Introduction
Renal disease can advance to tubulointerstitial
fibrosis and has been proposed as a final common pathway to
end-stage renal disease. Uunilateral ureteral obstruction (UUO)
results in a number of pathophysiological and morphological changes
in the renal parenchyma, including interstitial fibrosis,
inflammation and apoptotic changes in tubular and interstitial
cells (1). Among the mechanisms
responsible for renal fibrogenesis, the transforming growth
factor-β (TGF-β) signalling pathway is known to play a pivotal role
in renal tubulointerstitial fibrosis. Through the autocrine and
paracrine stimulation of cells by TGF-β1, connective tissue growth
factor (CTGF) is released and synthesised, which plays a role in
fibrogenesis (2).
Reactive oxygen species (ROS) are also believed to
play a key role in fibrosis. The renal tubulointerstitium requires
high oxygen consumption for active solute transport and
reabsorption, and the tubulointerstitium is susceptible to
oxidative stress. Increased ROS accumulation leads to renal
fibrosis and is involved in various kidney diseases (3). Oxidative stress caused by ROS
mediates TGF-β1-induced cellular responses in various cells
(4,5). Maintaining the redox balance and
using antioxidants can delay the progression of renal interstitial
fibrosis.
The therapeutic potential of Sirt1 activators has
been explored in several diseases. Mammalian sirtuin 1 (Sirt1), a
nicotinamide adenine dinucleotide (NAD+)-dependent
protein class III deacetylase, is widely expressed throughout
nearly all mammalian organs. Sirt1 is a key modulator of cell
proliferation, hormone responses, stress responses, apoptosis and
cell metabolism (6,7). A recent study demonstrated that
Sirt1 activation attenuates renal fibrosis by inhibiting the
TGF-β1/Smad3 pathway (8). Sirt1
also interacts with TGF-β1 signalling and inhibits apoptosis in
TGF-β1-treated mesangial cells (9). Moreover, a recent study highlighted
the interaction of Sirt1 with CTGF in cancer (10). However, it is unclear as to
whether Sirt1 activation exerts protective effects against
UUO-induced tubulointerstitial fibrosis by suppressing renal
oxidative stress and the TGF-β1/CTGF signalling pathway.
Thus, in this study, we investigated the effects of
the Sirt1 activator, SRT1720, a small molecule activator of Sirt1
that is more potent than resveratrol (11), on UUO-induced renal
tubulointerstitial fibrosis and we aimed to determine whether these
effects are mediated by the inhibition of oxidative stress and the
TGF-β1/CTGF pathway.
Materials and methods
Animal model and experimental design
Male 12-week-old CD-1 mice (Charles River
Laboratories, Vital River, Beijing, China) were housed in the
animal facilities at the Hebei Medical University with free access
to food and water. UUO was created by the ligation of the left
ureter with 4–0 silk through a left flank incision; during the
procedure, the animals were under anesthesia induced with sodium
pentobarbital. Sham-operated mice underwent the same procedure,
except that the ureter was not ligated. The animals were randomly
divided into 3 groups as follows: the sham-operated (sham) group,
the group subjected to UUO (UUO) and the group subjected to UUO and
treated with SRT1720 (SRT1720) (n=6/group). The Sirt1 activator,
SRT1720 (100 mg/kg) (12)
(Selleck Chemicals, Houston, TX, USA), was administered by
intraperitoneal injection for 3 days prior to the operation and was
continued for 7 days after UUO. The animals were sacrificed 7 days
after UUO or sham operation, and the kidneys were harvested. All
procedures were performed in compliance with the guidelines
established by the Ethics Review Committee for Animal
Experimentation of Hebei Medical University. This study was
specifically approved by the Ethics Review Committee for Animal
Experimentation of Hebei Medical University (Permit no.
2013-0666).
Histological analysis
The kidneys were fixed in 4% paraformaldehyde
overnight and embedded in paraffin. Sections (2-µm-thick)
were stained with Masson's trichrome (Sigma, St. Louis, MO, USA).
The fibrotic area was measured quantitatively using the HPIAS-2000
image analysis software (Champion Image Co., Wuhan, China). The
results were expressed as the percentage area of interstitial
fibrosis, as previously described (13).
Immunohistochemistry
The kidney sections (4-µm-thick) were
deparaffinised with xylene and rehydrated in graded ethanol.
Endogenous horseradish peroxidase (HRP) activity was blocked with
3% H2O2 in methanol for 30 min. Antigen
retrieval was performed by microwave treatment in 10 mM citrate
buffer for 15 min. The sections were incubated with Sirt1
(sc-74465), α-smooth muscle actin (α-SMA; sc-53015), tissue
inhibitor of metalloproteinase (TIMP)-1 (sc-21734) and plasminogen
activator inhibitor-1 (PAI-1; sc-8979) (Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA) antibodies overnight at 4°C. The
sections were then washed and incubated with biotinylated secondary
antibody (sc-2364 and sc-2363; Santa Cruz Biotechnology, Inc.) and
HRP-conjugated streptavidin. Labelling was visualised with
3,3-diaminobenzidine (DAB; Sigma) to produce a brown colour, and
the sections were counterstained with hematoxylin (Abcam,
Cambridge, UK).
Detection of apoptotic cells
Apoptotic cells in the kidneys were identified using
the Dead End™ Colorimetric terminal
deoxynucleotidyltransferase-mediated dUTP nick-end labelling
(TUNEL) system (Promega, Madison, WI, USA). Briefly,
4-µm-thick paraffin-embedded sections were treated with
freshly diluted 20 µg/ml proteinase K for 15 min at room
temperature. Following treatment with equilibration buffer for 10
min, the rTdT reaction mix was added for 60 min at 37°C. Endogenous
peroxidase activity was blocked by incubating the sections in 0.3%
H2O2 in phosphate-buffered saline (PBS) for 5
min. Streptavidin HRP solution was applied to each slide followed
by incubation for 30 min at room temperature. The sections were
developed with DAB. In randomly selected entire cortex fields, we
captured 6 non-repeating micrographs (at ×400 magnification) for
each sample using a Leica DM2000 LED light microscope (Leica,
Heidelberg, Germany). The number of TUNEL-positive apoptotic cells
was counted per 100 tubular epithelial cells.
Western blot analysis
The whole kidneys were homogenised in ice-cold RIPA
lysis buffer and centrifuged at 14,000 × g for 25 min at 4°C. The
supernatants were collected. The protein concentration was measured
by Bradford's method, and the supernatant was stored at −80°C. The
cell lysates (50 µg of protein/lane) were loaded, separated
by sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE), and transferred onto polyvinylidenedifluoride (PVDF)
membranes (Millipore, Billerica, MA, USA). The membranes were
incubated overnight at 4°C with the primary antibodies.
Subsequently, the membranes were incubated with goat anti-rabbit
IgG (sc-2004; Santa Cruz Biotechnology, Inc.) or goat anti-mouse
IgG HRP conjugate (sc-2005; Santa Cruz Biotechnology, Inc.), and
then immersed in ECL Plus Western Blotting detection reagent and
exposed to Hyperfilm ECL (both from Amersham, Piscataway, NJ, USA).
The intensity of the bands was measured using Lab Works 4.5
software (UVP, Upland, CA, USA). The primary antibodies were CTGF
(ab6992; Abcam), TGF-β1 (sc-130348), Sirt1 (sc-74465), collagen
type I (Col1; sc-293182), α-SMA (sc-53015), TIMP-1 (sc-21734),
PAI-1 (sc-8979), E-Cadherin (E-cad; sc-71007), Bcl-2 (sc-56015),
Bax (sc-20067) and β-actin (sc-130300) (Santa Cruz Biotechnology,
Inc.).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated and cDNA was prepared from
whole kidney samples using the SV Total RNA Isolation system and
RT-PCR kits (both from Promega, Madison, WI, USA). Quantitative PCR
(qPCR) was performed using SYBR Premix Ex Taq™ II. Real-time PCR
reactions were performed on the Agilent Mx3000P QPCR system
(Agilent, Santa Clara, CA, USA). The mRNA levels were normalised to
the eukaryotic 18s rRNA expression level and were calculated using
the 2(−ΔΔCq) method (14). The
cycling conditions were as follows: initial denaturation at 95°C
for 30 sec, followed by 40 cycles at 95°C for 5 sec, 60°C for 30
sec and 72°C for 30 sec. The primer sequences for Col1, α-SMA,
TIMP-1, PAI-1, E-cad, Bcl-2, Bax, CTGF, TGF-β1, Sirt1 and 18S rRNA
are shown in Table I.
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Primer | Sequence |
|---|
| Col1 | F:
5′-CGCCATCAAGGTCTACTGC-3′ |
| R:
5′-GAATCCATCGGTCATGCTCT-3′ |
| α-SMA | F:
5′-GAAGAGCATCCGACACTG-3′ |
| R:
5′-GGCATAGCCCTCATAGATAG-3′ |
| TIMP-1 | F:
5′-TGGGAAATGCCGCAGATA-3′ |
| R:
5′-GCCAGGGAACCAAGAAGC-3′ |
| PAI-1 | F:
5′-AAAAGTGTTTCAGCAGGTGGTC-3′ |
| R:
5′-ACAAAGATGGCATCCGCAGT-3′ |
| E-cad | F:
5′-CGACCCTGCCTCTGAATC-3′ |
| R:
5′-AGCCAGCCTGTCTCCCTCT-3′ |
| Bcl-2 | F:
5′-GCTACCGTCGTGACTTCGC-3′ |
| R:
5′-TCCCAGCCTCCGTTATCC-3′ |
| Bax | F:
5′-CCAGGATGCGTCCACCAA-3′ |
| R:
5′-AAGTAGAAGAGGGCAACCAC-3′ |
| CTGF | F:
5′-GCCCAGACCCAACTATGATGC-3′ |
| R:
5′-GCCCTTCTTAATGTTTTCCTCC-3′ |
| TGF-β1 | F:
5′-GGCGGTGCTCGCTTTGTA-3′ |
| R:
5′-TTTCTCATAGATGGCGTTGTT-3′ |
| Sirt1 | F:
5′-TAGGGAACCTTTGCCTCATCT-3′ |
| R:
5′-CATTGTTGTTTGTTGCTTGGTC-3′ |
| 18sRNA | F:
5′-AAACGGCTACCACATCCAAGG-3′ |
| R:
5′-CCGCTCCCAAGATCCAACTAC-3′ |
Sirt1 deacetylase activity assay
For the measurement of Sirt1 deacetylase activity,
the nuclear proteins were extracted and Sirt1 deacetylase activity
was colourimetrically measured using the Sirt1 Deacetylase Activity
assay kit (Genmed Scientifics Inc., Arlington, MA, USA) according
to the manufacturer's instructions). The kit provides a Sirt1
substrate with an acetylated peptide fragment derived from p53,
which is known to be deacetylated by Sirt1, that was pre-labeled by
p-Nitroaniline. Following deacetylation by Sirt1, the
amino-peptidase cleaved the deacetylated substrate and generated a
highly chemiluminescent molecule group of p-Nitroaniline. The
optical densities (OD) at a wavelength of 405 nm were recorded
using a spectrophotometer (Diareader ELX800G; Dialab GmbH, Vienna,
Austria).
Measurement of SOD, MDA, GSH and GPx
levels
The superoxidedismutase (SOD; A001-1), malonaldehyde
(MDA; A003-1), glutathione (GSH; A006-2) and glutathione peroxidase
(GPx; A005) kits were purchased from Jiancheng Bioengineering
Institute (Nanjing, China). First, the prepared fresh kidney tissue
samples were homogenised in a saline solution to produce 10% kidney
tissue homogenates. The homogenates were centrifuged at 3,500 rpm
for 20 min. The supernatant was collected to measure the levels of
SOD (hydroxylamine method), MDA (thiobarbituric acid method), GSH
(microplate assay) and GPx (colorimetric method) using the specific
kits according to the manufacturer's instructions.
Statistical analysis
The data are presented as the means ± SD.
Statistical analysis was performed by one-way analysis of variance
(ANOVA) using SPSS 11.0 statistical software. Significance was
defined at a value of P<0.05.
Results
Sirt1 activation attenuates UUO-induced
fibrosis
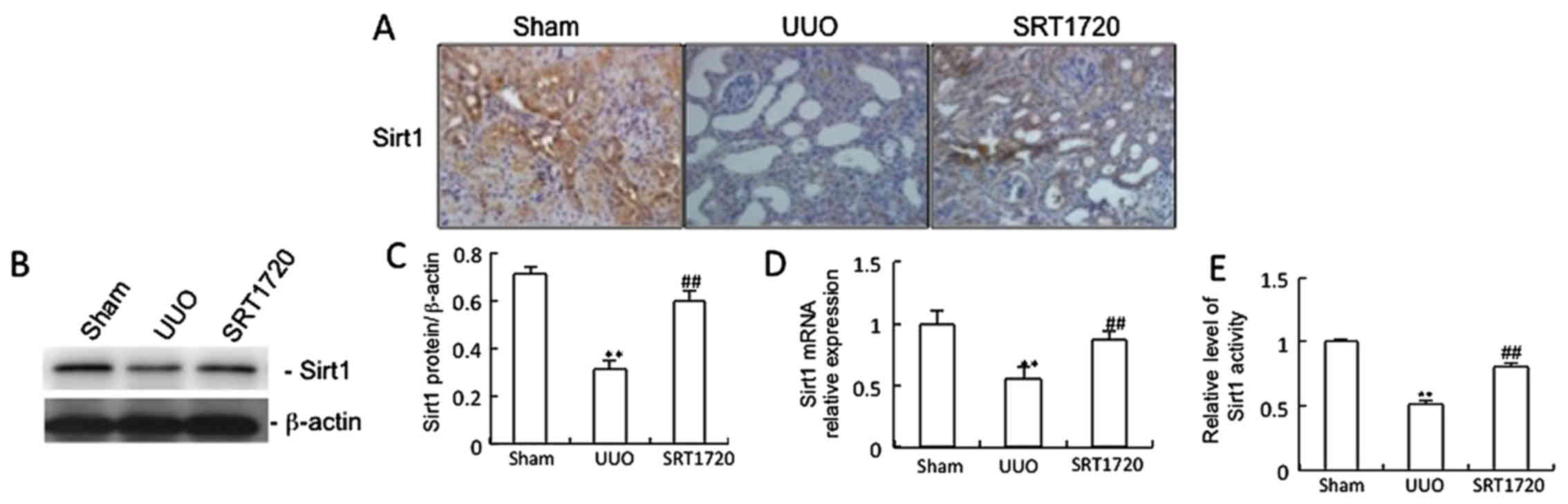
The Sirt1 protein (Fig. 1A–C), mRNA (Fig. 1D) and activity levels (Fig. 1E) were markedly decreased after
UUO compared with the sham group. Treatment with the Sirt1
activator, SRT1720, increased the Sirt1 mRNA, protein and activity
levels compared with the UUO group (Fig. 1). The percentage area of
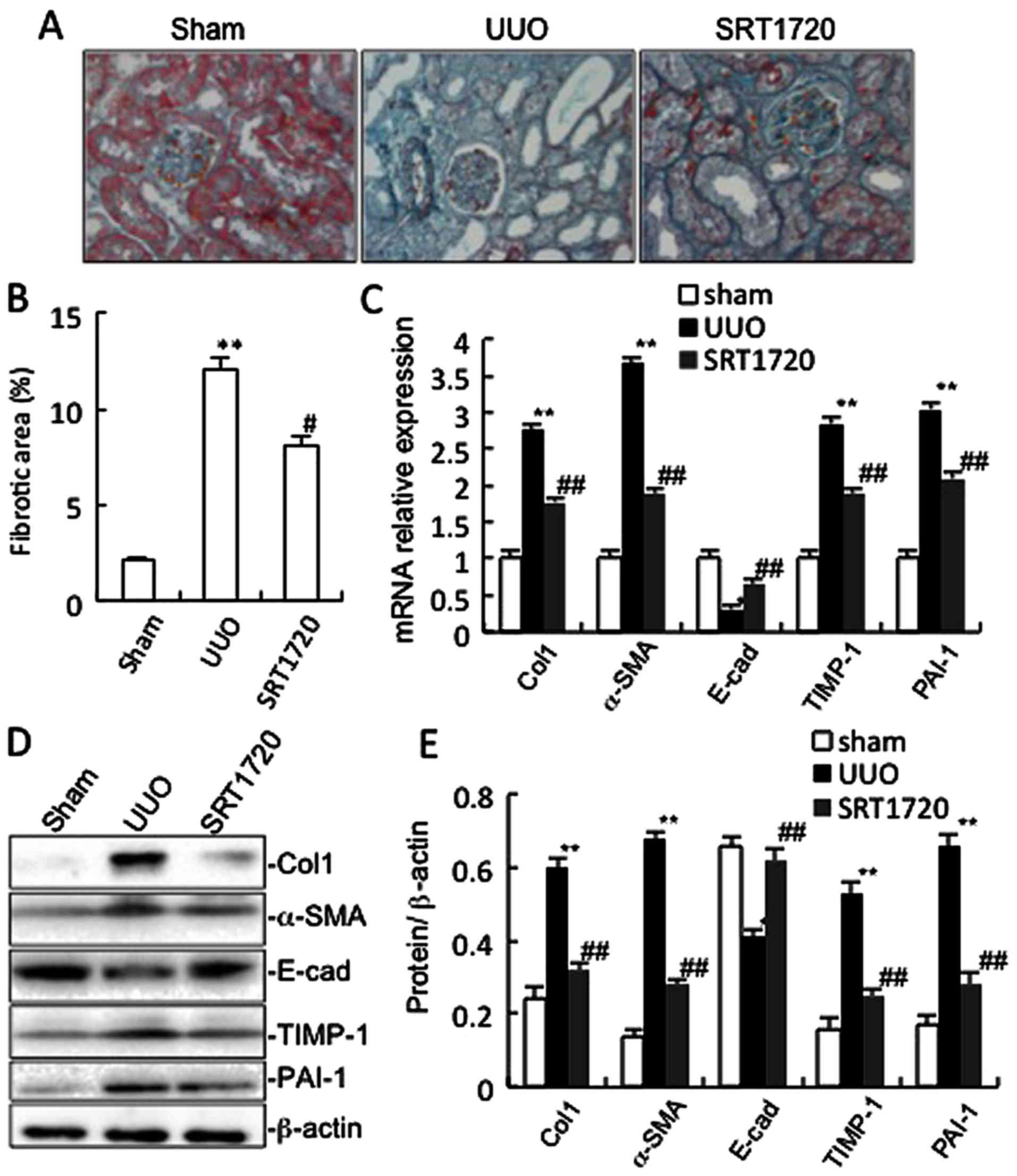
interstitial fibrosis in the kidneys from the sham-operated mice
exhibited very little (if any) fibrosis in the tubules or
interstitium. The percentage fibrotic area in the kidneys from the
mice subjected to UUO was much higher than that of the sham group
(Fig. 2A and B). Masson's
trichrome staining revealed increased collagen deposition in the
kidneys from the mice subjected to UUO compared with the
sham-operated mice (Fig. 2A). We
also analysed the total kidney tissue lysates by western blot
analysis for Col1. An increase in Col1 protein expression was
observed in the obstructed kidneys compared with the sham group
(Fig. 2D and E). Consistent with
the protein expression of Col1, the mRNA level of Col1 was
increased in the mice subjected to UUO (Fig. 2C). SRT1720 substantially reversed
these changes (Fig. 2).
A variety of cytokines are known to affect the
process of interstitial fibrosis, such as PAI-1 and TIMP-1
(15). Tubular EMT occurs when
losing epithelial markers, such as E-cad, and acquiring mesenchymal
ones, such as α-SMA (16). In
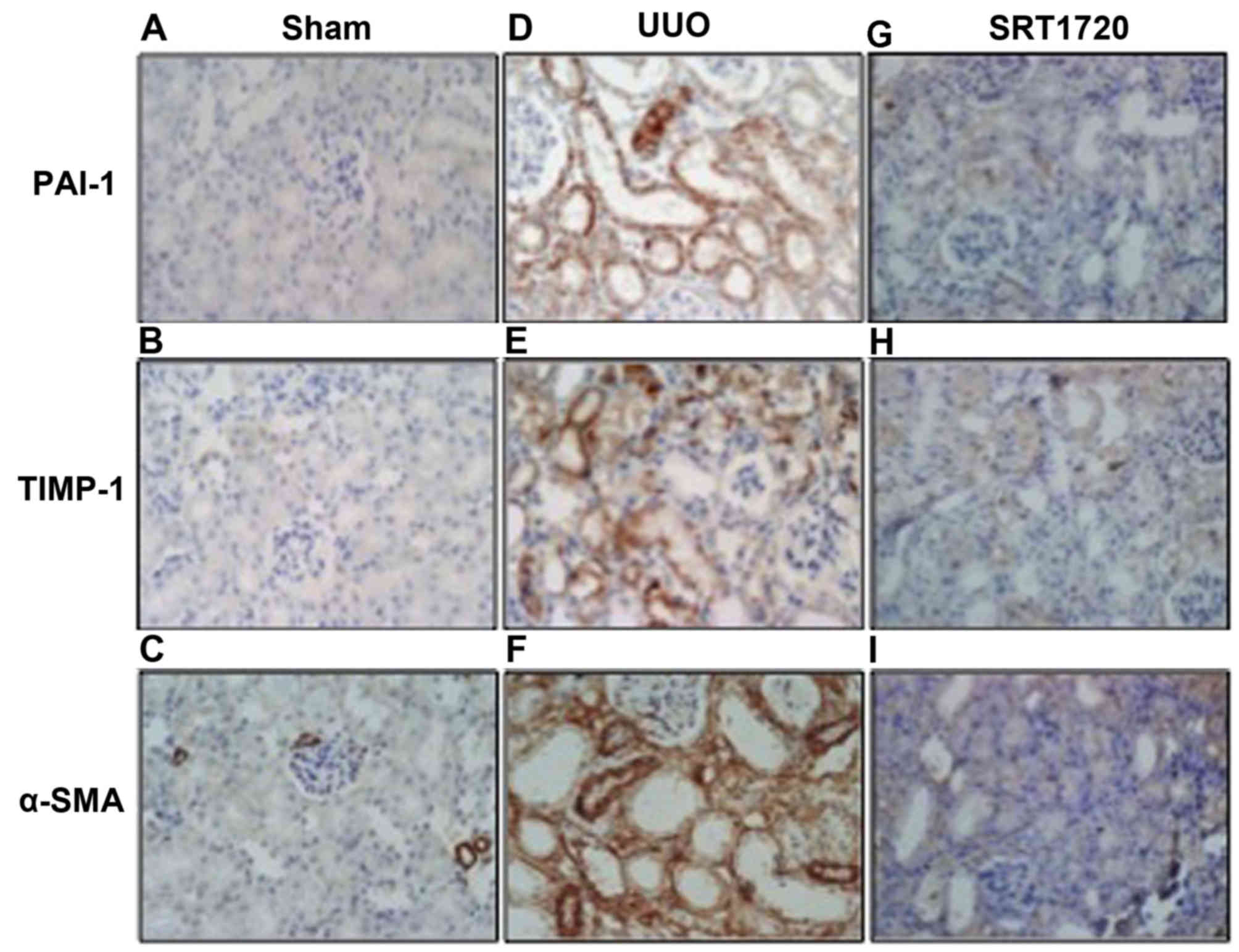
this study, immunohistochemistry (Fig. 3) indicated that positive staining
for PAI-1, TIMP-1 and α-SMA was mainly found in the tubular
epithelial cells in the kidneys from the mice subjected to UUO.
Moreover, our results demonstrated that the mRNA and protein levels
of PAI-1, TIMP-1 and α-SMA all increased significantly in the
obstructed kidneys of the mice in the UUO group compared to those
of the sham group, as shown by western blot analysis and RT-qPCR
(Fig. 2C–E). UUO led to decreased
E-cad mRNA and protein expression. SRT1720 notably decreased the
expression of α-SMA, PAI-1 and TIMP-1, and increased E-cad
expression in the mice subjected to UUO (Fig. 2C–E).
Sirt1 activation attenuates UUO-induced
apoptosis in the kidneys of mice with UUO-induced fibrosis
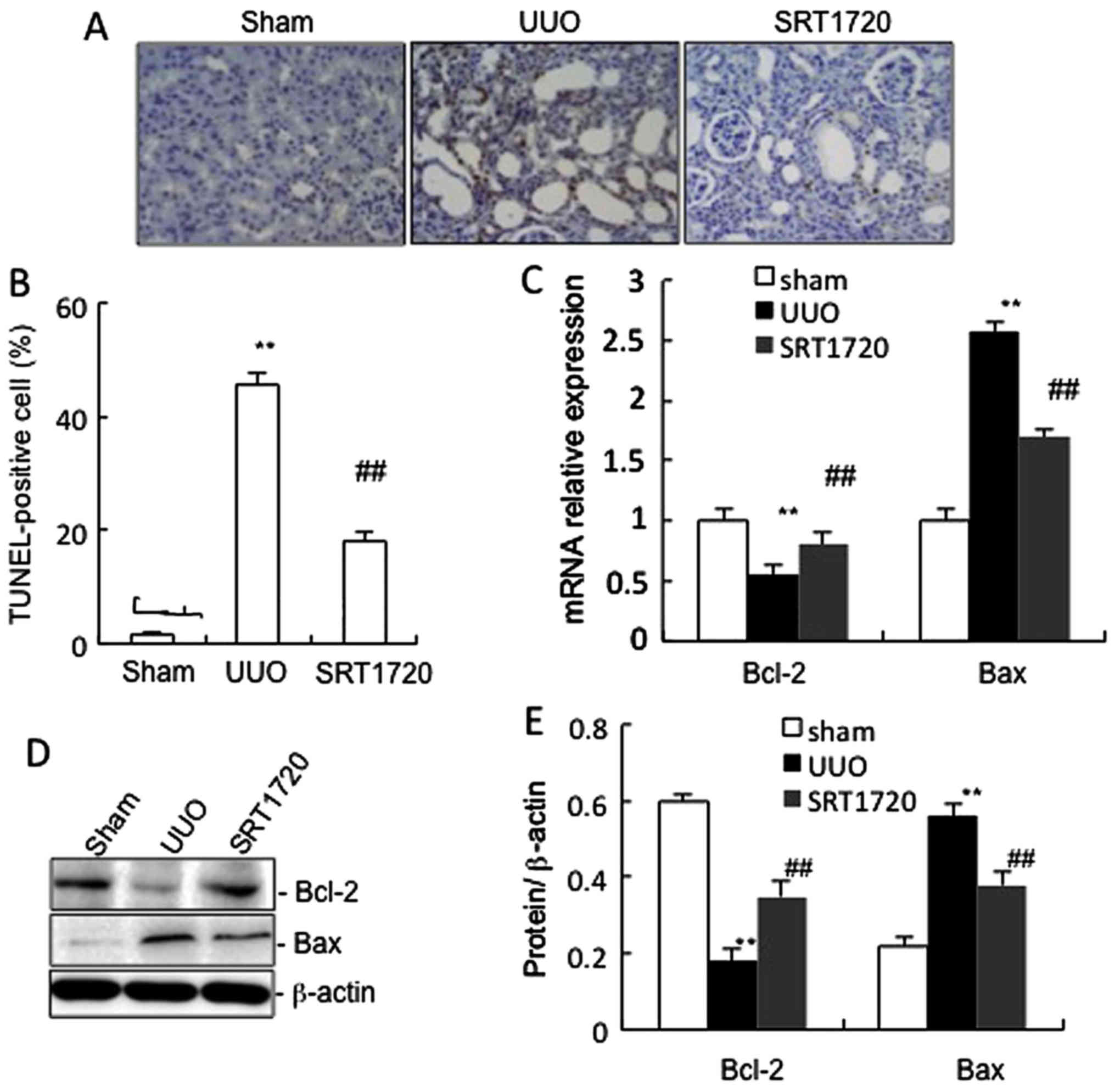
Fig. 4A and B
present the number of TUNEL-positive tubular epithelial cells in
the mice subjected to UUO and treated with or without SRT1720. UUO
significantly increased tubular epithelial cell apoptosis, which
was markedly decreased by the delivery of SRT1720, compared with
the sham-operated mice. Furthermore, in the obstructed kidneys from
the mice in the UUO group, 7 days of UUO resulted in a significant
decrease in the protein and mRNA expression of Bcl-2, and in an
increase in Bax expression compared with the sham-operated group.
SRT1720 administration attenuated the changes in Bcl-2 and Bax
expression (Fig. 4C–E).
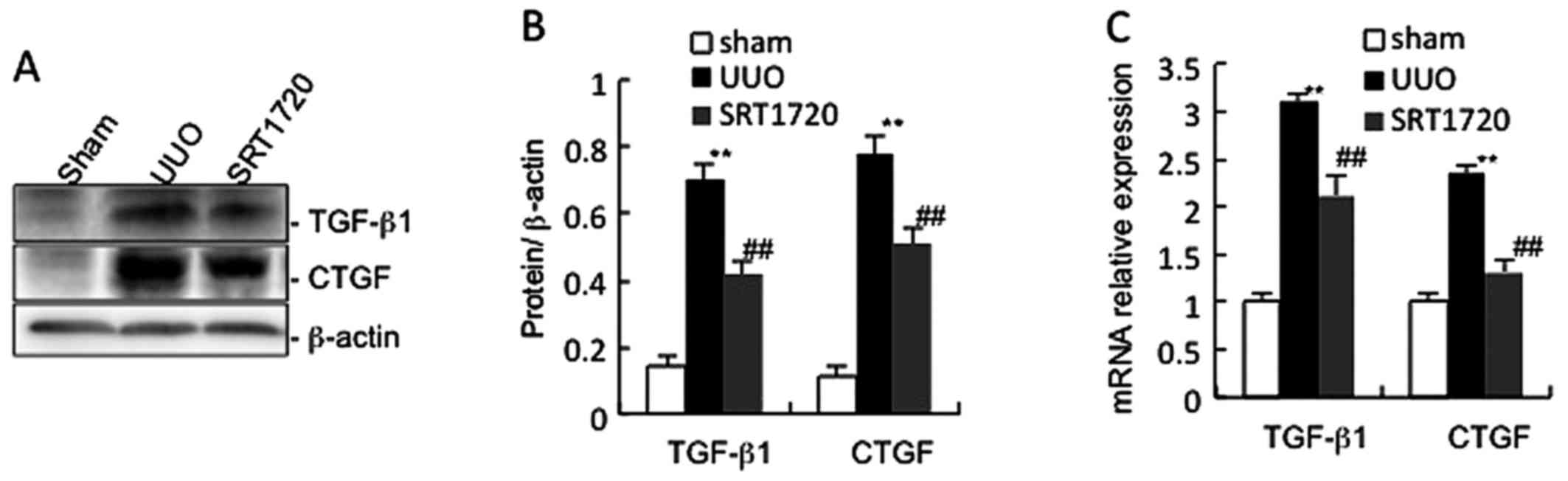
Sirt1 activation inhibits the TGF-β1/CTGF
signalling pathway
It is well known that TGF-β1 plays an important role
in the physiology and pathology of kidney fibrosis (2). In this study, the TGF-β1 level in
the mice subjected to UUO was increased compared with the sham
group. SRT1720 attenuated the UUO-induced increase in TGF-β1
expression (Fig. 5). We then
evaluated the TGF-β1 downstream signalling molecule, CTGF. SRT1720
treatment also significantly blunted the increase in CTGF
expression induced by UUO in mice.
Sirt1 activation inhibits oxidative
stress in kidneys
Oxidative stress has been postulated as an important
contributing factor to fibrosis. Antioxidant enzymes, including SOD
and GPx, protect against oxidative stress by converting free
radicals or reactive oxygen intermediates to non-radical products.
GSH is the major intracellular thiol that plays a critical role in
the cellular defense against oxidative stress. It is clearly
evident from Table II that UUO
led to decreased SOD and GPx activity in the kidneys compared to
the sham group. No significant change in the GSH level occurred
after UUO. However, SRT1720 enhanced the renal SOD, GPx and GSH
levels compared to the sham group. The increased activity of SOD,
GPx and GSH in this study reflects a compensatory mechanism to the
increased oxidative stress. With respect to the pro-oxidative
biomarker, MDA (a major product of lipid peroxidation), the renal
levels were significantly higher in the UUO group compared to the
sham group, and the MDA levels decreased in the SRT1720 group
compared with the UUO group (Table
II).
| Table IIThe results of oxidative stress
analysis in sham and experimental groupsa. |
Table II
The results of oxidative stress
analysis in sham and experimental groupsa.
| Parameters | Unit | sham | UUO | SRT1720 |
|---|
| MDA | nmol/mg protein | 7.12±1.06 | 12.01±1.94b | 8.01±1.85c |
| SOD | U/mg protein | 15.78±1.25 | 10.34±0.96b | 14.36±1.44c |
| GSH | µmol/mg
protein | 5.44±1.01 | 4.89±1.20d | 8.62±0.87c |
| GPx | mU/mg protein | 378.34±28.25 |
257.75±40.66b |
409.30±41.74c |
Discussion
Renal fibrosis, whether the origin is inflammatory
or immunological, obstructive, metabolic or systemic, inevitably
progresses to end-stage renal disease with a progressive,
irreversible decline in renal function. Therefore, the regulation
of the renal fibrotic process provides new insight into the
treatment and prevention of chronic kidney disease. The present
study demonstrated that Sirt1 deficiency accentuated renal tubular
cell apoptosis and tubulointerstitial fibrosis. Conversely, the
Sirt1 activator, SRT1720, promoted the resistance of renal cells to
oxidative stress during UUO. One of the primary mechanisms by which
SRT1720 mediates its protective effects in the kidneys was by
inhibiting TGF-β1/CTGF expression.
Growing evidence indicates that sirtuins play a
regulatory role in the process of different renal diseases. The
antifibrotic function of Sirt1 has been reported in heart failure,
cardiac fibrosis, hepatic steatosis, diabetic nephropathy and UUO
(17). Furthermore, it has been
demonstrated that Sirt1 is associated with TGF-β in endothelial,
vascular smooth muscle and bronchial epithelial cells. The
overexpression of Sirt1 attenuates TGF-β1-induced extracellular
matrix expression in cultured mesangial cells (8). In this study, we observed that in
the UUO-obstructed mouse kidneys, Sirt1 expression and activity
both markedly decreased. The the Sirt1 activator, SRT1720, was
administered. Our results demonstrated that SRT1720 increased the
Sirt1 levels and activity, and attenuated the renal upregulation of
matrix accumulation and epithelial-mesenchymal transition. We also
demonstrated that SRT1720 also effectively attenuated apoptosis
induced by UUO. The resutls of TUNEL assay also indicated that UUO
increased the apoptotic rate, which was effectively inhibited by
treatment with SRT1720. The results of western blot analysis and
RT-qPCR revealed that Sirt1 overexpression increased Bcl-2 protein
and mRNA expression, and decreased the protein and mRNA levels of
Bax. The protective effects of Sirt1 on apoptosis have been
reported in several studies in multiple organs and tissues, against
injury from diverse causes. H2O2-induced
apoptosis has been shown to be significantly attenuated in
Sirt1-overexpressing murine mesangial cells, but to be enhanced
with Sirt1-knockdown in MMCs (18). The downregulation of Sirt1
expression in diabetes promotes podocyte apoptosis (19). In another study, the UUO model of
kidney injury markedly induced more renal apoptosis and fibrosis in
Sirt1+/− mice compared to the wild-type controls, and
the pharmacological Sirt1 activation substantially attenuated
apoptosis and fibrosis in wild-type mice (17). These data further indicate that
Sirt1 may be a promising therapeutic target as a common pathway in
renal apoptosis and fibrosis.
The pathogenesis of renal fibrosis has been depicted
as a continuum of 4 overlapping phases: priming (localised
inflammatory response), activation (activation and recruitment of
matrix producing cells), execution (accumulation of matrix protein)
and progression (loss of cells and renal function) (20). TGF-β1 and CTGF, key mediators of
kidney fibrosis, were identified based on their contribution to
these phases (2). TGF-β1, an
ubiquitously expressed cytokine, also promotes tissue repair, but
its excessive action can lead to diverse profibrotic events in the
liver, lung, kidney and breast (21). CTGF has many diverse functions,
such as angiogenesis, fibrosis and apoptosis (22). In this study, we found that the
expression of CTGF and TGF-β1 increased in the UUO-obstructed
kidneys, which was accompanied by a significant upregulation of
matrix accumulation, EMT and apoptosis. This result is consistent
with the observation that the expression of TGF-β1 and CTGF is
increased in human and experimental models of chronic kidney
disease (correlating with the severity of renal fibrosis) (23–25).
Oxidative stress results from an imbalance between
ROS production and elimination. Modi et al demonstrated that
the MDA levels were significantly higher in rats with ureteral
obstruction compared with the sham-treated group (26). In response to ROS, antioxidant
enzymes, including SOD, are thought to protect cellular function. A
deficiency in these antioxidant enzymes enhances ROS production and
renal tubulointerstitial fibrosis (27). Our results demonstrated that under
normal conditions, kidney cells contain low levels of the
pro-oxidative biomarker, MDA. Abundant antioxidant enzymes (such as
SOD and GPx) are likely to play an important role in protecting the
kidneys against oxidative stress. However, UUO led to decreased SOD
and GPx activity compared to the normal group. There was no
significant difference in the GSH levels. The MDA levels were
markedly higher than the normal group.
In the present study, to the best of our knowledge,
we demonstrated for the first time that SRT1720 treatment decreased
TGF-β1 production and CTGF activation in kidney tissues obtained
from mice subjected to UUO. In addition, SRT1720 treatment reversed
the oxidative stress induced by UUO.
Taken together, these data suggest that the Sirt1
activator, SRT1720, exerts protective effects against UUO-induced
tubulointerstitial fibrosis, in part through the suppression of
renal oxidative stress and the TGF-β1/CTGF signalling pathway.
Targeting Sirt1 with pharmacological activators may thus improve
renal function during conditions that induce tubulointerstitial
fibrosis, including ureteral obstruction.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (no. 81200532) and the
preferred fund of Hebei Province Postdoctoral Scientific Research
Projects (no. B2015003028).
References
|
1
|
Klahr S and Morrissey J: Obstructive
nephropathy and renal fibrosis. Am J Physiol Renal Physiol.
283:F861–F875. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Leask A and Abraham DJ: TGF-beta signaling
and the fibrotic response. FASEB J. 18:816–827. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Devarajan P: Update on mechanisms of
ischemic acute kidney injury. J Am Soc Nephrol. 17:1503–1520. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chiu C, Maddock DA, Zhang Q, Souza KP,
Townsend AR and Wan Y: TGF-beta-induced p38 activation is mediated
by Rac1-regulated generation of reactive oxygen species in cultured
human keratinocytes. Int J Mol Med. 8:251–255. 2001.PubMed/NCBI
|
|
5
|
Herrera B, Alvarez AM, Sánchez A,
Fernández M, Roncero C, Benito M and Fabregat I: Reactive oxygen
species (ROS) mediates the mitochondrial-dependent apoptosis
induced by transforming growth factor (β) in fetal hepatocytes.
FASEB J. 15:741–751. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Michan S and Sinclair D: Sirtuins in
mammals: Insights into their biological function. Biochem J.
404:1–13. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim DH, Jung YJ, Lee JE, Lee AS, Kang KP,
Lee S, Park SK, Han MK, Lee SY, Ramkumar KM, et al: SIRT1
activation by resveratrol ameliorates cisplatin-induced renal
injury through deacetylation of p53. Am J Physiol Renal Physiol.
301:F427–F435. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang XZ, Wen D, Zhang M, Xie Q, Ma L,
Guan Y, Ren Y, Chen J and Hao CM: Sirt1 activation ameliorates
renal fibrosis by inhibiting the TGF-β/Smad3 pathway. J Cell
Biochem. 115:996–1005. 2014. View Article : Google Scholar
|
|
9
|
Kume S, Haneda M, Kanasaki K, Sugimoto T,
Araki S, Isshiki K, Isono M, Uzu T, Guarente L, Kashiwagi A, et al:
SIRT1 inhibits transforming growth factor beta-induced apoptosis in
glomerular mesangial cells via Smad7 deacetylation. J Biol Chem.
282:151–158. 2007. View Article : Google Scholar
|
|
10
|
Mao B, Hu F, Cheng J, Wang P, Xu M, Yuan
F, Meng S, Wang Y, Yuan Z and Bi W: SIRT1 regulates YAP2-mediated
cell proliferation and chemoresistance in hepatocellular carcinoma.
Oncogene. 33:1468–1474. 2014. View Article : Google Scholar
|
|
11
|
Milne JC, Lambert PD, Schenk S, Carney DP,
Smith JJ, Gagne DJ, Jin L, Boss O, Perni RB, Vu CB, et al: Small
molecule activators of SIRT1 as therapeutics for the treatment of
type 2 diabetes. Nature. 450:712–716. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gano LB, Donato AJ, Pasha HM, Hearon CM
Jr, Sindler AL and Seals DR: The SIRT1 activator SRT1720 reverses
vascular endothelial dysfunction, excessive superoxide production,
and inflammation with aging in mice. Am J Physiol Heart Circ
Physiol. 307:H1754–H1763. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shimizu H, Maruyama S, Yuzawa Y, Kato T,
Miki Y, Suzuki S, Sato W, Morita Y, Maruyama H, Egashira K, et al:
Antimonocyte chemoattractant protein-1 gene therapy attenuated
renal injury induced by protein-overload proteinuria. J Am Soc
Nephrol. 14:1496–1505. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−ΔΔC(T)) method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
15
|
Duymelinck C, Deng JT, Dauwe SE, De Broe
ME and Verpooten GA: Inhibition of the matrix metalloproteinase
system in a rat model of chronic cyclosporine nephropathy. Kidney
Int. 54:804–818. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang J and Liu Y: Dissection of key events
in tubular epithelial to myofibroblast transition and its
implications in renal interstitial fibrosis. Am J Pathol.
159:1465–1475. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
He W, Wang Y, Zhang MZ, You L, Davis LS,
Fan H, Yang HC, Fogo AB, Zent R, Harris RC, et al: Sirt1 activation
protects the mouse renal medulla from oxidative injury. J Clin
Invest. 120:1056–1068. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kume S, Haneda M, Kanasaki K, Sugimoto T,
Araki S, Isono M, Isshiki K, Uzu T, Kashiwagi A and Koya D: Silent
information regulator 2 (SIRT1) attenuates oxidative stress-induced
mesangial cell apoptosis via p53 deacetylation. Free Radic Biol
Med. 40:2175–2182. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chuang PY, Dai Y, Liu R, He H, Kretzler M,
Jim B, Cohen CD and He JC: Alteration of forkhead box O (foxo4)
acetylation mediates apoptosis of podocytes in diabetes mellitus.
PLoS One. 6:e235662011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Y: Cellular and molecular mechanisms
of renal fibrosis. Nat Rev Nephrol. 7:684–696. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Franklin TJ: Therapeutic approaches to
organ fibrosis. Int J Biochem Cell Biol. 29:79–89. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sánchez-López E, Rodrigues Díez R,
Rodríguez Vita J, Rayego Mateos S, Rodrigues Díez RR, Rodríguez
García E, Lavoz Barria C, Mezzano S, Egido J, Ortiz A, Ruiz-Ortega
M and Selgas R: Connective tissue growth factor (CTGF): a key
factor in the onset and progression of kidney damage. Nefrologia.
29:382–391. 2009.In Spanish.
|
|
23
|
Border WA and Noble NA: Transforming
growth factor beta in tissue fibrosis. N Engl J Med. 331:1286–1292.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ito Y, Aten J, Bende RJ, Oemar BS,
Rabelink TJ, Weening JJ and Goldschmeding R: Expression of
connective tissue growth factor in human renal fibrosis. Kidney
Int. 53:853–861. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yokoi H, Sugawara A, Mukoyama M, Mori K,
Makino H, Suganami T, Nagae T, Yahata K, Fujinaga Y, Tanaka I, et
al: Role of connective tissue growth factor in profibrotic action
of transforming growth factor-beta: A potential target for
preventing renal fibrosis. Am J Kidney Dis. 38(Suppl 1): S134–S138.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Modi KS, Morrissey J, Shah SV, Schreiner
GF and Klahr S: Effects of probucol on renal function in rats with
bilateral ureteral obstruction. Kidney Int. 38:843–850. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sunami R, Sugiyama H, Wang DH, Kobayashi
M, Maeshima Y, Yamasaki Y, Masuoka N, Ogawa N, Kira S and Makino H:
Acatalasemia sensitizes renal tubular epithelial cells to apoptosis
and exacerbates renal fibrosis after unilateral ureteral
obstruction. Am J Physiol Renal Physiol. 286:F1030–F1038. 2004.
View Article : Google Scholar : PubMed/NCBI
|