Introduction
Intrinsic and extrinsic aging are two basic
processes of skin aging. Extrinsic aging is generally referred to
as photoaging and is characterized by severe wrinkling and
pigmentary changes, such as solar lentigo and mottled pigmentation
on exposed areas, such as the face, neck and forearms. Solar
ultraviolet (UV) irradiation is a major environmental hazard that
generates reactive oxygen species (ROS), induces DNA damage, and
ultimately results in skin inflammation, photoaging, and cancer
development (1).
Oxidative stress is considered a primary feature in
aging and age-related diseases, including cataracts,
atherosclerosis, diabetes and Alzheimer’s disease. Aging is
considered to be the consequence of free radical damage by various
endogenous ROS, according to the original free radical theory of
aging. ROS production and release can be affected by environmental
factors such as UV radiation and exogenous toxins. Cytoprotective
responses are characterized by the upregulation of antioxidant
enzymes and decreased sensitivity to oxidative stress damage
(2). Various compounds with
differential antioxidant properties are found in plants, which may
be applicable as therapeutics to decrease and prevent free radical
damage. Several medical plants have been screened and assessed for
properties in antagonizing free radical-induced oxidative stress,
and their natural products are used to treat 87% of all classified
human diseases (3,4).
Recently, it was suggested that excessive matrix
degradation by UV-induced matrix metalloproteinases (MMPs), which
are secreted by various cells including keratinocytes, fibroblasts
and inflammatory cells, contributes to connective tissue damage
during photoaging (5–8). MMPs are a family of enzymes
responsible for degrading connective tissue. They are structurally
related endopeptidases that mediate the degradation of different
macromolecular components of the extracellular matrix (ECM) and the
basement membrane, including collagen. The UV-induced synthesis of
MMPs present on dermal fibroblasts contributes to the breakdown of
dermal interstitial collagen and other connective tissue
components. These results are indicative of the MMP-mediated
degradation of collagen in photodamaged skin. In particular, in
photodamaged aging skin, increased ROS leads to the induction of
AP-1 and NF-κB transcription factors, which consequently induce
collagen degradation by the upregulation of MMPs. These properties
make MMPs an attractive target for anti-photoaging compounds. ECM
degrades naturally over time due to intrinsic aging. However, this
breakdown is accelerated by external factors (especially UV
irradiation) and the resultant oxidative stress, as well as
increases in activity of MMPs.
Fibroblasts are the major cell component of the
dermis that produces ECM proteins, as well as structural proteins
(collagen and elastin), adhesive proteins (laminins and
fibronectin), glycosaminoglycans (GAG) and proteoglycans. The most
abundant structural protein in skin connective tissue is type I
collagen (90% of ECM in dermis), which is synthesized primarily by
fibroblasts and is responsible for conferring strength and
resilience to cells (9). Elastin
is an essential part of various human tissues that depend on
elasticity, including the skin, lung and arteries. Elastin provides
these elastic tissues with the ability to stretch and recoil, and
it plays a critical role in supporting and maintaining healthy
cells (10).
Mycosporine-like amino acids (MAAs) possess
significant chemoprotective effects against photo-induced skin
senescence (11). MAAs found in
and isolated from a number of marine organisms, such as
cyanobacteria, algae, and heterotrophic bacteria, have attracted a
great deal of interest, especially for potential UV protection. In
a recent study, it was suggested that MAAs have antioxidant
properties and UV absorbance activity (12). An important MAA is porphyra-334,
which has been reported to act mainly in photoprotection, but it
also posseses antioxidation abilities. Results of a recent study
have shown that algae extracts prevent UV-induced photodamage in
human keratinocytes (13).
Although the photo-protective effect of MAAs in algae and tissue
has been reported (14,15), little is known regarding its
effects on the aging process of skin cells. This study investigated
whether supplementation with porphyra-334, an active MAA from
Porphyra (P.) yezoensis, inhibits UVA-induced cellular
senescence in human skin fibroblasts.
Materials and methods
Extraction and isolation of water-soluble
porphyra-334
The porphyra-334 extraction method was performed as
previously described (16) with
minor modifications. Briefly, dried P. yezoensis (100 g) was
ground and extracted in hydrophilic solvent consisting of 80%
aqueous methanol (v/v) at 45°C for 2 h. The extract was filtered
(no. 3, 90 mm; Advantec, Tokyo, Japan) to remove powder particles,
and the residual aqueous suspension was evaporated to dryness under
vacuum at 41°C (EYELA N-1100; Tokyo Rikakikai Co., Ltd., Nihonbashi
Honcho, Japan). The dried extract was dissolved in 150 ml ultrapure
water and transferred to a separating funnel containing 666 ml
chloroform-methanol-ultrapure water (2:1:1, v/v/v). The upper layer
containing crude MAAs was collected. The water layer was then
filtered through 0.2-μm pore-sized syringe filters (Woongki
Science, Seoul, Korea) and loaded onto a Strata C18-E cartridge
(Phenomenex, Inc., Torrance, CA, USA) (previously equilibrated with
ultrapure water) for analysis.
High-performance liquid chromatography
(HPLC) analysis
Porphyra-334 was purified using an Agilent 1100
series HPLC system equipped with a diode array detector (DAD;
Agilent Technologies, Inc., Palo Alto, CA, USA). The HPLC
conditions used were: column, Gemini-NX 5 μ C18 (i.d. 250x21.20 mm;
Phenomenex); column temperature, RT; flow rate, 30 ml/min; mobile
phase, 0.1% acetic acid in H2O; and wavelength for
detection, 334 nm. Purified porphyra-334 was stored in the dark at
−70°C until analysis. Identification of MAAs was performed using UV
absorption spectra and mass spectrometry.
Electrospray ionization-mass spectrometry
(ESI-MS) analysis
To identify the peaks of fingerprints, the Agilent
1100 series (Agilent Technologies) ion-trap mass spectrometer with
an electrospray ionization (ESI) source was used for the HPLC/MS
method. ESI-MS conditions of each HPLC peak were set as follows:
scan mass range, m/z 100–600; fragmentor voltage, 70 V;
drying gas N2 flow rate, 12 l/min; sheath gas flow rate,
60 arbitrary units; drying gas temperature, 350°C; and capillary
voltage, 3,000 V.
Cell culture
Human skin fibroblasts (CCD-986sk) were obtained
from the American Type Culture Collection (ATCC, Manassas, VA,
USA). Cells were grown in Dulbecco’s modified Eagle’s medium (DMEM;
Gibco, Grand Island, NY, USA) containing 10% (v/v) fetal bovine
serum (FBS; HyClone, Logan, UT, USA) and 1% (v/v)
penicillin-streptomycin (Gibco) under a humidified atmosphere of 5%
CO2 at 37°C.
UVA irradiation and treatment
Prior to UV irradiation, cells were washed with PBS
and exposed to a radiation dose of 10 J/cm2 of UVA light
(BLX-254; Vilber Lourmat, Marne La Vallée, France) in PBS.
Subsequent to irradiation, the treated cells were washed with PBS
and replaced with different concentrations of porphyra-334 for 24
h. Concomitantly, no irradiation control cells were treated in the
same manner, although the wells were covered with aluminum foil to
prevent irradiation.
Cell viability
Cell viability was determined using a
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxy-phenyl)-2-
(4-sulfophenyl)-2H-tetrazolium (MTS) assay (Promega Corp., Madison,
WI, USA) according to the manufacturer’s instructions. Briefly,
2x104 cells/well were plated and allowed to attach to
96-well plates. The cells were then exposed to serial
concentrations of porphyra-334 for 24 h. After 24 h, a soluble MTS
reagent was added and absorbance of the formazan was measured
directly in 96-well plates at 490 nm using a multi-plate reader
(SpectraMAX 340PC; Molecular Devices, Sunnyvale, CA, USA). Relative
cell viability was calculated as the percent viability relative to
the untreated control cells. Each experiment was performed in
triplicate.
Senescence-associated β-galactosidase
(SA-β-gal) staining
SA-β-gal activity was determined at 24 h after UVA
irradiation. A cellular senescence assay kit (Cell Biolabs, Inc.,
San Diego, CA, USA) was performed according to the manufacturer’s
instructions. Briefly, the cells were washed twice in PBS and
incubated at room temperature for 5 min with fixing solution. The
cells were washed three times with PBS, the final wash was
aspirated, and the cells were completely covered with freshly
prepared cell staining working solution. The cells were then
incubated in the dark overnight at 37°C. Following removal of the
cell staining solution, the cells were washed twice with PBS and
blue-stained senescence cells were observed using a light
microscope (Olympus Microscope System IX51; Olympus, Tokyo,
Japan).
Intracellular ROS production
The production of intracellular ROS was measured
using the redox-sensitive fluorescent dye 2′-7′-dichlorofluorescein
diacetate [DCF-DA
(C24H14Cl2O7);
Sigma-Aldrich, Inc., St. Louis, MO, USA]. The ability of cells to
produce ROS was measured by fluorescence. The cells were treated
with 25 μM DCF-DA for 30 min at 37°C and washed twice in PBS.
Representative images were obtained using a fluorescence microscope
(Olympus Microscope System IX51; Olympus).
Elastase activity
Elastase activity using the synthetic substrate
N-Succinyl-Ala-Ala-Ala-p-nitroanilide (STANA; Sigma-Aldrich) was
measured as previously described (17). Briefly, 100 μl of enzyme solution
was dispensed into 96-well plates, which were pre-incubated for 15
min at 37°C. Following the addition of 2 μl 55.3 mM STANA, the
plates were further incubated for 1 h at 37°C. The release of
p-nitroaniline was measured by absorbance at 410 nm and enzymatic
activity was expressed as a percentage of total p-nitroaniline.
Total collagen
Total collagen synthesis in fibroblasts was measured
using the Procollagen Type I C-Peptide (PIP) EIA kit (Takara Bio
Inc., Otsu, Japan) according to the manufacturer’s instructions.
Briefly, 100 μl of antibody-POD conjugate solution were added to
appropriate wells, after which 20 μl of cell culture medium was
added to the wells within 5 min and incubated for 3 h at 37°C. The
contents were removed by suction and the wells were washed four
times with 300 μl of washing buffer. Substrate solution (100 μl)
was added to each well and incubated at room temperature for 15
min. Subsequently, 100 μl of stop solution was added to all the
wells, and absorbance was read at 450 nm.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA from each sample was extracted using
TRIzol reagent (Invitrogen, Carlsbad, CA, USA). According to the
manufacturer’s instructions, total RNA (1 μg) was subjected to
first strand cDNA synthesis using a Reverse Transcriptase PreMix
kit (Intron Biotechnology, Inc., Gyeonggi-do, Korea). PCR
amplification of the cDNA products was performed with 2X TOPsimple™
DyeMIX(aliquot)-nTaq (Enzynomics, Daejeon, Korea) and primer
pairs. Amplified products were separated by 1% agarose gel
electrophoresis and visualized with 1 mg/ml ethidium bromide. mRNA
levels were normalized using GAPDH as an internal control.
Western blot analysis
After treatment, cells were washed twice with PBS,
harvested and lysed in RIPA buffer [50 mM Tris (pH 7.4), 1 mM
ethylene glycol tetraacetic acid (EGTA), 150 mM NaCl, 1% Triton
X-100, 0.25% sodium deoxycholate]containing protease inhibitor
cocktail (Geno Technology, Inc., St. Louis, MO, USA). The lysates
were centrifuged at 13,475 x g for 15 min at 4°C (Smart-R17; Hanil
Science Industrial, Incheon, Korea). Supernatants were collected
and their protein concentrations were determined using a BCA
protein assay kit (Pierce Biotechnology, Inc., Rockford, IL, USA).
Equal amounts of protein (30 μg) were boiled for 10 min and
separated using 7.5–15% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE). The resolved proteins were then
transferred to polyvinylidene difluoride (PVDF) membranes
(Millipore Corp., Billerica, MA, USA). The membranes were blocked
by incubation with 1% bovine serum albumin (BSA) in TBS-T [10 mM
Tris-HCl, 150 mM NaCl (pH 7.5) containing 0.1% Tween-20] at room
temperature for 1 h and incubated with specific primary antibody
(Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) for 3 h. The
membranes were washed three times with TBS-T and incubated for 2 h
with the appropriate HRP-conjugated goat anti-rabbit, goat
anti-mouse or rabbit anti-goat secondary antibody (Santa Cruz
Biotechnology) diluted at 1:10,000 in TBS-T containing 1% BSA. The
respective proteins were detected with SuperSignal® West
Pico (Thermo Fisher Scientific, Inc., Rockford, IL, USA). Equal
protein loading was assessed by the detection of GAPDH levels.
Statistical analysis
The results were presented as means ± SEM from at
least three independent experiments. Data were analyzed using
one-way analysis of variance (ANOVA) with the Student’s t-test
using SPSS 10.0 (SPSS, Inc., Chicago, IL, USA). Differences were
considered significant at p<0.05.
Results
Identification of MAAs
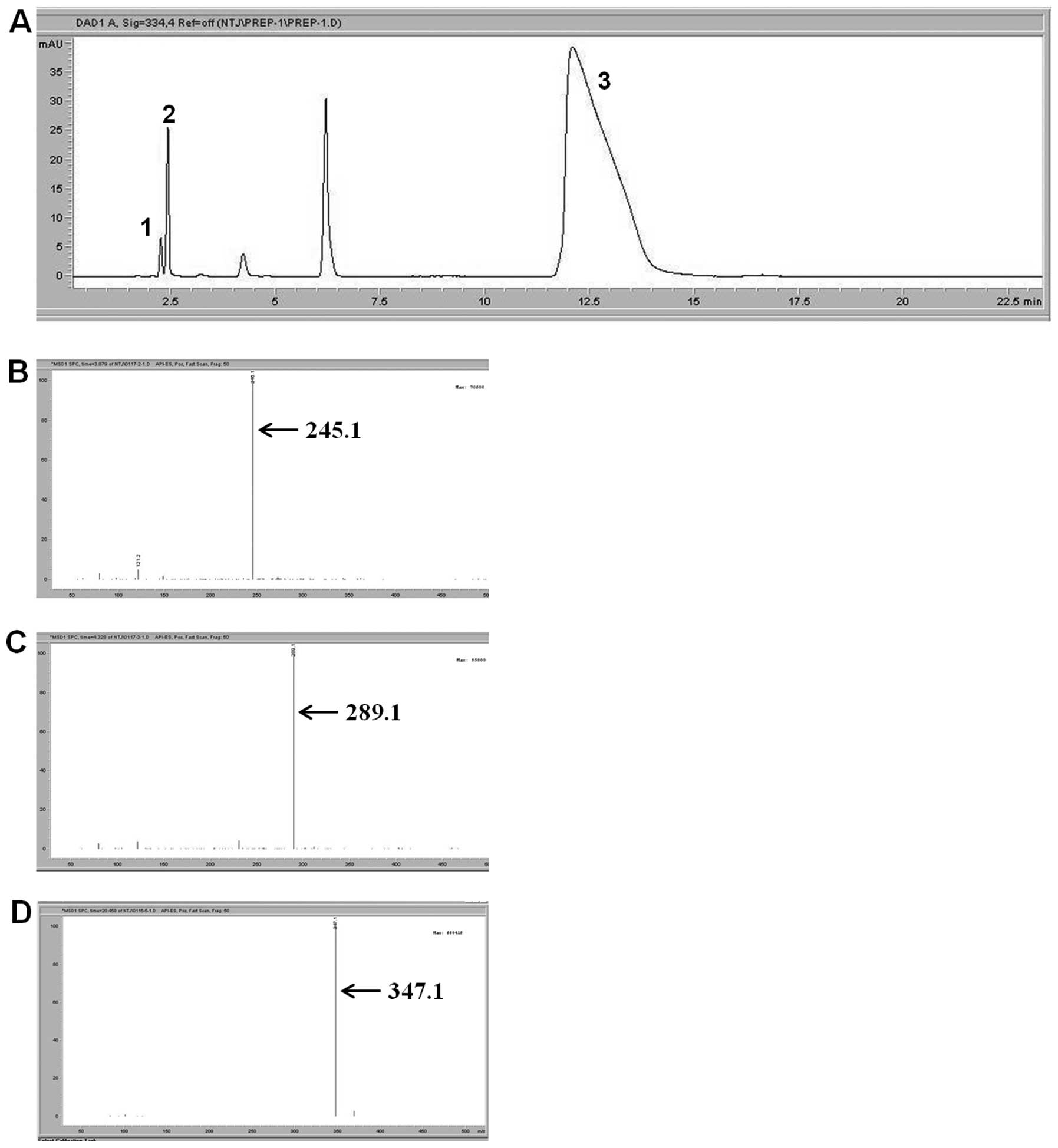
As shown in Fig.
1A, the HPLC analysis of P. yezoensis revealed various
absorption peaks at 334 nm. Peaks were identified tentatively by
comparing retention times with known standards (peak 1, 2.29 min;
peak 2, 2.493 min; and peak 3, 11.53 min). We then identified the
three peaks using LC/MS analysis. Peak 1, 2 and 3 showed
[M+H]+ ions at m/z 245.1, 289.1 and 347.1. Peak 1
was tentatively identified as palythine, peak 2 as asterina-330 and
peak 3 as porphyra-334. Porphyra-334 was the most abundant MAA in
P. yezoensis. Therefore, the following photoaging protection
experiments were conducted using porphyra-334.
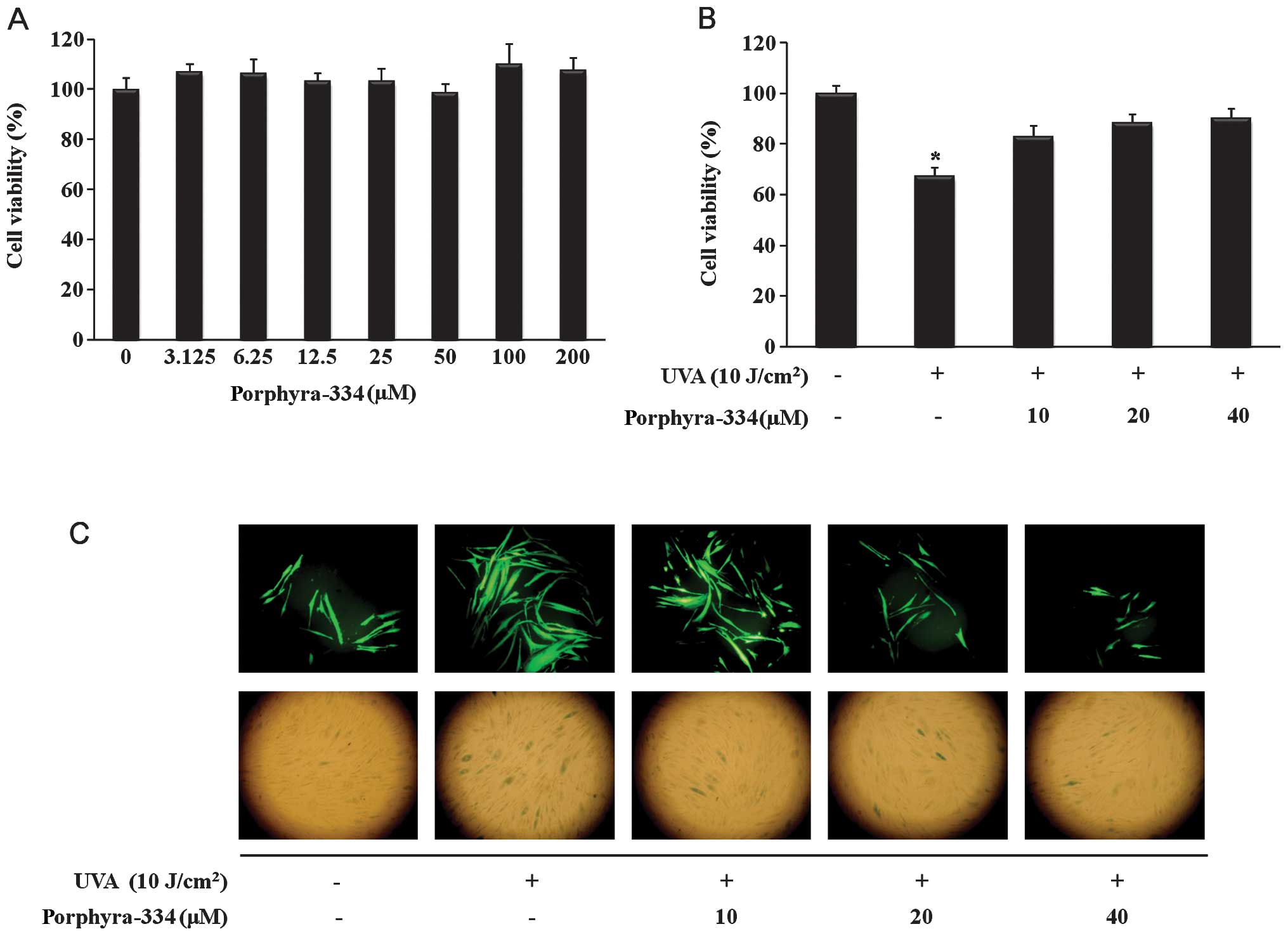
Effect of porphyra-334 on cell
viability
To determine the cytotoxic effect of porphyra-334 on
human skin fibroblasts, cell viability was measured using the MTS
assay. Human skin fibroblasts were incubated with or without
porphyra-334 at concentrations of 0–200 μM. As shown in Fig. 2A, no significant toxicity was
observed in the cells treated with porphyra-334 for 24 h. To
explore the protective effect of porphyra-334 on UVA-induced cell
damage, human skin fibroblasts (previously exposed to UVA
irradiation) were incubated with various concentrations of
porphyra-334 (0–40 μM). As shown in Fig. 2B, treatment with porphyra-334
protected against cell damage in a dose-dependent manner.
Effect of porphyra-334 on ROS
generation
To determine whether porphyra-334 functions as a
scavenger of UVA-induced ROS generation, intracellular ROS levels
were measured. As shown in Fig.
2C (upper panel), the level of ROS in UVA-irradiated cells
increased compared with non-irradiated cells. UVA-exposed cells
showed a significant reduction in DCF-DA staining in the presence
of porphyra-334, indicating that porphyra-334 inhibited the
intracellular accumulation of ROS in human skin fibroblasts damaged
by UVA-induced oxidant stress.
Effect of porphyra-334 on SA-β-gal
activity
SA-β-gal staining was performed to observe SA-β-gal
activity, which is a biomarker of senescence. Images clearly
indicated that the cells were induced to a senescence-like state by
UVA irradiation. The inhibitory activity of porphyra-334 was also
observed and found to effectively suppress the expression of
SA-β-gal in a dose-dependent manner (Fig. 2C lower panel).
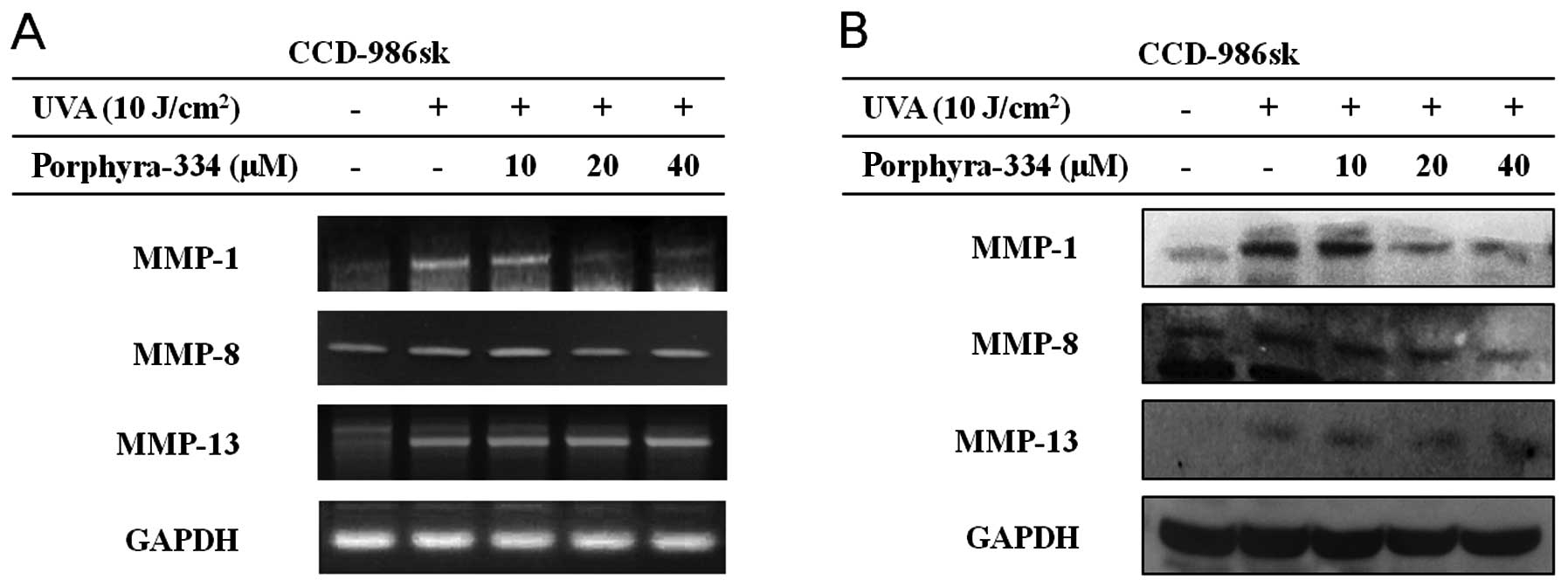
Effect of porphyra-334 on MMP
expression
To determine whether porphyra-334 inhibited MMP
expression induced by UVA irradiation, human skin fibroblasts were
irradiated with UVA (10 J/cm2) and treated with
porphyra-334 for 24 h. Based on RT-PCR, UVA irradiation
significantly increased MMP-1 mRNA expression in culture medium
(Fig. 3A). To investigate the
dose-dependent effect of porphyra-334, the cells were treated with
different concentrations of porphyra-334 ranging from 10 to 40 μM.
The highest concentration of porphyra-334 inhibited MMP-1 mRNA
expression in UVA irradiated human skin fibroblasts up to 56.2%,
and the inhibition was dose-dependent. The inhibition of MMP-8 was
similar to MMP-1, but not MMP-13. MMP-13 expression in the presence
of porphyra-334 was similar to the UVA-irradiation control.
Porphyra-334 reduced the elevated MMP expression, with the
exception of MMP-13, at the gene and protein levels compared with
the control groups, which were irradiated without treatment. Of the
three MMPs, MMP-1 was the most active compared with the positive
control. Thus, porphyra-334 exerted a protective effect on
UVA-induced collagen degradation via the negative regulation of MMP
expression.
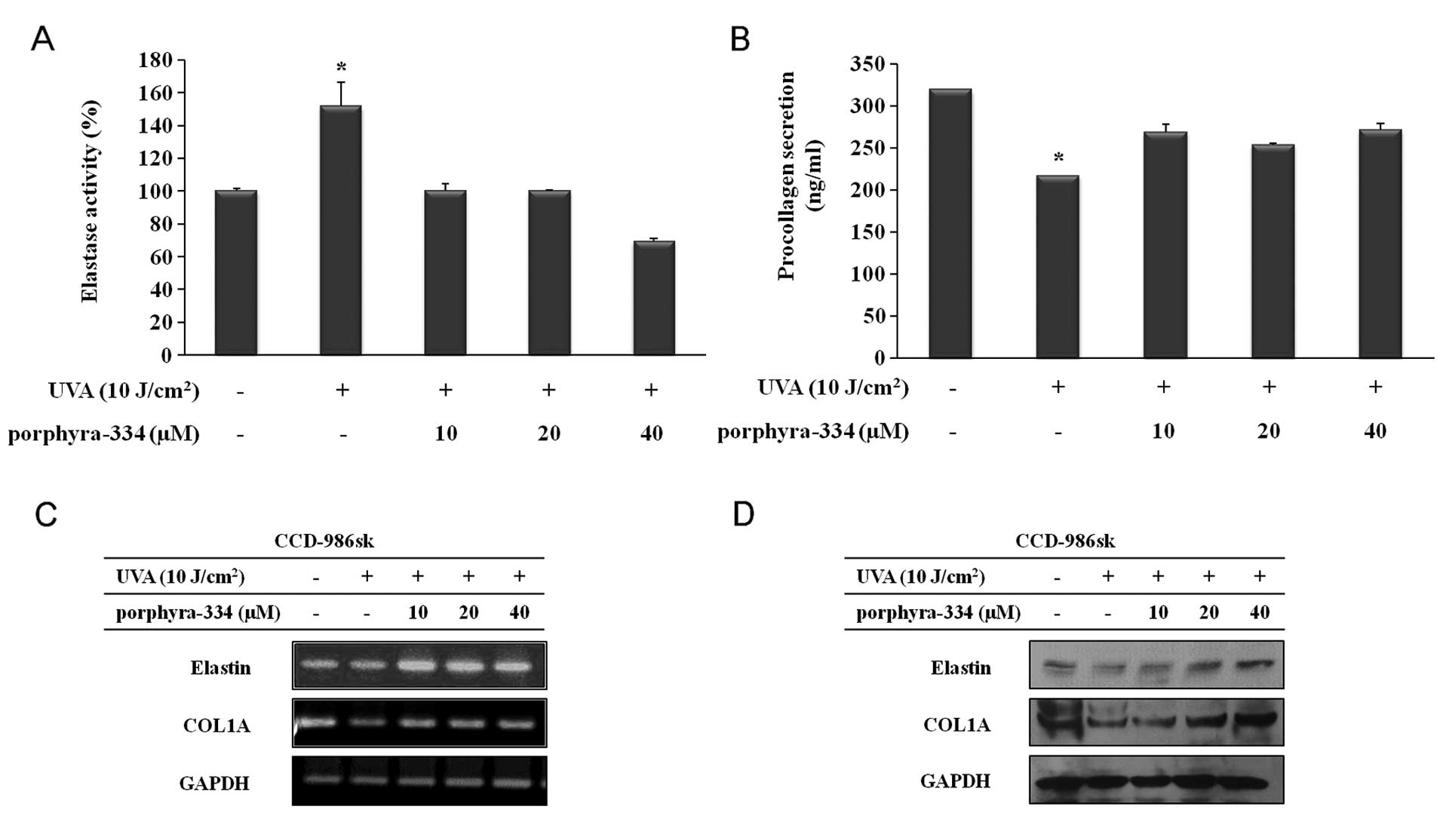
Effect of porphyra-334 on UVA-induced
intracellular procollagen
To evaluate the effect of porphyra-334 on collagen
synthesis, human skin fibroblasts were treated with various
concentrations of porphyra-334 (0–40 μM) for >24 h. The secreted
procollagen level was measured in the culture medium using an ELISA
assay, as described in the ‘Materials and methods’. As shown in
Fig. 4A, the collagen content was
319.5±0.069 ng/ml in the non-irradiated cells but 217.333±0.177
ng/ml in the UVA-irradiated cells. Procollagen secretion levels
increased by 269.167±9.090, 253.833±1.464 and 271.833±7.224 ng/ml
in the presence of porphyra-334 at concentrations of 10, 20 and 40
μM, respectively. Thus, porphyra-334 has a protective effect on
collagen degradation by enhancing collagen synthesis in
photodamaged human skin fibroblasts.
Effect of porphyra-334 on UVA-induced
elastase activity
UV irradiation is known to cause elastin degradation
by activating elastase (18), and
loss of skin elastin caused by UVA irradiation results in wrinkle
formation. In this study, the elastase activity of human skin
fibroblasts in response to porphyra-334 was confirmed based on the
release of p-nitroaniline. As shown in Fig. 4B, elastase activity increased by
51.9% in the supernatant and intracellular contents, respectively,
following UVA irradiation compared with the non-irradiated cells.
This increase in the elastase inhibitory effect was decreased by
porphyra-334 treatment at 10, 20 and 40 μM after UVA irradiation by
51.9, 51.9 and 82.5% compared with the UVA-irradiated cells,
respectively.
Effect of porphyra-334 on UVA-induced
collagen and elastin degradation
To examine the effect of porphyra-334 on collagen
and elastin degradation, human skin fibroblasts previously
stimulated with UVA irradiation were incubated with various
concentrations of porphyra-334 (0–40 μM). The expression of the
specific elastin and type I collagen at the mRNA and protein levels
was determined by RT-PCR and western blot analysis, respectively.
The mRNA and protein levels of type I collagen and elastin are
shown in Fig. 4C and D.
Expression of type I collagen was decreased in UVA-irradiated
cells, while the decrease in cellular collagen levels following UVA
exposure was prevented in a dose-dependent manner in the presence
of porphyra-334. Similarly, the expression of elastin was enhanced
following porphyra-334 treatment after UVA irradiation. Results of
the western blot analysis were in concordance with those of RT-PCR.
These results indicated that porphyra-334 may be involved in
collagen synthesis by regulating collagen-degrading MMP expression
and elastinase activity.
Discussion
Porphyra sp. has been used to protect against
a variety of diseases. However, the intracellular signaling and
protective effects of porphyra-334 against UVA-induced photodamage
in human skin fibroblasts remains poorly understood. Numerous
studies (19–21) have explored the impact of UV
irradiation on skin cells, but the beneficial effect of
photoprotective agents on UVA-induced damage in human skin is not
well characterized. UVA is a potent inducer of various ROS, and it
causes lipid peroxidation in cell membranes. Antioxidant defense
mechanisms may be overwhelmed by excessive free radical generation,
which damages the cells and increases the chances of
photocarcinogenesis. Development of novel antioxidant strategies to
supplement the natural defense mechanism of the skin may be an
important strategy to reduce UV-induced effects. This study showed
that porphyra-334, a rich source of MAA derived from P.
yezoensis, is capable of reducing the adverse effects of
UVA-mediated cutaneous damage.
Photoaging associated with UV irradiation is thought
to play a central role in initiating and driving the signaling
events that lead to cell responses. UV radiation of skin decreases
antioxidant enzyme concentrations (22) and increases hydrogen peroxides and
other ROS (23,24). UV irradiation initiates the
generation of ROS and alters gene and protein structure and
function, leading to skin damage. Porphyra-334 was shown to
dose-dependently decrease intracellular UVA-induced ROS generation
in human skin fibroblasts based on a modified DCF-DA fluorescence
assay. Since the generation of MMPs is significantly induced by ROS
(25), we showed that the ROS
scavenging of porphyra-334 was comparable to the non-UV irradiated
control. This suggests that porphyra-334 controls the expression of
MMPs by scavenging excess ROS in damaged skin fibroblasts.
UV irradiation enhances the decomposition of skin
connective tissue by activating MMPs responsible for the
degradation of skin collagen and inhibiting collagen synthesis of
ECM in connective tissues (26).
In this study, we observed the upregulation of MMPs (especially
MMP-1) and the degradation of dermal collagen following UV
irradiation. Porphyra-334 was a potent suppressor of UVA-induced
MMP generation, and it also inhibited the MMP-1-initiated
degradation of type I collagen. Therefore, inhibition of
collagenase MMP expression or activation of collagen synthesis may
be an effective strategy to prevent wrinkle formation following UVA
irradiation. This mechanism predicts that a free radical scavenger
may prevent UV-induced dermal damage by inhibiting MMP induction.
These findings suggest that the regulation of MMPs and type I
collagen levels in UV-irradiated fibroblasts may be related to the
inhibition of ROS generation by porphyra-334. MAAs from red algae
are known to effectively decrease MMPs and increase type I collagen
expression levels to prevent premature skin aging (27). Results of this study suggest that
porphyra-334 increases procollagen production by suppressing MMP
gene expression and reducing MMP production, which can reduce
cytokine secretion by human skin fibroblasts. Many algae have been
screened for potential MMP inhibitors. For example, Corallina
pilulifera (28), Ecklonia
cava (29), Laurencia
undulata (30) and
Amphiroa dilatata (31)
exhibited inhibitory effects on MMP activities. In this study,
porphyra-334 was safe to human skin fibroblasts and significantly
decreased the expression of MMPs induced by UVA irradiation.
Wrinkle formation in the skin is closely associated
with the degradation of ECM, and UV irradiation is known to induce
the degradation of ECM (32). UV
irradiation enhances collagenase activity and contributes to
wrinkle formation through the degradation of collagen in the dermal
ECM (33,34). Therefore, collagenase inhibitors
have been identified as potential therapeutic agents that protect
against photoaging and wrinkle formation (35). Collagen is the main component of
the ECM of dermal connective tissue, and its concentration
decreases with photoaging. Once collagen is initially cleaved by
MMP-1, MMP-13 and other MMPs, collagen breakdown is further
promoted. The enzyme mainly responsible for collagen breakdown in
skin is MMP-1, which cleaves types I, III, VII, VIII and X
collagen. As reported previously with increasing photoaging, MMP-1
levels increase and collagen synthesis decreases in sun-protected
human skin in vivo (36).
In this study, the exposure of human skin fibroblasts to UVA
significantly decreased type I collagen levels and increased MMP
secretion, both of which were reversed by porphyra-334. Elastin is
important in the dermis (37),
and UV exposure has been shown to cause elastin degradation by
activating elastase (38). As
shown in Fig. 4B, the inhibitory
effect of porphyra-334 on elastase activity was significant at a
concentration of ≥10 μM.
Overall, porphyra-334 of P. yezoensis
significantly inhibits ROS production, reduces MMP expression, and
induces type I collagen, elastin at the mRNA and protein levels in
a dose-dependent manner. These data suggest that porphyra-334 is a
potential candidate for the prevention and treatment of skin
photoaging. Additionally, porphyra-334 can be used to characterize
the signal transduction pathways and molecular mechanisms involved
in the anti-photoaging process.
Acknowledgements
This study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education (2012R1A6A1028677).
References
|
1
|
Ichihashi M, Ueda M, Budiyanto A, et al:
UV-induced skin damage. Toxicology. 189:21–39. 2003. View Article : Google Scholar
|
|
2
|
Jaiswal AK: Nrf2 signaling in coordinated
activation of antioxidant gene expression. Free Radic Biol Med.
36:1199–1207. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Marchioli R, Schweiger G, Tavazzi L and
Valagussa F: Antioxidant vitamins and prevention of cardiovascular
disease: epidemiological and clinical trial data. Lipids.
36:S53–S63. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chin YW, Balunase MJ, Chai HB and Kinghorn
AD: Drug discovery from natural sources. AAPS J. 8:E239–E253.
2006.PubMed/NCBI
|
|
5
|
Cho S, Lee MJ, Kim MS, et al: Infrared
plus visible light and heat from natural sunlight participate in
the expression of MMPs and type I procollagen as well as
infiltration of inflammatory cell in human skin in vivo. J Dermatol
Sci. 50:123–133. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gogly B, Ferré FC, Cherifi H, Naveau A and
Fournier BP: Inhibition of elastin and collagen networks
degradation in human skin by gingival fibroblast. In vitro, ex vivo
and in vivo studies. J Cosmet Dermatol Sci Appl. 1:4–14. 2011.
|
|
7
|
Zhang M, Dang L, Guo F, Wang X, Zhao W and
Zhao R: Coenzyme Q10 enhances dermal elastin expression, inhibits
IL-1α production and melanin synthesis in vitro. Int J Cosmet Sci.
34:273–279. 2012.PubMed/NCBI
|
|
8
|
Chiang HM, Chen HC, Lin TJ, Shih IC and
Wen KC: Michelia alba extract attenuates UVB-induced
expression of matrix metalloproteinases via MAP kinase pathway in
human dermal fibroblasts. Food Chem Toxicol. 50:4260–4269. 2012.
View Article : Google Scholar
|
|
9
|
Gelse K, Pöschl E and Aigner T:
Collagens-structure, function, and biosynthesis. Adv Drug Delivery
Rev. 55:1531–1546. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Almine JF, Bax DV, Mithieux SM, et al:
Elastin-based materials. Chem Soc Rev. 39:3371–3379. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yuan YV and Walsh NA: Antioxidant and
antiproliferative activities of extracts from a variety of edible
seaweeds. Food Chem Toxicol. 44:1144–1150. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Andreguetti D, Stein EM, Pereira CM, Pinto
E and Colepicolo P: Antioxidant properties and UV absorbance
pattern of mycosporine-like amino acids analogs synthesized in an
environmentally friendly manner. J Biochem Mol Toxicol. 27:305–312.
2013. View Article : Google Scholar
|
|
13
|
Piao MJ, Hyun YJ, Cho SJ, et al: An
ethanol extract derived from Bonnemaisonia hamifera
scavenges ultraviolet B (UVB) radiation-induced reactive oxygen
species and attenuates UVB-induced cell damage in human
keratinocytes. Mar Drugs. 10:2826–2845. 2012.
|
|
14
|
Sommaruga R, Whitehead K, Shick JM and
Lobban CS: Mycosporine-like amino acids in the zooxanthella-ciliate
symbiosis Maristentor dinoferus. Protist. 157:185–191. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rastogi RP and Incharoensakdi A: UV
radiation-induced accumulation of photoprotective compounds in the
green alga Tetraspora sp. CU2551. Plant Physiol Bioch.
70:7–13. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tao C, Sugawara T, Maeda S, Wang X and
Hirata T: Antioxidative activities of a mycosporine-like amino
acid, porphyra-334. Fish Sci. 74:1166–1172. 2008. View Article : Google Scholar
|
|
17
|
Nakagawa K, Tsuji T, Kadoya A and Hamada
T: Elastase-like enzyme activity in cultured human fibroblast. Skin
Res. 29:793–797. 1987.
|
|
18
|
Seite S, Zucchi H, Septier D,
Igondjo-Tchen S, Senni K and Godeau G: Elastin changes during
chronological and photo-ageing: the important role of lysozyme. J
Eur Acad Dermatol Venereol. 20:980–987. 2006.PubMed/NCBI
|
|
19
|
Tournier C, Hess P, Yang DD, et al:
Requirement of JNK for stress-induced activation of the cytochrome
c-mediated death pathway. Science. 288:870–874. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kulms D and Schwarz T: Molecular
mechanisms involved in UV-induced apoptotic cell death. Skin
Pharmacol Physiol. 15:342–347. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Šitum M, Buljan M, Bulat V, Lugović Mihić
L, Bolanča Ž and Šimić D: The role of UV radiation in the
development of basal cell carcinoma. Coll Antropol. 32:167–170.
2008.PubMed/NCBI
|
|
22
|
Yamamoto Y: Role of active oxygen species
and antioxidants in photoaging. J Dermatol Sci. 27(Suppl 1): S1–S4.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Masaki H, Atsumi T and Sakurai H:
Detection of hydrogen peroxide and hydroxyl radicals in murine skin
fibroblasts under UVB irradiation. Biochem Biophys Res Commun.
206:474–479. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yasui H and Sakurai H: Chemiluminescent
detection and imaging of reactive oxygen species in live mouse skin
exposed to UVA. Biochem Biophys Res Commun. 269:131–136. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nalson KK and Melendez JA: Mitochondrial
redox control of matrix metalloproteinase. Free Radical Biol Med.
37:768–784. 2004. View Article : Google Scholar
|
|
26
|
Wlaschek M, Heinen G, Poswig A, Schwarz A,
Krieg T and Scharffetter-Kochanek K: UVA-induced autocrine
stimulation of fibroblast-derived collagenase/MMP-1 by interrelated
loops of interleukin-1 and interleukin-6. J Photochem Photobiol.
59:550–556. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pallela R, Yoon NY and Kim SK:
Anti-photoaging and photoprotective compounds derived from marine
organisms. Mar Drugs. 8:1189–1202. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ryu B, Qian ZJ, Kim MM, Nam KW and Kim SK:
Anti-photoaging activity and inhibition of matrix metalloproteinase
(MMP) by marine red alga, Corallina pilulifera methanol
extract. Radiat Phys Chem. 78:98–105. 2009. View Article : Google Scholar
|
|
29
|
Zhang C, Li Y, Shi X and Kim SK:
Inhibition of the expression on MMP-2, 9 and morphological changes
via human fibrosarcoma cell line by 6,6′-bieckol from marine alga
Ecklonia cava. BMB Rep. 43:62–68. 2010.PubMed/NCBI
|
|
30
|
Li YX, Li Y, Lee SH, Qian ZJ and Kim SK:
Inhibitors of oxidation and matrix metalloproteinases, floridoside,
and D-isofloridoside from marine red alga Laurencia
undulata. J Agric Food Chem. 58:578–586. 2009.
|
|
31
|
Khan SB, Kong CS, Kim JA and Kim SK:
Protective effect of Amphiroa dilatata on ROS induced
oxidative damage and MMP expressions in HT1080 cells. Biotechnol
Bioproc Eng. 15:191–198. 2010.
|
|
32
|
Rittié L and Fisher GJ: UV-light-induced
signal cascades and skin aging. Ageing Res Rev. 1:705–720.
2002.PubMed/NCBI
|
|
33
|
Uitto J: The role of elastin and collagen
in cutaneous aging: intrinsic aging versus photoexposure. J Drugs
Dermatol. 7:S12–S16. 2008.PubMed/NCBI
|
|
34
|
Uitto J, Fazio MJ and Olsen DR: Molecular
mechanisms of cutaneous aging: age-associated connective tissue
alterations in the dermis. J Am Acad Dermatol. 21:614–622. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Inomata S, Matsunaga Y, Amano S, et al:
Possible involvement of gelatinases in basement membrane damage and
wrinkle formation in chronically ultraviolet B-exposed hairless
mouse. J Invest Dermatol. 120:128–134. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chung JH, Seo JY, Choi HR, et al:
Modulation of skin collagen metabolism in aged and photoaged human
skin in vivo. J Invest Dermatol. 117:1218–1224. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Labat-Robert L and Robert L: Aging of the
extracellular matrix and its pathology. Exp Gerontol. 23:5–18.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Getie M, Schmelzer CE and Neubert RH:
Characterization of peptides results from digestion of human skin
elastin with elastase. Proteins. 61:649–657. 2005. View Article : Google Scholar : PubMed/NCBI
|