cells after renal ishemia/reperfusion. J Immunol.
178:1819–1828. 2007.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Ahamed J, Venkatesha RT, Thangam EB and
Ali H: C3a enhances nerve growth factor-induced NFAT activation and
chemokine production in a human mast cell line, HMC-1. J Immunol.
172:6961–6968. 2004.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Martin CB, Ingersoll SA and Martin BK:
Transcriptional control of the C3a receptor gene in glial cells:
Dependence upon AP-1 but not Ets. Mol Immunol. 44:703–712.
2007.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Jo DH, Cho CS and Kim JH and Kim JH:
Intracellular amyloid-β disrupts tight junctions of the retinal
pigment epithelium via NF-κB activation. Neurobiol Aging.
95:115–122. 2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Feng Q, Yang W, Gao Z, Ruan X and Zhang Y:
Up-regulation of P-gp via NF-κB activation confers protection
against oxidative damage in the retinal pigment epithelium cells.
Exp Eye Res. 181:367–373. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Hu Y, Lin H, Dib B, Atik A, Bouzika P, Lin
C, Yan Y, Tang S, Miller JW and Vavvas DG: Cholesterol crystals
induce inflammatory cytokines expression in a human retinal pigment
epithelium cell line by activating the NF-κB pathway. Discov Med.
18:7–14. 2014.PubMed/NCBI
|
|
24
|
Armento A, Schmidt TL, Sonntag I, Merle
DA, Jarboui MA, Kilger E, Clark SJ and Ueffing M: CFH loss in human
RPE cells leads to inflammation and complement system dysregulation
via the NF-κB pathway. Int J Mol Sci. 22(8727)2021.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Machemer R, Aaberg TM, Freeman HM, Irvine
AR, Lean JS and Michels RM: An updated classification of retinal
detachment with proliferative vitreoretinopathy. Am J Ophthalmol.
112:159–165. 1991.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Bröker K, Terzenbach R, Bentzien F, Lüth S
and Dammermann W: Complement factors C3a and C5a mimick a
proinflammatory microenvironment and increase HBV IGRA sensitivity.
J Transl Med. 17(6)2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Huang Y, Enzmann V and Ildstad ST: Stem
cell-based therapeutic applications in retinal degenerative
diseases. Stem Cell Rev Rep. 7:434–445. 2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Murakami Y, Notomi S, Hisatomi T, Nakazawa
T, Ishibashi T, Miller JW and Vavvas DG: Photoreceptor cell death
and rescue in retinal detachment and degenerations. Prog Retin Eye
Res. 37:114–140. 2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Strainic MG, Liu J, Huang D, An F, Lalli
PN, Muqim N, Shapiro VS, Dubyak GR, Heeger PS and Medof ME: Locally
produced complement fragments C5a and C3a provide both
costimulatory and survival signals to naive CD4+ T cells. Immunity.
28:425–435. 2008.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Ricklin D, Hajishengallis G, Yang K and
Lambris JD: Complement: A key system for immune surveillance and
homeostasis. Nat Immunol. 11:785–797. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
31
|
Wu KY, Zhang T, Zhao GX, Ma N, Zhao SJ,
Wang N, Wang JX, Li ZF, Zhou W and Li K: The C3a/C3aR axis mediates
anti-inflammatory activity and protects against uropathogenic E
coli-induced kidney injury in mice. Kidney Int. 96:612–627.
2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Luo JL, Kamata H and Karin M:
IKK/NF-kappaB signaling: Balancing life and death-a new approach to
cancer therapy. J Clin Invest. 115:2625–2632. 2005.PubMed/NCBI View
Article : Google Scholar
|
|
33
|
Sun SC and Liu ZG: A special issue on
NF-κB signaling and function. Cell Res. 21:1–2. 2011.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Liu T, Zhang L, Joo D and Sun SC: NF-κB
signaling in inflammation. Signal Transduct Target Ther.
2(17023)2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ling J and He P: miR-361-5p regulates
ovarian cancer cell proliferation and apoptosis by targeting TRAF3.
Exp Ther Med. 21(199)2021.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Liu Y, Wang K, Liang X, Li Y, Zhang Y,
Zhang C, Wei H, Luo R, Ge S and Xu G: Complement C3 produced by
macrophages promotes renal fibrosis via IL-17A secretion. Front
Immunol. 9(2385)2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Dewan SMR, Osaka M, Deushi M and Yoshida
M: Complement C5a-triggered differentiated HL-60 stimulates
migration of THP-1 monocytic leukocytes via secretion of CCL2. FEBS
Open Bio. 11:1374–1381. 2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Luo C, Zhao J, Chen M and Xu H: The
expression of C1 inhibitor (C1INH) in macrophages is upregulated by
retinal pigment epithelial cells-implication in subretinal immune
privilege in the aging eye. Aging (Albany NY). 10:1380–1389.
2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhao Q, Ji M and Wang X: IL-10 inhibits
retinal pigment epithelium cell proliferation and migration through
regulation of VEGF in rhegmatogenous retinal detachment. Mol Med
Rep. 17:7301–7306. 2018.PubMed/NCBI View Article : Google Scholar
|