Introduction
Peripheral nerve damage is common worldwide, and
patients suffering from this type of injury may partially or
completely lose motor, sensory and autonomic function (1). Nevertheless, the treatment of
peripheral nerve injury (PNI) remains a majory medical concern due
to the lack of satisfactory treatments (2,3).
Following PNI in mammals, the nerve distal to the
site of injury may undergo a process known as Wallerian
degeneration, during which Schwann cells lose contact with axons
and dedifferentiate into stem-like cells or repair cells that play
a vital role in repairing PNI (4,5) by
forming a bundle that provides a permissive microenvironment for
axon regeneration. Along with Schwann cells, macrophages gather at
the site of PNI to clear myelin debris. It has also been widely
reported that Schwann cells may secrete trophic support molecules,
such as neurotrophin-3 (NT-3), fibroblast growth factor (FGF) and
nerve growth factor (NGF) (6).
Once Schwann cells reach the regrowing axons, they begin to
remyelinate the axon apprxomately 7 days following nerve injury
(7,8). The molecular mechanism of the
regenerative process has not yet been fully elucidated. A better
understanding of the mechanisms of Wallerian degeneration would
help us to improve the repair process after PNI.
Long non-coding RNAs (lncRNAs) are
non-protein-coding RNA molecules ranging from 200 nt to
approximately 100 kb in length (9,10).
lncRNAs have been proven to regulate gene expression at almost
every level of transcription and translation, including genomic
imprinting, chromatin modification and cytoplasmic mRNA translation
(11,12). Data from an increasing number of
studies have indicated that lncRNAs are associated with important
regulatory functions during many biological processes. In
neurobiology, lncRNAs are well known to be related to
neurodevelopmental disorders, neurodegeneration and brain cancers
(13). For example, the knockdown
of linc-Brn1b has been shown to result in the reduction of
intermediate progenitor cells in the brain, indicating that this
lncRNA may play an important role in cortical development (14). In another previous study, lncRNAs
that were differentially expressed following sciatic nerve
resection in rats were found to be involved in regenerating dorsal
root ganglion (DRG) neurons, and the downregulation of lncRNA
BC089918 was found to promote neurite regeneration in DRG neurons
(15). Although lncRNA studies
are now common in various fields, the lncRNA expression signature
and the possible roles of lncRNAs following Wallerian degeneration
in the distal end of the peripheral nerve have not yet been
reported, at least to the best of our knowledge.
In this study, the expression of lncRNAs and mRNAs
in the distal segment of the sciatic nerve following PNI was
profiled using microarray analysis. We identified a group of
lncRNAs that were significantly dysregulated at different time
points during Wallerian degeneration. The classification of the
lncRNAs, validation by RT-qPCR, target prediction, gene ontology
(GO) analysis and target prediction were performed. In particular,
we predicted several pairs of lncRNAs and their related mRNAs.
RT-qPCR analysis was used for the preliminary verification of the
lncRNA-mRNA pairs. The overexpression of NONMMUG014387 was also
found to promote the proliferation of mouse Schwann cells (MSCs).
This study may provide a basis for the further investigation of the
function of lncRNAs in peripheral nerve regeneration following
injury.
Materials and methods
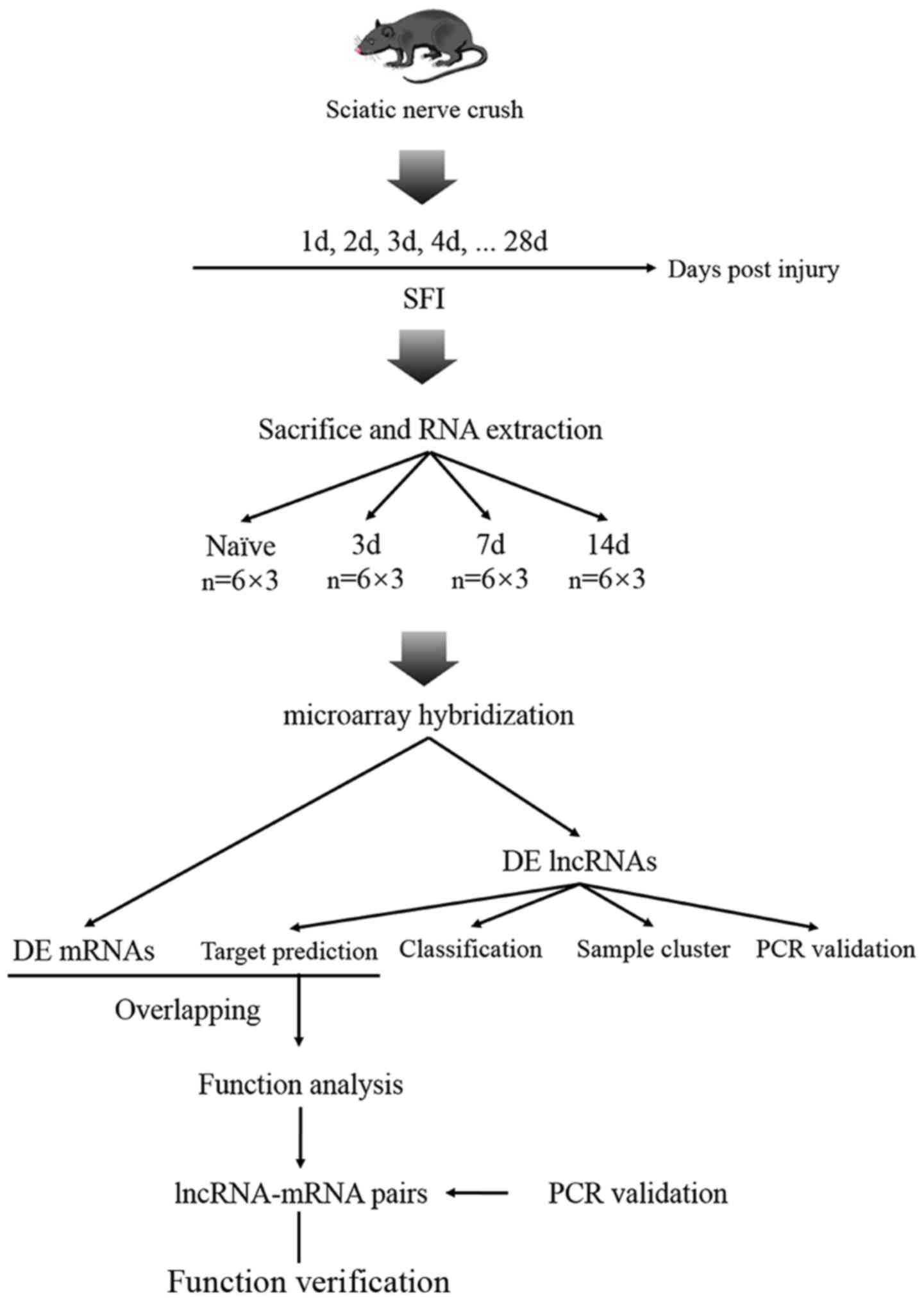
The complete research process used in the present
study is summarized and presented in Fig. 1.
Experimental animals
A total of 99 mice were used in this study; 15 mice
were used for the functional assessment of sciatic functional index
(SFI), 72 mice were used for microarray analysis and 12 mice were
used for further PCR verification. All the mice were obtained from
the Laboratory Animal Research Center, Academy of Military Medical
Sciences, of the Chinese People's Liberation Army, Beijing,
China.
Functional assessment of SFI
The right sciatic nerves of 15 C57Bl6 mice were
crushed using an ultra-fine, smooth, straight hemostat (tip width,
0.6 mm, Fine Science Tools) for 20 sec, as previously descrbied
(16). After this procedure, the
SFI was measured in each mouse daily for 28 days. The hind paws of
the mice were immersed in non-toxic ink. The mice moved without
assistance along a walking track and generated footprints were
recorded. As previously described (17), several parameters were calculated
for the SFI: print length (PL), toe spread (TS) and intermediate
toe spread (ITS). All parameters were measured for normal (N) and
experimental (E) animals. The SFI was determined according to the
formula described by Inserra et al (17) as follows: SFI, 118.8 (ETS −
NTS/NTS) − 51.2 (EPL − NPL/NPL) − 7.5, where NTS is normal toe
spread, ETS is experimental toe spread, EPL is experimental print
length, and NPL is normal print length.
Animal preparation and nerve lesion
experiments
A total of 72 C57Bl6 mice, approximately 2 months of
age were selected and randomly classified into 4 groups according
to the time points of 0, 3, 7 and 14 days post-surgery. The 0 day
group was used as a control, while all the other groups were the
experimental groups. Due to the insufficient volume of a single
sciatic nerve, 6 mice were pooled into one sample, and 3 samples
for each time point. After the mice were anaesthetized via an
intraperitoneal injection of chloral hydrate, an incision was
created on the right lateral thigh to expose and lift the sciatic
nerve. As described in a previous study (18), right sciatic nerve crush was
performed at the upper thigh level using an acutenaculum. The nerve
was compressed for 30 sec to ensure that the axon was disconnected
while the epineurium remained intact. The crush site was then
ligated to generate a marker for the damage site. The incision was
then closed. To relieve painful mechanical stimulation and
discomfort, the mice were housed in clean cages with sawdust
bedding. We provided free access to food and water. These
experiments were performed according to the NIH Guidelines for the
Care and Use of Laboratory Animals (http://oacu.od.nih.gov/regs/index.htm). All procedures
were approved by the Ethics Committee of Tianjin Medical
University, Tianjin, China.
Microarray experiment
The microarray experiment was conducted using an
Agilent-074622 Mouse lncRNA micro-array (Agilent Technologies,
Santa Clara, CA, USA). The mice were sacrificed by cervical
dislocation. Tissue perfusion was performed to remove the blood in
the sciatic nerve. Distal segments of crushed sciatic nerves (0.5
cm) were isolated and harvested from the mice of each group 0, 3, 7
and 14 days post-surgery. Total RNA was extracted using a Takara
RNAiso Plus kit (#9109) according to the manufacturer's
instructions. The quality of the total RNA was assessed by the RNA
integrity number (RIN) on an Agilent Bioanalyzer 2100 (Agilent
Technologies). The total RNA was purified using an RNase-Free DNase
kit and an RNeasy micro kit (both Qiagen GmBH, Hilden, Germany).
The total RNA samples had a RIN of ≥7.0 and a 28S/18S ratio of
≥0.7.
The preparation of One-Color Spike Mix was performed
using an Agilent One-Color RNA Spike-In kit. We amplified and
labeled total RNA using a Low Input Quick Amp WT Labeling kit
(Agilent Technologies) and purified cDNA using an RNeasy mini kit
(Qiagen GmBH).
A total of 1.65 µg of Cy3-labeled cDNA was
hybridized onto each microarray using a Gene Expression
Hybridization kit in a hybridization oven (both from Agilent
Technologies). Microarray slides were washed after a 17-h
hybridization using a Gene Expression Wash Buffer kit (Agilent
Technologies). All aforementioned procedures were performed in
accordance with the manufacturer's instructions.
After scanning the microarrays with an Agilent
Microarray Scanner we used Feature Extraction software v10.7 (both
from Agilent Technologies) to extract the data. Raw data were
normalized by a quantile algorithm in GeneSpring software v11.0
(Agilent Technologies).
Bioinformatics analysis
The microarray data generated in our study were
deposited into the NCBI Gene Expression Omnibus (GEO) under
accession no. GSE74087 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE74087).
Raw data were extracted using Feature Extraction
10.7 software, and the quantiles were normalized using GeneSpring
GX 11.0 software (Agilent Technologies). The global distribution
characteristics of the sample data were normalized and visualized
using a box plot. To comprehensively and clearly depict the
associations and differences among the samples, clustering into
expressed genes or differentially expressed genes was performed.
Samples with similar characteristics may present in the same
cluster after processing, and genes presenting in the same cluster
may have similar biological functions. Following the normalization
of the raw data, the fold change was determined, and multiple
hypothesis testing was performed to identify differentially
expressed genes. Genes with a fold change (linear) ≤0.5 or ≥2 and a
P-value <0.01 in a t-test were selected. Differentially
expressed genes that were identified were visually presented using
scatter plots and heat maps.
To visualize the time dependence of the differential
lncRNA expression, we performed a time series analysis in which
expression levels were summed to obtain relative expression
patterns over time. lncRNAs were clustered into several distinct
profiles using k-means clustering with a distance matrix based on
Pearson's correlation.
lncRNAs evidently influence the expression of
protein-coding genes; thus, we predicted the target genes of
differentially expressed lncRNAs to further investigate their
functions in biological processes. The possible target genes for
cis- or trans-regulating lncRNAs were predicted by
applying two independent algorithms.
For cis-regulating prediction, the genomic
positional associations between the lncRNAs and their potential
paired target genes were determined from the RefSeq and UCSC Known
Genes databases (19). Based on
the distance between each lncRNA gene and its neighboring known
protein-coding gene, algorithms, including an ORF-Predictor and
BLASTP pipeline were used to identify cis-regulating
lncRNA-mRNA potential pairs. We selected 10 kb as the cut-off of
the distance between lncRNAs and mRNAs based on a previous study
(19).
For trans-regulating prediction, the RNAplex
method (http://www.tbi.univie.ac.at/RNA/RNAplex.1.html) was
applied to identify possible target genes for lncRNAs based on a
previous study (20). RNAplex,
which is specifically designed to rapidly identify possible
hybridization sites for an RNA query in large RNA databases, adopts
a slightly different energy model that reduces the computational
time significantly.
To predict the function of differentially expressed
lncRNAs, we selected predicted target genes that overlapped with
the differentially expressed mRNAs for further GO analysis. GO
analysis was performed to analyze the primary function of the
target coding genes according to the GO database, which defines the
terms 'cellular components', 'molecular functions', and 'biological
process' for each gene. These analyses were used to determine the
functional enrichment of the target mRNA to analyze the functions
of the lncRNAs involved in the response to nerve injury.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
To validate the microarray results, the expression
level of three randomly selected lncRNAs, ENSMUSG00000087366,
NONMMUG018386 and NONMMUG018381, were profiled at different time
points after injury. For the preliminary validation of the
lncRNA-mRNA pairs, the NONMMUG014387-Cthrc1, NONMMUG042364-Ntm and
ENSMUSG00000097535-Icam1 pairs were detected. Glyceraldehyde
3-phosphate dehydrogenase (GAPDH) was used as a control. In total,
12 mice were used for PCR verification. Total RNA was extracted
from the sciatic nerve as described above. Reverse-transcribed
cDNAs were synthesized using a PrimeScript RT Reagent kit according
to the manufacturer's instructions (Shanghai Biotechnology Corp.,
Shanghai, China). PCR reactions were performed using SYBR Premix Ex
Taq according to the manufacturer's instructions in a Rotor-Gene
6000 instrument. The PCR reaction used the following cycling
conditions: 10 min at 95°C; 95°C (10 sec), 60°C (60 sec) and 95°C
(10 sec) for a total of 40 cycles; and a final temperature increase
from 60 to 99°C. The relative quantity of RNA was calculated and
analyzed using the 2ΔΔCq method. All experiments were performed in
triplicate. The primer sequences are presented in Table I.
 | Table IPCR primers used for expression
analyses. |
Table I
PCR primers used for expression
analyses.
| Gene name and
primer sequences (5′→3′) |
|---|
| NONMMUG014387 |
| Forward:
AAAGGGATTACAGGCACACG |
| Reverse:
CCAGGCCATTTACTCAGCAT |
| Cthrc1 |
| Forward:
CGAAATAAAGCCTCTGACGA |
| Reverse:
TTAACTTTGCTTTTTCATTCAGC |
| NONMMUG018381 |
| Forward:
GCTCTTCTAAAGGTCATGGGTTCA |
| Reverse:
CTTGGCTCCCCTGGAACTG |
| NONMMUG018386 |
| Forward:
GGTCACATTTCCACATCAGC |
| Reverse:
GGTCACTCGGGAATCTTGAA |
|
ENSMUSG00000087366 |
| Forward:
ACATTTATGGGACCCCCTCT |
| Reverse:
AACCACCAACACCACTACCAA |
| Ntm |
| Forward:
AGTGCCCCACCATGAAACA |
| Reverse:
TTCTTGCTCTGTGCTTGTGTCTATG |
| NONMMUG042364 |
| Forward:
TCCTGAAGAGAAGCTGCAAA |
| Reverse:
TTCTTCTACCCCAGCTTCCA |
|
ENSMUSG00000097535 |
| Forward:
TAAGACTCGGGGAATGTGGA |
| Reverse:
GGCTTGTCAACACTGAATGC |
| Icam1 |
| Forward:
GTTCTCTGCTCCTCCACATCCT |
| Reverse:
GGCTGACATTGGGAACAAAAGTAG |
| Gapdh |
| Forward:
CGTGTTCCTACCCCCAATGT |
| Reverse:
TGTCATCATACTTGGCAGGTTTCT |
Cell culture
Mouse Schwann cells (MSCs) were puchased from
Shanghai Cellbio Co. (Shanghai, China). The cells were inoculated
in a new culture flask, prepared medium was added, 90% HyClone
Dulbecco's modified Eagle's medium (DMEM) [10% fetal bovine serum
(FBS); Gibco, Grand Island, NY, USA] 1–1.5%
penicillin-streptomycin. The cells were then placed in an incubator
chamber at 37°C over a period of 24 h and the density was observed
under a microscope. The cells were passaged when they reached 80%
confluence. The medium was then removed and the culture flask was
washed with 3–5 ml D-Hanks or phosphate-buffered saline (PBS). This
was followed by the addition of 0.5–1 ml trypsin with 0.25% EDTA
for digestion. The digestion was terminated by the addition of 3–5
ml medium, followed by centrifugaqtion at 1,000 rpm/5 min. The
supernatant was then discarded and passaged according to the amount
of cells.
Construction of the recombinant
adenoviruses used in this study
NONMMUG014387 was inserted into the pHBAd-MCMV-GFP
vector. One day prior to transfection, 293 cells were seeded in a
60 mm dish and incubated at 37°C with 5% CO2 overnight
with Dulbecco's modified Eagle's medium (DMEM) supplemented with
10% FBS. The cells were transfected pHBAd-MCMV-GFP-NONMMUG014387 at
70–80% confluence. Following transfection, recombinant adenoviruses
carrying NONMMUG014387 were harvested. All of the viral particles
were purified by cesium chloride density gradient centrifugation
and tittered by the TCID50 method.
Cell proliferation assay
The MSCs were plated in 6-well plate (cell/well),
and different adenoviruses (Ad-GFP and Ad-NONMMUG014387) were added
to each well at an MOI of 20. at 48 h post-transfection, the MSCs,
and the MSC-GFP- and MSC-NONMMUG014387-transfected cells were
digested into a single cell suspension. Each group of cells was
equipped with a single cell suspension with 3×104
cells/ml concentration. Following cell adherence, 10 µl of
CCK-8 (Hanheng Biotechnology Corp., Shanghai, China) were aded to
the wells at 0, 24, 48, 72 and 96 h. The optical denstity (OD) was
at 450 nm using a spectrophotometer.
Statistical analysis
The statistical methods employed in this study were
performed using the SPSS 20.0 software packages (SPSS, Inc.,
Chicago, IL, USA). The statistical significance of the differential
expression of various lncRNAs in the micro-array analysis and
RT-qPCR validation was determined using an independent sample
t-test. GO and pathway analyses were conducted using Fisher's exact
test. Our data are expressed as the means ± standard deviation
(SD). Statistical significance was defined as P<0.01.
Results
Assessment of sciatic nerve function
index
To assess the degree of functional impairment and
recovery of locomotion following sciatic nerve injury in mice, the
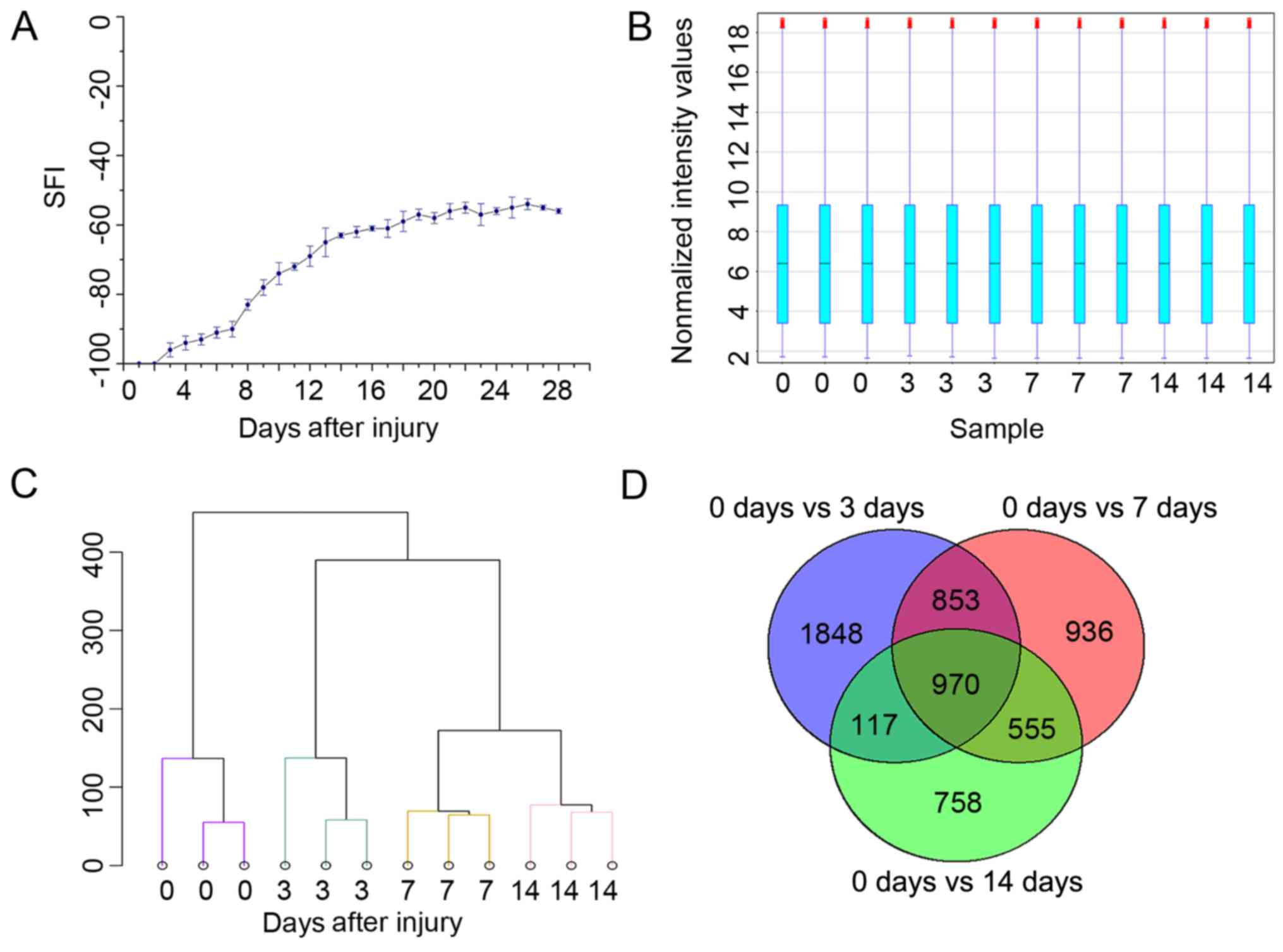
SFI was calculated daily for 28 days. The SFI values were almost
−100 immediately following PNI; after 7 days, there was rapid
neural recovery; however, after 14 days, nerve function recovery
had decelerated (Fig. 2A).
Differential expression of lncRNAs
The Agilent-074622 mouse lncRNA microarray consisted
of 64,221 lncRNA probes collected from public databases, including
GENCODE v21/Ensembl, Noncode v4 and UCSC. A total of 26,531 mRNA
probes were also present on this array. Based on the SFI results,
microarray experiments were performed at 0, 3, 7 and 14 days after
sciatic nerve crush to identify lncRNAs that were differentially
expressed during Wallerian degeneration.
The 12 specimens shared a similar level of total
characteristics, which is shown in a box plot (Fig. 2B). Sample clustering revealed that
the samples from each time group were grouped into the same cluster
(Fig. 2C). A total of 5,354
lncRNAs were identified as significantly differentially expressed
among all groups following injury (P<0.01; fold change >2).
We also compared lncRNA expression independently. A comparison
between the 0 and 3 days data revealed that 3,788 lncRNAs were
differentially expressed. Among these, 1,521 were upregulated,
while 2,267 were downregulated. A comparison between the 0 and 7
days data revealed that 3,314 lncRNAs were differentially
expressed. Among these, 1,754 were upregulated, while 1,560 were
downregulated. A comparison of the data between 0 and 14 days
revealed a total of 2,400 differentially expressed lncRNAs, 1,401
of which were upregulated and 999 down-regulated (Fig. 2D). The top 10 downregulated and
upregulated lncRNAs between the different groups are shown in
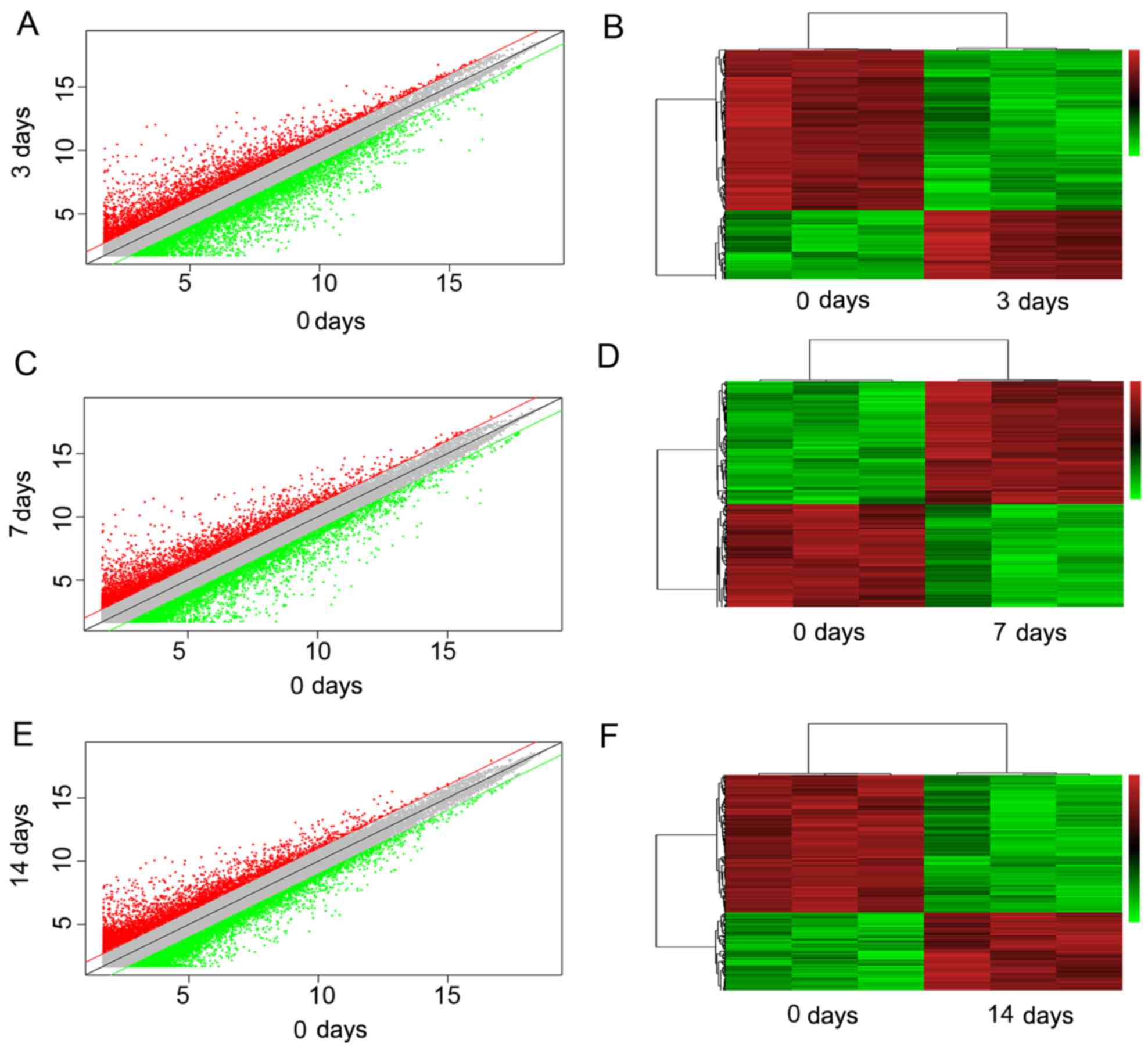
Tables IITable III–IV. A scatter plot was generated to
assess the variation in lncRNA expression among the different
groups (Fig. 3A, C and E). In
addition, hierarchical cluster analysis revealed lncRNA expression
patterns. In hierarchical clustering analysis, we used a heatmap to
indicate that differentially expressed lncRNAs at different
post-injury time -points were segregated into different clusters
(Fig. 3B, D and F). The results
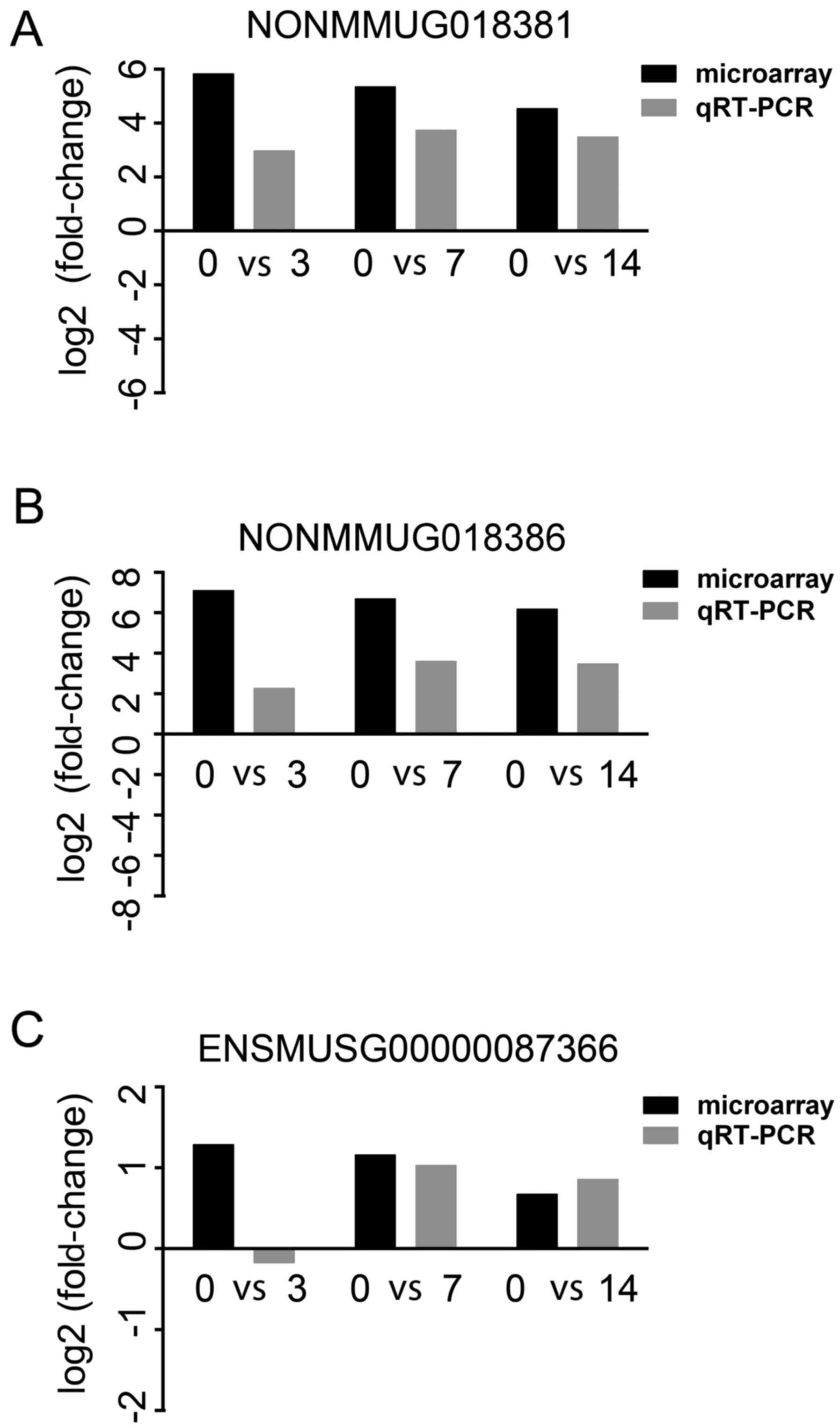
of RT-qPCR for NONMMUG018381, NONMMUG018386 and ENSMUSG00000087366
were consistent with those of the microarray analysis (Fig. 4).
| Table IITop 10 differentially expressed
lncRNAs post-injury at day 0 vs day 3. |
Table II
Top 10 differentially expressed
lncRNAs post-injury at day 0 vs day 3.
Upregulated lncRNAs
| Downregulated
lncRNAs
|
|---|
| lncRNA | P-value | Fold change | lncRNA | P-value | Fold change |
|---|
| NONMMUG032014 | 0.006677 | 365.7848 | NONMMUG013007 | 6.48E-05 | 172.2607 |
| NONMMUG032012 | 0.000117 | 342.0147 | 1500009C09Rik | 0.000838 | 97.30048 |
| NONMMUG032010 | 6.50E-05 | 328.0504 | NONMMUG013007 | 0.000995 | 83.27552 |
| NONMMUG032016 | 0.000539 | 311.5205 | NONMMUG042554 | 0.001978 | 82.2234 |
| NONMMUG010768 | 0.001339 | 188.556 | NONMMUG027331 | 0.00012 | 80.19379 |
| Gm11351 | 0.000147 | 176.1802 | NONMMUG027331 | 0.000143 | 73.40636 |
| NONMMUG013417 | 1.50E-06 | 166.4564 | NONMMUG013007 | 9.24E-07 | 63.55268 |
| NONMMUG018385 | 0.000157 | 159.6788 |
ENSMUSG00000101344 | 0.000463 | 56.43617 |
| NONMMUG018386 | 1.29E-05 | 138.9562 | NONMMUG025392 | 0.000777 | 55.75199 |
| NONMMUG027492 | 0.00123 | 117.741 | NONMMUG023470 | 0.000209 | 55.61781 |
| Table IIITop 10 differentially expressed
lncRNAs post-injury at day 0 vs day 7. |
Table III
Top 10 differentially expressed
lncRNAs post-injury at day 0 vs day 7.
Upregulated lncRNAs
| Downregulated
lncRNAs
|
|---|
| lncRNA | P-value | Fold change | lncRNA | P-value | Fold change |
|---|
| NONMMUG002067 | 0.001319 | 265.9105 | 1500009C09Rik | 0.001432 | 178.838 |
| NONMMUG032016 | 6.48E-07 | 248.2987 | NONMMUG013117 | 0.001561 | 131.4498 |
| NONMMUG032014 | 0.007602 | 225.2465 | NONMMUG042554 | 0.000353 | 109.4421 |
| NONMMUG032010 | 0.000118 | 216.8008 | NONMMUG027331 | 0.001987 | 101.779 |
| NONMMUG032012 | 0.000162 | 203.7992 | 1500009C09Rik | 0.001267 | 98.69802 |
| Bach2os | 0.000424 | 143.7661 | NONMMUG027426 | 0.00085 | 68.50972 |
| NONMMUG027492 | 0.000211 | 128.3399 | NONMMUG027331 | 0.000138 | 66.7439 |
| NONMMUG018385 | 0.000129 | 125.3089 |
ENSMUSG00000101344 | 0.002055 | 63.9863 |
| NONMMUG018386 | 1.89E-05 | 103.5612 | NONMMUG019812 | 0.000344 | 60.7132 |
| NONMMUG004899 | 3.19E-06 | 102.7997 |
ENSMUSG00000099137 | 5.63E-05 | 56.10263 |
| Table IVTop 10 differentially expressed
lncRNAs at post-injury day 0 vs 14. |
Table IV
Top 10 differentially expressed
lncRNAs at post-injury day 0 vs 14.
Upregulated lncRNAs
| Downregulated
lncRNAs
|
|---|
| lncRNA | P-value | Fold change | lncRNA | P-value | Fold change |
|---|
| NONMMUG032016 | 4.02E-06 | 92.85173 | A330015K06Rik | 0.00014 | 36.64583 |
| NONMMUG032010 | 0.000687 | 83.81547 | 1500009C09Rik | 0.00557 | 33.69628 |
| Bach2os | 0.000716 | 79.68822 | 1500009C09Rik | 0.006163 | 28.85862 |
| NONMMUG018385 | 0.000206 | 78.50652 | NONMMUG027331 | 0.000373 | 28.00702 |
| NONMMUG032012 | 0.00025 | 76.64838 | NONMMUG025427 | 0.002157 | 26.34975 |
| NONMMUG018386 | 1.79E-05 | 73.27124 | NONMMUG043073 | 0.003822 | 24.84478 |
| NONMMUG005793 | 0.00107 | 55.83829 | NONMMUG013117 | 0.006125 | 23.96197 |
| NONMMUG010751 | 3.14E-06 | 48.46715 |
ENSMUSG00000086253 | 0.001054 | 23.74235 |
| NONMMUG029505 | 0.00412 | 47.81979 | NONMMUG010960 | 0.000317 | 22.64306 |
| NONMMUG005794 | 9.66E-05 | 47.10508 | NONMMUG016086 | 0.007818 | 22.17701 |
Classification of differentially
expressed lncRNAs
The dysregulated lncRNAs were characterized by their
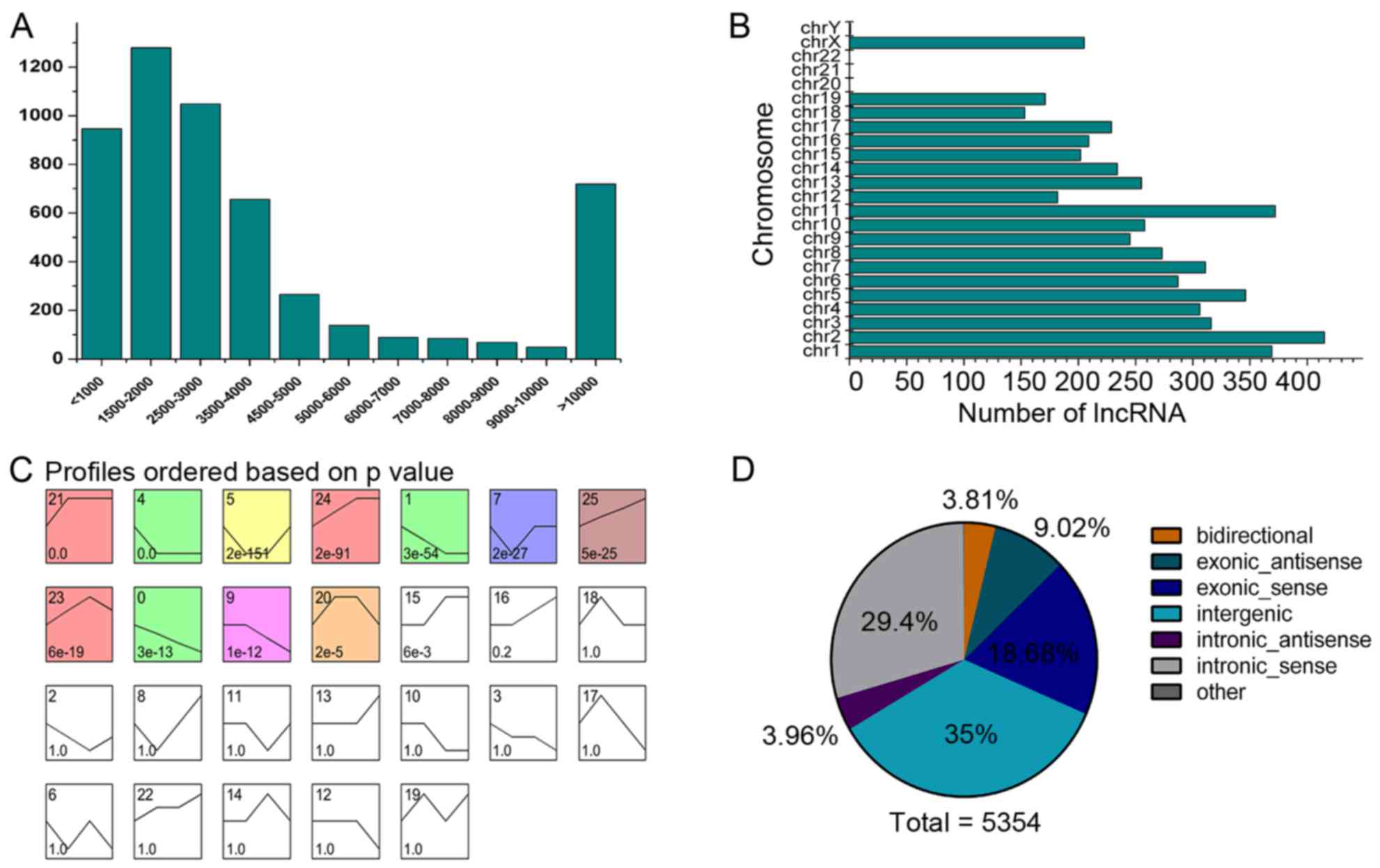
lengths and chromosomal distribution (Fig. 5A and B). Several cellular
biological processes, such as dedifferentiation, demyelination,
redifferentiation and remyelination, are involved in Wallerian
degeneration, which results from a cut or crush injury (21–23). Furthermore, lncRNA expression in
response to injury may be time-dependent. We therefore performed a
time series analysis to classify these dysregulated lncRNAs.
Consequently, 11 lncRNA profiles had significant P-values (Fig. 5C). Recent studies have
demonstrated that lncRNA transcription is often initiated
simultaneously with the expression of their overlapping or
interspersed sequences (20,21). This finding implies that the
lncRNAs that are associated with adjacent genes may be involved
inregulating gene expression. To obtain a better understanding of
this association, lncRNAs were crudely classified into five
categories according to their association with adjacent genes: i)
sense lncRNAs overlap with the exons of coding genes and are
transcribed from the same strand; ii) antisense lncRNAs overlap
with the exons of coding genes and are transcribed from the
antisense strand; iii) bidirectional lncRNAs are transcribed with a
coding transcript that is transcribed in close proximity on the
opposite strand; iv) intronic lncRNAs are derived from the introns
of coding genes; and v) intergenic lncRNAs are located within the
interval between two genes (24).
In this study, we analyzed the categorical distribution of the
5,354 lncRNAs that were differentially expressed following sciatic
nerve injury (Fig. 5D).
lncRNA target prediction and functional
analysis
To further explore the dysregulated lncRNAs, we
predicted the target genes that would be regulated using cis
and trans mechanisms. The target genes overlapped with mRNAs
that were differentially expressed at different time-points. We
chose the overlapping mRNAs for further GO analysis. For further
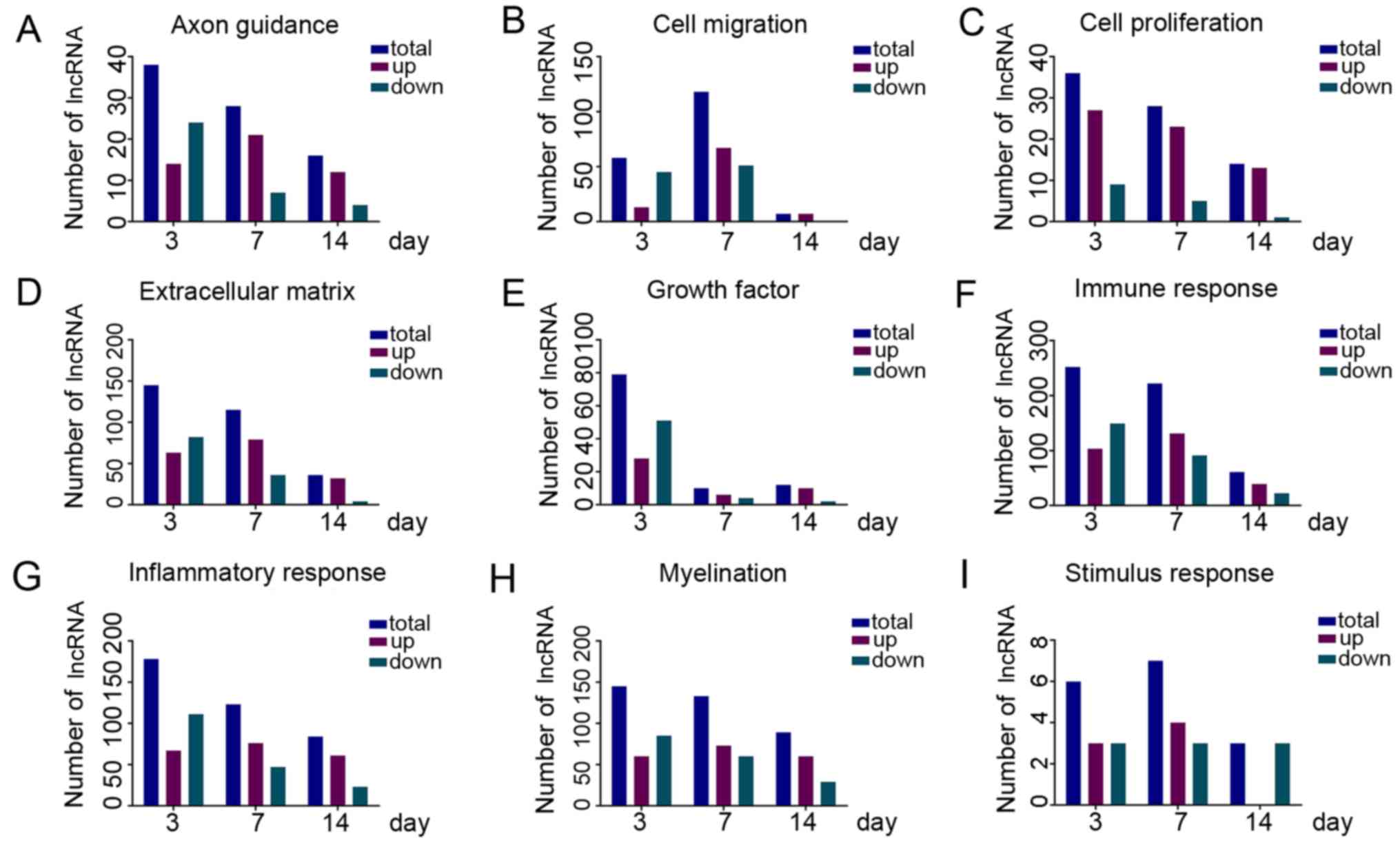
analysis, we selected 9 classes of biological processes that are
closely related to nerve regeneration following PNI, including
stimulation responses, inflammatory responses, immune responses,
cell proliferation, cell migration, axon guidance, myelination,
extracellular matrix processes, protein kinase activity and growth
factor activity. The numbers of dysregulated lncRNAs that were
predicted to participate in these processes are shown in Fig. 6.
Functional prediction of lncRNA-mRNA
pairs
Based on the classification of lncRNAs in relation
to different biological processes, we predicted the lncRNA-mRNA
pairs in the 9 biological processes that may be important for
peripheral nerve regeneration following injury, according to the
following criteria: i) the lncRNA was predicted to be involved in
the 9 biological processes according to the results described
above; ii) the lncRNA was predicted to use a cis or
trans mechanism to target mRNAs that have been reported to
be involved in peripheral nerve regeneration; iii) the mRNA has
been reported to be involved in peripheral nerve regeneration; iv)
both the lncRNA and mRNA were differentially expressed in the
microarray experiment. A list of these lncRNA-mRNA pairs is
provided in Table V.
| Table VDysregulated lncRNA transcripts and
their potential target mRNA transcripts. |
Table V
Dysregulated lncRNA transcripts and
their potential target mRNA transcripts.
| Gene symbol | Chr | Start | End | TargetGene | Regulation
type |
|---|
| NONMMUG036610 | Chr6 | 125479051 | 125480980 | CD9 | Intronic sense |
| NONMMUG036611 | Chr6 | 125493330 | 125507715 | CD9 | Exonic
antisense |
| NONMMUG036609 | Chr6 | 125476049 | 125478755 | CD9 | Intronic sense |
| LOC100504703 | Chr10 | 127070480 | 127071101 | Cdk4 | Bidirectional |
| NONMMUG014387 | Chr15 | 39076901 | 39079473 | Cthrc1 | Exonic sense |
| NONMMUG035232 | Chr6 | 47584681 | 47585827 | Ezh2 | Intronic sense |
|
ENSMUSG00000054779 | Chr3 | 37335332 | 37349740 | FGF2 | Exonic
antisense |
|
ENSMUSG00000097535 | Chr9 | 21034290 | 21037782 | Icam1 | Exonic
antisense |
| NONMMUG042235 | Chr9 | 21034152 | 21036784 | Icam1 | Exonic
antisense |
|
ENSMUSG00000087366 | Chr4 | 95052951 | 95060658 | Jun | Bidirectional |
| NONMMUG020461 | Chr18 | 82575341 | 82577314 | MBP | Exonic sense |
|
ENSMUSG00000100811 | Chr1 | 62714723 | 62718073 | Nrp2 | Intronic
antisense |
| NONMMUG042364 | Chr9 | 29327912 | 29329715 | Ntm | Intronic sense |
|
ENSMUSG00000073394 | Chr17 | 44735845 | 44737612 | Runx2 | Exonic
antisense |
| NONMMUG026340 | Chr3 | 34638297 | 34680814 | Sox2 | Exonic sense |
| SOX2OT | Chr3 | 34560380 | 34677993 | Sox2 | Exonic sense |
| NONMMUG014942 | Chr15 | 79166059 | 79216796 | SOX10 | Exonic
antisense |
|
ENSMUSG00000075555 | Chr15 | 79166066 | 79227524 | SOX10 | Exonic
antisense |
| GM10863 | Chr15 | 79166065 | 79216401 | SOX10 | Exonic
antisense |
| NONMMUG027334 | Chr3 | 96559498 | 96560013 | TXNIP | Exonic sense |
| NONMMUG036332 | Chr6 | 114683284 | 114685489 | ATG7 | trans
regulation |
| NONMMUG036586 | Chr6 | 125119767 | 125120865 | CHD4 | trans
regulation |
|
ENSMUSG00000085185 | Chr17 | 24208861 | 24221547 | FGD4 | trans
regulation |
| NONMMUG026576 | Chr3 | 52349362 | 52353221 | FOXO1 | trans
regulation |
| NONMMUG003088 | Chr10 | 14530916 | 14544995 | Gpr126 | trans
regulation |
| NONMMUG004321 | Chr10 | 87859068 | 87862787 | IGF-I | trans
regulation |
|
ENSMUSG00000084785 | Chr2 | 114654425 | 114697839 | Mbp | trans
regulation |
| NONMMUG022363 | Chr2 | 26498155 | 26503824 | Notch1 | trans
regulation |
| NONMMUG042365 | Chr9 | 29941548 | 29963141 | Ntm | trans
regulation |
| NONMMUG045895 | ChrX | 136833251 | 136834685 | Plp1 | trans
regulation |
| NONMMUG010575 | Chr13 | 28949083 | 28951671 | Sox4 | trans
regulation |
Validation of lncRNA-mRNA pairs by
RT-qPCR
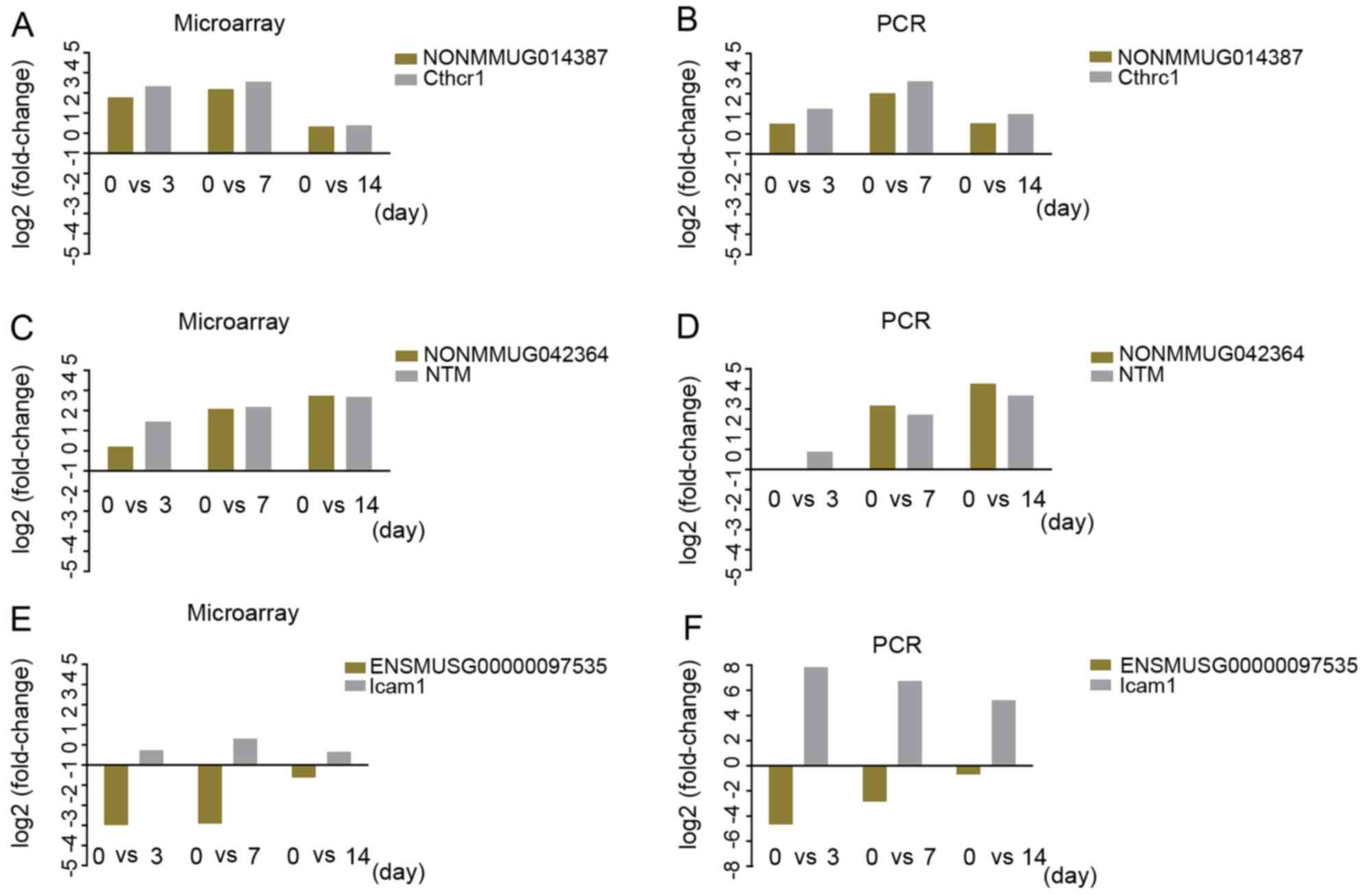
For the preliminary validation of the predicted
lncRNA-mRNA pairs, we selected three lncRNA-mRNA pairs for RT-qPCR
verification. NONMMUG014387 was predicted to have
cis-regulating potential with Cthrc1, NONMMUG042364 was
predicted to have cis-regulating potential with Ntm, and
ENSMUST00000180870 was predicted to have cis-regulating
potential with Icam1. The results of RT-qPCR and microarray
analysis revealed that NONMMUG014387 and NONMMUG042364 were
upregulated following PNI, and Cthrc1 and Ntm were also upregulated
following PNI. The expression trends of the two lncRNAs at
different time points were consistent with the separate trends of
the two mRNAs. ENSMUSG00000097535 was downregulated following PNI,
while Icam1 was upregulated following PNI, although in a different
direction; the expression level of ENSMUSG00000097535 was mostly
consistent with that of Icam (Fig.
7).
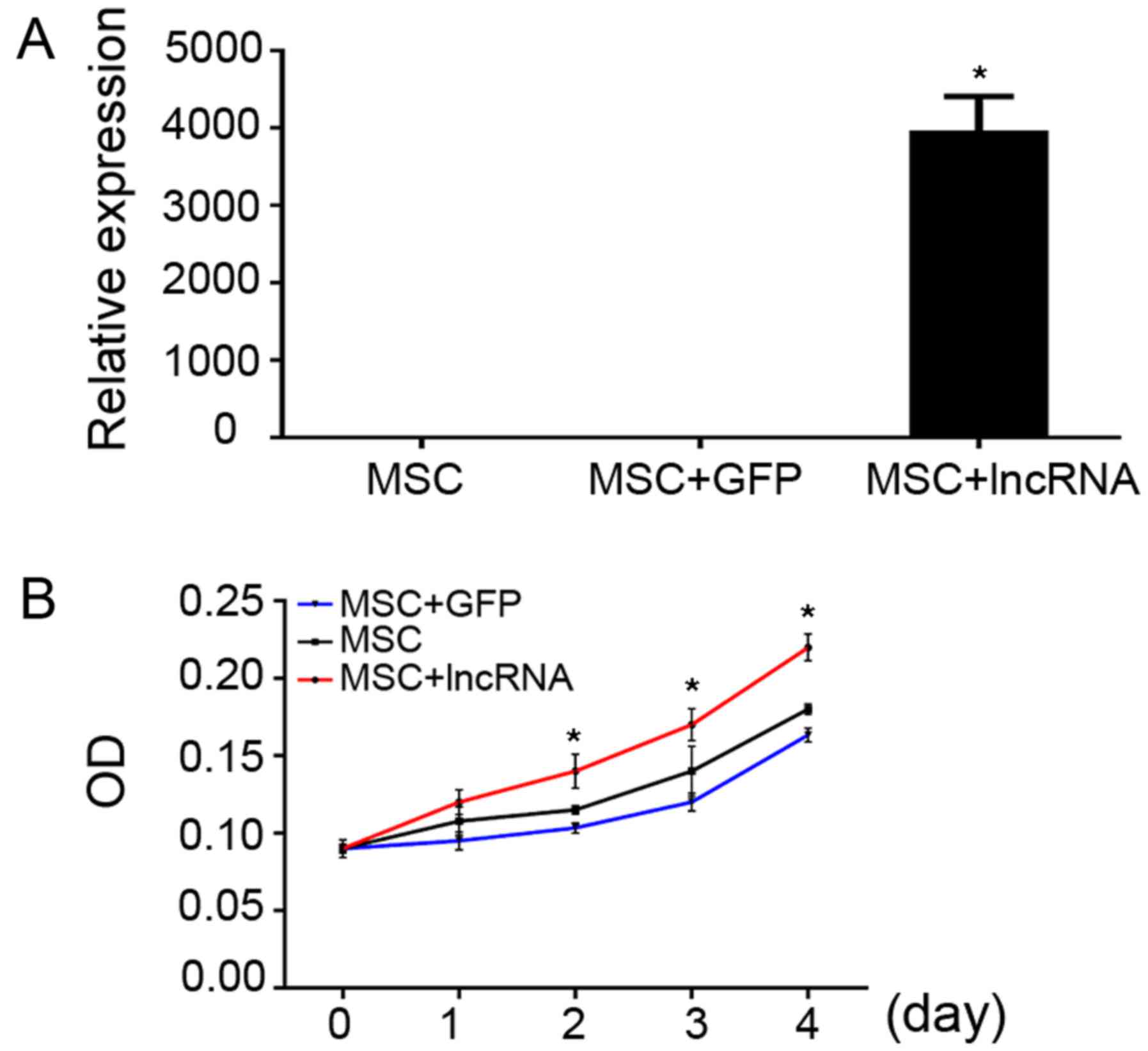
Overexpression of NONMMUG014387 enhances
MSC proliferation
NONMMUG014387 was overexpressed through recombinant
adenoviruses, the relative level of NONMMUG014387 was detected by
RT-qPCR, and the value of the MSCs transfected with NONMMUG014387
was higher than that of the control MSCs or the MSCs + GFP group
(Fig. 8A). CCK8 reavealed that
the overexpression of NONMMUG014387 in the MSCs increased cell
proliferation compared with the control MSCs and the MSCs + GFP
group d (Fig. 8B).
Discussion
To the best of our knowledge, this study is the
first to obtain a genome-wide microarray profile of lncRNA
expression at the distal end of the sciatic nerve 0, 3, 7 and 14
days following nerve crush injury and to further predict the
possible lncRNA functions using bioinformatics approaches. A total
of 5,354 lncRNAs were dysregulated following PNI, with 3,788, 3,314
and 2,400 lncRNAs dysregulated at 3, 7 and 14 days following nerve
crush injury, respectively. Validation by RT-qPCR demonstrated that
genes selected from the differentially expressed lncRNAs were
dysregulated in a manner consistent with the microarray data at
different time points. Therefore, many lncRNAs are predicted to
target key mRNAs during peripheral nerve regeneration.
It has been demonstrated that transcriptional
changes occur following sciatic nerve crush (25). Li et al demonstrated that
the expression of immune response-related genes was significantly
upregulated following nerve crush injury and rapidly peaked at 7
days post-crush (26). During the
early stage of Wallerian degeneration, immune responses leads to
debris clearance and cell death. They also found that the
expression of genes associated with cell proliferation and
migration showed a similar pattern to that of the immune response
genes, and genes associated with guidance and regeneration were
rapidly upregulated between 3 and 7 days post-crush. Li et
al also demonstrated that genes related to myelination were
downregulated following nerve crush injury and were then
upregulated between at 7 and 14 days post-injury. In this study,
SFI indicated the rapid recovery of sciatic nerve function at 7
days post-injury. However, the speed of recovery decelerated after
14 days. Overall, the gene expression data at 0, 3, 7 and 14 days
following injury may reflect the process of nerve regeneration.
As mentioned above, lncRNA-mRNA pairs were predicted
to be involved in peripheral nerve regeneration following injury.
Several lncRNA-mRNA pairs caught our attention.
Many key genes were cis-regulated by lncRNAs.
Our analysis revealed that Jun was cis-regulated by lncRNA
ENSMUSG00000087366). Jun is a protein encoded by an oncogene that
combines with c-Fos to form the AP-1 transcription factor. A
previous study demonstrated that Jun is a central regulator of
Schwann cells in response to injury (27). The knockdown of Jun has been shown
to result in strikingly compromised axonal regeneration and
functional repair (28,29). ENSMUSG00000075555, GM10863 and
NONMMUG014942 were paired with SOX10. These three lncRNAs are
located upstream of the SOX10 gene on the sense strand. Previous
studies have shown that SOX10 plays an important role in the
myelination of Schwann cells during development and is required for
myelination maintenance in adults (30,31). Following sciatic nerve crush
injury, SOX10 downregulation exerts a demyelination effect involved
in the phenotypic transition of Schwann cells (30). In addition to the effects of SOX10
effects, the downregulated expression of Sox2 also has an tight
association with lncRNAs. As shown by our results, Sox2 was
encompassed or overlapped by two differentially expressed lncRNAs
(SOX2OT and NONMMUG026340). Sox2 has been reported to be a mediator
of EphB signaling, which aids in Schwann cell sorting and guides
axon regeneration (32).
NONMMUG042235 and ENSMUSG00000097535 were paired with intercellular
adhesion molecule-1 (Icam1). It has recently been demonstrated that
Icam1 is related to both inflammation and cell recruitment in
peripheral nerve degeneration following injury, as well as to the
function of myelinogenesis in Schwann cell (33). ENSMUSG00000054779 was paired with
fibroblast growth factor-2 (FGF2). FGF2 is highly expressed in
Schwann cells and may exert paracrine actions following PNI
(21). FGF2 has also been found
to benefit motor neuron regeneration following sciatic nerve injury
and can induce Schwann cell proliferation via transforming growth
factor (TGF)-β signaling (34).
LOC100504703 was paired with cyclin-dependent kinase 4 (Cdk4),
which has been reported to be involved in the differentiation and
proliferation of Schwann cells after sciatic nerve injury (35). NONMMUG036611, NONMMUG036609 and
NONMMUG036610 were paired with CD9. CD9 promotes the migration of
Schwann cells in vitro. CD9 has been shown to participate in
the regulation of Schwann cells in response to PNI (36,37). NONMMUG027334 was paired with
thioredoxin interacting protein (TXNIP). TXNIP expression is
closely associated with the process of peripheral nerve
regeneration in vivo. TXNIP is required for advanced
glycation end products receptor (RAGE)-induced p38
mitogen-activated protein kinase (MAPK) activation. The silencing
of TXNIP affects the migration of Schwann cells and blocks
interleukin (IL)-1 and fibronectin (FN) expression (38). NONMMUG042364 was paired with
neurotrimin (Ntm). Ntm is a member of the neural cell adhesion
molecule (NCAM) family (39). The
involvement of NCAMs in cell differentiation, growth and migration
is widely accepted (40). Of
note, miR-182 reduces the migration ability of Schwann cells by
targeting Ntm at an early stage following sciatic nerve injury
(41). Whether Ntm is targeted by
both lncRNAs and microRNAs simultaneously in a competing endogenous
RNA network remains to be clarified.
Although overlapping or neighboring locations of
mRNAs and lncRNAs could establish a tight association through
cis-regulation, many lncRNAs target mRNA through
trans-regulation mechanisms (42,43). In contrast to
cis-regulating lncRNAs, trans-regulating lncRNAs
dissociate from the primary locus of transcription and influence
gene expression from a great distance. In this study, we predicted
several trans-regulatory lncRNA-mRNA pairs.
ENSMUSG00000084785 was paired with myelin basic protein (MBP)
through a trans-regulatory mechanism. MBP has been reported
to promote nerve regeneration by cleaving the neural cell adhesion
molecule L1 (44). NONMMUG004321
was paired with insulin-like growth factor-I (IGF-I). IGF-I is not
only an important mediator of growth hormone action, but is also a
neurotrophic factor for a variety of neurons. Furthermore, IGF-I
plays a key role in the development and growth of the peripheral
nervous system; systemic IGF-I treatment can promote peripheral
nerve regeneration (45).
NONMMUG003088 was paired with G-protein-coupled receptor 126
(Gpr126). The knockdown of Gpr12 has been shown to result in limb
posture abnormalities in mice, and Gpr126 is essential for
peripheral nerve development in mice (46).
All the lncRNAs discussed above were closely
associated with the target mRNAs. This study raises the possibility
that these lncRNAs may promote nerve regeneration by targeting this
set of coding genes.
This study has some limitations. First, this study
was limited by the disadvantages of microarray technology, which
can only detect the expression of known sequences;therefore, some
unknown but crucial lncRNAs involved in axon regeneration may have
been omitted. Second, due to the properties of lncRNAs and their
unclear functional mechanisms involving gene expression, it is not
possible to accurately predict the role of a specific lncRNA in an
overt biological process based on its expression level or sequence
(47). Third, the predictions of
the potential function of differentially expressed lncRNAs were not
fully verified; the determination of the definite roles of lncRNAs
depends on further experimental validation. Despite its
limitations, this study describes the lncRNA expression levels in
the distal end of the sciatic nerves for the first time;
predictions of lncRNA function are essential for future
investigations of the important role of lncRNAs in nerve
regeneration.
In conclusion, the present study, to the best of our
knowledge, provides the first evidence for temporally regulated
genome-wide lncRNA expression patterns in the sciatic nerve
following crush injuury. We predicted the function of these altered
lncRNAs based on their target mRNAs and identified lncRNAs that may
play an important role in peripheral nerve regeneration.
Acknowledgments
This study was supported by the State Key Program of
the National Natural Science Foundation of China (no. 81330042),
the Special Program for Sino-Russian Joint Research Sponsored by
the Ministry of Science and Technology, China (no. 2014DFR31210)
and the Key Program Sponsored by the Tianjin Science and Technology
Committee, China (nos. 13RCGFSY19000 and 14ZCZDSY00044). The
authors would like to thank Xiong Hui of Tianjin Medical University
(Department of Pathophysiology) and Wen-Yuan Shen of Nankai
University (School of Medicine) for providing technical
assistance.
References
|
1
|
Rochkind S: Phototherapy in peripheral
nerve regeneration: From basic science to clinical study. Neurosurg
Focus. 26:E82009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ertem K, Ceylan F, Zorludemir S, Karakoc Y
and Yologlu S: Impairment of peripheral nerve healing after nerve
repair in rats chronically exposed to alcohol. Arch Med Res.
40:325–330. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gao Y, Wang YL, Kong D, Qu B, Su XJ, Li H
and Pi HY: Nerve autografts and tissue-engineered materials for the
repair of peripheral nerve injuries: A 5-year bibliometric
analysis. Neural Regen Res. 10:1003–1008. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thomas PK and King RHM: The degeneration
of unmyelinated axons following nerve section: An ultrastructural
study. J Neurocytol. 3:497–512. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Court FA and Alvarez J: Local regulation
of the axonal phenotype, a case of merotrophism. Biol Res.
38:365–374. 2005. View Article : Google Scholar
|
|
6
|
Gao R, Wang L, Sun J, Nie K, Jian H, Gao
L, Liao X, Zhang H, Huang J and Gan S: MiR-204 promotes apoptosis
in oxidative stress-induced rat Schwann cells by suppressing
neuritin expression. FEBS Lett. 588:3225–3232. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fawcett JW and Keynes RJ: Peripheral nerve
regeneration. Annu Rev Neurosci. 13:43–60. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Akassoglou K and Strickland S: Nervous
system pathology: The fibrin perspective. Biol Chem. 383:37–45.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee JT: Epigenetic regulation by long
noncoding RNAs. Science. 338:1435–1439. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cao G, Zhang J, Wang M, Song X, Liu W, Mao
C and Lv C: Differential expression of long non-coding RNAs in
bleomycin-induced lung fibrosis. Int J Mol Med. 32:355–364.
2013.PubMed/NCBI
|
|
11
|
Lee JT and Bartolomei MS: X-inactivation,
imprinting, and long noncoding RNAs in health and disease. Cell.
152:1308–1323. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Marchese FP and Huarte M: Long non-coding
RNAs and chromatin modifiers: Their place in the epigenetic code.
Epigenetics. 9:21–26. 2014. View Article : Google Scholar :
|
|
13
|
Roberts TC, Morris KV and Wood MJ: The
role of long non-coding RNAs in neurodevelopment, brain function
and neurological disease. Philos Trans R Soc Lond B Biol Sci.
369:201305072014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mercer TR, Dinger ME, Sunkin SM, Mehler MF
and Mattick JS: Specific expression of long noncoding RNAs in the
mouse brain. Proc Natl Acad Sci USA. 105:716–721. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu B, Zhou S, Hu W, Qian T, Gao R, Ding G,
Ding F and Gu X: Altered long noncoding RNA expressions in dorsal
root ganglion after rat sciatic nerve injury. Neurosci Lett.
534:117–122. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Morrison BM, Tsingalia A, Vidensky S, Lee
Y, Jin L, Farah MH, Lengacher S, Magistretti PJ, Pellerin L and
Rothstein JD: Deficiency in monocarboxylate transporter 1 (MCT1) in
mice delays regeneration of peripheral nerves following sciatic
nerve crush. Exp Neurol. 263:325–338. 2015. View Article : Google Scholar :
|
|
17
|
Inserra MM, Bloch DA and Terris DJ:
Functional indices for sciatic, peroneal, and posterior tibial
nerve lesions in the mouse. Microsurgery. 18:119–124. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stassart RM, Fledrich R, Velanac V,
Brinkmann BG, Schwab MH, Meijer D, Sereda MW and Nave KA: A role
for Schwann cell-derived neuregulin-1 in remyelination. Nat
Neurosci. 16:48–54. 2013. View
Article : Google Scholar
|
|
19
|
Jia H, Osak M, Bogu GK, Stanton LW,
Johnson R and Lipovich L: Genome-wide computational identification
and manual annotation of human long noncoding RNA genes. RNA.
16:1478–1487. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tafer H and Hofacker IL: RNAplex: A fast
tool for RNA-RNA interaction search. Bioinformatics. 24:2657–2663.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Duobles T, Lima Tde S, Levy Bde F and
Chadi G: S100ß and fibroblast growth factor-2 are present in
cultured Schwann cells and may exert paracrine actions on the
peripheral nerve injury. Acta Cir Bras. 23:555–560. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ørom UA, Derrien T, Beringer M, Gumireddy
K, Gardini A, Bussotti G, Lai F, Zytnicki M, Notredame C, Huang Q,
et al: Long noncoding RNAs with enhancer-like function in human
cells. Cell. 143:46–58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kartha RV and Subramanian S: Competing
endogenous RNAs (ceRNAs): New entrants to the intricacies of gene
regulation. Front Genet. 5:82014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li D, Chen G, Yang J, Fan X, Gong Y, Xu G,
Cui Q and Geng B: Transcriptome analysis reveals distinct patterns
of long noncoding RNAs in heart and plasma of mice with heart
failure. PLoS One. 8:e779382013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang N, Li H, Sun Y, Yin D, Zhao Q, Cui S
and Yao D: Differential gene expression in proximal and distal
nerve segments of rats with sciatic nerve injury during Wallerian
degeneration. Neural Regen Res. 9:1186–1194. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li S, Liu Q, Wang Y, Gu Y, Liu D, Wang C,
Ding G, Chen J, Liu J, Gu X and Li S: Differential gene expression
profiling and biological process analysis in proximal nerve
segments after sciatic nerve transection. PLoS One. 8:e570002013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Parkinson DB, Bhaskaran A, Arthur-Farraj
P, Noon LA, Woodhoo A, Lloyd AC, Feltri ML, Wrabetz L, Behrens A,
Mirsky R, et al: c-Jun is a negative regulator of myelination. J
Cell Biol. 181:625–637. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vargas ME and Barres BA: Why is Wallerian
degeneration in the CNS so slow? Annu Rev Neurosci. 30:153–179.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fontana X, Hristova M, Da Costa C, Patodia
S, Thei L, Makwana M, Spencer-Dene B, Latouche M, Mirsky R, Jessen
KR, et al: c-Jun in Schwann cells promotes axonal regeneration and
motoneuron survival via paracrine signaling. J Cell Biol.
198:127–141. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Glenn TD and Talbot WS: Signals regulating
myelination in peripheral nerves and the Schwann cell response to
injury. Curr Opin Neurobiol. 23:1041–1048. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fujiwara S, Hoshikawa S, Ueno T, Hirata M,
Saito T, Ikeda T, Kawaguchi H, Nakamura K, Tanaka S and Ogata T:
SOX10 transactivates S100B to suppress Schwann cell proliferation
and to promote myelination. PLoS One. 9:e1154002014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Webber C and Zochodne D: The nerve
regenerative microenvironment: Early behavior and partnership of
axons and Schwann cells. Exp Neurol. 223:51–59. 2010. View Article : Google Scholar
|
|
33
|
Yang J, Gu Y, Huang X, Shen A and Cheng C:
Dynamic changes of ICAM-1 expression in peripheral nervous system
following sciatic nerve injury. Neurol Res. 33:75–83. 2011.
View Article : Google Scholar
|
|
34
|
Allodi I, Mecollari V, González-Pérez F,
Eggers R, Hoyng S, Verhaagen J, Navarro X and Udina E: Schwann
cells transduced with a lentiviral vector encoding Fgf-2 promote
motor neuron regeneration following sciatic nerve injury. Glia.
62:1736–1746. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Atanasoski S, Boentert M, De Ventura L,
Pohl H, Baranek C, Beier K, Young P, Barbacid M and Suter U:
Postnatal Schwann cell proliferation but not myelination is
strictly and uniquely dependent on cyclin-dependent kinase 4
(cdk4). Mol Cell Neurosci. 37:519–527. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Anton ES, Hadjiargyrou M, Patterson PH and
Matthew WD: CD9 plays a role in Schwann cell migration in vitro. J
Neurosci. 15:584–595. 1995.PubMed/NCBI
|
|
37
|
Hadjiargyrou M and Patterson PH: An
anti-CD9 monoclonal antibody promotes adhesion and induces
proliferation of Schwann cells in vitro. J Neurosci. 15:574–583.
1995.PubMed/NCBI
|
|
38
|
Sbai O, Devi TS, Melone MA, Feron F,
Khrestchatisky M, Singh LP and Perrone L: RAGE-TXNIP axis is
required for S100B-promoted Schwann cell migration, fibronectin
expression and cytokine secretion. J Cell Sci. 123:4332–4339. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Grijalva I, Li X, Marcillo A, Salzer JL
and Levi AD: Expression of neurotrimin in the normal and injured
adult human spinal cord. Spinal Cord. 44:280–286. 2006. View Article : Google Scholar
|
|
40
|
Bachelin C, Zujovic V, Buchet D, Mallet J
and Baron-Van Evercooren A: Ectopic expression of polysialylated
neural cell adhesion molecule in adult macaque Schwann cells
promotes their migration and remyelination potential in the central
nervous system. Brain. 133:406–420. 2010. View Article : Google Scholar :
|
|
41
|
Yu B, Qian T, Wang Y, Zhou S, Ding G, Ding
F and Gu X: miR-182 inhibits Schwann cell proliferation and
migration by targeting FGF9 and NTM, respectively at an early stage
following sciatic nerve injury. Nucleic Acids Res. 40:10356–10365.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lutz D, Kataria H, Kleene R, Loers G,
Chaudhary H, Guseva D, Wu B, Jakovcevski I and Schachner M: Myelin
basic protein cleaves cell adhesion molecule L1 and improves
regeneration after injury. Mol Neurobiol. 53:3360–3376. 2016.
View Article : Google Scholar
|
|
45
|
Gao WQ, Shinsky N, Ingle G, Beck K, Elias
KA and Powell-Braxton L: IGF-I deficient mice show reduced
peripheral nerve conduction velocities and decreased axonal
diameters and respond to exogenous IGF-I treatment. J Neurobiol.
39:142–152. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Monk KR, Oshima K, Jörs S, Heller S and
Talbot WS: Gpr126 is essential for peripheral nerve development and
myelination in mammals. Development. 138:2673–2680. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bassett AR, Akhtar A, Barlow DP, Bird AP,
Brockdorff N, Duboule D, Ephrussi A, Ferguson-Smith AC, Gingeras TR
and Haerty W: Considerations when investigating lncRNA function in
vivo. eLife. 3:e030582014. View Article : Google Scholar : PubMed/NCBI
|