Introduction
Human articular cartilage tissue is characterized by
a rich extracellular matrix (ECM) and a low cell density. The few
chondrocytes are embedded in the ECM which is produced and retained
by the former (1). Characteristic
components of the ECM are collagen fibrils of collagen type II
(Col2) and the proteoglycans aggrecan and chondroitin sulphate, and
sulphated glycosaminoglycans (sGAG) (2).
Control of the chondrocytic phenotype within the
physiological microenvironment is the main challenge in cartilage
tissue engineering and cartilage repair (3). A current strategy in orthopedic
surgery is autologous chondrocyte transplantation (ACT) and its
advancements. In this two-step procedure, chondrocytic cells are
isolated from an unaffected part of the patient's articular knee,
expanded in monolayer cell culture, and finally implanted in the
cartilage lesion. The implantation of the autologous cells can be
carried out either with biomaterial constructs, or as suspension or
3D spheroid cultures without the use of biomaterials. For the
phenotypical appearance and function of chondrocytes, the
microenvironment of the cartilage tissue is substantial (4). Although the technique of ACT opens
up new regenerative opportunities, this therapeutic approach is
limited by the availability of unaffected cells (5) as well as their dedifferentiation
potential to fibroblasts during in vitro expansion. This
dedifferentiation process is characterized by an increased
expression of characteristically fibrogenous proteins such as type
I collagen (Col1) whereas cartilage-specific markers, such as
collagen types II, XI, IX and proteoglycans like aggrecan (ACAN)
are decreased (6,7). Fibroblastic cells are not capable to
assemble an ECM which resists the mechanical forces within the
joint.
An enhanced approximation of in vitro
conditions to physiological conditions, such as culturing in 3D or
reduced oxygen condition appears to promote the process of
redifferentiation of the cells into a cartilaginous phenotype
(3,8). For this reason, the further
development of the in vitro cultivation of chondrocytic
cells in 3D could be beneficial, since spheroid cultures should
sustain the chondrocytic phenotype during the process of ACT
(9). Nevertheless, the balance
between production and availability of essential growth factors for
cartilage formation should not be neglected. Since growth factors
are stored in the ECM and are released under specific conditions to
regulate cellular behaviour (10), the isolation of chondrocytic cells
from their natural environment reduces the availability of these
growth factors. Exogenous growth factors are able to induce the
formation of cartilage-like tissue (7,11).
Therefore, knowledge concerning the interaction of specific growth
factors for cartilage regeneration is a prerequisite for
high-quality hyaline cartilage.
Chondrogenic growth factors such as the insulin-like
growth factor (IGF)-1 and transforming growth factor (TGF)-β1 are
able to influence the expression of Col2 and GAGs within the
cartilage tissue and thus influence chondrogenic redifferentiation
(12). IGF-1 is an important
anabolic factor for the stimulation of chondrocyte proliferation
and matrix synthesis (13-15).
Its activity is mediated via membranous IGF-1 receptors (IGFR1)
(16).
TGF-β1 is known to be an important stimulator of
proteoglycans and Col2 synthesis in chondrocytes (17–19) and is able to induce the
chondrogenic differentiation of mesenchymal stem cells in
vitro (8). Effects emerging
from TGF-β1 are carried out via TGF-β receptors existing as
homodimeric or heterodimeric isoforms. Signals are transmitted by
forming complexes consisting of dimers of TGF-β receptor 1
(TGFBR1) and 2 (TGFBR2) (20,21).
In previous studies, we showed that the addition of
the chondrogenic growth factors TGF-β1 and IGF-1 can influence
chondrogenic redifferentiation in 3D cultures of alginate, spheroid
and matrix-associated cultures (22–24). Moreover, we demonstrated that
reduction of the oxygen content to more physiological conditions
(5%, hypoxia) improved the effects of growth factors on the
redifferentiation process, especially for IGF-1 (22). In the present study, it was
assumed that the reduced IGF-1 activity in chondrocytes under
normoxia resulted from the enrichment of reactive oxygen species
(22). Nevertheless, we observed
donor-specific reactions on growth factor treatment, suggesting
donor-specific expression profiles of IGF-1 and TGF-β receptors on
chondrocytic cells. Therefore, the aim of the present study was to
evaluate IGF-1 and TGF-β receptor mRNA expression levels in human
chondrocytes in monolayer and 3D pellet cultures. A comparison of
monolayer and pellet cultures should clarify whether a gene
expression profile is suitable for therapeutic strategies and what
type of cultivation is more effective. Doing this, we hypothesized
that a targeted application of chondrogenic growth factors could
improve cartilage tissue engineering strategies to optimize repair
techniques such as ACT. Hence, we analyzed the impact of IGF-1 and
TGF-β1 as well as a combination of both on the gene expression
rates of collagen type I αI (COL1A1), type II (COL2A1), as
well as alkaline phosphatase (ALP) and collagen type X αI
(COL10A1) as hypertrophic markers in pellet cultures over a
period of 5 weeks. Additionally, amounts of Col2 protein as well as
GAGs were quantified. We also examined the reduction of the oxygen
content during cultivation with respect to the effect of different
growth factors on chondrogenic cell behaviour.
Materials and methods
Isolation and cultivation of human
chondrocytes
Human primary chondrocytes (female: n=3, age 36–44
years; male: n=2, age 44–47 years) were isolated post mortem from
unaffected articular knee cartilage within 24 h to 6 days of death.
All tissues did not show any signs of osteoarthritis or
degeneration which was examined macroscopically. Isolation of cells
was performed as described in the protocol of Jonitz et al
(22). Cells were separated into
two groups: i) hypoxia (5% O2) and ii) normoxia (21%
O2) and incubated in a humidified atmosphere at 37°C in
5% CO2. The tissue removal was approved by the Local
Ethics Committee (registration no. A2011-138).
Stimulation of monolayer culture with
chondrogenic growth factors
In passage three, chondrocytes were transferred to
48-well plates with 10,000 cells/well in Dulbecco's modified
Eagle's medium (DMEM) containing 10% fetal calf serum (FCS), 1%
penicillin/streptomycin, 1% amphotericin B (all from Invitrogen,
Darmstadt, Germany) and ascorbic acid (final concentration, 50
µg/ml; Sigma, St. Louis, MO, USA).
After 24 h of adherence under the described
conditions (normoxia and hypoxia), the medium was replaced by
serum-free medium containing ascorbic acid (final concentration: 50
µg/ml; Sigma), dexamethasone (final concentration, 100 nM;
Sigma), ITS™ (complete medium and ITS™ in a 100:1 ratio;
Becton-Dickinson, Heidelberg, Germany) and chondrogenic growth
factors, IGF-1 (R&D Systems, Wiesbaden, Germany) and/or TGF-β1
(Tebu-Bio, Offenbach, Germany) in three different combinations
(Table I). The chosen growth
factor concentrations are based on previous studies (22–24). The stimulation was performed in
duplicates, at 37°C in a humidified atmosphere of 5% CO2
and the appropriate oxygen concentration for 24, 48 and 72 h,
respectively. The beginning of the chondrogenic stimulation was
defined as time point 0.
 | Table IThree different growth factor
combinations in the cell medium. |
Table I
Three different growth factor
combinations in the cell medium.
| Combination 1 | Combination 2 | Combination 3 |
|---|
| TGF-β1 | 50 ng/ml | – | 50 ng/ml |
| IGF-1 | – | 50 ng/ml | 50 ng/ml |
Stimulation of pellet culture with
chondrogenic growth factors
From 3 of the 5 donors, the stimulation was
additionally performed as 3D pellet cultures. For pellet formation,
5×104 cells/well (passage three) were transferred into
96-well V-bottom suspension-plates suspended in the same three
medium combinations as mentioned above (Table I). After centrifugation at 400 × g
for 5 min for sedimentation, the cells were incubated for 24, 48,
72 h as well as 14 and 35 days under standard culture conditions
and a 5 or 21% oxygen level, respectively. The medium was changed
every 2–3 days. For the start point (named as 0) 5×104
cells suspended in serum-free medium with ITS were used and the
same pellet formatting procedure was used as described above.
RNA extraction and cDNA synthesis
For RNA isolation, the monolayer cells were directly
lysed in the well. Pellet cultures were transferred with lysis
buffer into 2 ml homogenisation tubes containing small steel beads
(Precellys-Steel kit, 2.8 mm; PeqLab Biotechnology GmbH, Erlangen,
Germany) and homogenized for 30 sec at 5,000 × g.
Isolation of RNA was performed using the peqGOLD
Total RNA kit (PeqLab Biotechnology GmbH) which relies on the
principle of column purification of RNA probes according to the
supplier's recommendations.
For cDNA synthesis, the High Capacity cDNA kit
(Applied Biosystems, Foster City, CA, USA) was used. An amount of
10 ng isolated RNA of each sample was used and operated according
to the manufacturer's instructions.
Quantitative (real-time) PCR (qPCR)
Relative quantification of target cDNA levels was
performed by qPCR (qTower 2.0; Analytik Jena AG, Jena, Germany)
using the InnuMIX qPCR MasterMix SyGreen (Analytik Jena AG) and the
cDNA primers (Sigma-Aldrich) which are listed in Table II. PCR was performed following
the manufacturer's instructions undergoing the following process:
95°C for 2 min, 40 cycles of 95°C for 5 sec and 65°C for 25 sec.
The reactions were performed as duplicates.
| Table IIcDNA primers for qPCR. |
Table II
cDNA primers for qPCR.
| Primer | Sequence 5′→3′
(forward and reverse) |
|---|
| ALP |
CATTGTGACCACCACGAGAG |
|
CCATGATCACGTCAATGTCC |
| COL1A1 |
ACGAAGACATCCCACCAATC |
|
AGATCACGTCATCGCACAAC |
| COL2A1 |
AATGGTGGCTTCCATTTCAG |
|
GTGATGTTCTGGGAGCCTTC |
| COL10A1 |
GAACTCCCAGCACGCAGAATC |
|
AGTGGGCCTTTTATGCCTGT |
| IGF1R |
CGAGTGGAGAAATCTGCGGG |
|
ACTCGGTAATGACCGTGAGC |
| TGFBR1 |
TTCCAACTACTGGTTTACCATTGC |
|
AATCTCTGCCTCACGGAACC |
| TGFBR2 |
GTCACTGACAACAACGGTGC |
|
TCTTTCTCCATACAGCCACACAG |
| ACTB |
CTTCCTGGGCATGGAGTC |
|
AGCACTGTGTTGGCGTACAG |
For further analyses the ΔΔCq method was applied.
The relative expression of each mRNA compared to the housekeeper
β-actin was calculated according to the equation: ΔCq =
Cqtarget − Cqβ-actin.
The relative amount of target mRNA at each time
point (24, 48, 72 h and at 14 and 35 days) was normalized to the
mRNA level of unstimulated cells at time point 0 using the equation
ΔΔCqstimulation = ΔCqgene −
ΔCqcontrol. The resulting gene expression is presented
as the 2−ΔΔCq value.
The gene expression of the following chondrogenic
marker genes was analyzed: COL1A1 and COL2A1,
IGF-1-receptor (IGF1R), TGF-β-receptor 1 and 2
(TGFBR1/TGFBR2). Furthermore, gene expression of ALP
and collagen type X α1 (COL10A1) was determined to detect
cell hypertrophy. Each gene was analyzed in duplicates.
Pro-Col2 quantification
Protein amounts of Col2 synthesized by chondrocytic
pellet cultures were analyzed using an ELISA (Ibex Pharmaceuticals,
Inc., Mont-Royal, QC, Canada). The principal of the ELISA is based
on the cleavage of the carboxy propeptides of newly formed collagen
which were measured in the supernatant of pellet cultures. Hence,
supernatants of every medium combination from pellet cultures were
collected after 24, 48, 72 h as well as after 14 and 35 days of
incubation and stored at −20°C. The ELISA was performed according
to the manufacturer's instructions. Absorbance was measured at 450
nm using Opsys MR™ microplate reader (Dynex Technologies,
Denkendorf, Germany). The content of Col2 was normalized to the
total protein content (ng Col2/mg proteintotal).
Total protein determination
Total protein content of the collected supernatants
was determined using the Qubit® protein assay kit
(Thermo Fisher Scientific, Waltham, MA, USA) following the
manufacturer's instructions manual. Protein content was calculated
using a defined standard curve. The supernatants of each pellet
culture at every time point were collected and stored at −20°C. A
cell-free medium combination incubated for 3 days served as
background control.
Blyscan™ GAG αssay
The Blyscan™ assay (Biocolor Ltd., Carrickfergus,
Northern Ireland, UK) is a quantitative dye-binding method used to
measure sulfated proteoglycans and GAGs. The supernatants of the
stimulated pellet cultures were collected after 24, 48, 72 h as
well as 14 and 35 days. After papain digestion at 60°C overnight
(20 units/mg papain from papaya latex; Sigma-Aldrich) in 50 mM
phosphate buffer [1 M NaCl, 5 mM cystein-HCL, 1 mM EDTA (Merck
Millipore, Darmstadt, Germany)] the Blyscan™ assay was performed
according to the manufacturer's instructions. Absorbance was
measured at 656 nm using the Opsys microplate reader (Dynex
Technologies). The content of sGAG was normalized to total protein
content (µg sGAG/mg proteintotal).
Histology
For histological investigations, human chondrocytes
were cultured as pellet cultures and stimulated with TGF-β1 and/or
IGF-1 either under normoxia or hypoxia. After 35 days of
incubation, the pellet cultures were embedded in
Tissue-Tek® O.C.T.™ compound, stored at −80°C and
afterwards sectioned to 7–10 µm slices using a Leica
cryostat. Sections were directly mounted on pretreated slides
(Superfrost Plus; Menzel Glaser, Braunschweig, Germany), air dried,
and fixed in acetone/methanol (1:2) at room temperature for 10 min.
The slices were analyzed using Heidenhain's AZAN trichrome
staining. The conventional dyes azocarmine G and aniline blue were
used providing red nuclei as well as blue collagen-rich ECM and
acid mucus substances.
Statistical analysis
Data in the diagrams are presented as box plots.
Boxes denote interquartile ranges, horizontal lines within the
boxes denote medians, and whiskers denote minimum and maximum
values (n=5). All statistical analyses were performed using SPSS
statistics 22 (IBM Deutschland, Ehningen, Germany). Since all
cultures of each specific donor were obtained from the same
starting cultures, measurements could be estimated as repeated ones
from a single sample. Therefore, the statistical significance was
calculated with the Wilcoxon signed rank rest. P<0.05 was
considered to indicate a statistically significant difference.
Results
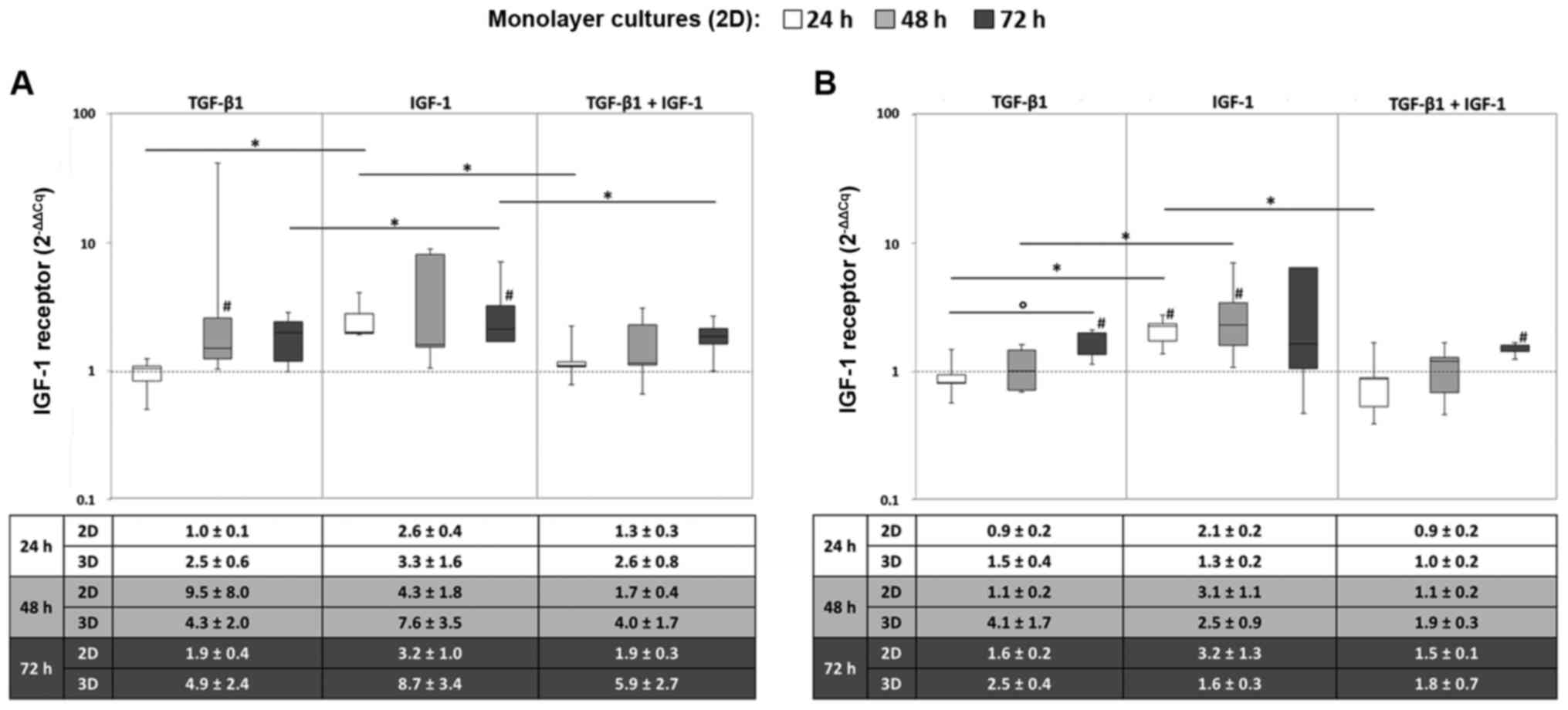
Gene expression rates of the IGF-1
receptor
In monolayer cultures, the IGF1R gene was
mainly expressed when chondrocytes were stimulated with IGF-1
alone. Here, the mRNA levels after 72 h were significantly higher
after IGF-1 stimulation compared to TGF-β1 and/or IGF-1 stimulation
under hypoxic culture conditions (all: P=0.043) (Fig. 1A). Under normoxia, significant
differences between the sole stimulation with TGF-β1 and IGF-1 were
also detectable after 24 and 48 h (both: P=0.043) (Fig. 1B).
Additionally, an increased expression level of
IGF1R was observed over time, which was more marked for the
stimulation with TGF-β1 and the combination of TGF-β1 and IGF-1.
Indeed, this can be shown by a significant increase under normoxia
and TGF-β1 stimulation from 24 to 72 h (P=0.043) (Fig. 1). Furthermore, comparable
tendencies in gene expression levels between monolayer and pellet
cultures regarding IGF1R expression were detected.
Nevertheless, the resulting differences were more decisive for
cells cultured in monolayer. This behaviour is exemplified by the
mean values ± SEM presented in the tables below the diagrams
(Fig. 1).
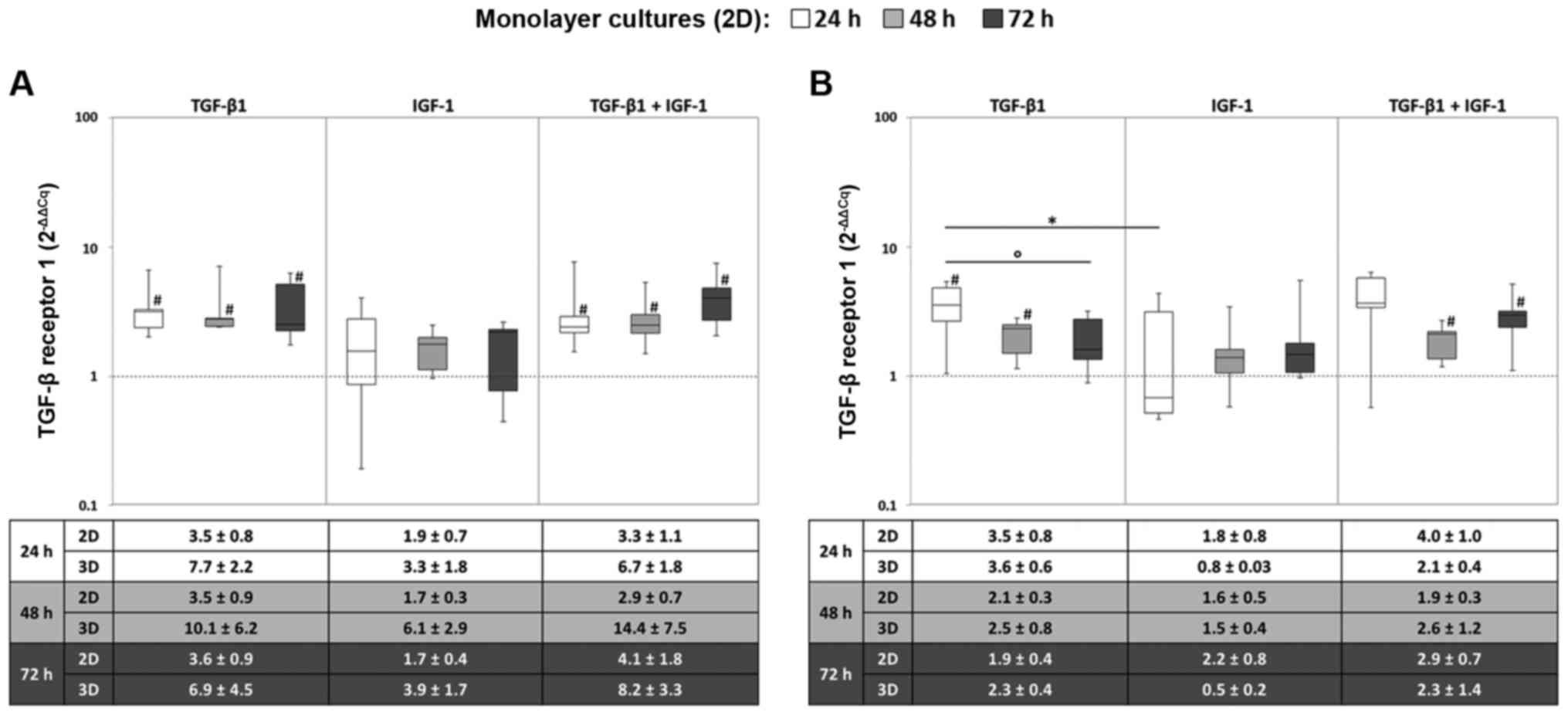
Gene expression rates of the TGF-β-
receptors
The expression of the TGFBR1 gene was
enhanced after stimulation with TGF-β1 alone or in combination with
IGF-1. It was apparent that the highest TGFBR1 mRNA level
was reached after 24 h with no marked changes over time. Hence,
under hypoxic conditions, the expression levels were significantly
enhanced in comparison to the starting point for TGF-β1 and the
combined stimulation (all: P=0.043) (Fig. 2A). However, a significant
difference between the different growth factors was observed under
normoxia for TGF-β1 and IGF-1 stimulation after 24 h (P=0.043)
(Fig. 2B). Moreover, comparing
mRNA levels of monolayer and pellet cultures, similar patterns
could be detected for the TGFBR1 gene expression profile,
although differences in the expression levels were more pronounced
in the monolayer culture.
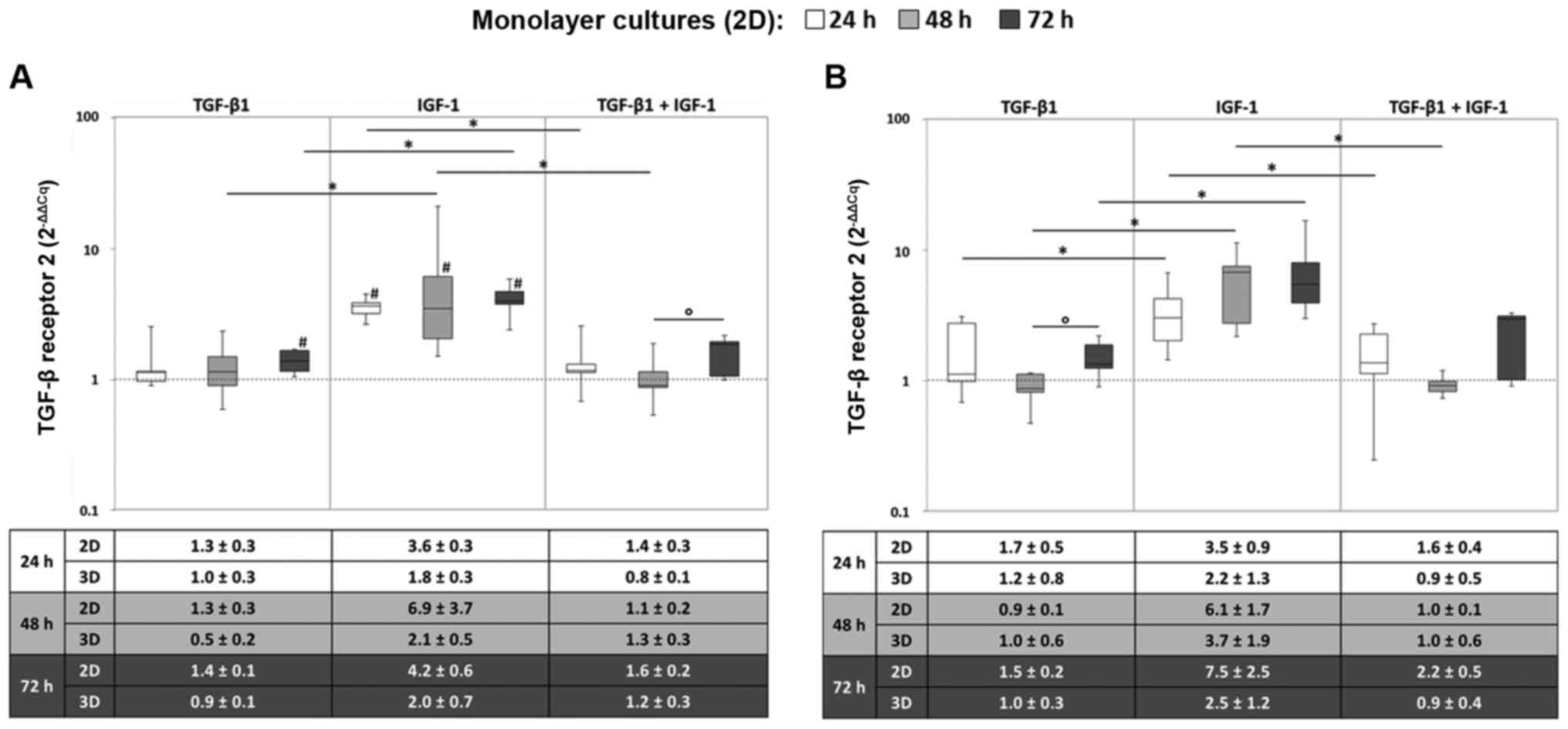
Regarding TGFBR2 gene expression, a notable
increase in the mRNA levels was observed under IGF-1-stimulation.
As in the same way observed for TGFBR1, TGFBR2
reached maximum expression levels already after 24 h, which did not
considerably change during further cultivation. Thus, compared to
the starting point, significantly enhanced expression levels for
all measured time points were determined under hypoxic conditions
after IGF-1 stimulation (all: P=0.043). Additionally, TGF-β1
stimulation resulted in significantly enhanced expression levels
after 72 h in comparison to the starting point (P=0.043) (Fig. 3A). Under normoxic culture
onditions, TGFBR2 was significantly expressed at a higher level
under IGF-1 stimulation compared to the sole stimulation with
TGF-β1 at all measured time points and compared to the combined
stimulation TGF-β1 and IGF-1 after 24 and 48 h (all: P=0.043)
(Fig. 3B). Again, the gene
expression levels between monolayer and pellet cultures revealed
comparable tendencies.
Overall, no significant differences in receptor gene
expression behaviour comparing monolayer and pellet culture was
determined. Characteristic gene expression profiles of the growth
factor receptors were determined in regards to the applied growth
factors, although the values were partially widely scattered among
the 5 donors. Moreover, it was apparent that IGF1R showed an
increase in mRNA levels over time whereas both TGFBRs
already reached maximum expression levels after 24 h and remained
almost constant over time.
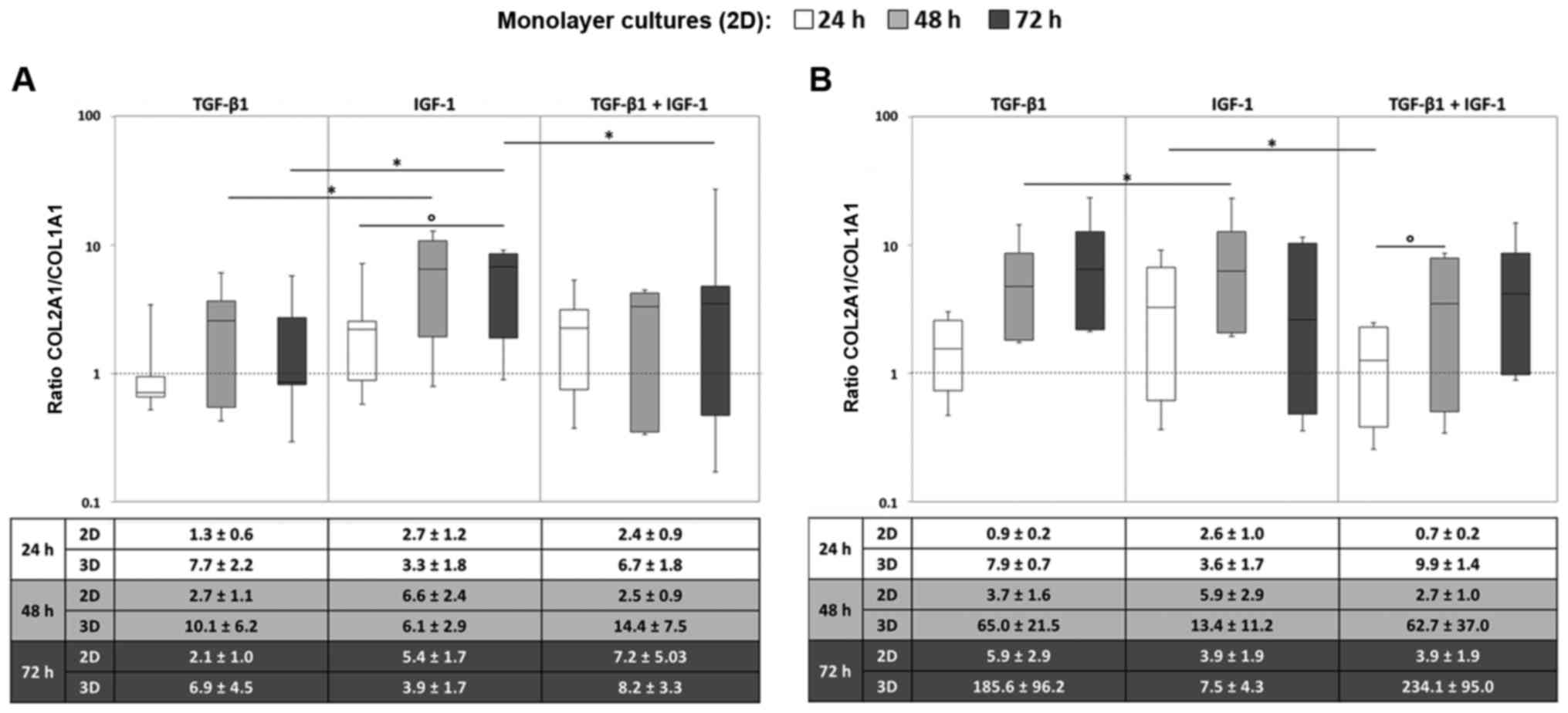
Gene expression of COL1A1 and COL2A1
In order to analyze the relation between
COL1A1 and COL2A1, and therefore the quality of
cartilogenous pellet cultures, COL2A1/COL1A1 ratios
were determined.
In most cases, we observed an increase in the ratio
over time. In general, the ratios for the monolayer cultures
reached higher values under stimulation with IGF-1 in comparison to
the stimulation with TGF-β1 or the combination of TGF-β1 and IGF-1.
This was observed for both oxygen conditions. Most of the donors
showed a time-dependent increase in the ratios, especially when
they were stimulated with TGF-β1 or the combination. In contrast to
that, the ratios under IGF-1 stimulation altogether fluctuated at
higher values, but rarely increased in a steady manner as was the
case for TGF-β1 and the combination (Fig. 4).
Upon comparing the monolayer and pellet cultures,
the COL2A1/COL1A1 ratios reached higher values for the
pellet cultures especially when they were stimulated with TGF-β1 or
with both growth factors. Similar to the monolayer culture under
hypoxic conditions, a time-dependent increase in the ratios in
chondrocytic pellets was detected after treatment with TGF-β1 or
the combination (Fig. 4A). This
increase over time was even more marked for the pellet cultures.
Nonetheless, regarding the COL2A1/COL1A1 ratios, no
significant differences for the ratios between the two culturing
methods were found.
In addition, a prominent increase in the ratios in
the pellet cultures after 14 and 35 days after stimulation with
TGF-β1 and the combination was detected. After 35 days we observed
higher ratios for the combination of IGF-1 and TGF-β1 compared to
the sole stimulation. This was detectable for both oxygen
conditions but was more pronounced under hypoxia. Nevertheless, the
ratios were widely scattered among the three donors, which is
indicated by the high standard errors of the mean (SEM) (Fig. 4).
Gene expression of hypertrophic
markers
In order to detect hypertrophy of cells, the gene
expression of ALP and COL10A1 was analyzed from
pellet cultures after 24, 48 and 72 h as well as after 14 and 35
days.
Gene expression of ALP was not induced in the
chondrocytic pellet cultures. In most of the cases the gene
expression level was lower than that of the unstimulated cells at
time point 0 or did not extend detectable ranges. The highest
detected value was a 1.7-fold increase compared to the unstimulated
control obtained for TGF-β1 stimulation after 48 h of cultivation
under hypoxia (for donor 2), but the expression level decreased
over time to a 0.07-fold value (35 days, TGF-β1 stimulation).
Furthermore, the expression levels did not differ between the
different stimulation types or between the two oxygen conditions.
COL10A1 gene expression was not detectable in most of the
samples. There were no detectable expression levels at time point
0, hence 2−ΔΔCq values could not be calculated. Instead,
expression levels were related to the expression of the housekeeper
gene β-actin. Some low expression rates were detected in cultures
treated with TGF-β1 or the combination of TGF-β1 and IGF-1.
Nevertheless, the detected rates did not exceed 0.1% of the
expression level of the housekeeper.
Synthesis rates of hyaline cartilage-like
ECM components
In addition to gene expression analyses, levels of
the hyaline-characteristic proteins Col2 as well as sGAG were
quantified from supernatants of the chondrocytic pellet cultures to
determine the redifferentiation potential.
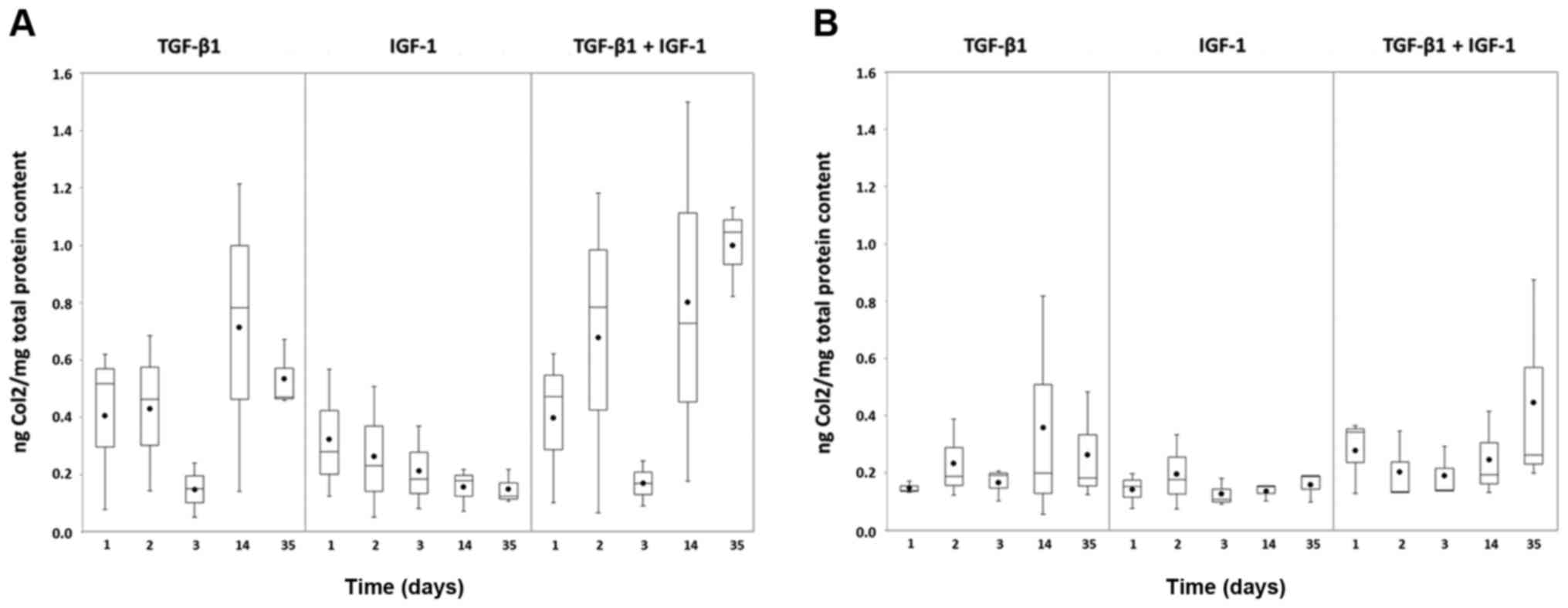
Overall the measured Col2 amounts were higher under
hypoxia (Fig. 5A) than under
normoxia (Fig. 5B). Comparing the
different stimulations, greatest protein amounts were reached with
TGF-β1 and the combination of TGF-β1 and IGF-1. Nevertheless, a
conspicuous decrease was observable for these two stimulations
under hypoxia at day 3 with increasing values at the later measured
points (14 and 35 days) (Fig.
5A).
Only less Col2 protein could be detected from the
pellet cultures stimulated with IGF-1. Under hypoxia the amount
decreased over time (Fig. 5A),
whereas under normoxia it remained at a low level (Fig. 5B).
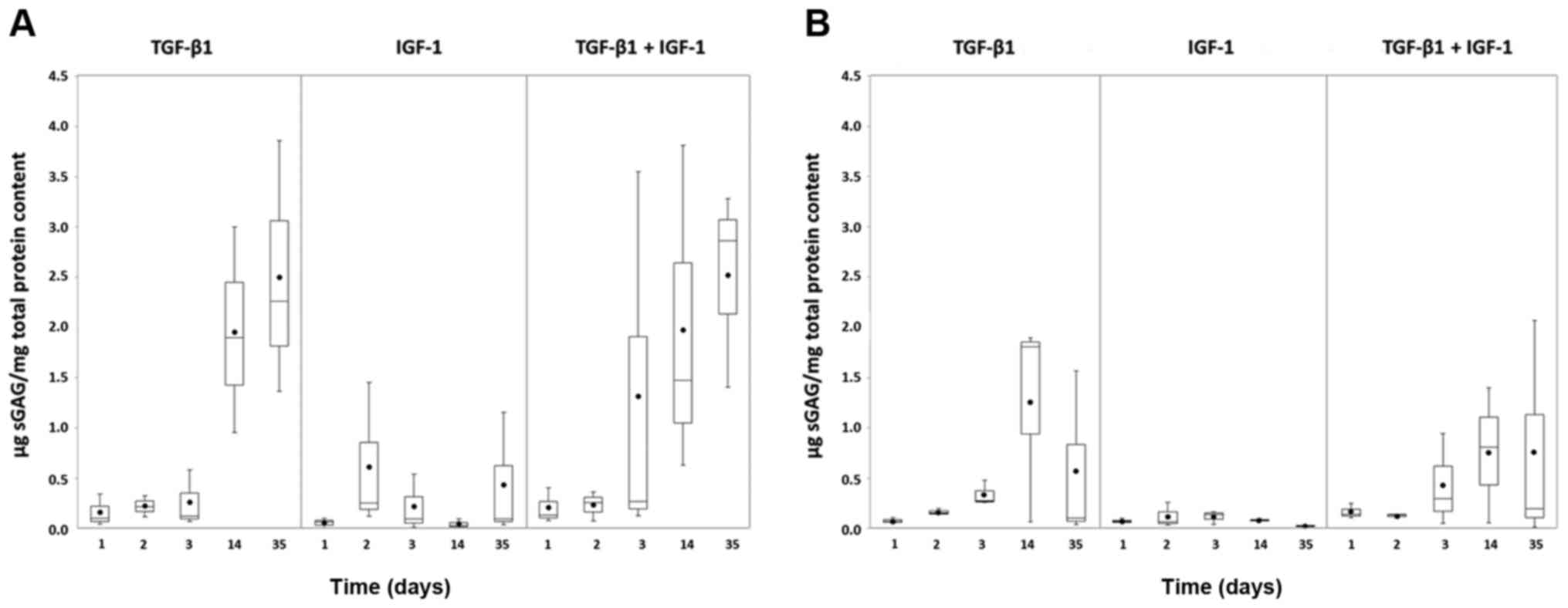
Similar to Col2 protein amounts, the sGAG reached
higher levels under hypoxia (Fig.
6A) than under normoxia (Fig.
6B). The amount of sGAG produced by the pellet cultures
stimulated with TGF-β1 alone or in combination with TGF-β1 and
IGF-1 increased over time. This tendency was more visible under
hypoxia (Fig. 6A). In contrast,
under both oxygen conditions, only low amounts of sGAG were
detected under IGF-1 stimulation which remained almost stable over
time.
Morphological and histological
characterization of pellet cultures
During incubation of the chondrocyte pellet cultures
over 35 days under different growth factor combinations and oxygen
conditions the cell pellets developed unequal sizes. After 35 days
of incubation, the pellets stimulated with TGF-β1 or the
combination of TGF-β1 and IGF-1 were approximately twice as large
as pellets stimulated with IGF-1 alone. For pellets cultured under
normoxia, we could even observe a reduction in pellet size over
time. This difference in size after 35 days of culturing is
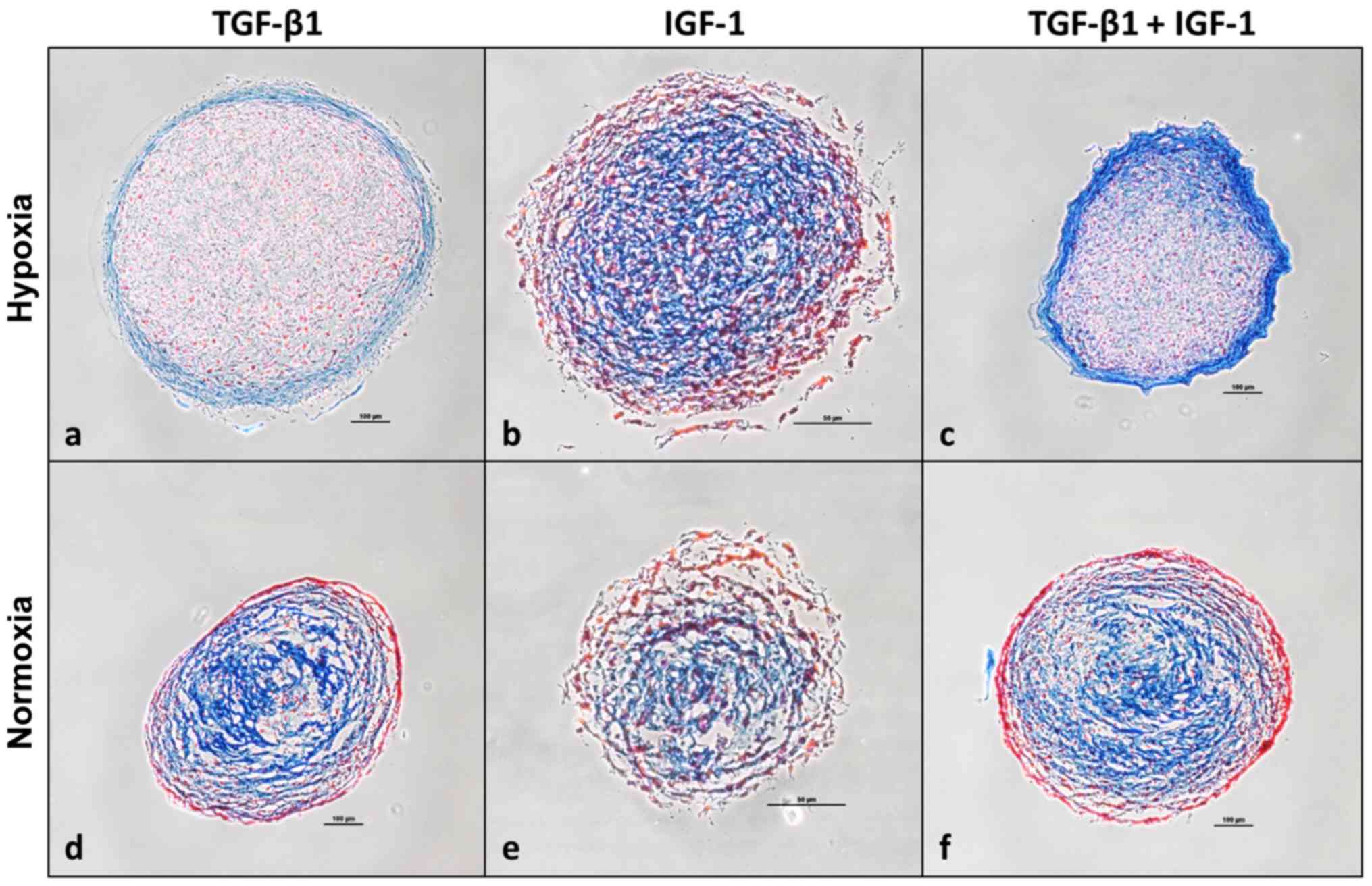
indicated in Fig. 7.
Additionally, an enhanced robustness of the pellet cultures against
homogenisation was observed over increasing culturing time. This
enhancement in robustness was more pronounced for pellets cultured
under hypoxia combined with TGF-β or TGF-β and IGF-1
stimulation.
Additionally, pellet cultures were histologically
analyzed after 35 days of incubation using Heidenhain's AZAN
trichrome staining indicating collagen-rich ECM in blue and nuclei
as well as cytoplasm in red. Pellets stimulated under hypoxia
(Fig. 7a–c) appeared more compact
and homogenous compared to those stimulated under normoxia
(Fig. 7d–f). Blue stained
marginal sides of pellets stimulated with TGF-β1 or the combination
of TGF-β1 and IGF-1 under hypoxia indicate the localization of
collagen-rich ECM as well as acid mucus substances (Fig. 7a and c). Referring to Fig. 7c, representing a section of a
pellet which was cultured under hypoxia and stimulated with the
combination of TGF-β1 and IGF-1, it is prominent that among all
stained sections its periphery is mostly deep-blue stained. This is
in contrast to IGF-1 stimulated pellet cultures from hypoxia
(Fig. 7b) and to the pellet
cultures cultured under normoxia (Fig. 7a–f). Here blue collagen-rich ECM
and acid mucus substances are rather located in the centre of the
section than in the periphery.
Discussion
Previous in vitro studies have already
examined growth factor receptor expression on TGF-β1- and/or
IGF-1-stimulated bovine (25,26) or leporine chondrocytes (27). To our knowledge, no studies have
been carried out to date dealing with donor-specific growth factor
responsiveness of human chondrocytes. Accordingly, our study
presents a new approach to investigate whether the targeted use of
growth factors deduced from specific receptor gene expression
profiles on the surface of human chondrocytes can positively
influence the redifferentiation potential of dedifferentiated
chondrocytic cells.
Gene expression of IGF-1 and TGF-β
receptors
In monolayer cultures, significant differences in
receptor mRNA levels in dependency to the applied growth factor
were determined.
These results indicate an upregulation of
IGF1R and TGFBR1 gene expression by the corresponding
growth factors IGF-1 and TGF-β1, respectively. Several in
vitro studies have previously demonstrated the capacity of
TGF-β1 to regulate the expression of its own receptors in rabbit
periosteal explants (27), human
hepatoma cell lines (28) or
human small cell lung cancer cell lines (29), respectively. Moreover, a positive
autoregulatory action was also demonstrated for IGF-1 on
IGF1R by Davies et al (26) in bovine chondrocytes and in
granulosa cells by deMoura et al (30). In contrast, TGFBR2 gene
expression was not upregulated by its corresponding growth factor
TGF-β1. As described by Barbara et al (31), TGF-β1 acts through an heterologous
complex composed of TGFBR1 and TGFBR2, suggesting
that both complex partners should be present in an adequate amount
to upregulate the total receptor complex. Nevertheless, we could
not demonstrate that the heterologous TGF-β-receptor complex
partners were simultaneously expressed at the mRNA level. The gene
expression of both receptors changed rather in dependency on growth
factor stimulation. It could be assumed that TGFBR2 was already
expressed at a higher level at the starting point, thus further
upregulation was not required. TGF-β1 may compensate the deficiency
of the limiting receptor subunit TGFBR1 by its upregulation
in order to improve sensitivity of chondrocytes towards TGF-β1
stimulation. The higher expression of TGFBR2 under IGF-1
stimulation could also be explained by gene regulation mechanisms.
Possible, due to a deficit of the corresponding ligand (here,
TGF-β1), the receptor is upregulated in chondrocytes in order to
increase sensitivity.
Moreover, it was apparent that the IGF1R
rather showed an increase in mRNA level over time whereas both
TGFBRs already reached maximum expression levels after 24 h
and remained almost constant over time. A possible explanation for
this behaviour could be a faster degradation time of TGFBRs
mRNA compared to IGF1R mRNA indicating mRNA accumulation
over time. As a consequence, the amount of transcribed and degraded
mRNA is in balance, leaving the TGFBRs mRNA level unchanged.
In the present study, we only analyzed growth factor receptor
expression at the mRNA level. In further studies, we must examine
how the receptor mRNA levels correlate with corresponding protein
levels.
However, our findings denote that the chondrogenic
growth factors could influence the gene expression profile of
IGF1R, TGFBR1 and TGFBR2. Moreover, the values
were partially widely scattered among the donors indicating the
existence of donor-specific responsiveness towards growth factor
stimulation.
Comparing the two different oxygen concentrations
(21 vs. 5%), we observed more pronounced gene expression profiles
under hypoxia implying better response ability under physiological
conditions. Furthermore, no significant differences in receptor
gene expression behaviour comparing monolayer and pellet culture
were determined. Therefore, pellet culturing for analysis of
receptor gene expression patterns seems to have no benefit,
suggesting monolayer culturing suitable for the establishment of an
analysis tool to deduce growth factor responsiveness.
Differentiation capacity of chondrocytic
cells
Although the ratios for the monolayer cultures
reached higher values under stimulation with IGF-1 compared to the
stimulation with TGF-β1 or the combination of TGF-β1 and IGF-1,
most of the cultures showed an increase in the ratios over time
under the latter stimulations. Assuming further increases in the
ratios under stimulation with TGF-β1 or the combination for
prolonged cultivation, it might be possible that the ratios even
exceed the ones under stimulation with IGF-1. This assumption could
be supported by the prominent increase in ratios in pellet cultures
after 14 and 35 days after stimulation with TGF-β1 and the
combination but not for IGF-1 alone, suggesting a greater impact of
TGF-β1 on the hyaline cartilage characteristic COL2A1 gene
expression.
Furthermore, after 35 days, higher
COL2A1/COL1A1 ratios were detected for the
combination of IGF-1 and TGF-β1 compared to the sole stimulation.
This was observed for both oxygen conditions, but was more
pronounced under hypoxia. This finding suggests an additive effect
of both growth factors, which was also observed in previous studies
(23) as well as by other
research groups (18,32,33). Additionally, it was previously
reported that intracellular signalling pathways of TGF-β1 and IGF-1
are partially linked (7,34). The connection of both pathways
could be a hint for an additive effect of both growth factors on
the chondrogenic differentiation. Nonetheless the signalling
pathways are not fully understood to date (7).
The detection of higher COL2A1/COL1A1
ratios in the pellet cultures in comparison to those noted in the
monolayer cultures suggests the dependency of three-dimensional
culturing as a chondrogenic stimulus itself. It is already known
that phenotypes and gene expression of chondrocytes are highly
dependent on the enclosing ECM and cell-to-cell contact (35). The close three-dimensional contact
of chondrocytes enables efficient autocrine stimulation and
enhances sythesis of ECM components (36). Our results support the previously
described findings. Nevertheless, varying
COL2A1/COL1A1 ratios among the donors, dependent on
the applied growth factors presumably denote donor-specific
redifferentiation potentials.
Comparing Col2 and sGAG synthesis rates of pellet
cultures, enhanced amounts of ECM components were detected under
hypoxia and stimulation with TGF-β1 or with the combination with
IGF-1, whereas they were low for the sole stimulation with IGF-1.
This finding also suggests an anabolic effect of TGF-β1 on the
synthesis of Col2 and sGAG and again a promotive effect of
physiological oxygen content. The results for the Col2 protein
expression were consistent with the mRNA expression levels of
COL2A1 after 14 and 35 days, confirming the correlation of
gene and protein expression. However, no additive effect of both
growth factors could be found on the protein level similar to the
case for the mRNA level. A delay in the translation from mRNA to
the protein could be a possible explanation for the absent additive
effect on the protein level.
The increased protein amounts of Col2 and sGAG in
pellet cultures after incubation with TGF-β1 or the combination of
TGF-β1 and IGF-1 were in relation to enhanced TGFBR1
expression of the different donors, supposing the possibility to
derive the responsiveness to TGF-β1 and/or IGF-1 from the increase
in mRNA levels of the corresponding receptors.
Altogether, the detection of higher
COL2A1/COL1A1-ratios as well as higher Col2 and sGAG
amounts after TGF-β1 stimulation alone or in combination with IGF-1
under hypoxia confirm the positive impact of physiologically
approximated culture conditions on the redifferentiation potential.
Moreover, gene expression of ALP and COL10A1 was not
induced in pellet cultures, giving no evidence for the emergence of
hyper-trophic processes and the development of fibrocartilage.
Furthermore, we examined
COL2A1/COL1A1-ratios in unstimulated cultures, using
only ITS-supplement in the cell culture media. These control
cultures showed almost similar results as the cultures treated with
IGF-1. We suppose that these effects may be explained by the
supplemented insulin in the ITS-reagent. Because of molecular
similarities between insulin and IGF-1, both interact on same
receptor binding sites (37).
Therefore, insulin probably simulated the effects of IGF-1 by the
upregulation of same pathways.
Morphological and histological
characterization of pellet cultures
The variation of pellet size and metabolic activity
in relation to different growth factor stimulation and different
oxygen conditions confirm the dependency of pellet morphology on
defined culturing conditions. An enhancement in pellet size under
stimulation with TGF-β1 or the combination of TGF-β1 and IGF-1
suggests an enhancing effect of the applied growth factors either
on proliferation or ECM production of human chondrocytes. The same
can be estimated for the culturing under hypoxia due to an increase
in pellet size. The reduction in pellet size under normoxia could
probably be explained by an intensification of oxidative stress,
driving cells towards apoptosis (38,39). Bohensky et al (40) previously showed that a reduced
oxygen concentration prevents apoptosis of chondrocytes due to the
expression of reactive oxygen species protective hypoxic-inducible
transcription factors. Babur et al (41) compared human chondrocytic
macropellets and micropellets under different oxygen conditions and
likewise observed larger volumes as well as greater metabolic
activity, sGAG production and higher Col2 expression of pellets
cultivated under hypoxia, indicating a redifferentiation
process.
Observed enhanced robustness under hypoxia combined
with TGF-β or TGF-β and IGF-1 stimulation may indicate that pellets
gained robustness due to the production of hyaline-like ECM. This
assumption would resemble the detected anabolic effect of TGF-β1 on
the Col2 and sGAG expression.
Analysis of histological sections of the pellet
cultures after 35 days indicated that pellets cultured under
hypoxia appeared more compact. This finding was consistent with the
morphological observations and enhanced levels of hyaline-like ECM
components Col2 and sGAG. For pellets stimulated with TGF-β1 or the
combination of TGF-β1 and IGF-1 under hypoxia, collagen-rich ECM as
well as acid mucus substances were rather present at the marginal
sides indicating a capsule-like formation of ECM. This capsule
formation could also explain the higher resistance of these pellets
to homogenization. Although the staining did not distinguish
between fibrous-cartilaginous typical Col1 and hyaline
characteristic Col2, the enhancement in robustness may give a hint
for the production of more mechanically resistant Col2.
In contrast, in IGF-1-stimulated pellets under
hypoxia and in all pellet cultures under normoxia, the ECM was
rather located in the centre of the histological sections.
One could speculate that the capsule-like formation
could lead to a higher concentration of autocrine secreted growth
factors (and other substances) within the pellet itself due to a
reduced diffusion out of the capsule. This enhanced autocrine
stimulation could promote the redifferentiation of the chondrocytes
within the pellet construct. Therefore, the capsule-like formation
might be more favourable than the central location of ECM in the
pellets. On the other hand, less diffusion of nutrients into the
centre of the pellets may impair cell viability. Thus, it remains
unclear which ECM formation of the pellets is more beneficial to
generate functional in vivo-like cartilage. This should be
clarified in further studies.
In conclusion, we found characteristic gene
expression profiles of TGFBRs and IGF1R according to
the applied growth factors. These gene expression profiles were
even more pronounced under hypoxia. Our findings support the
assumption that the chondrogenic growth factors influence the gene
expression of their corresponding receptors IGF1R,
TGFBR1 and TGFBR2. Moreover, increased amounts of
Col2 protein and sGAG after incubation with TGF-β1 and/or IGF-1
were validated in pellet cultures. Further histological
investigations also indicated that reduced oxygen conditions
combined with the use of chondrogenic growth factors supported the
generation of hyaline-like tissue. Nevertheless, further
experiments with higher sample sizes and prolonged culturing time
must be conducted to prove the applicability of a screening method
on growth factor receptors. Moreover, the further examination of
intracellular signalling pathways that are related to the
redifferentiation process of human chondrocytes can be helpful to
analyze possible interactions of growth factors at the molecular
level. In further research, it is recommended to investigate
protein levels of specific growth factor receptors either by flow
cytometry or western blot analysis in order to examine the
correlation of mRNA and protein levels. It may also be enlightening
to sort TGF-β1 and/or IGF-1 receptor-positive cells and determine
their ability for redifferentiation.
Nonetheless, constructing a gene expression profile
regarding mRNA levels of specific growth factor receptors in
monolayer cultures is a valuable tool to predict the individual
response of donors in growth factor application in vitro.
The results of the present study could be a basis for further
research on donor-specific cartilage regeneration to make the
therapeutic attempt of chondrogenic redifferentiation more
efficient in ex vivo cell expansion procedures.
Acknowledgments
The authors would like to thank Mrs. Frauke Winzer,
Institute for Anatomy, University Medical Centre Rostock (Rostock,
Germany) for supporting the sample preparation for the histological
staining.
References
|
1
|
Loeser RF Jr: Aging cartilage and
osteoarthritis - What's the link? Sci Aging Knowl Env.
2004:pe312004. View Article : Google Scholar
|
|
2
|
Chiang H and Jiang C-C: Repair of
articular cartilage defects: Review and perspectives. J Formos Med
Assoc. 108:87–101. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shi Y, Ma J, Zhang X, Li H, Jiang L and
Qin J: Hypoxia combined with spheroid culture improves cartilage
specific function in chondrocytes. Integr Biol. 7:289–297. 2015.
View Article : Google Scholar
|
|
4
|
Tekari A, Luginbuehl R, Hofstetter W and
Egli RJ: Transforming growth factor beta signaling is essential for
the autonomous formation of cartilage-like tissue by expanded
chondrocytes. PLoS One. 10:e01208572015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Arzi B, DuRaine GD, Lee CA, Huey DJ,
Borjesson DL, Murphy BG, Hu JC, Baumgarth N and Athanasiou KA:
Cartilage immunoprivilege depends on donor source and lesion
location. Acta Biomater. 23:72–81. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schulze-Tanzil G: Activation and
dedifferentiation of chondrocytes: Implications in cartilage injury
and repair. Ann Anat. 91:325–338. 2009. View Article : Google Scholar
|
|
7
|
Duan L, Ma B, Liang Y, Chen J, Zhu W, Li M
and Wang D: Cytokine networking of chondrocyte dedifferentiation in
vitro and its implications for cell-based cartilage therapy. Am J
Transl Res. 7:194–208. 2015.PubMed/NCBI
|
|
8
|
Bhardwaj N, Devi D and Mandal BB:
Tissue-engineered cartilage: The crossroads of biomaterials, cells
and stimulating factors. Macromol Biosci. 15:153–182. 2015.
View Article : Google Scholar
|
|
9
|
Fickert S, Gerwien P, Helmert B,
Schattenberg T, Weckbach S, Kaszkin-Bettag M and Lehmann L:
One-year clinical and radiological results of a prospective,
investigator-initiated trial examining a novel, purely autologous
3-dimensional autologous chondrocyte transplantation product in the
knee. Cartilage. 3:27–42. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
van der Kraan PM, Buma P, van Kuppevelt T
and van den Berg WB: Interaction of chondrocytes, extracellular
matrix and growth factors: Relevance for articular cartilage tissue
engineering. Osteoarthritis Cartilage. 10:631–637. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
van Osch GJ, van den Berg WB, Hunziker EB
and Häuselmann HJ: Differential effects of IGF-1 and TGF beta-2 on
the assembly of proteoglycans in pericellular and territorial
matrix by cultured bovine articular chondrocytes. Osteoarthritis
Cartilage. 6:187–195. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Giovannini S, Diaz-Romero J, Aigner T,
Mainil-Varlet P and Nesic D: Population doublings and percentage of
S100-positive cells as predictors of in vitro chondrogenicity of
expanded human articular chondrocytes. J Cell Physiol. 222:411–420.
2010. View Article : Google Scholar
|
|
13
|
Chubinskaya S, Hakimiyan A, Pacione C,
Yanke A, Rappoport L, Aigner T, Rueger DC and Loeser RF:
Synergistic effect of IGF-1 and OP-1 on matrix formation by normal
and OA chondrocytes cultured in alginate beads. Osteoarthritis
Cartilage. 15:421–430. 2007. View Article : Google Scholar
|
|
14
|
McQuillan DJ, Handley CJ, Campbell MA,
Bolis S, Milway VE and Herington AC: Stimulation of proteoglycan
biosynthesis by serum and insulin-like growth factor-I in cultured
bovine articular cartilage. Biochem J. 240:423–430. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guerne PA, Sublet A and Lotz M: Growth
factor responsiveness of human articular chondrocytes: Distinct
profiles in primary chondrocytes, subcultured chondrocytes, and
fibroblasts. J Cell Physiol. 158:476–484. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jones JI and Clemmons DR: Insulin-like
growth factors and their binding proteins: Biological actions.
Endocr Rev. 16:3–34. 1995.PubMed/NCBI
|
|
17
|
Giannoni P and Cancedda R: Articular
chondrocyte culturing for cell-based cartilage repair: Needs and
perspectives. Cells Tissues Organs. 184:1–15. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fukumoto T, Sperling JW, Sanyal A,
Fitzsimmons JS, Reinholz GG, Conover CA and O'Driscoll SW: Combined
effects of insulin-like growth factor-1 and transforming growth
factor-beta1 on periosteal mesenchymal cells during chondrogenesis
in vitro. Osteoarthritis Cartilage. 11:55–64. 2003. View Article : Google Scholar
|
|
19
|
Redini F, Galera P, Mauviel A, Loyau G and
Pujol JP: Transforming growth factor beta stimulates collagen and
glycosaminoglycan biosynthesis in cultured rabbit articular
chondrocytes. FEBS Lett. 234:172–176. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shi Y and Massagué J: Mechanisms of
TGF-beta signaling from cell membrane to the nucleus. Cell.
113:685–700. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bassing CH, Howe DJ, Segarini PR, Donahoe
PK and Wang XF: A single heteromeric receptor complex is sufficient
to mediate biological effects of transforming growth factor-beta
ligands. J Biol Chem. 269:14861–14864. 1994.PubMed/NCBI
|
|
22
|
Jonitz A, Lochner K, Peters K, Salamon A,
Pasold J, Mueller-Hilke B, Hansmann D and Bader R: Differentiation
capacity of human chondrocytes embedded in alginate matrix. Connect
Tissue Res. 52:503–511. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jonitz A, Lochner K, Tischer T, Hansmann D
and Bader R: TGF-β1 and IGF-1 influence the redifferentiation
capacity of human chondrocytes in 3D pellet cultures in relation to
different oxygen concentrations. Int J Mol Med. 30:666–672.
2012.PubMed/NCBI
|
|
24
|
Pasold J, Zander K, Heskamp B, Grüttner C,
Lüthen F, Tischer T, Jonitz-Heincke A and Bader R: Positive impact
of IGF-1-coupled nanoparticles on the differentiation potential of
human chondrocytes cultured on collagen scaffolds. Int J
Nanomedicine. 10:1131–1143. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang Y-H and Barabino GA: Differential
morphology and homogeneity of tissue-engineered cartilage in
hydrodynamic cultivation with transient exposure to insulin-like
growth factor-1 and transforming growth factor-β1. Tissue Eng Part
A. 19:2349–2360. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Davies LC, Blain EJ, Gilbert SJ, Caterson
B and Duance VC: The potential of IGF-1 and TGFbeta1 for promoting
'adult' articular cartilage repair: An in vitro study. Tissue Eng
Part A. 14:1251–1261. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mizuta H, Sanyal A, Fukumoto T,
Fitzsimmons JS, Matsui N, Bolander ME, Oursler MJ and O'Driscoll
SW: The spatiotemporal expression of TGF-beta1 and its receptors
during periosteal chondrogenesis in vitro. J Orthop Res.
20:562–574. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Inagaki M, Wang Z and Carr BI:
Transforming growth factor beta 1 selectively increases gene
expression of the serine/threonine kinase receptor 1 (SKR1) in
human hepatoma cell lines. Cell Struct Funct. 19:305–313. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nørgaard P, Spang-Thomsen M and Poulsen
HS: Expression and autoregulation of transforming growth factor
beta receptor mRNA in small-cell lung cancer cell lines. Br J
Cancer. 73:1037–1043. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
deMoura MD, Chamoun D, Resnick CE and
Adashi EY: Insulin-like growth factor (IGF)-I stimulates IGF-I and
type 1 IGF receptor expression in cultured rat granulosa cells:
Autocrine regulation of the intrafollicular IGF-I system.
Endocrine. 13:103–110. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Barbara NP, Wrana JL and Letarte M:
Endoglin is an accessory protein that interacts with the signaling
receptor complex of multiple members of the transforming growth
factor-β super-family. J Biol Chem. 274:584–594. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Longobardi L, O'Rear L, Aakula S,
Johnstone B, Shimer K, Chytil A, Horton WA, Moses HL and Spagnoli
A: Effect of IGF-I in the chondrogenesis of bone marrow mesenchymal
stem cells in the presence or absence of TGF-beta signaling. J Bone
Miner Res. 21:626–636. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yaeger PC, Masi TL, de Ortiz JL, Binette
F, Tubo R and McPherson JM: Synergistic action of transforming
growth factor-beta and insulin-like growth factor-I induces
expression of type II collagen and aggrecan genes in adult human
articular chondrocytes. Exp Cell Res. 237:318–325. 1997. View Article : Google Scholar
|
|
34
|
Shakibaei M, John T and Rahmanzadeh R:
Collaboration with the insulin-like growth factor-I receptor.
Biochem J. 342:615–623
|
|
35
|
Landry J, Bernier D, Ouellet C, Goyette R
and Marceau N: Spheroidal aggregate culture of rat liver cells:
Histotypic reorganization, biomatrix deposition, and maintenance of
functional activities. J Cell Biol. 101:914–923. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Anderer U and Libera J: In vitro
engineering of human autogenous cartilage. J Bone Miner Res.
17:1420–1429. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Czech MP: Signal transmission by the
insulin-like growth factors. Cell. 59:235–238. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Iwasa K, Hayashi S, Fujishiro T, Kanzaki
N, Hashimoto S, Sakata S, Chinzei N, Nishiyama T, Kuroda R and
Kurosaka M: PTEN regulates matrix synthesis in adult human
chondrocytes under oxidative stress. J Orthop Res. 32:231–237.
2014. View Article : Google Scholar
|
|
39
|
Sakata S, Hayashi S, Fujishiro T, Kawakita
K, Kanzaki N, Hashimoto S, Iwasa K, Chinzei N, Kihara S, Haneda M,
et al: Oxidative stress-induced apoptosis and matrix loss of
chondrocytes is inhibited by eicosapentaenoic acid. J Orthop Res.
33:359–365. 2015. View Article : Google Scholar
|
|
40
|
Bohensky J, Terkhorn SP, Freeman TA, Adams
CS, Garcia JA, Shapiro IM and Srinivas V: Regulation of autophagy
in cartilage: HIF-2 suppresses chondrocyte autophagy. Arthritis
Rheum. 60:1406–1415. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Babur BK, Ghanavi P, Levett P, Lott WB,
Klein T, Cooper-White JJ, Crawford R and Doran MR: The interplay
between chondrocyte redifferentiation pellet size and oxygen
concentration. PLoS One. 8:e588652013. View Article : Google Scholar : PubMed/NCBI
|