Introduction
The neuropeptide substance P (SP) is an important
mediator of neurogenic inflammation within the central and
peripheral nervous systems. SP has been implicated in pain, and
also plays important roles in cancer (e.g., tumor cell
proliferation, anti-apoptotic effects on tumor cells, angiogenesis,
tumor cell invasion and metastasis) (1–8).
SP is released from primary afferent nociceptors and sympathetic
post-ganglionic neurons, and activates neighboring receptors,
thereby triggering and spreading activation (6–11).
Moreover, SP has been shown to induce the expression of
pro-inflammatory cytokines, such as interleukin (IL)-6 and IL-8
(12,13), which are involved in the
pathogenesis of several disorders of the human brain (such as
multiple sclerosis, dementia complex, and Alzheimer's disease)
(14), although it is currently a
matter of debate as to whether SP plays a pathogenic or a
protective role in these disorders. Previous research has indicated
that the activation of an SP receptor [neurokinin-1 receptor
(NK-1R)] elicits signals affecting the gene expression of some
inflammatory cytokines (13). In
addition, it has been reported that nanomolar concentrations of SP
potently trigger the activation of nuclear factor-κB (NF-κB), an
important transcriptional activator, which regulates the production
of various cytokines and other proinflammatory mediators (13).
The mitogen-activated protein kinase (MAPK) family
of protein Ser/Thr kinases consists of at least three major
subfamilies: i) the p42/44 MAPKs, which are also known as the
extracellular signal-regulated kinases (ERKs)1/2; ii) the c-Jun
NH2-terminal kinase/stress-activated protein kinases
(JNK/SAPKs), including p46 JNK-1 and p54 JNK2; and iii) the p38
MAPK subfamily. MAPKs are activated under conditions of stress in
response to a number of extracellular stimuli, including oxidative
stress. Among these, the phosphorylation of p38 MAPK is induced in
the dorsal horn of the spinal cord and in the dorsal root ganglia
following peripheral nerve injury or inflammation (15–17), and the phosphorylation of ERK1/2
is induced by inflammatory stimuli in the trigeminal nucleus and
the dorsal horn (16,18,19).
The release of glutamate and SP from the primary
afferents, activates N-methyl-D-asparate (NMDA) receptors
and NK-1Rs, respectively, thereby resulting in the increase of
intracellular calcium concentrations in the dorsal horn neurons,
the activation of phospholipase A2 (PLA2)
(20,21) and the production of prostaglandins
by constitutively expressed spinal cyclooxygenase-2 (COX-2)
(22). Ketamine, an intravenous
anesthetic agent, functions as a competitive antagonist of the
excitatory neurotransmission NMDA receptor (23) and also antagonizes the NK-1R by
interfering with the binding of SP (24). Notably, the anti-inflammatory
effect of ketamine has also been demonstrated in various animal
models, where it was observed that ketamine markedly suppressed the
production of tumor necrosis factor-α (TNF-α) and IL-6 following
the stimulation of macrophages and peripheral leucocytes by
lipopolysaccharide (LPS) (25).
Moreover, it has been reported that ketamine inhibits the systemic
production of inflammatory molecules by inhibiting the NF-κB
signaling pathway (26). Based on
these findings, we hypothesized that ketamine may also act on NK-1R
(SP) and exert anti-inflammatory effects by modulating the MAPK and
NF-κB signaling pathways.
Thus, in the present study, we examined the
anti-inflammatory effects of ketamine on the SP-induced activation
of the human astrocytoma cell line, U373MG, which expresses high
levels of NK-1R.
Materials and methods
Materials
The U373MG cell line (Uppsala; ECACC 08061901) was
purchased from the European Collection of Authenticated Cell
Cultures (ECACC; Salisbury, UK). SP was obtained from Sigma-Aldrich
(St. Louis, MO, USA); ketamine was purchased from Daiichi Sankyo
Co., Ltd. (Tokyo, Japan); minimum essential medium (MEM),
non-essential amino acids (NEAAs), sodium pyruvate and fetal bovine
serum (FBS) were puchased from Gibco BRL Life Technologies (Grand
Island, NY, USA). RIPA buffer with protease inhibitor cocktail,
sample buffer solution with reducing reagent (6X) for SDS-PAGE,
running buffer solution (10X) for SDS-PAGE, Blocking One and
Protein Ladder One Multi-color (Broad Range) for SDS-PAGE were
obtained from Nacalai Tesque, Inc. (Kyoto, Japan). A BCA protein
assay reagent kit and enhanced chemiluminescence reagent,
SuperSignal West Dura were purchased from Thermo Fisher Scientific
(Rockford, IL, USA). Mini-PROTEAN® TGX™ precast gel and
Trans-Blot® Turbo™ Mini PVDF transfer packs were
purchased from Bio-Rad Laboratories, Inc. (Hercules, CA, USA), and
a Cytometric Bead Array (Human Inflammatory Cytokine kit) was
obtained from BD Biosciences (San Jose, CA, USA).
Antibodies
Anti-phospho-ERK1/2 MAPK (Thr202/Tyr204) rabbit
antibody (#9101), anti-ERK1/2 MAPK rabbit antibody (#9102),
anti-phospho-NF-κB p65 (Ser536) rabbit monoclonal antibody (mAb;
#3033), and anti-NF-κB p65 rabbit mAb (#8242) were purchased from
Cell Signaling Technology, Inc. (Danvers, MA, USA); anti-phospho
p38 MAPK rabbit antibody (Thr180/Tyr182; V121A) was obtained from
Promega Corp. (Madison, WI, USA); anti-p38 MAPK (p38/SAPK2α;
612168) mouse mAb was purchased from BD Biosciences. Horseradish
peroxidase (HRP)-conjugated goat anti-rabbit IgG (AP132P) and
HRP-conjugated goat anti-mouse IgG/IgM (AP308P) were obtained from
Chemicon International (Temecula, CA, USA).
Cell culture
The U373MG cells were cultured in MEM supplemented
with 1% (v/v) penicillin/streptomycin, NEAA, 1 mM sodium pyruvate
and 10% heat-inactivated FBS. The cells were maintained at 37°C in
a 5% CO2 humidified atmosphere.
Western blot analysis
The U373MG cells were plated into 12-well tissue
culture plates at a density of 2×105 cells/well and
incubated in MEM supplemented with 10% FBS for 12 h, followed by
incubation in MEM supplemented with 0.5% FBS for 12 h at 37°C.
Subsequently, the cells were incubated with ketamine (0.1 and 1 mM)
for 60 min, and then stimulated with SP (100 nM) for 10 min.
Thereafter, the cells were washed 3 times with ice-cold
phosphate-buffered saline (PBS) and lysed in 0.1 ml of RIPA buffer
(50 mmol/l Tris-HCl pH 7.6, 150 mmol/l NaCl, 1% Nonidet P40, 0.5%
sodium deoxycholate, protease inhibitor cocktail and 0.1% SDS). The
protein concentrations of cell lysates were measured using the
Bradford assay (Thermo Fisher Scientific). The lysates were mixed
with SDS-PAGE sample buffer (62.5 mM Tris-HCl, pH 6.8, 2% SDS, 10%
glycerol, 0.05% bromphenol blue, and 5% 2-mercaptoethanol), and
applied to SDS-PAGE 10% gels (Mini-PROTEAN TGX precast gel; 15
μg protein/lane). Thereafter, the separated proteins were
electroblotted onto polyvinylidine fluoride membranes (Trans-Blot
Turbo Mini PVDF Transfer packs). Following incubation with Blocking
One (Nacalai Tesque, Inc.), the blots were probed with a 1,000-fold
dilution of rabbit anti-phospho ERK1/2, anti-phospho NF-κB or
anti-phospho p38 MAPK antibody, and further probed with a
2,000-fold dilution of HRP-conjugated goat anti-rabbit IgG/IgM. The
signals were detected by SuperSignal West Pico/Dura
Chemiluminescent substrate (Thermo Fisher Scientific), and
quantified using an LAS-3000 luminescent image analyzer and
MultiGauge software (both from Fujifilm, Tokyo, Japan). Thereafter,
the antibody was stripped using WB Stripping Solution Strong
(Naclai Tesque, Inc.) at room temperature for 15 min. The blots
were probed with a 1,000-fold dilution of rabbit anti-ERK1/2,
anti-NF-κB or anti-p38 MAPK antibody, and further probed with a
2,000-fold dilution of HRP-conjugated goat anti-rabbit IgG/IgM or
HRP-conjugated goat anti-mouse IgG/IgM.
Quantification of IL-6 and IL-8
levels
The U373MG cells were plated into 12-well tissue
culture plates at a density of 2×105 cells/well and
incubated in MEM supplemented with 10% FBS for 12 h, which was
followed by incubation in MEM supplemented with 0.5% FBS for 12 h
at 37°C. The cells were then incubated with ketamine (1 mM) for 60
min, and stimulated with SP (100 nM) for 24 h. The culture
supernatants were recovered, centrifuged at 12,000 × g for 10 min,
and the levels of IL-6 and IL-8 in the medium were measured using a
Cytometric Bead Array Human Inflammatory Cytokine kit (BD
Biosciences), according to the manufacturer's instructions.
Statistical analysis
Data are expressed as the means ± SD, and analyzed
for significant differences using a one-way ANOVA followed by a
multiple comparison test using GraphPad Prism version 6.0 for
Windows (GraphPad Software, San Diego, CA, USA). A P-value <0.05
was considered to indicate a statistically significant
difference.
Results
Suppression of SP-induced ERK1/2
activation by ketamine
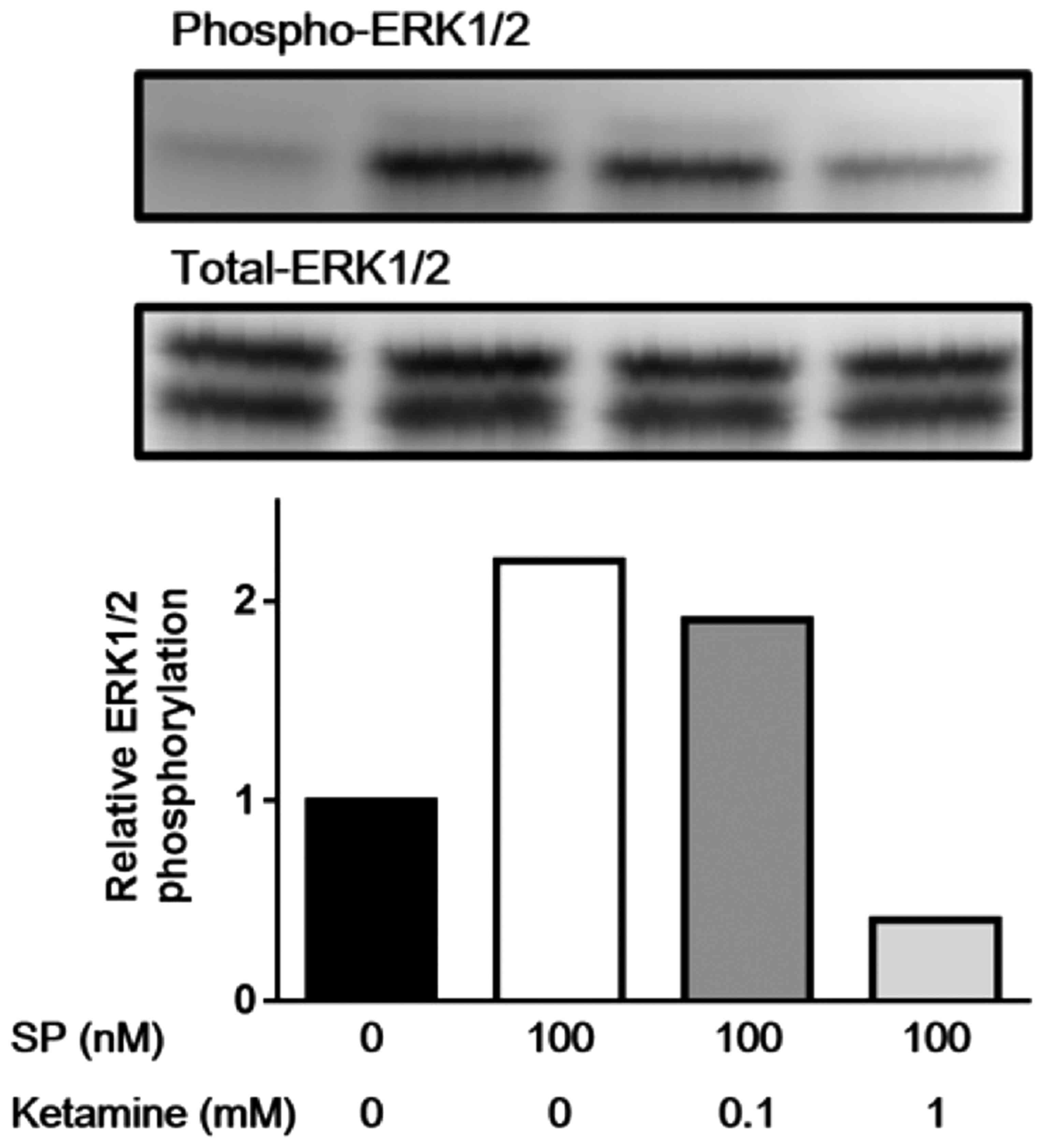
Firstly, we examined the dose-dependent effect of
ketamine on the phosphorylation of ERK1/2. As shown in Fig. 1, SP stimulation (100 nM) markedly
induced the phosphorylation of ERK1/2 in the U373MG cells, and
ketamine suppressed the SP-induced phosphorylation of ERK1/2 in a
dose-dependent manner; the phosphorylation of ERK1/2 was only
minimally inhibited by 0.1 mM ketamine, whereas the concentration
of 1 mM ketamine caused marked inhibition. Thus, the concentration
of 1 mM ketamine was used to repeatedly evaluate the effect of
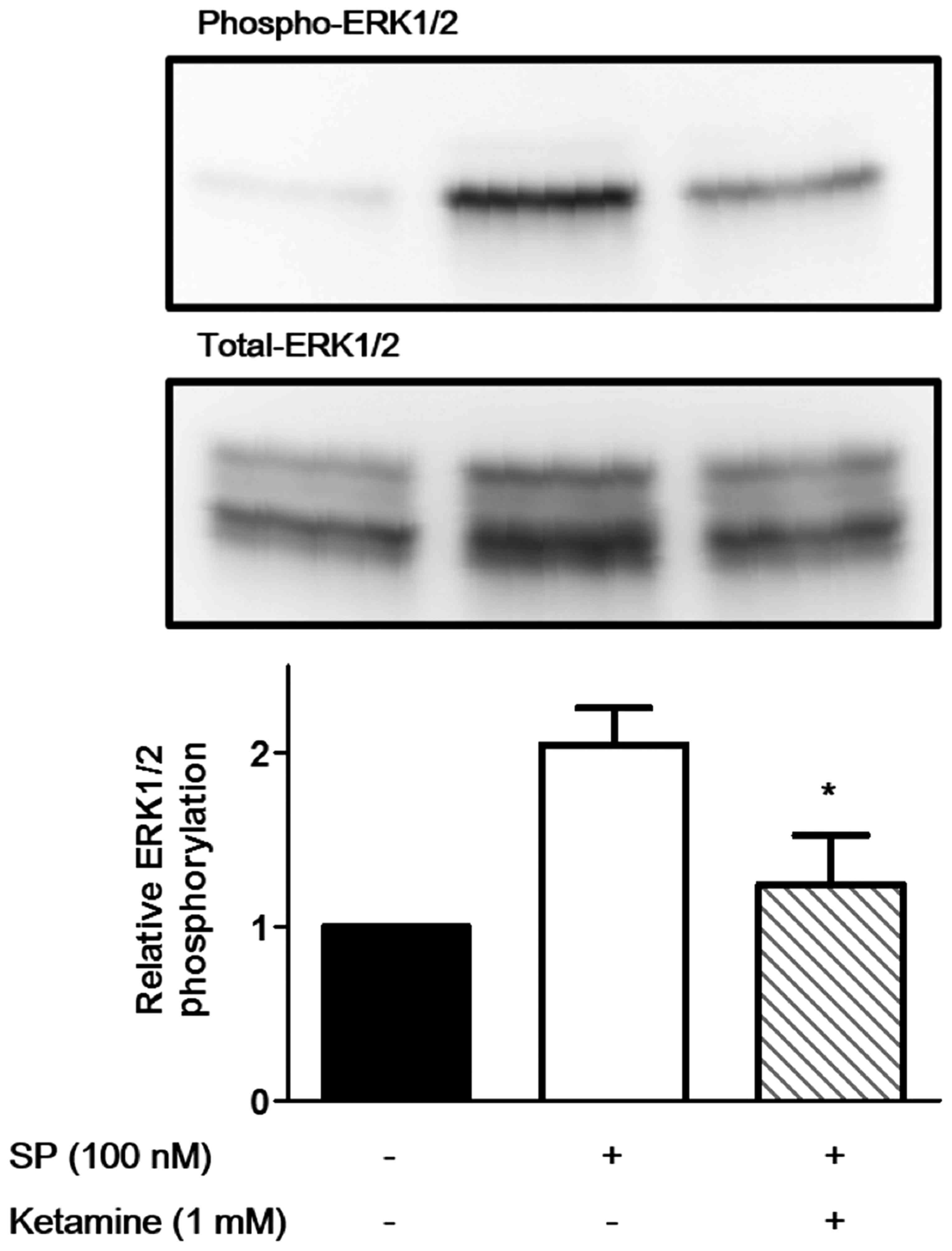
ketamine on the SP-induced phosphorylation of ERK1/2. As shown in
Fig. 2, 1 mM ketamine significantly suppressed
the SP-induced phosphorylation of ERK1/2 in the U373MG cells
(P<0.05).
Suppression of the SP-induced activation
of p38 MAPK and NF-κB by ketamine
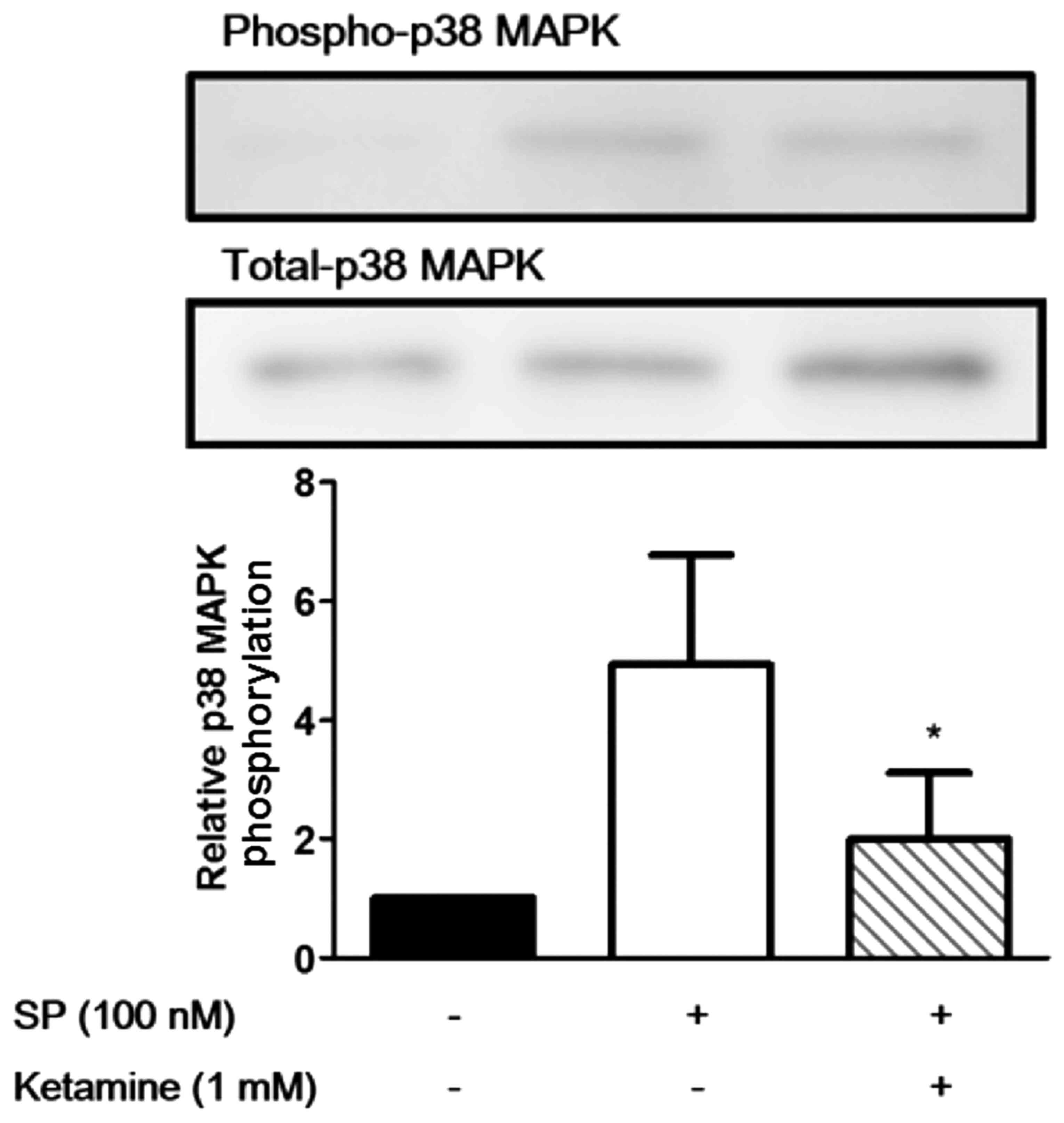
We then evaluated the effects of ketamine on the
phosphorylation of p38 MAPK and NF-κB. As shown in Fig. 3, stimulation of the U373MG cells
with SP (100 nM) markedly induced the phosphorylation of p38 MAPK,
and ketamine (1 mM) significantly suppressed the SP-induced
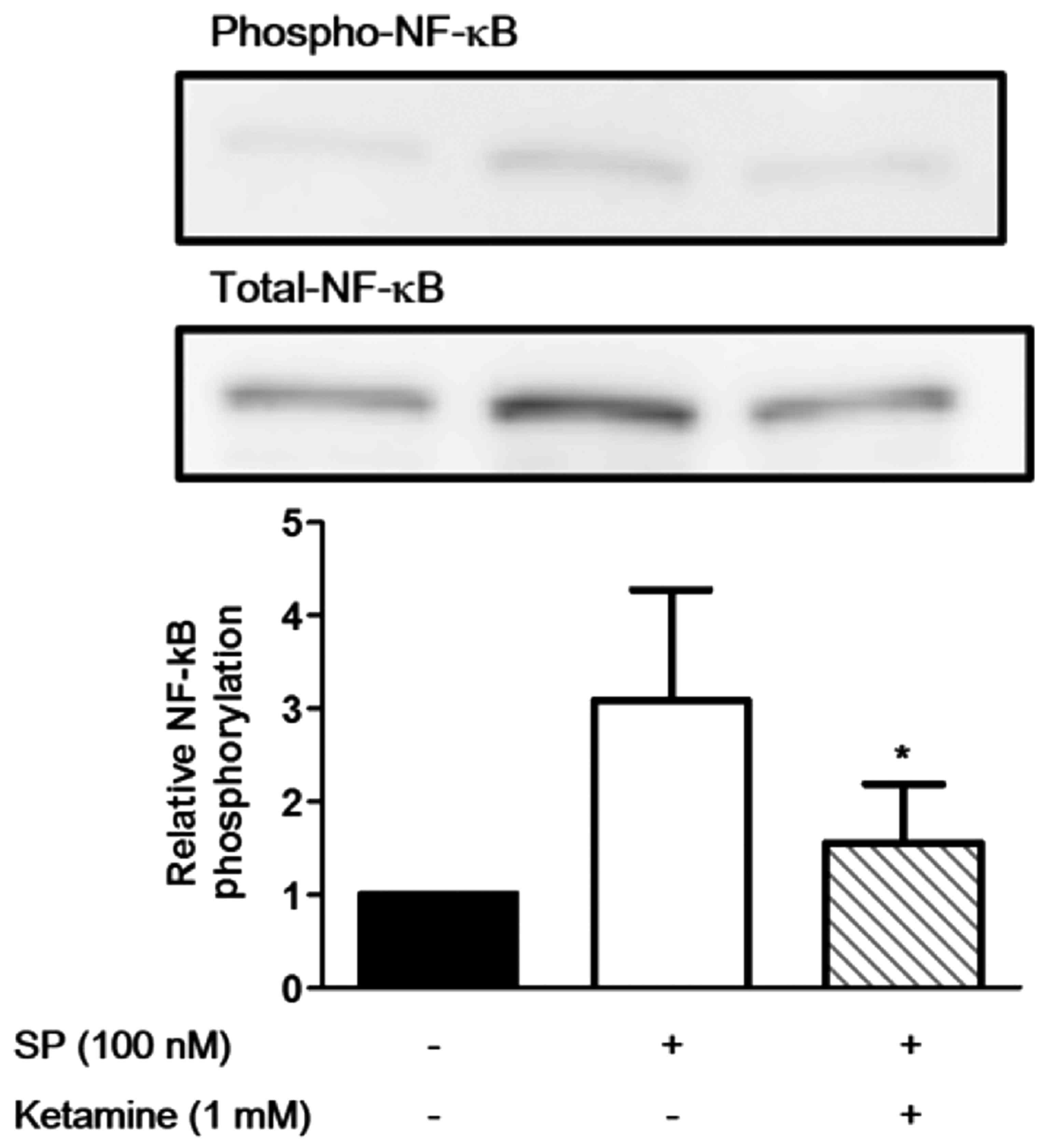
phosphorylation of p38 MAPK (P<0.05). Similarly, stimulation of
the U373MG cells with SP (100 nM) markedly induced the phospho
rylation of NF-κB, and ketamine (1 mM) significantly suppressed the
SP-induced phosphorylation of NF-κB (Fig. 4; P<0.05).
Effects of ketamine on the SP-induced
production of IL-6 and Il-8 by U373MG cells
Finally, we evaluated the effects of ketamine on the
production of IL-6 and IL-8 by U373MG cells. As shown in Fig. 5, the levels of IL-6 and IL-8 in
the cell culture supernatants were markedly increased following
stimulation with SP (100 nM). Notably, ketamine (1 mM)
significantly suppressed the production of both IL-6 and IL-8
(P<0.05).
Discussion
To the best of our knowledge, this is the first
study to demonstrate the effects of ketamine on SP-induced
inflammatory responses in U373MG glioblastoma/astrocytoma cells. In
the present study, we revealed that ketamine suppressed the
production of IL-6 and IL-8 in U373MG cells. Furthermore, ketamine
inhibited the SP-induced activation of ERK1/2, p38 MAPK and NF-κB.
Thus, ketamine may suppress the SP-induced activation (IL-6 and
IL-8 production) of U373MG cells by inhibiting the phosphorylation
of signaling molecules (namely ERK1/2, p38 MAPK and NF-κB), thereby
exerting anti-inflammatory effects.
U373MG astrocytoma cells express a functional
high-affinity SP receptor, NK-1R (27) and are capable of producing IL-6 in
response to stimulation with SP (12). Thus, this cell line is widely used
as an in vitro model in order to analyze the functions of
the NK-1R. Previously, it has been demonstrated that the plasma
levels of ketamine reached up to 25,800 ng/ml (108.4 μM) in
surgical patients 1 min after an intravenous injection of ketamine
at a dose of 2.0–2.2 mg/kg (28).
In the present study, we revealed that the phosphorylation of
ERK1/2 was only minimally inhibited by 0.1 mM ketamine, whereas the
dose of 1 mM ketamine caused a marked inhibition. Thus, it is
possible that clinically relevant (100 mM) or higher concentrations
of ketamine modulate the inflammatory responses of SP-stimulated
NK-1R-expressing cells in vivo by suppressing the activation
of signaling molecules (such as ERK1/2, p38 MAPK and NF-κB).
Neuroinflammation is involved in several diseases
affecting the central nervous system (CNS) in humans, such as
Parkinson's disease (PD) (29,30). As regards the pathogenesis of PD,
neuroinflammation is a common feature, which is primarily induced
by the long-term activation of glial cells (astrocytes and
microglia) in the brain (31). It
has also been demonstrated that activated glial cells produce
pro-inflammatory cytokines, which play roles in the initiation and
progression of neuroinflammation (29). Thus, anti-inflammatory drugs may
be expected to reduce the risk and delay the pathogenic process of
neuroinflammation by suppressing the activation of glial cells
(32). Ketamine has been
identified as a non-selective NMDA receptor antagonist. The
pharmacological actions of ketamine include the regulation of
inflammatory and immune responses in neural tissues, which leads to
decreases in the levels of proinflammatory cytokines including IL-6
and IL-8. Previously, ketamine has been reported to exert an
anti-inflammatory effect on LPS-stimulated macrophages in
vitro and in vivo by suppressing the MAPK pathways
(33,34). The glial cells of the CNS are
involved in regulating the immune response under conditions of
neuropathic pain (35) and
ketamine has been shown to reduce the LPS-induced production of
TNF-α and prostaglandin E2 by astrocytes (36). Moreover, ketamine exerts an
inhibitory effect on the activation of LPS-stimulated astrocytes by
suppressing NF-κB activation through the reduction of Toll-like
receptor 4 expression (26). As
regards the effect of ketamine on the NK-1R, Okamoto et al
demonstrated that ketamine inhibited NK-1R-mediated signaling by
interfering with the binding of SP using NK-1R-expressing
Xenopus oocytes using a whole-cell voltage clamp; however,
the binding site of ketamine in the NK-1R is probably different
from that of SP (24). Thus, the
precise mechanism through which ketamine modulates SP-induced
signaling remains unknown.
SP stimulates a number of intracellular signaling
molecules, including members of the MAPK family (ERK1/2 and p38
MAPK) via the action of NK-1R. In fact, SP has been demonstrated to
enhance the production of inflammatory chemokines by murine
macrophages through ERK/p38 MAPK-mediated NF-κB activation
(37). Moreover, it has been
previously demonstrated that SP induces the expression of IL-6
through the activation of p38 MAPK, ERK1/2 and NF-κB in the U373MG
astrocytoma cell line, which was also used in the present study
(14,38). Taken together, these observations
suggest that SP induces the production of cytokines/chemokines by
NK-1R-expressing cells through the NK-1R, a principal receptor for
SP, followed by the activation of signaling molecules (ERK1/2, p38
MAPK and NF-κB). Of note, ketamine may inhibit NK-1R-mediated
signaling by interfering with the binding of SP to the receptor
(24). Thus, ketamine may
suppress the SP-induced activation of signaling molecules (ERK1/2,
p38 MAPK and NF-κB), thereby suppressing the production of
cytokines/chemokines by exerting an inhibitory effect on the NK-1R
(interference of SP binding with the receptor).
In conclusion, our results demonstrate that ketamine
inhibits the SP-induced phosphorylation of MAPK (ERK1/2 and p38
MAPK) and NF-κB, and suppresses the production of the
proinflammatory cytokines IL-6 and IL-8. Moreover, the
anti-inflammatory effect of ketamine is potentially mediated
through the inhibition of signaling molecules (MAPK and NF-κB).
Thus, ketamine may modulate SP-induced inflammatory responses by
NK-1R-expressing cells through the suppression of signaling
molecules (such as ERK1/2, p38 MAPK and NF-κB); however, a detailed
examination of the anti-inflammatory effects of ketamine on glial
cells (such as astrocytes and microglia cells) is warranted in the
future.
Acknowledgments
The present study was supported by a Grant-in-Aid
for Scientific Research (grant no. 23592265) from the Japan Society
for the Promotion of Science.
References
|
1
|
Guha S, Eibl G, Kisfalvi K, Fan RS,
Burdick M, Reber H, Hines OJ, Strieter R and Rozengurt E:
Broad-spectrum G protein-coupled receptor antagonist,
[D-Arg1,D-Trp5,7,9,Leu11] SP: a dual inhibitor of growth and
angiogenesis in pancreatic cancer. Cancer Res. 65:2738–2745. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Esteban F, Muñoz M, González-Moles MA and
Rosso M: A role for substance P in cancer promotion and
progression: a mechanism to counteract intracellular death signals
following oncogene activation or DNA damage. Cancer Metastasis Rev.
25:137–145. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Muñoz M, Rosso M and Coveñas R: A new
frontier in the treatment of cancer: NK-1 receptor antagonists.
Curr Med Chem. 17:504–516. 2010. View Article : Google Scholar
|
|
4
|
Samsam M, Coveñas R, Ahangari R, Yajeya J,
Narváez JA and Tramu G: Simultaneous depletion of neurokinin A,
substance P and calcitonin gene-related peptide from the caudal
trigeminal nucleus of the rat during electrical stimulation of the
trigeminal ganglion. Pain. 84:389–395. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bang R, Sass G, Kiemer AK, Vollmar AM,
Neuhuber WL and Tiegs G: Neurokinin-1 receptor antagonists
CP-96,345 and L-733,060 protect mice from cytokine-mediated liver
injury. J Pharmacol Exp Ther. 305:31–39. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Muñoz M and Rosso M: The NK-1 receptor
antagonist aprepitant as a broad spectrum antitumor drug. Invest
New Drugs. 28:187–193. 2010. View Article : Google Scholar
|
|
7
|
Samsam M, Coveñas R, Csillik B, Ahangari
R, Yajeya J, Riquelme R, Narváez JA and Tramu G: Depletion of
substance P, neurokinin A and calcitonin gene-related peptide from
the contralateral and ipsilateral caudal trigeminal
nucleusfollowing unilateral electrical stimulation of the
trigeminal ganglion; apossible neurophysiological and
neuroanatomical link to generalized head pain. J Chem Neuroanat.
21:161–169. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Muñoz M and Coveñas R: Involvement of
substance P and the NK-1 receptor in cancer progression. Peptides.
48:1–9. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maeno H, Kiyama H and Tohyama M:
Distribution of the substance P receptor (NK-1 receptor) in the
central nervous system. Brain Res Mol Brain Res. 18:43–58. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Saffroy M, Beaujouan JC, Torrens Y,
Besseyre J, Bergström L and Glowinski J: Localization of tachykinin
binding sites (NK1, NK2, NK3 ligands) in the rat brain. Peptides.
9:227–241. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wolf SS, Moody TW, Quirion R and O'Donohue
TL: Biochemical characterization and autoradiographic localization
of central substance P receptors using [125I]physalaemin. Brain
Res. 332:299–307. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lieb K, Schaller H, Bauer J, Berger M,
Schulze-Osthoff K and Fiebich BL: Substance P and histamine induce
interleukin-6 expression in human astrocytoma cells by a mechanism
involving protein kinase C and nuclear factor-IL-6. J Neurochem.
70:1577–1583. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lieb K, Fiebich BL, Berger M, Bauer J and
Schulze-Osthoff K: The neuropeptide substance P activates
transcription factor NF-kappa B and kappa B-dependent gene
expression in human astrocytoma cells. J Immunol. 159:4952–4958.
1997.PubMed/NCBI
|
|
14
|
Horuk R, Martin AW, Wang Z, Schweitzer L,
Gerassimides A, Guo H, Lu Z, Hesselgesser J, Perez HD, Kim J, et
al: Expression of chemokine receptors by subsets of neurons in the
central nervous system. J Immunol. 158:2882–2890. 1997.PubMed/NCBI
|
|
15
|
Kim SY, Bae JC, Kim JY, Lee HL, Lee KM,
Kim DS and Cho HJ: Activation of p38 MAP kinase in the rat dorsal
root ganglia and spinal cord following peripheral inflammation and
nerve injury. Neuroreport. 13:2483–2486. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ji RR, Befort K, Brenner GJ and Woolf CJ:
ERK MAP kinase activation in superficial spinal cord neurons
induces prodynorphin and NK-1 upregulation and contributes to
persistent inflammatory pain hypersensitivity. J Neurosci.
22:478–485. 2002.PubMed/NCBI
|
|
17
|
Jin SX, Zhuang ZY, Woolf CJ and Ji RR: p38
mitogen-activated protein kinase is activated after a spinal nerve
ligation in spinal cord microglia and dorsal root ganglion neurons
and contributes to the generation of neuropathic pain. J Neurosci.
23:4017–4022. 2003.PubMed/NCBI
|
|
18
|
Huang WJ, Wang BR, Yao LB, Huang CS, Wang
X, Zhang P, Jiao XY, Duan XL, Chen BF and Ju G: Activity of p44/42
MAP kinase in the caudal subnucleus of trigeminal spinal nucleus is
increased following perioral noxious stimulation in the mouse.
Brain Res. 861:181–185. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ji RR, Baba H, Brenner GJ and Woolf CJ:
Nociceptive-specific activation of ERK in spinal neurons
contributes to pain hypersensitivity. Nat Neurosci. 2:1114–1119.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lazarewicz JW, Wroblewski JT and Costa E:
N-methyl-D-aspartate-sensitive glutamate receptors induce
calcium-mediated arachidonic acid release in primary cultures of
cerebellar granule cells. J Neurochem. 55:1875–1881. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim DK, Rordorf G, Nemenoff RA, Koroshetz
WJ and Bonventre JV: Glutamate stably enhances the activity of two
cytosolic forms of phospholipase A2 in brain cortical cultures.
Biochem J. 310:83–90. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen QR, Miyaura C, Higashi S, Murakami M,
Kudo I, Saito S, Hiraide T, Shibasaki Y and Suda T: Activation of
cytosolic phospholipase A2 by platelet-derived growth factor is
essential for cyclooxygenase-2-dependent prostaglandin E2 synthesis
in mouse osteoblasts cultured with interleukin-1. J Biol Chem.
272:5952–5958. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zeilhofer HU, Swandulla D, Geisslinger G
and Brune K: Differential effects of ketamine enantiomers on NMDA
receptor currents in cultured neurons. Eur J Pharmacol.
213:155–158. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Okamoto T, Minami K, Uezono Y, Ogata J,
Shiraishi M, Shigematsu A and Ueta Y: The inhibitory effects of
ketamine and pentobarbital on substance p receptors expressed in
Xenopus oocytes. Anesth Analg. 97:104–110. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mazar J, Rogachev B, Shaked G, Ziv NY,
Czeiger D, Chaimovitz C, Zlotnik M, Mukmenev I, Byk G and
Douvdevani A: Involvement of adenosine in the antiinflammatory
action of ketamine. Anesthesiology. 102:1174–1181. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu Y, Li W and Zhou C, Lu F, Gao T, Liu Y,
Cao J, Zhang Y, Zhang Y and Zhou C: Ketamine inhibits
lipopolysaccharide-induced astrocytes activation by suppressing
TLR4/NF-ĸB pathway. Cell Physiol Biochem. 30:609–617. 2012.
View Article : Google Scholar
|
|
27
|
Heuillet E, Ménager J, Fardin V, Flamand
O, Bock M, Garret C, Crespo A, Fallourd AM and Doble A:
Characterization of a human NK1 tachykinin receptor in the
astrocytoma cell line U 373 MG. J Neurochem. 60:868–876. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Domino EF, Zsigmond EK, Domino LE, Domino
KE, Kothary SP and Domino SE: Plasma levels of ketamine and two of
its metabolites in surgical patients using a gas chromatographic
mass fragmentographic assay. Anesth Analg. 61:87–92. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Niranjan R: The role of inflammatory and
oxidative stress mechanisms in the pathogenesis of Parkinson's
disease: focus on astrocytes. Mol Neurobiol. 49:28–38. 2014.
View Article : Google Scholar
|
|
30
|
Simões AP, Duarte JA, Agasse F, Canas PM,
Tomé AR, Agostinho P and Cunha RA: Blockade of adenosine A2A
receptors prevents interleukin-1β-induced exacerbation of neuronal
toxicity through a p38 mitogen-activated protein kinase pathway. J
Neuroinflammation. 9:2042012. View Article : Google Scholar
|
|
31
|
Minagar A, Shapshak P, Fujimura R, Ownby
R, Heyes M and Eisdorfer C: The role of macrophage/microglia and
astrocytes in the pathogenesis of three neurologic disorders:
HIV-associated dementia, Alzheimer disease, and multiple sclerosis.
J Neurol Sci. 202:13–23. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
García-Bueno B, Madrigal JL, Lizasoain I,
Moro MA, Lorenzo P and Leza JC: Peroxisome proliferator-activated
receptor gamma activation decreases neuroinflammation in brain
after stress in rats. Biol Psychiatry. 57:885–894. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu GJ, Chen TL, Ueng YF and Chen RM:
Ketamine inhibits tumor necrosis factor-alpha and interleukin-6
gene expressions in lipopolysaccharide-stimulated macrophages
through suppression of Toll-like receptor 4-mediated c-Jun
N-terminal kinase phosphorylation and activator protein-1
activation. Toxicol Appl Pharmacol. 228:105–113. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chang HC, Lin KH, Tai YT, Chen JT and Chen
RM: Lipoteichoic acid-induced TNF-α and IL-6 gene expressions and
oxidative stress production in macrophages are suppressed by
ketamine through downregulating Toll-like receptor 2-mediated
activation oF ERK1/2 and NFκB. Shock. 33:485–492. 2010.
|
|
35
|
Milligan ED and Watkins LR: Pathological
and protective roles of glia in chronic pain. Nat Rev Neurosci.
10:23–36. 2009. View
Article : Google Scholar :
|
|
36
|
Shibakawa YS, Sasaki Y, Goshima Y, Echigo
N, Kamiya Y, Kurahashi K, Yamada Y and Andoh T: Effects of ketamine
and propofol on inflammatory responses of primary glial cell
cultures stimulated with lipopolysaccharide. Br J Anaesth.
95:803–810. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sun J, Ramnath RD, Zhi L, Tamizhselvi R
and Bhatia M: Substance P enhances NF-kappaB transactivation and
chemokine response in murine macrophages via ERK1/2 and p38 MAPK
signaling pathways. Am J Physiol Cell Physiol. 294:C1586–C1596.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fiebich BL, Schleicher S, Butcher RD,
Craig A and Lieb K: The neuropeptide substance P activates p38
mitogen-activated protein kinase resulting in IL-6 expression
independently from NF-kappaB. J Immunol. 165:5606–5611. 2000.
View Article : Google Scholar : PubMed/NCBI
|