Introduction
Antigen-specific TCR gene transfer enables the
instantaneous generation of defined T cell immunity and TCR
gene-modified T cells are fully functional in vitro and in
murine models (1–6). The retroviral transfer of a
MART1-specific TCR for adoptive T cell therapy first used in
clinical trials on melanoma patients demonstrated the feasibility
of TCR gene therapy (7,8). However, several factors currently
limit the efficacy and hamper the application of TCR gene
immunotherapy. One of these critical issues is that the introduced
TCRα and β chain can potentially assemble with endogenous TCR
chains (i.e., TCR mispairing ), which not only reduces the
expression of the desired TCR pair, but can create a new TCR with
unknown specificity that can potentially cause autoimmunity and
off-target toxicity (9,10).
A number of strategies have been investigated to
prevent the mixed TCR dimer formation. Examples of such strategies
are the replacement of the C domains of the TCRα and β chains by
the corresponding murine domains (11,12), introduction of an additional
inter-chain disulfide bond between the constant domains of TCRα and
β chains (13,14), the inversion of amino acid
residues in the constant region of the TCRα and β chains that form
the TCR interface (15), and the
use of single-chain TCR (scTCR) chimeras, including three-domain
TCRs that contain other signaling domains, such as CD3ζ or FcɛRIγ
(VαVβCβCD3ζ) (16–18). However, the full extent to which
these strategies can prevent TCR chain mispairing is unclear.
In a recent study, we have isolated a TCR (Vα12 and
Vβ7) from tumor infiltrating lymphocytes (TILs) of patients
(HLA-A2+ and AFP+) with hepatocellular
carcinoma (19). We demonstrated
that T cells derived from central memory cells modified by
tumor-specific TCR gene transfer were more effective than T cells
derived from CD8+ T cells modified by TCR gene transfer
in inducing CTL activity and effector cytokine secretion. However,
wild-type (wt)TCRα12 and β7 expression in transduced human T cells
was lower than the levels of endogenous TCR chains, mispairing with
the endogenous TCR.
In the present study, we generated a chimeric
TCRαζβζ which was modified by fusing the original constant domains
downstream of the extracellular cysteine of wtTCRα and β chains to
complete human CD3ζ. Subsequently, we constructed genetically
encoded reporters coupled with a pair of fluorescent proteins to
monitor the expression of TCRαζβζ and pairing between TCRαζ and
TCRβζ using confocal laser scanning microscopy (CLSM) in living
cells (Jurkat and BEL-7402 cells). We demonstrate that these
reporters provide accurate images of TCRαζβζ expression and
pairing. Of note, we observed that the expression of TCRαζβζ was
markedly stronger and with evident microclusters accumulated at the
plasma membrane compared to wtTCRαβ. Fluorescence resonance energy
transfer (FRET) imaging analysis of the T cells and non-T cells
revealed that the expression of TCRαζβζ does not need the
engagement of CD3 subunits. Using these reporters, we demonstrate
that in addition to enhanced pairing, TCRαζβζ expression is greater
in T cells or non-T cells without CD3 subunits. Taken together,
these results suggest the reporters we used are usseful in studying
the expression/pairing of TCRαζβζ, which may be an effective
approach to preventing mispairing in TCR gene therapy.
Materials and methods
Cells, genes and reagents
Jurkat/E6-1 (ATCC TIB-152) cells were cultured in
RPMI-1640 medium (Gibco/Invitrogen, Carlsbad, CA, USA), human
embryonic kidney cells (HEK293) and human hepatocellular carcinoma
cells (BEL-7402; maintained in our laboratory) were cultured in
Dulbecco’s modified Eagle’s medium (DMEM) (Gibco/Invitrogen). All
cultures were supplemented with 10% fetal bovine serum (FBS;
Gibco/Invitrogen), 100 U/ml streptomycin and 100 U/ml penicillin.
The TCRα12 and β7 chains were isolated from TILs of patients
(HLA-A2+ and AFP+) with hepatocellular
carcinoma by our laboratory as previously described (19). CD3ζ chains were isolated from
peripheral blood mononuclear cells (PBMCs) of a healthy donor
(GenBank accession no. NM_000734.3). The pair of fluorescent
proteins [enhanced cyan fluorescent protein (ECFP)/enhanced yellow
fluorescent protein (EYFP)] was kindly provided by Dr G. Zhang
(Guangzhou University of Chinese Medicine, Guangzhou, China).
Monoclonal antibodies (mAbs) used for flow cytometry included
FITC-conjugated anti-TCRVα12.1 mAb (Pierce Biotechnology, Rockford,
IL, USA) and PE-conjugated anti-TCRVβ7.1 (Beckman Coulter, Brea,
CA, USA).
Splice overlap extension (SOE) PCR
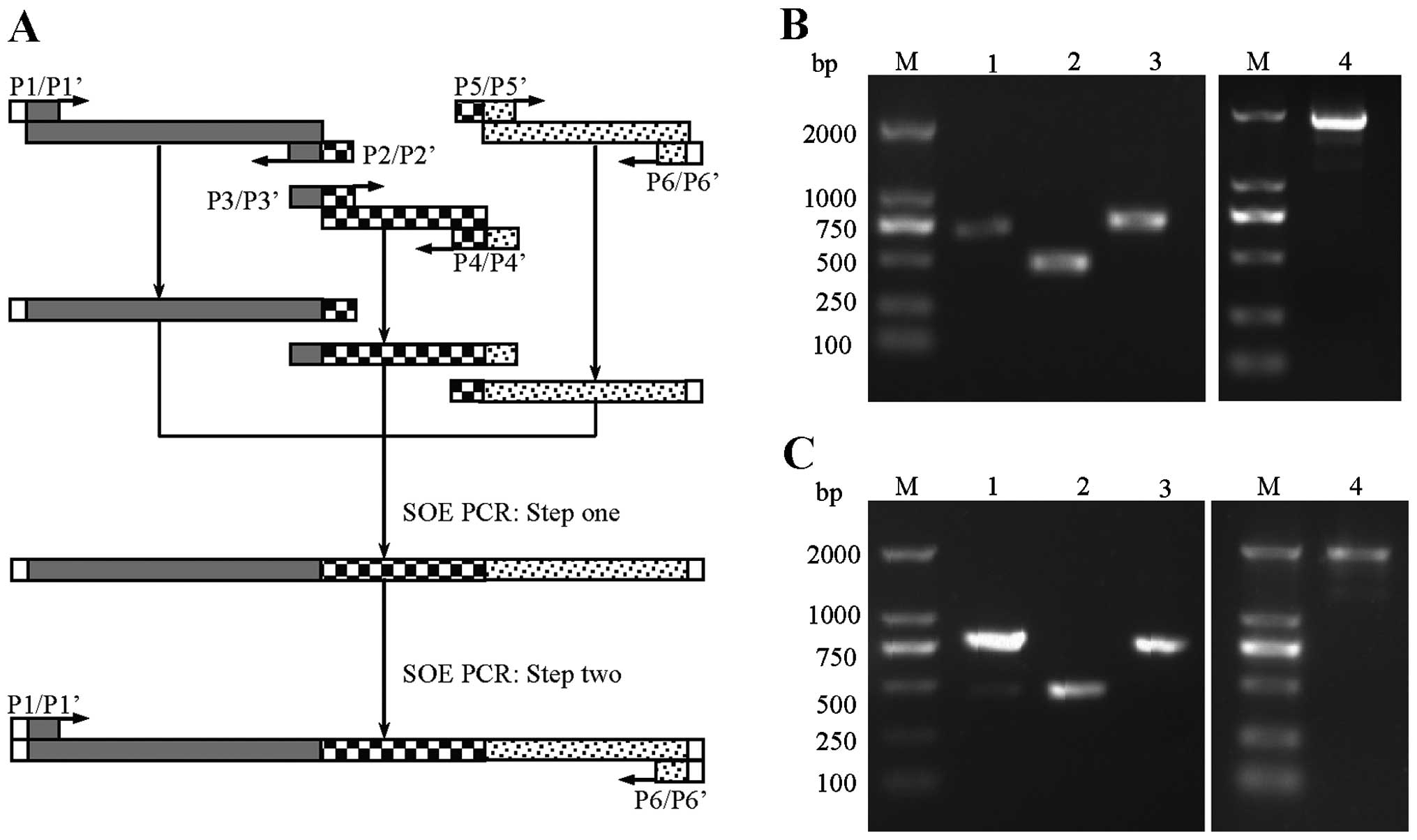
The strategy for the tripartite DNA fragment fusion
by SOE PCR is shown in Fig. 2A.
Variable fragments for generating TCRαζ-ECFP (including TCRα,
CD3ζ-a and EYFP) and TCRβζ-EYFP (including TCRβ, CD3ζ-b and ECFP)
were amplified using a set of forward primers and reverse primers.
The first step of SOE PCR reactions was performed with 100 ng (3
fragments were mixed with an equimolar ratio) of template without
primers, 10X Buffer, 2 mM dNTPs, 25 mM MgSO4, 0.5 U
KOD-Plus-Neo Polymerase in a 25 μl reaction volume. The PCR cycling
conditions were as follows: initial denaturation at 94°C for 2 min,
followed by 5 cycles at 94°C for 30 sec, at 57°C for 30 sec and at
68°C for 1.5 min and completed with a final extension at 68°C for 7
min. This initial PCRs generate overlapping gene segments that are
then used as template DNA for the second step of SOE PCR to create
a full-length product. Therefore, another 25 μl reaction mixture
(containing 10X Buffer, 2 mM dNTPs, 25 mM MgSO4, 0.5
KOD-Plus-Neo Polymerase and 1 μM forward primer P1/P1′ and reverse
primer P6/P6′) was added to the first reaction mixture for the
second step of SOE PCR. The reaction conditions were initial
denaturation at 94°C for 2 min, followed by 30 cycles at 94°C for
30 sec, at 62°C for 30 sec, and at 68°C for 1.5 min and a final
extension at 68°C for 7 min, 2 μl of the amplicons were analyzed by
agarose gel electrophoresis (1.0%) (Fig. 2B and C). Primer sequences for the
amplification of variable regions and fusion chains are presented
in Table I. The wtTCRα-ECFP and
wtTCRβ-EYFP fusing sequences were generated using the same
methods.
 | Table IPrimers for amplifying 6 gene
fragments (regular PCR and SOE PCR). |
Table I
Primers for amplifying 6 gene
fragments (regular PCR and SOE PCR).
| Fragments | Primers | Sequences
(5′→3′) |
|---|
| TCRα | P1 | ACGCCACAACCTTGGCCACCATGATATCCTTGAGAGTT |
| P2 |
CAGCAGGCCAAAGCTCTGTGGGCTGGGGAAGAAGGTGT |
| CD3ζ-a | P3 |
ACACCTTCTTCCCCAGCCCACAGAGCTTTGGCCTGCTG |
| P4 |
TCGCCCTTGCTCACCATGCGAGGGGGCAGGGCCTG |
| ECFP | P5 |
CAGGCCCTGCCCCCTCGCATGGTGAGCAAGGGCG |
| P6 | AGTGCGGCCGCTTACTTGTACAGCTCGTCCAT |
| TCRβ | P1′ | ATAGCTAGCGCCACCATGGGCTGCAGGCTGCTCTG |
| P2′ |
ATCCAGCAGGCCAAAGCTCTGTGCTCTACCCCAGGC |
| CD3ζ-b | P3′ |
GCCTGGGGTAGAGCACAGAGCTTTGGCCTGCTGGAT |
| P4′ |
TCGCCCTTGCTCACCATGCGAGGGGGCAGGGCCTG |
| EYFP | P5′ |
CAGGCCCTGCCCCCTCGCATGGTGAGCAAGGGCG |
| P6′ | CGGCGTCGACTTACTTGTACAGCTCGTC |
Vector construction
To construct a bicistronic vector expressing both
the TCRαζ-ECFP and TCRβζ-EYFP chains, the fusion sequences
TCRαζ-ECFP and TCRβζ-EYFP were cloned into the original pIRES2-EGFP
vector (Clontech Laboratories, Inc., Palo Alto, CA, USA). The
TCRαζ-ECFP fusion gene was inserted into the
BstXI/NotI restriction site of the pIRES2-EGFP vector
(Clontech Laboratories), located downstream of an internal
ribosomal entry site (IRES) sequence of the plasmid to replace the
EGFP gene, then the TRBζ-EYFP fusion gene was inserted into the
NheI/SalI restriction site, located upstream of IRES.
The other bicistronic vector expressing both non-modified
wtTCRα-ECFP and wtTCRβ-EYFP was constructed in a manner similar to
the above-mentioned procedure. The single vectors expressing
TCRα-ECFP or TCRβ-EYFP which were used to remove the spectral
bleed-through (SBT) contamination in the FRET images were
constructed as previously described (20). The identity of all constructs was
verified by direct sequencing.
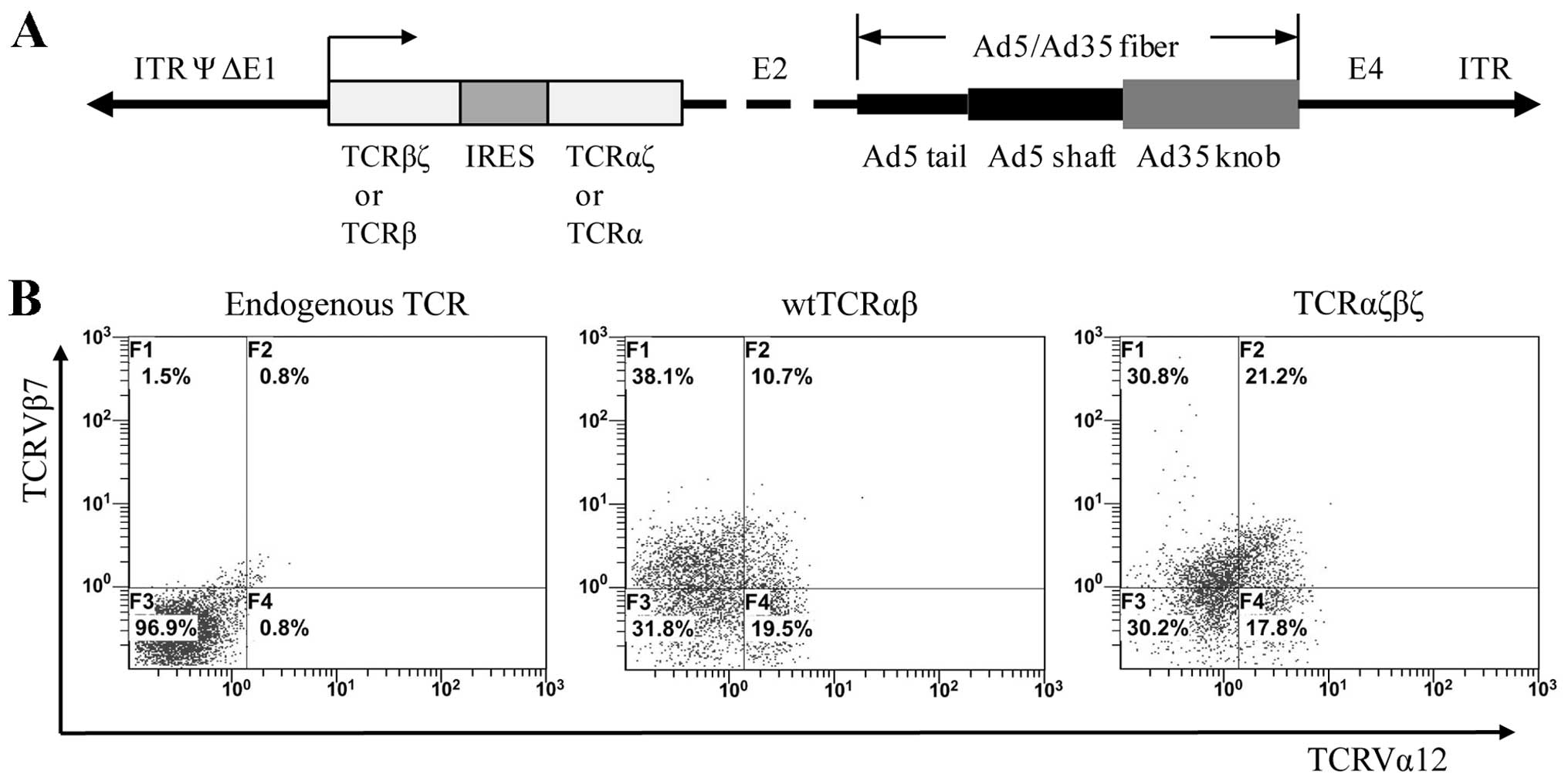
Production of adenoviral particles and
the transduction of TCR gene into Jurkat cells
We used the Ad5F35 chimeric adenoviral vector
(21) which contained the Ad35
fiber knob incorporated into an Ad5 capsid as the packaging vector,
and the whole wtTCRαβ cassette (TCRβ-IRES-TCRα) and TCRαζβζ
cassette (TCRβζ-IRES-TCRαζ) was cloned into the shuttle plasmid
(pDC315) (Fig. 4A). Adenoviral
particles were produced by co-transfection of the packaging cells,
HEK293, with Ad5F35 vector and recombinant pDC315, containing
wtTCRαβ or TCRαζβζ. The virus-containing supernatants were filtered
through 0.45-μm filters. Adenoviral particle titers were determind
by the 50% tissue culture infectious dose (TCID50) method and the
supernatants were directly used for the infection of the target
Jurkat T cells. For Jurkat T cell transduction, 6-well plates were
seeded with Jurkat cells at 1×106 cells/well in 2 ml of
RPMI 1640 medium supplemented with 2% FBS, cultured for 35 min, and
then transduced with MOI 100 PFU of wtTCRαβ or TCRαζβζ viral
supernatants. Following culture for 24 h at 37°C with 5%
CO2, the culture medium was replaced with RPMI 1640
medium supplemented with 10% FBS. After a further 48-h culture
period, the cells were analyzed for TCR expression by flow
cytometry.
Flow cytometry
The TCR-transduced Jurkat T cells (5×105)
were analyzed for transgene expression by flow cytometry using
FITC-conjugated anti-TCRVα12 mAb and PE-conjugated anti-TCRVβ7
mAbs. The cells were incubated with the mAbs on ice for 30 min.
Subsequently, the Jurkat cells were washed twice with PBS and
centrifuged for 10 min at 1000 × g, and the cell pellet was
resuspended and fixed with 2% paraformaldehyde before taking
measurements on an Epics XL flow cytometer (Beckman Coulter). The
samples were analyzed using EXPO32 software (Beckman Coulter) and
are displayed as dotplots.
Imaging by CLSM
The recombinant vectors, pIRES-TCRβ-EYFP/TCRα-ECFP
and pIRES-TCRβζ-EYFP/TCRαζ-ECFP, were transfected into the Jurkat
and BEL-7402 cells using Lipofectamine LTX/PLUS (Invitrogen)
according to the manufacturer’s instructions. The cells transfected
with pIRES-TCRα-ECFP and pIRES-TCRβ-EYFP were used as controls to
remove the SBT contamination in FRET analysis. After 24 h of
transfection, the Jurkat cells grown on glass-bottom dishes were
washed twice with PBS and immobilized with 0.05% low metling
agarose for 15 min and the BEL-7402 cells were washed twice with
PBS solution. Confocal images of the cells were acquired using an
Olympus FluoView 1000 confocal laser scanning microscope with
FV10-ASW 1.7 software (Olympus, Tokyo, Japan). The FRET donor
(TCRα-ECFP or TCRαζ-ECFP) was excited with a 458 nm Ar-laser and
the acceptor (TCRβ-EYFP or TCRβζ-EYFP) was excited with a 515 nm
Ar-laser. The cells were scanned from 475 to 585 nm with a 10 nm
step-size and 20 nm band-width to obtain original images.
FRET analysis
FRET efficiency was analyzed using the sensitized
acceptor emission (SE) method. FRET occurs when 2 fluorophores
(donor and acceptor) have sufficiently large spectral overlap which
results in the energy of the donor fluorescence transferring to the
acceptor. The spectral overlap between donor and acceptor
fluorophores also causes FRET signal contamination, termed SBT. It
is important to remove SBT in FRET efficiency measurements. The
pFRET images were corrected as shown in the equation and as
previously described (22): pFRET
= uFRET − DSBT − ASBT, where uFRET is uncorrected FRET, ASBT is the
acceptor spectral bleed-through signal, and DSBT is the donor
spectral bleed-through signal. The final FRET efficiency equation
was calculated as E = 1 − IDA/{IDA + pFRET ×
[(ψdd/ψaa) × (Qd/Qa,
where IDA is the intensity of the donor in the presence
of the acceptor, ψdd and ψaa are the
collection efficiency in the donor and acceptor channel, and
Qd and Qa are the quantum yield of the donor
and acceptor, respectively (22,23).
Statistical analysis
Mean FRET efficiency was calculated from multiple
(n=6) cell images in each group and 6 random regions of interest
(ROI) in each cell image. Statistical analysis was performed using
two-way ANOVA with a Bonferroni’s multiple comparisons test using
Graphpad Prism 6 software (GraphPad Software Inc., San Diego, CA,
USA). Differences with P-values <0.05 were considered
statistically significant. Data are expressed as the means ±
SD.
Results
TCRαζβζ generation and location on the
surface of cells
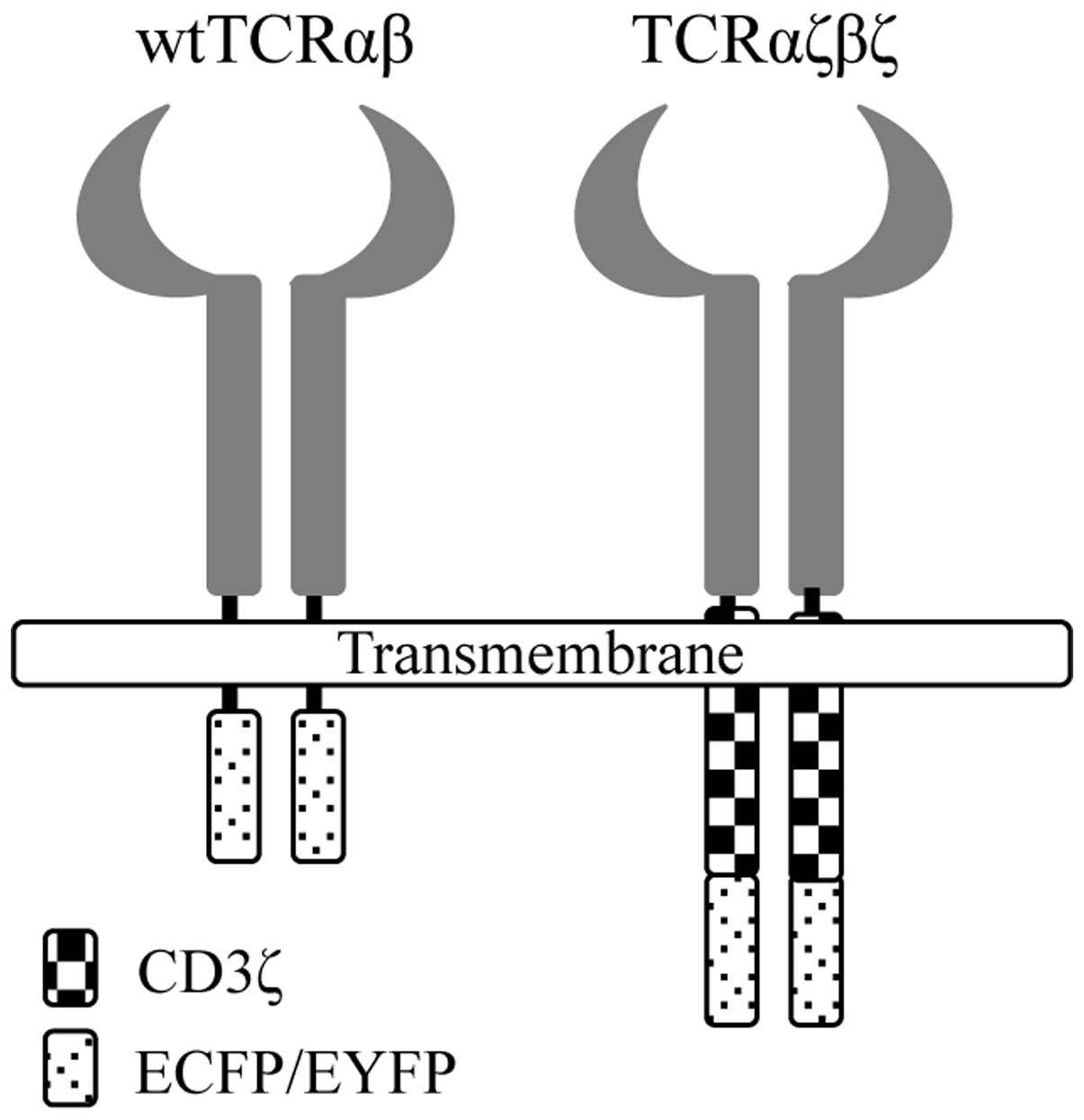
We designed a modified TCR in which the
extracellular and transmembrane domain of TCRα or β chains were
exchanged for CD3ζ chains at a structurally favorable position. To
better observe the expression of the modified TCRαζβζ in living
cells, we fused the C terminus of TCRαζ and TCRβζ chains to a pair
of cyan and yellow fluorescent proteins, ECFP and EYFP,
respectively (Fig. 1). The fusion
genes, TCRαζ-ECFP and TCRβζ-EYFP, were obtained by SOE PCR
(Fig. 2B and C). To compare the
relative levels of surface expression with wtTCRαβ, we also fused
the C terminus of the wtTCRα and wtTCRβ chains to a pair of ECFP
and EYFP fluorescent proteins (Fig.
1). The fusion genes were then cloned into the pIRES2-EGFP
vector.
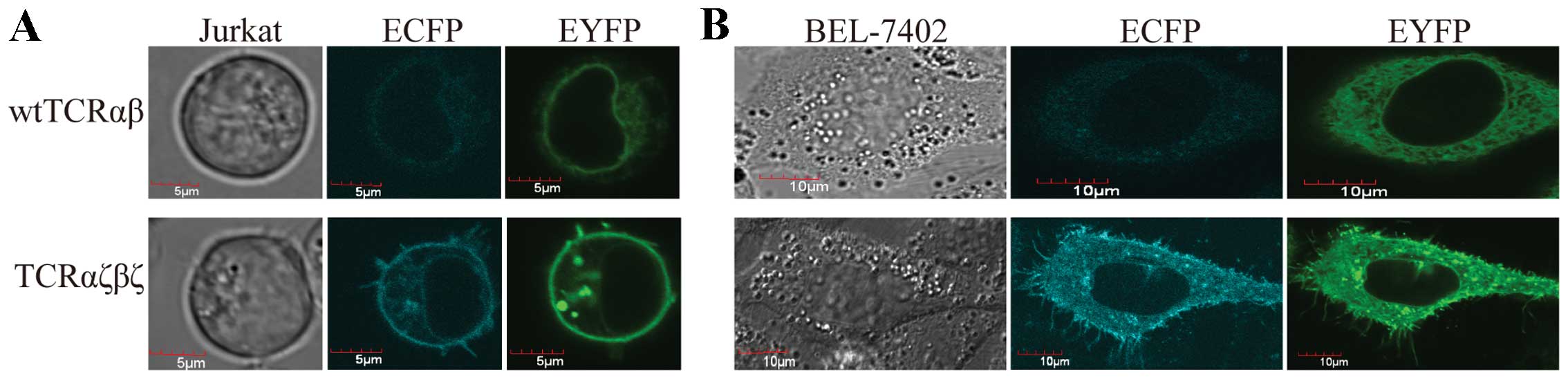
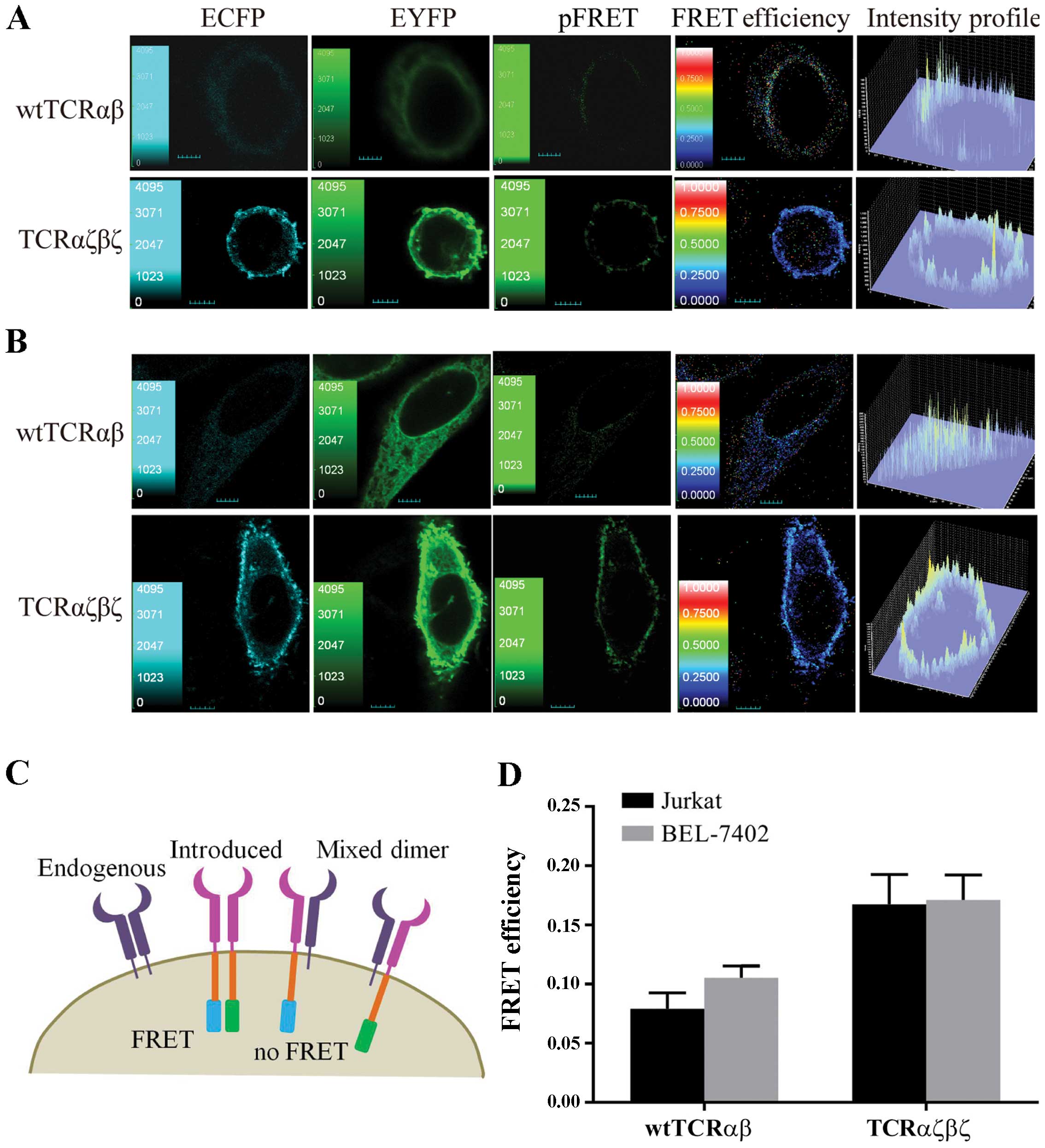
We first examined whether the TCRαζ-ECFP and
TCRβζ-EYFP fusion chains are expressed on the surface of the cells.
We transfected the recombinant plasmids into Jurkat cells, as well
as BEL-7402 cells, and 24 h after transfection, the fluorescence
observation by CLSM revealed that the introduced fusion genes,
TCRαζ-ECFP and TCRβζ-EYFP, were located at the plasma membrane of
the cells and were effectively expressed in T cells and non-T cells
(Fig. 3).
TCRαζβζ enhanced surface expression in T
cells and non-T cells
We observed a very interesting phenomenon, namely
that in the TCRαζβζ-transduced cells, but not in the
TCRαβ-transduced cells, microclusters were clearly evident at the
plasma membrane and the cilia of the plasma membrane (Fig. 3). This observation indicated that
TCRαζβζ expression was stronger in the Jurkat cells and BEL-7402
cells compared to the expression of wtTCRαβ. To confirm this
result, we inserted the wtTCRαβ and TCRαζβζ genes separately into
the shuttle plasmid, pDC315, to produce Ad5F35 adenovirus (Fig. 4A). Ad5F35 adenovirus which
contained the Ad35 fiber knob incorporated into an Ad5 capsid was
able to effectively transduce human T cells. The wtTCRαβ and
TCRαζβζ adenoviral vectors were introduced into the Jurkat T cells
followed by FACS analysis. Double immunofluorescent staining with
anti-TCRVα12 mAbFITC and anti-Vβ7 mAbPE
showed that the Jurkat cells transduced with wtTCRαβ displayed
little surface co-expression (up to 10.7%). By contrast, there was
a clear increased surface coexpression (up to 21.2%) in the
TCRαζβζ-transduced Jurkat cells (Fig.
4B). These data are in accordance with the imaging evidence
showing the enhanced surface expression of TCRαζβζ in the cells.
These results demonstrated that the FRET reporter,
TCRαζ-ECFP/TCRβζ-EYFP, can be effectively used to express TCRαζβζ
may thus be used for monitoring the expression and interaction of
TCRαζβζ.
Highly preferred pairing between TCRαζ
and βζ, but not wtTCRα and β in Jurkat cells
To investigate the interaction of TCRαζ and βζ in T
cells, we used Jurkat cells (clone E6-1) expressing endogenous TCR
as a recipient TCR cell model. We hypothesized that the introduced
wtTCRα and β chains or TCRαζ and βζ chains composing heterodimers
would result in FRET efficiency between the donor (ECFP) and
acceptor (EYFP) fluorescent proteins. Once the introduced TCRαζ and
βζ chains paired with endogenous TCRβ and α chains, it would fail
to detect FRET between the mispaired TCR (Fig. 5C). We observed that the TCRαζβζ
displayed a higher FRET signal compared to wtTCRαβ in Jurkat cells.
(Fig. 5A). Six independent cell
images and 6 ROI in each cell image were selected for FRET
efficiency analysis in each group. The results from statistical
analysis showed that the average FRET efficiency between TCRαζ-ECFP
and TCRβζ-EYFP (16.7±2.5%) was significantly increased compared to
wtTCRα-ECFP and wtTCRβ-EYFP (7.9±1.3%) in the Jurkat cells
(P<0.0001) (Fig. 5D). These
data suggested that the fusion of wtTCRα12 and β7 with CD3ζ
improved pairing.
TCRαζβζ assembles independently of CD3
subunits in T cells and non-T cells
To better characterize the highly preferred pairing
of TCRαζ and βζ, we selected BEL-7402 cells which are deficient in
TCR and CD3 molecules as the next recipient cell model to
investigate the interaction of TCRαζβζ and endogenous CD3 subunits.
We hypothesized that since there was no endogenous TCR in BEL-7402
cells, the pairing of introduced wtTCRα and β or TCRαζ and βζ would
not be interfered with and would result in the same FRET signal.
Unexpectedly, we observed that TCRαζβζ still displayed a higher
FRET signal compared to wtTCRαβ in BEL-7402 cells (Fig. 5B). The average FRET efficiency
between wtTCRα and β chains (10.5±1.0%) was significantly reduced
compared to the TCRαζ and βζ chains (17.1±2.1%) in BEL-7402 cells
(P<0.0001) (Fig. 5D). The
results indicated that the assemble of wtTCRα and β chains was
impaired in the absence of CD3 subunits, in spite of no endogenous
TCR in the BEL-7402 cells. Of note, we found that the average FRET
efficiency between TCRαζ-ECFP and TCRβζ-EYFP in the BEL-7402 cells
(17.1±2.1%) was not reduced compared to the Jurkat cells
(16.7±2.5%) (P>0.05) (Fig.
5D). These results demonstrated that the expression and
assembly of TCRαζβζ was not compromised even without the CD3γɛ, Δɛ
and ζζ signaling dimers in the BEL-7402 cells.
Discussion
In the present study, we generated several ECFP and
EYFP fusions as a pair of reporters that allow the monitoring of
the expression and pairing between TCRαζ and TCRβζ in living cells.
First, we used a wtTCRαβ (Vα12 and Vβ7) isolated from TILs of
patients fused to complete human CD3ζ molecule to generate a pair
of chimeric TCRαζ and TCRβζ, then ECFP fused to TCRαζ, and EYFP
fused to TCRβζ via overlap PCR, respectively. This suggested
strategy using these reporters offers several advantages over
conventional immunofluorescent staining using specific antibodies:
it provides accurate imaging of modified TCRαβζ expression in
Jurkat T cells, and can be used in combination with FRET methods to
study the interaction of TCRαζ and TCRβζ in living cells.
Using these reporters, we observed that the
expression of the introduced TCRαζβζ was enhanced when compared to
the non-modified wtTCRαβ in Jurkat cells, which was in line with
our FACS analysis. Our results showed that the fusion gene,
TCRαζβζ, coupled with ECFP/EYFP was able to be expressed normally
on the surface of cells. Furthermore, we found that wtTCRαβ and
TCRαζβζ were expressed not only in Jurkat cells but also in non-T
cells (BEL-7402). Of note, we observed that the TCRαζβζ-transduced
cells, but not the wtTCRαβ-transduced cells showed evident
microclusters at the plasma membrane and cilia of the plasma
membrane. These phenomena indicated that the surface expression of
TCRαζβζ was markedly enhanced due to the fusion CD3ζ molecules,
whereas the expression of wtTCRαβ was impaired in the absence of
CD3 molecules.
The fluorescence images of the reporters were then
subjected to FRET analysis which allowed the detection of the
interaction of TCRαβ in the living cells. The FRET efficiencies
between wtTCRα-ECFP and β-EYFP or TCRαζ-ECPF and βζ-EYFP in the
Jurkat cells revealed that the introduction of TCRαζβζ improved
paring compared to the wtTCRαβ majority of which mispaired with
endogenous TCR. The FRET efficiency of TCRαζ and βζ in the Jurkat
and BEL-7402 cells did not show any significant differences,
suggesting that TCRαζβζ may be expressed/or assembled in BEL-7402
cells without endogenous CD3 subunits. The assembly of the TCR-CD3
complex is in part determined by the transmembrane charges, the
positive charges at the transmembrane regions of TCRα interacting
with the negative charges at the CD3Δɛ and CD3ζζ dimers, while TCRβ
interacts with CD3γɛ (24–26).
In the TCRαζ and βζ chains, the extracellular connecting peptide
motif, transmembrane and intracellular domains of TCRα and β are
replaced by CD3ζ. Thereby, CD3γɛ or CD3Δɛ cannot be associated with
the modified TCRαζβζ, which explains why TCRαζβζ can expressed
independently of CD3 components.
In conclusion, using the strategy of fluorescent
proteins fused to the TCRαζ and βζ chains we monitored and measured
the expression and interaction of TCRαζ and TCRβζ in living cells
accurately. The ECFP and EYFP fusion reporters revealed that the
TCRαβ chains fused to CD3ζ enhanced the expression and prevented
mispairing, paving the way to potential immunological studies
dealing with TCR mispairing.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (nos. 31100664,
31300737 and 81303292), the National Major Scientific and
Technological Special Project for ‘Significant New Drugs
Development’ (no. 2009ZX09103-708), the Natural Science Foundation
of Guangdong Province (no. 10151022401000024), and a grant from the
Faculty Development and Research Funds of GDPU and The Science and
Technology Research Project of Dongguan City (no.
2011105102027).
References
|
1
|
Heemskerk MH, Hoogeboom M, Hagedoorn R, et
al: Reprogramming of virus-specific T cells into leukemia-reactive
T cells using T cell receptor gene transfer. J Exp Med.
199:885–894. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hughes MS, Yu YY, Dudley ME, et al:
Transfer of a TCR gene derived from a patient with a marked
antitumor response conveys highly active T-cell effector functions.
Hum Gene Ther. 16:457–472. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Abad JD, Wrzensinski C, Overwijk W, et al:
T-cell receptor gene therapy of established tumors in a murine
melanoma model. J Immunother. 31:1–6. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Coccoris M, Swart E, de Witte MA, et al:
Long-term functionality of TCR-transduced T cells in vivo. J
Immunol. 180:6536–6543. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
de Witte MA, Bendle GM, van den Boom MD,
et al: TCR gene therapy of spontaneous prostate carcinoma requires
in vivo T cell activation. J Immunol. 181:2563–2571.
2008.PubMed/NCBI
|
|
6
|
Dossett ML, Teague RM, Schmitt TM, et al:
Adoptive immunotherapy of disseminated leukemia with
TCR-transduced, CD8+ T cells expressing a known
endogenous TCR. Mol Ther. 17:742–749. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Morgan RA, Dudley ME, Wunderlich JR, et
al: Cancer regression in patients after transfer of genetically
engineered lymphocytes. Science. 314:126–129. 2006. View Article : Google Scholar
|
|
8
|
Johnson LA, Morgan RA, Dudley ME, et al:
Gene therapy with human and mouse T-cell receptors mediates cancer
regression and targets normal tissues expressing cognate antigen.
Blood. 114:535–546. 2009. View Article : Google Scholar
|
|
9
|
Bendle GM, Linnemann C, Hooijkaas AI, et
al: Lethal graft-versus-host disease in mouse models of T cell
receptor gene therapy. Nat Med. 16:565–570. 2010. View Article : Google Scholar
|
|
10
|
van Loenen MM, de Boer R, Amir AL, et al:
Mixed T cell receptor dimers harbor potentially harmful
neoreactivity. Proc Natl Acad Sci USA. 107:10972–10977.
2010.PubMed/NCBI
|
|
11
|
Cohen CJ, Zhao Y, Zheng Z, et al: Enhanced
antitumor activity of murine-human hybrid T-cell receptor (TCR) in
human lymphocytes is associated with improved pairing and TCR/CD3
stability. Cancer Res. 66:8878–8886. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sommermeyer D and Uckert W: Minimal amino
acid exchange in human TCR constant regions fosters improved
function of TCR gene-modified T cells. J Immunol. 184:6223–6231.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kuball J, Dossett ML, Wolfl M, et al:
Facilitating matched pairing and expression of TCR chains
introduced into human T cells. Blood. 109:2331–2338. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cohen CJ, Li YF, El-Gamil M, et al:
Enhanced antitumor activity of T cells engineered to express T-cell
receptors with a second disulfide bond. Cancer Res. 67:3898–3903.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Voss RH, Willemsen RA, Kuball J, et al:
Molecular design of the Cαβ interface favors specific pairing of
introduced TCRαβ in human T cells. J Immunol. 180:391–401.
2008.PubMed/NCBI
|
|
16
|
Willemsen R, Weijtens M, Ronteltap C, et
al: Grafting primary human T lymphocytes with cancer-specific
chimeric single chain and two chain TCR. Gene Ther. 7:1369–1377.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang T, He X, Tsang TC, et al: Transgenic
TCR expression: comparison of single chain with full-length
receptor constructs for T-cell function. Cancer Gene Ther.
11:487–496. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schaft N, Lankiewicz B, Drexhage J, et al:
T cell re-targeting to EBV antigens following TCR gene transfer:
CD28-containing receptors mediate enhanced antigen-specific IFNγ
production. Int Immunol. 18:591–601. 2006.PubMed/NCBI
|
|
19
|
Wu F, Zhang W, Shao H, et al: Human
effector T cells derived from central memory cells rather than
CD8+ T cells modified by tumor-specific TCR gene
transfer possess superior traits for adoptive immunotherapy. Cancer
Lett. 339:195–207. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shao H, Zhang W, Hu Q, et al: TCR
mispairing in genetically modified T cells was detected by
fluorescence resonance energy transfer. Mol Biol Rep. 37:3951–3956.
2010. View Article : Google Scholar
|
|
21
|
Schroers R, Hildebrandt Y, Hasenkamp J, et
al: Gene transfer into human T lymphocytes and natural killer cells
by Ad5/F35 chimeric adenoviral vectors. Exp Hematol. 32:536–546.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Elangovan M, Wallrabe H, Chen Y, et al:
Characterization of one-and two-photon excitation fluorescence
resonance energy transfer microscopy. Methods. 29:58–73. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wallrabe H and Periasamy A: Imaging
protein molecules using FRET and FLIM microscopy. Curr Opin
Biotech. 16:19–27. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Call ME, Wucherpfennig KW and Chou JJ: The
structural basis for intramembrane assembly of an activating
immunoreceptor complex. Nat Immunol. 11:1023–1029. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kuhns MS, Girvin AT, Klein LO, et al:
Evidence for a functional sidedness to the αβTCR. Proc Natl Acad
Sci USA. 107:5094–5099. 2010.
|
|
26
|
Call ME, Schnell JR, Xu C, et al: The
structure of the ζζ transmembrane dimer reveals features essential
for its assembly with the T cell receptor. Cell. 127:355–368.
2006.
|