Introduction
Obesity has a partly genetic basis; genes of the
endocrine system and metabolism may favor obesity. However, the
direct cause of obesity is an imbalance between energy storage and
consumption, and thus an imbalance between calorie intake and
consumption (1,2). The incidence and severity of
obesity-related diseases, including circulatory diseases, such as
diabetes, cancer and high blood pressure render obesity one of the
leading causes of mortality (3,4).
Physiologically, obesity is associated with increased levels of fat
within adipocytes and/or an increase in the number of adipocytes.
Accumulated intracellular triglycerides (TGs) can be broken down by
exercise or diet; however, obesity caused by increased numbers of
adipocytes is difficult to treat as the fat cells must be destroyed
or removed. The differentiation of pre-adipocytes into adipocytes
may be triggered by changes in specific hormone levels and/or
excessive nutritional intake during the growth process. Adipocyte
differentiation may occur in particular regions of the body. In
addition, due to limitations in the size of adipocytes, the number
of such cells increases with excess energy consumption, in order to
provide storage for these excess energy levels (5). The 3T3-L1 cell line is a
pre-adipocyte line derived from mouse embryonic fibroblasts and is
the most commonly used line in obesity research.
In the case that glucose and TGs accumulate and the
CCAAT/enhancer-binding protein (C/EBP)-β or -δ is induced during
early adipocyte differentiation, the activation of C/EBP-α, a
peroxisome proliferator-activated receptor (PPAR) transcription
factor, occurs late in differentiation, in turn causing the
transcriptional activation of various adipocyte-specific mRNAs
encoding fatty acid synthase (FAS), lipoprotein lipase (LPL),
acetyl-CoA carboxylase (ACC), stearoyl-CoA desaturase-1 (SCD-1) and
PPAR co-activator-1 (PGC-1). These enzymes are involved in
lipogenesis and adipogenesis, and induce the synthesis of fat
globules. Wnts are a family of secreted glycoproteins with a high
number of conserved cysteines. They regulate various morphogenetic
processes by controlling the expression of a number of genes
through several steps triggered when Wnt receptors are activated by
binding to the receptors in surrounding cells (6–8).
The Wnt canonical signal transmission system plays a major role in
the differentiation of adipocytes. As previously demonstrated, when
a canonical signal transmission system, such as Wnt is activated in
pre-adipocytes and Wnt-10b is overexpressed, adipocyte formation is
prevented through the inhibition of C/EBPs and PPAR-γ (key factors
in adipocyte cell differentiation), in turn suppressing the
expression of adipocyte-specific genes (9). In addition, the balance between
β-catenin and PPAR-γ levels is important in terms of adipocyte
differentiation; maintaining this equilibrium depends on
proteolysis following the phosphorylation of β-catenin by glycogen
synthase kinase-3β (GSK-3β) (10).
Tuna is known to have anti-arteriosclerotic and
anti-obesity properties, and to reduce the levels of cholesterol in
the blood. Processed tuna is principally muscle tissue. In a
previous study of ours, we analyzed boiled tuna extract, identified
a relevant peptide in that extract, synthesized that peptide
(11), and measured the
anti-obesity effects thereof. We found that the peptide inhibited
the adipocyte differentiation of 3T3-L1 cells.
Materials and methods
Preparation of desalinated boiled tuna
extract
The desalinated boiled tuna extract used in this
study was prepared in 2014 in Korea. First, the boiled tuna extract
was centrifuged to remove suspended solids that may interfere with
desalination. This process changed the Brix status from 55 to 45
Brix, and the salinity from 12 to 13%. Membrane filtration was
performed (cut-off, 200 Da). We obtained a solution of 30 Brix and
1% salinity and sterilized the material using a heat exchanger
(conditions: 110°C for 10 sec). The sample was then placed in a
1.5-ml tube and stored at −70°C until use.
Preparation of soluble/insoluble tuna
protein and synthesis of the tuna peptide
The desalinated boiled tuna extract was incubated in
a Tween-20 extraction buffer overnight at room temperature,
followed by centrifugation at 5,000 rpm for 10 min at 4°C. The
upper phase was mixed with cold methanol/chloroform to induce
protein separation. The solution was then centrifuged at 12,000 rpm
for 5 min at 4°C. The aqueous layer was removed, and cold methanol
was added, followed by centifugation at 12,000 rpm for 10 min at
4°C. The supernatant was removed and dried.
The molecular weights of the tuna proteins were
investigated by Coomassie blue (7% acetic acid, 40% methanol, and
0.1% bromophenol blue; all v/v) staining of SDS-PAGE gels and
destaining (7% acetic acid, 20% methanol; both v/v). Tuna proteins
approximately 10 kDa in size were analyzed by
quadrupole-time-of-flight tandem mass spectrometry (Q-TOF MS/MS).
The peptide mixture was desalinated and concentrated in a C18
column packed with POROS R2 (20–30 µm bead size; PerSeptive
Biosystems, Inc., Framingham, MA, USA). MS/MS involved
nano-electrospray ionization (ESI) and micro Q-TOF MS (Bruker
Daltonics, Bremen, Germany). The tuna-derived peptide
D-I-V-D-K-I-E-I (TP-D) was synthesized by Peptron (Daejeon, Korea),
and the purified tuna peptide was synthesized on a C18 column
(Shiseido Capcell Pak; Shiseido, Tokyo, Japan) using a Shimadzu
Prominence HPLC system (Shimadzu, Kyoto, Japan). The eluent was
placed in 0.1% (v/v) trifluoroacetic acid (TFA)/water with a
gradient of 0–90% (v/v) acetonitrile. The flow rate was 1 ml/min,
and detection was carried out via UV at 220 nm. The molecular
weight of TP-D was 944 Da (determined using an HP 1100 Series
LC/MSD system; Agilent Technologies, Santa Clara, CA, USA).
Cell culture and adipocyte
differentiation
3T3-L1 mouse pre-adipocytes (derived from
fibroblasts; American Type Culture Collection, Manassas, VA, USA)
were maintained at a temperature of 37°C in a humidified atmosphere
containing 5% (v/v) CO2. The cells were cultured in
Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10%
(v/v) bovine calf serum (BCS;HyClone, Logan, UT, USA) and
penicillin/streptomycin (100 U/ml/100 mg/ml). The cells were
cultured to 60–80% confluence in a 6-well plate and, upon reaching
confluence, were allowed to grow for an additional 2–4 days in DMEM
with 10% (v/v) fetal bovine serum (FBS; HyClone). Cell
differentiation was initiated by treatment with MDI [0.5 mM
3-isobutyl-1-methylxanthine (IBMX), 0.25 µM dexamethasone
and 10 mg/l insulin] for 48 h. The medium was then replaced with
DMEM containing 10 mg/l insulin and changed every 2 days.
Cell proliferation assays
The 3T3-L1 cell proliferation was measured using the
CellTiter 96® AQueous Non-Radioactive Cell Proliferation
assay (Promega Corp., Madison, WI, USA). The assay is based on the
cleavage of
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxy-phenyl)-2-(4-sulfonyl)-2H-tetra-zolium
(MTS) to a formazan product soluble in the cell culture medium. The
cells were seeded into 96-well plates at 2×104
cells/well in 100 µl of medium and maintained for 24 h; the
medium was then replaced with serum-free medium (SFM). After 24 h,
the medium was replaced with SFM containing TP-D (125, 250, 500 or
1,000 µg/ml) for 24 h. Subsequently, the cells were
incubated with MTS solution for 30 min at 37°C. Cell proliferation
was determined by measuring the absorbance at 490 nm using a
Benchmark enzyme-linked immunosorbent assay (ELISA) plate reader
(Bio-Rad Laboratories, Hercules, CA, USA).
Glucose uptake assay
The 3T3-L1 pre-adipocytes were incubated in DMEM
containing 10% (v/v) BCS. Cell differentiation was induced by
treatment with MDI in fresh DMEM containing 10% (v/v) FBS.
Following differentiation, the medium was replaced with SFM
containing TP-D at concentrations of 125, 250, 500 or 1,000 ng/ml
for 48 h prior to the glucose uptake assay using a glucose kit
protocol (Asan Pharmaceutical Co., Ltd., Hwaseong, Korea). The test
enzyme was added to the culture medium maintained at 37°C for 5 min
in a humidified atmosphere containing 5% (v/v) CO2. The
absorbance at 500 nm was measured within 40 min.
TG assay
The 3T3-L1 pre-adipocytes were incubated in DMEM
containing 10% (v/v) BCS. Cell differentiation was induced by
treatment with MDI in fresh DMEM with 10% (v/v) FBS. Following the
induction of cell differentiation, the medium was replaced with SFM
containing TP-D at concentrations of 125, 250, 500 and 1,000 ng/ml
for 48 h prior to the TG assay. Cell pellets were ruptured with
phosphate-buffered saline (PBS), and the TG levels were assayed
using a TG kit protocol (obtained from Asan Pharmaceutical Co.,
Ltd.). The test enzyme (TG-measured solution) was added to the
lysate supernatants, and the cells were maintained at 37°C for 5
min in a humidified atmosphere containing 5% CO2. The
absorbance at 550 nm was measured within 60 min.
mRNA expression by RT-PCR
The 3T3-L1 pre-adipocytes were seeded into 6-well
plates at 2×104 cells/well in 2 ml of medium. Cell
differentiation was then induced by treatment with MDI, as
described in the experiments above (11). Following the induction of cell
differentiation, the medium was replaced with SFM containing TP-D
(500 or 1,000 ng/ml) for 48 h and the cells were then treated with
1 ml of TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA,
USA). Subsequently, the samples were centrifuged at 14,000 rpm for
15 min at 4°C, after adding 200 µl of chloroform. The
samples were divided into supernatant and pellet (0.1% of DEPC was
added to 50 µl of water). The mRNA levels in the cell
supernatants were quantified using an Oligo(dT) primer (Intron
Biotechnology, Seoul, Korea) and the cDNA was synthesized at 50°C
for 1 h, 95°C for 5 min. The cDNA was added to 2X TOPsimple™
DyeMIX-nTaq (Enzynomics, Daejeon, Korea), and primers were added to
0.1% (v/v) diethylpyrocarbonate (DEPC) in water. The PCR reactions
for the amplification of the DNA were run on 1% (w/v) agarose gels
and the nuclei were stained using RedSafe Nucleic Acid Staining
solution (Intron Biotechnology). Initial denaturation was performed
at 95°C for 2 min and the denaturation step at 95°C for 30 sec. In
order to allow annealing of the primers to the single-stranded DNA
template, the temperature was lowered to 50–55°C for 30 sec. The
elongation step was performed at 72°C for 1 min and the final
elongation at 72°C for 5 min.
Western blot analysis
The 3T3-L1 pre-adipocytes were seeded into 6-well
plates at 2×104 cells/well in 2 ml of medium. Cell
differentiation was induced by treatment with MDI, as described
above (11). Following
differentiation, the medium was replaced with SFM containing TP-D
(500 or 1,000 ng/ml) for 48 h. The cells were then washed with PBS,
and lysis buffer was added [20 mM Tris Base (pH 8), 150 mM NaCl,
100 µM sodium vanadate, 100 µM ammonium molybdate,
10% (v/v) glycerol, 0.1% (v/v) Nonidet P-40, 0.1% (w/v) SDS, 1 mM
glycerophos-phate, 1 µg/ml aprotinin, 1 µg/ml
leupeptin, 1 µg/ml pepstatin A and 1 mM
phenylmethanesulfonyl fluoride (PMSF)]. Proteins were separated by
7–15% (w/v) SDS-PAGE and transferred onto polyvinylidene fluoride
membranes (Millipore, Billerica, MA, USA), which were blocked at
room temperature with 1% (w/v) bovine serum albumin in TBS-T [10 mM
Tris-HCl (pH 7.5), 150 mM NaCl, 0.1% (v/v) Tween-20] and then
incubated with the following antibodies while being shaken:
anti-C/EBP-α (sc-9314, anti-rabbit; 1:1,000), anti-C/EBP-β (sc-150,
anti-rabbit; 1:1,000), anti-C/EBP-δ (sc-151, anti-rabbit; 1:1,000),
anti-PPAR-γ (sc-1984, anti-goat; 1:1,000), anti-CD36 (sc-7641,
anti-goat; 1:1,000), anti-sterol regulatory element-binding protein
1 (SREBP-1; sc-366, anti-rabbit; 1:1,000), anti-FAS (sc-7886,
anti-mouse; 1:1,000), anti-ACC (sc-271965, anti-mouse; 1:1,000),
anti-LPL (sc-32382, anti-goat; 1:1,000), anti-suppressor of
cytokine signaling-3 (SOCS-3; sc-73045, anti-mouse; 1:1,000),
anti-fatty acid binding protein (FABP; sc-18661, anti-goat;
1:1,000), anti-uncoupling protein (UCP)-1 (sc-6529, anti-goat;
1:1,000), anti-UCP-2 (sc-6525, anti-goat; 1:1,000), anti-glucose
transporter type 4 (GLUT4; sc-1606, anti-goat; 1:1,000),
anti-Wnt-10b (sc-25524, anti-rabbit; 1:1,000), anti-Frizzled
(sc-130758, anti-rabbit; 1:1,000), anti-lipoprotein
receptor-related protein-6 (LRP6; sc-25317, anti-mouse; 1:1,000),
anti-dishevelled (Dvl; sc-166303, anti-mouse; 1:1,000),
anti-β-catenin (sc-1496, anti-goat; 1:1,000), anti-GSK-3β
(sc-377213, anti-mouse; 1:1,000), anti-T cell factor (TCF;
sc-271453, anti-mouse; 1:1,000), anti-lymphoid enhancer-binding
factor (LEF; sc-28687, anti-rabbit; 1:1,000), anti-cyclin D1
(CCND1; sc-753, anti-rabbit; 1:1,000) or anti-glyceraldehyde
3-phosphate dehydrogenase (GAPDH; sc-25778, anti-rabbit; 1:1,000)
antibodies (all from Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA). The secondary antibodies were peroxidase-conjugated goat
(sc-2741), mouse (sc-2032) or rabbit (sc-2031) antibody (1:10,000;
GE Healthcare Bio-Sciences, Piscataway, NJ, USA). Proteins were
visualized by exposure to SuperSignal West Pico Stable Peroxide
solution and SuperSignal West Pico Luminol/Enhancer solution (both
from Thermo Fisher Scientific, Rockford, IL, USA) and Kodak X-ray
film.
Assays of total and high-molecular weight
(HMW) adiponectins
Adiponectin and HMW adiponectin levels were measured
using an ELISA kit (ALPCO Diagnostics, Salem, NH, USA). The 3T3-L1
pre-adipocytes were incubated with DMEM containing 10% (v/v) BCS in
a 6-well plate. Cell differentiation was induced by treatment with
MDI, as described above (11).
Following differentiation, the medium was replaced with SFM
containing TP-D (500 or 1,000 ng/ml) for 48 h. A total of 50
µl of medium was assayed. The culture medium and cell lysate
were incubated with 100 µl of protease buffer, which was
then neutralized with 100 µl of pre-treatment buffer. The
absorbances of total and HMW adiponectin at 492 nm were
measured.
Statistical analysis
The results are presented as the means ± SD and were
analyzed using SPSS version 10.0 software (SPSS,Inc., Chicago, IL,
USA). Data were validated by ANOVA, and a P-value <0.05 as
indicated by Duncan’s multiple range test was considered to
indicate a statistically significant difference.
Results
TP-D is not toxic to 3T3-L1
pre-adipocytes
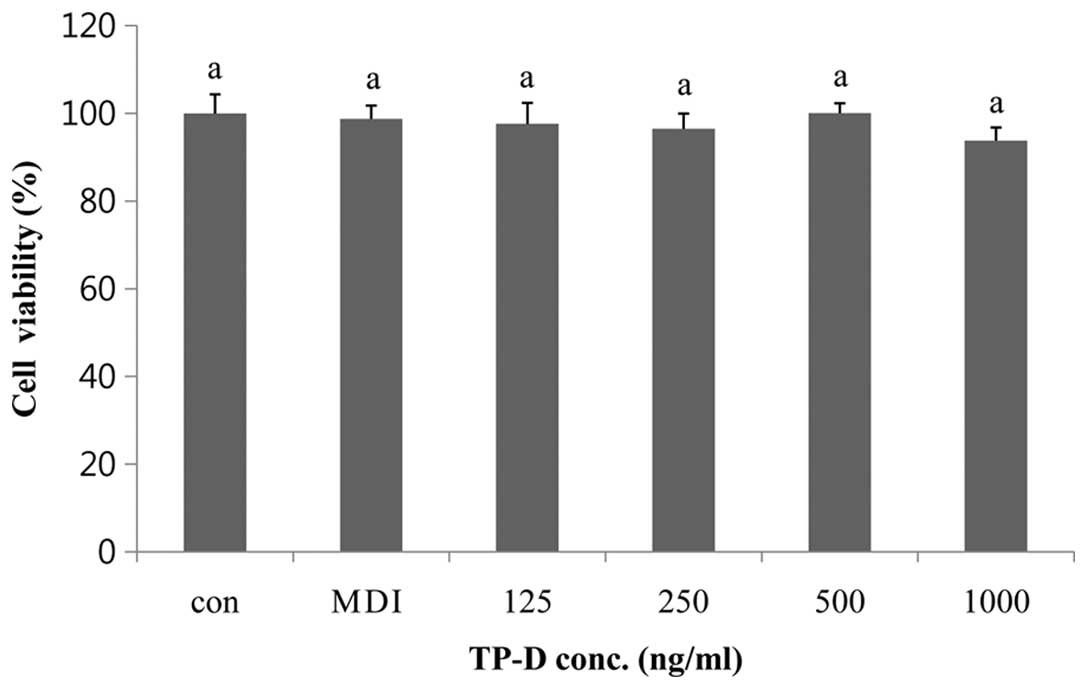
The effect of TP-D on the viability of the 3T3-L1
pre-adipocytes was investigated by MTS assay. The cells were seeded
into 96-well plates at 2×104 cells/well in 100 µl
of medium and allowed to attach for 24 h. The medium was replaced
with SFM for 4 h, followed by treatment of the 3T3-L1 cells with
various concentrations of TP-D (125, 250, 500 and 1,000
µg/ml) for 24 h. MTS solution was then added and cell
viability was measured. TP-D was not found to be toxic to the
cells, as no significant decrease in cell viability was observed at
any of the concentrations used (Fig.
1).
Effect of TP-D on glucose uptake by
3T3-L1 cells
Glucose consumption is a prerequisite for 3T3-L1
cell differentiation. We compared glucose consumptions of
differentiated and undifferentiated cells treated with TP-D. The
cells were incubated with TP-D at concentrations of 125, 250, 500
and 1,000 ng/ml for 48 h. In the TP-D-treated group, glucose uptake
decreased compared with the MDI-reated group. Glucose uptake was
significantly decreased with all the tested concentrations of TP-D,
indicating that TP-D inhibited glucose consumption and thus the
differentiation of 3T3-L1 cells (Fig.
2A).
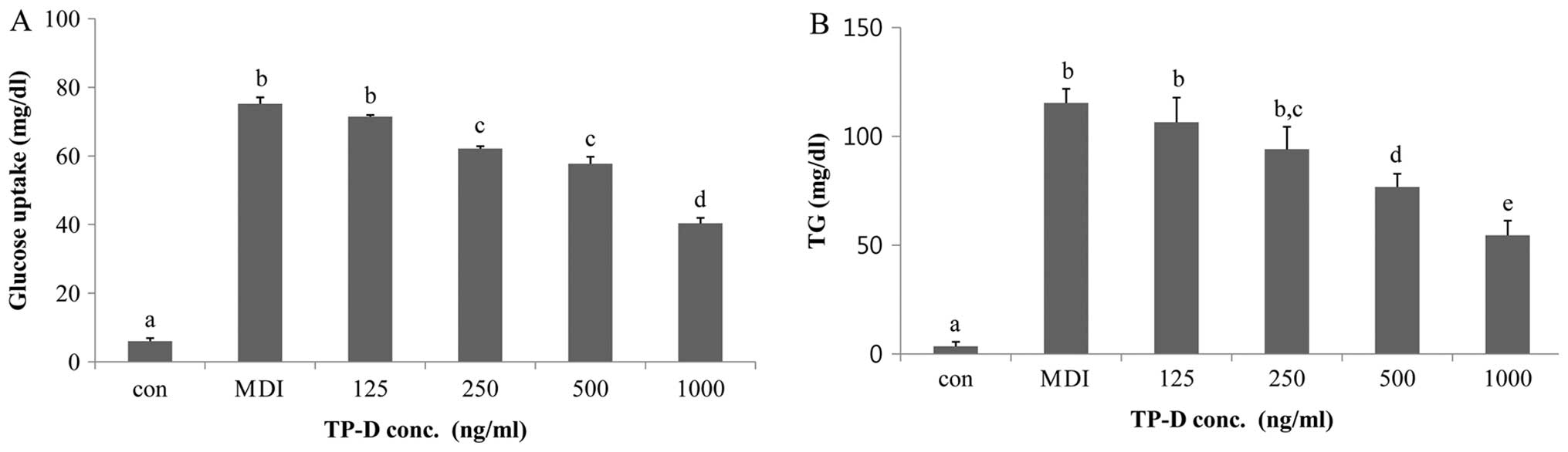 | Figure 2Effect of the tuna-derived peptide,
D-I-V-D-K-I-E-I (TP-D), on glucose uptake and triglyceride (TG)
levels in differentiated 3T3-L1 adipocytes. (A) The 3T3-L1 cells
were treated with TP-D at various concentrations (125, 250, 500 or
1,000 ng/ml) for 48 h. Glucose levels in the culture medium were
measured by a glucose uptake assay using an ELISA reader. (B) The
3T3-L1 cells were treated with TP-D at various concentrations (125,
250, 500 or 1,000 ng/ml) for 48 h. TG levels in the culture medium
were measured by a TG assay using an ELISA reader. Values represent
the means ± SD; P<0.05 as shown by ANOVA. Bars labeled with
different letters indicate significant differences according to
Duncan’s multiple range test. Con, control (undifferentiated
cells); conc., concentration. |
Effects of TP-D on TG levels in 3T3-L1
cells
In order to evaluate the effects of TP-D on TG
levels, the differentiated 3T3-L1 cells were treated with various
concentrations of TP-D (125, 250, 500 and 1,000 g/ml). Glucose
consumption induced cell differentiation and the accumulation of
TGs. As noted in our previous experiment descrbied above, glucose
uptake decreased when the cells were treated with TP-D (11). Therefore, we measured the TG
levels under the same treatment conditions. The TG levels in the
3T3-L1 cells decreased significantly and in a dose-dependent manner
following treatment with TP-D at 125, 250, 500 and 1,000 ng/ml for
48 h. The most significant decrease in the TG levels was observed
at the dose of 500 and 1,000 ng/ml. Therefore, all further
experiments were performed after 48 h of TP-D treatment (at 500 or
1,000 ng/ml; Fig. 2B).
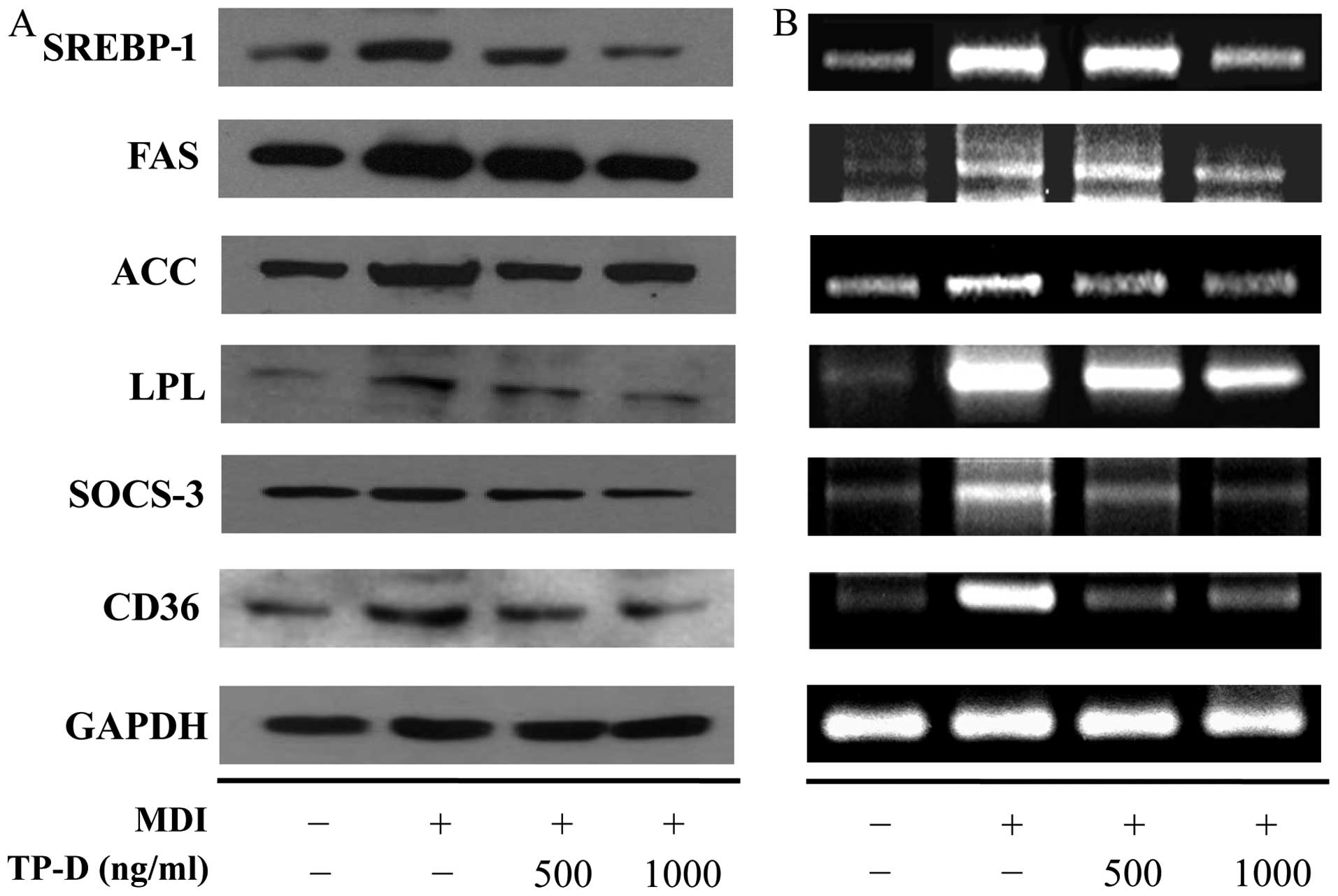
Effects of TP-D on the expression of
lipogenic and adipogenic genes during the differentiation of 3T3-L1
cells
C/EBP-β and -δ were overexpressed during the initial
differentiation of 3T3-L1 cells, which was triggered by adipogenic
inducers secreted by pre-adipocytes. This induced the expression of
C/EBP-α and PPAR-γ, transcription factors that are important in
differentiation (11). Western
blot analysis and RT-PCR were used to measure the protein and mRNA
expression levels, respectively. TP-D inhibited adipocyte formation
by downregulating the expression of C/EBPs and PPAR-γ (Fig. 3). Liver X receptors (LXRs) are
major nuclear receptors involved in the regulation of lipid
synthesis. They form LXR/retinoid X receptor (RXR) heterodimers
with RXR receptors and induce the activation of SREBP-1 (12–14), triggering lipid synthesis.
Activated SREBP-1 then upregulates ACC, FAS, FABP and LPL.
Adipogenesis and lipogenesis are thus accelerated, leading to the
accumulatin of lipids (15). In
addition, LXRs mediate the SREBP-1 pathway through the activation
of the uptake transporter, CD36 (16). SOCS-3 is overexpressed during
insulin resistance, thereby inducing SREBP-1 epxression, which is
involved in the homeostatic regulation of lipid levels (17,18). Thus, in this study, we measured
the expression levels of ACC, FAS, LPL, FABP, SREBP-1, SOCS-3 and
CD36 by western blot analysis and RT-PCR. The expression of levels
of all these factors was lower in the TP-D-treated group compared
with the MDI-treated group (Fig.
4). UCPs are mitochondrial membrane transporters of brown
adipose tissue. UCP-1 has a heat release function (lipids are
oxidized upon the upregulation of UCP-1). UCP-2 is distributed in
white adipose tissue and its expression increases as lipid cells
accumulate (19). In this study,
treatment with TP-D increased UCP-1 expression and decreased UCP-2
expression levels (Fig. 5). GLUT4
is involved in the active transport of glucose and its expression
is increased by C/EBPs (20). In
this study, treatment with TP-D decreased GLUT4 expression
(Fig. 5). The inhibition of GLUT4
plays a role in insulin sensitivity by selectively inhibiting the
activation of C/EBP-α by PPAR-γ.
Effect of TP-D on the expression of
Wnt-10b and the Wnt signaling pathway during 3T3-L1
differentiation
β-catenin, a member of the canonical Wnt signaling
pathway, is known to an anti-adipogenic regulator by inhibiting the
expression of C/EBP-α and PPAR-γ, which are play a major role in
lipid formation (10,21). Low-density LRP6 combines with Wnt
and Frizzled to bind to β-catenin, and the activation of
Wnt-10b/LRP6/Frizzled induces the upregulation of Dvl (22). This occurs during the degradation
of GSK-3β and catenin stabilization (23). The accumulation of catenin in the
cytoplasm coincides with shuttling of the transcription factor,
TCF/LEF, into the nucleus and the induction of CCND1. CCND1
inhibits C/EBP-α and PPAR-γ through a mechanism involving direct
interaction with PPAR (24). In
this study, we observed changes in Wnt signaling-related protein
and mRNA expression by western blot analysis and RT-PCR. As shown
in Fig. 6, the CCND1 and
β-catenin/TCF/LEF levels increased upon the activation of
Wnt-10b/LRP6/Frizzled in the cells treated with TP-D, and the
GSK-3β levels decreased. These results indicated that adipogenesis
and lipogenesis were decreased by the inhibition of PPAR through
the upregulation of CCND1.
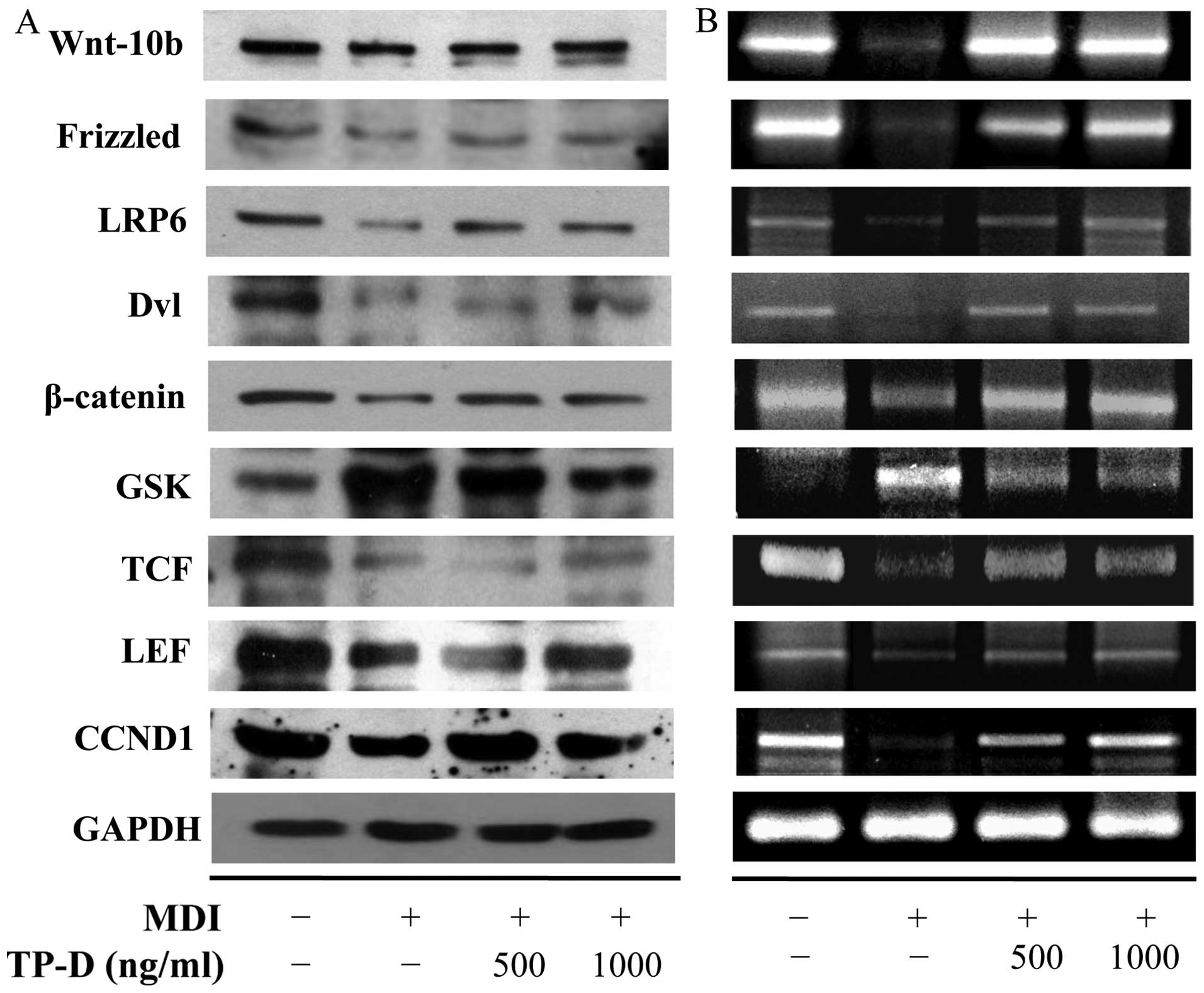 | Figure 6Expression levels of Wnt signaling
pathway-related proteins in differentiated 3T3-L1 adipocytes
following treatment with the tuna-derived peptide, D-I-V-D-K-I-E-I
(TP-D). Cells were treated with TP-D (125, 250, 500 or 1,000 ng/ml)
for 48 h. (A) Protein expression levels were measured by western
blot analysis. (B) cDNA was subjected to RT-PCR and mRNA expression
was analyzed by electrophoresis on a 1% (w/v) agarose gel and
staining with RedSafe Nucleic Acid staining solution. LRP6,
lipoprotein receptor-related protein-6; Dvl, dishevelled; GSK,
glycogen synthase kinase; TCF, T cell factor; LEF, lymphoid
enhancer-binding factor; CCND1, cyclin D1. |
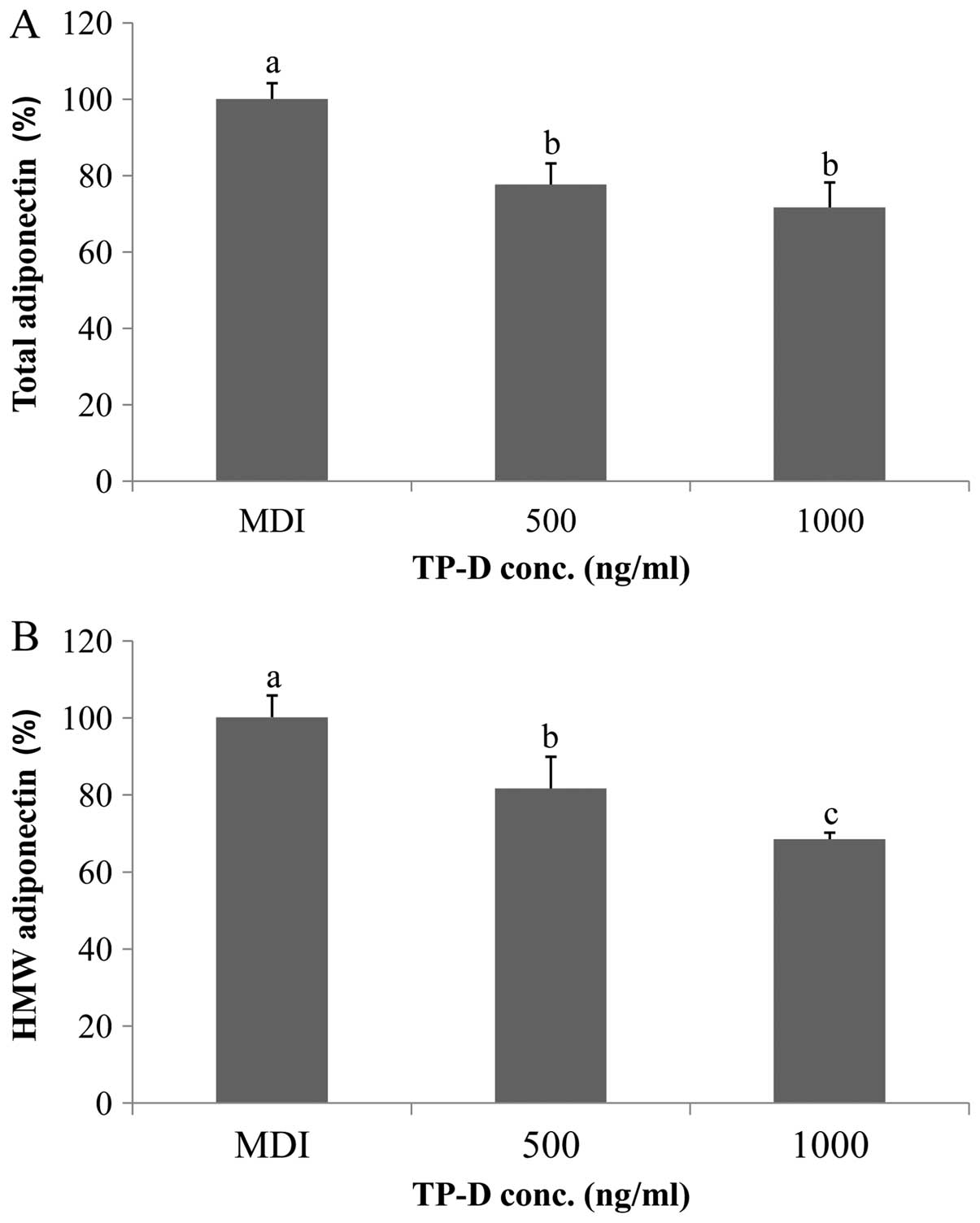
Effect of TP-D on the total and HMW
adiponectin levels during 3T3-L1 differentiation
In general, adiponectin forms multimers (25). HMW and total adiponectin are
markers of the key transcriptional regulator, PPAR-γ (26). In this study, we examined whether
TP-D affects the total and/or HMW adiponectin levels. As shown in
Fig. 7, the levels of total and
HMW adiponectin decreased upon treatment with TP-D, in a
dose-dependent manner, indicating that PPAR-γ synthesis was
inhibited.
Discussion
Obesity is the state in which excess body fat
accumulates, and it is directly caused by an increase in the number
and size of abnormal adipocytes. Various attempts to prevent and
manage obesity have been reported (27). The proliferation of adipocytes is
closely associated with adipogenesis and lipogenesis, the processes
through which pre-adipocytes differentiate into adipocytes
(28). Several transcription
factors regulate this process directly or indirectly; of these,
PPAR-γ and C/EBPs are important factors in controlling adipogenesis
during the early stages of differentiation. Therefore, it is
important to control these activities during early adipogenesis in
order to prevent obesity.
Recently, it has been suggested that food derived
from biologically active substances can prevent obesity. Tuna is a
high-protein food with anticancer and anti-atherosclerotic effects,
and is known to reduce blood cholesterol levels. Boiled tuna
extract contains carnosine and taurine, collagen-derived proteins,
and a number of free amino acids (29). However, boiled tuna extract is
typically considered a byproduct, and few studies on the material
have been published to date (30,31). In this study, we evaluated the
anti-obesity effects of a peptide of boiled tuna extract (sequence
D-I-V-D-K-I-E-I; termed TP-D) in 3T3-L1 pre-adipocytes. We found
that treatment of the cells with the peptide inhibited
differentiation and decreased glucose uptake and TG levels.
During the differentiation of 3T3-L1 pre-adipocytes
into adipocytes, the consumption of glucose increases and TGs
accumulate. Upon the induction of the expression of PPAR and C/EBP,
inducers of early differentiation, SREBP-1 is activated via SOCS-3.
SREBP-1 is directly involved in lipogenesis and adipogenesis.
During PPAR expression, LXR (a major nuclear receptor involved in
the regulation of lipogenensis) is activated by oxysterol, a
process that is also involved in SREBP expression and that may also
mediate SREBP-independent lipogenesis by activating fatty acid
translocase (FAT)/CD36) (12–16), an absorption carrier. Apart from
the traditional adipogenic pathway, the inhibition of lipogenesis
through the Wnt pathway is an anti-obesity effect. During the
formation of adipocytes, the canonical Wnt signaling pathway is
activated in pre-adipocytes; however, its activity decreases as
cells differentiate (32). The
main component of the canonical Wnt signaling pathway is β-catenin,
which has a variety of functions during the growth and
differentiation of several cell types (33). β-catenin activates LEF/TCF
transcription factors and, in the absence of the Wnt signal
transmission, is decomposed by the AXIN/GSK-3β/APC complex. The
activation of the Wnt/β-catenin pathway through the inhibition of
PPAR-γ is essential to the inhibition of the differentiation of
pre-adipocytes. CCND1 is the direct target of the β-catenin/LEF
complex and binds to the complex (34). CCND1 also inhibits PPAR-γ
activation through ligand binding; the mechanisms involve direct
interaction with PPAR-γ. Thus, as the inhibition of lipogenic and
adipogenic pathways reduces obesity, it is crucial to inhibit
C/EBPs and PPAR-γ and activate the Wnt/β-catenin pathway. In the
present study, we found that a peptide derived from boiled tuna
inhibited C/EBPs and PPAR-γ expression and activated the
Wnt/β-catenin pathway; as a result, the process through which
pre-adipocytes differentiate into fat globule cells was also
inhibited.
In conclusion, the findings of our present study
indicate that the tuna peptide, TP-D, may prove to be an effective
strategy with which to reduce glucose uptake and TG levels and to
prevent the adipocyte differentiation of 3T3-L1 cells.
Acknowledgments
This study was part of the project entitled
ʽFunctional materials and foods using fisheries by-productsʼ,
funded by the Ministry of Oceans and Fisheries, Korea
(20130279).
References
|
1
|
Kim DM, Choi HR, Park A, Shin SM, Bae KH,
Lee SC, Kim IC and Kim WK: Retinoic acid inhibits adipogenesis via
activation of Wnt signaling pathway in 3T3-L1 preadipocytes.
Biochem Biophys Res Commun. 434:455–459. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ordovas JM and Mooser V: Nutrigenomics and
nutrigenetics. Curr Opin Lipidol. 15:101–108. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ahn J, Lee H, Kim S and Ha T:
Curcumin-induced suppression of adipogenic differentiation is
accompanied by activation of Wnt/β-catenin signaling. Am J Physiol
Cell Physiol. 298:C1510–C1516. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Symposium of obesity. The ministry of
health welfare korea; 2006
|
|
5
|
Dani C, Smith AG, Dessolin S, Leroy P,
Staccini L, Villageois P, Darimont C and Ailhaud G: Differentiation
of embryonic stem cells into adipocytes in vitro. J Cell Sci.
110:1279–1285. 1997.PubMed/NCBI
|
|
6
|
Reya T and Clevers H: Wnt signalling in
stem cells and cancer. Nature. 434:843–850. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Moon RT, Kohn AD, De Ferrari GV and Kaykas
A: WNT and beta-catenin signalling: diseases and therapies. Nat Rev
Genet. 5:691–701. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Logan CY and Nusse R: The Wnt signaling
pathway in development and disease. Annu Rev Cell Dev Biol.
20:781–810. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu J and Farmer SR: Regulating the
balance between peroxisome proliferator-activated receptor gamma
and beta-catenin signaling during adipogenesis. A glycogen synthase
kinase 3beta phosphor-ylation-defective mutant of beta-catenin
inhibits expression of a subset of adipogenic genes. J Biol Chem.
279:45020–45027. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Moldes M, Zuo Y, Morrison RF, Silva D,
Park BH, Liu J and Farmer SR: Peroxisome-proliferator-activated
receptor gamma suppresses Wnt/beta-catenin signalling during
adipogenesis. Biochem J. 376:607–613. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim YM, Kim EY, Kim IH and Nam TJ: Peptide
derived from desalinated boiled tuna extract inhibits adipogenesis
through the downregulation of C/EBP-α and PPAR-γ in 3T3-L1
adipocytes. Int J Mol Med. 35:1362–1368. 2015.PubMed/NCBI
|
|
12
|
Browning JD and Horton JD: Molecular
mediators of hepatic steatosis and liver injury. J Clin Invest.
114:147–152. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Larter CZ and Farrell GC: Insulin
resistance, adiponectin, cytokines in NASH: which is the best
target to treat? J Hepatol. 44:253–261. 2006. View Article : Google Scholar
|
|
14
|
Mitro N, Mak PA, Vargas L, Godio C,
Hampton E, Molteni V, Kreusch A and Saez E: The nuclear receptor
LXR is a glucose sensor. Nature. 445:219–223. 2007. View Article : Google Scholar
|
|
15
|
Horton JD, Goldstein JL and Brown MS:
SREBPs: activators of the complete program of cholesterol and fatty
acid synthesis in the liver. J Clin Invest. 109:1125–1131. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou J, Febbraio M, Wada T, Zhai Y, Kuruba
R, He J, Lee JH, Khadem S, Ren S, Li S, et al: Hepatic fatty acid
transporter Cd36 is a common target of LXR, PXR, and PPARgamma in
promoting steatosis. Gastroenterology. 134:556–567. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ueki K, Kondo T and Kahn CR: Suppressor of
cytokine signaling 1 (SOCS-1) and SOCS-3 cause insulin resistance
through inhibition of tyrosine phosphorylation of insulin receptor
substrate proteins by discrete mechanisms. Mol Cell Biol.
24:5434–5446. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Musso G, Gambino R and Cassader M: Recent
insights into hepatic lipid metabolism in non-alcoholic fatty liver
disease (NAFLD). Prog Lipid Res. 48:1–26. 2009. View Article : Google Scholar
|
|
19
|
Klaus S, Ely M, Encke D and Heldmaier G:
Functional assessment of white and brown adipocyte development and
energy metabolism in cell culture. J Cell Sci. 108:3171–3180.
1995.
|
|
20
|
Stephens JM and Pekala PH: Transcriptional
repression of the GLUT4 and C/EBP genes in 3T3-L1 adipocytes by
tumor necrosis factor-alpha. J Biol Chem. 266:21839–21845.
1991.PubMed/NCBI
|
|
21
|
Qin L, Chen Y, Niu Y, Chen W, Wang Q, Xiao
S, Li A, Xie Y, Li J, Zhao X, et al: A deep investigation into the
adipogenesis mechanism: profile of microRNAs regulating
adipogenesis by modulating the canonical Wnt/beta-catenin signaling
pathway. BMC Genomics. 11:3202010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cadigan KM and Liu YI: Wnt signaling:
Complexity at the surface. J Cell Sci. 119:395–402. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ross SE, Hemati N, Longo KA, Bennett CN,
Lucas PC, Erickson RL and MacDougald OA: Inhibition of adipogenesis
by Wnt signaling. Science. 289:950–953. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang C, Pattabiraman N, Zhou JN, Fu M,
Sakamaki T, Albanese C, Li Z, Wu K, Hulit J, Neumeister P, et al:
Cyclin D1 repression of peroxisome proliferator-activated receptor
gamma expression and transactivation. Mol Cell Biol. 23:6159–6173.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Crouch E, Persson A, Chang D and Heuser J:
Molecular structure of pulmonary surfactant protein D (SP-D). J
Biol Chem. 269:17311–17319. 1994.PubMed/NCBI
|
|
26
|
McCormack FX, Pattanajitvilai S, Stewart
J, Possmayer F, Inchley K and Voelker DR: The Cys6 intermolecular
disulfide bond and the collagen-like region of rat SP-A play
critical roles in interactions with alveolar type II cells and
surfactant lipids. J Biol Chem. 272:27971–27979. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Choi TB and Kim YJ: Effect of obesity
reduction by exercise therapy and drug therapy. Kor J Aesthet
Cosmetol. 5:133–143. 2010.
|
|
28
|
Kim HO, Hwang YI and Kim GN: Anti-obese
function of demethoxycurcumin through suppression of adipogenesis
in 3T3-L1 cells. Kor J Aesthet Cosmetol. 11:29–34. 2013.
|
|
29
|
Lee HS, Kim HJ, Choi JI, Kim JH, Kim JG,
Chun BS, Ahn DH, Chung YJ, Kim YJ, Byun MW and Lee JW: Antioxidant
activity of the ethanol extract from cooking drips of Thunnus
thynnus by gamma irradiation. J Korean Soc Food Sci Nutr.
37:810–814. 2008. View Article : Google Scholar
|
|
30
|
Carroll KK: Biological effects of fish
oils in relation to chronic disease. Lipids. 21:731–732. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mehta J, Lopez LM and Wargovich T:
Eicosapentaenoic acid: its relevance in atherosclerosis and
coronary artery disease. Am J Cardiol. 59:155–159. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bennett CN, Ross SE, Longo KA, Bajnok L,
Hemati N, Johnson KW, Harrison SD and Macdougald OA: Regulation of
Wnt signaling during adipogenesis. J Biol Chem. 277:30998–31004.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Willert K and Nusse R: Beta-catenin: a key
mediator of Wnt signaling. Curr Opin Genet Dev. 8:95–102. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shtutman M, Zhurinsky J, Simcha I,
Albanese C, D’Amico M, Pestell R and Ben-Ze’ev A: The cyclin D1
gene is a target of the beta-catenin/LEF-1 pathway. Proc Natl Acad
Sci USA. 96:5522–5527. 1999. View Article : Google Scholar : PubMed/NCBI
|