Introduction
Myocardial edema occurs in various myocardial
pathologies, particularly in ischemia and reperfusion, and
contributes to cell dysfunction and death (1). Edema is one of the consequences of a
short period myocardial ischemia, which is reversible in the
initial stage (2). Following
myocardial reperfusion, due to abnormal fluid accumulation in
cardiomyocytes and interstitial compartments, there is an marked
edematous reaction in the myocardium, causing myocardial
dysfunction and cell death (3,4).
Myocardial edema is divided into cellular edema and interstitial
edema (5,6), and its measurement is primarily
based on the noninvasive evaluation of cardiac magnetic resonance
imaging (7) and tissue water
content (8). A number of studies
have focused on global myocardial edema during ischemia-reperfusion
(I/R). However, few studies have described the dynamic change in
terms of cardiomyocyte edema.
It is well established that water movement across
cell membranes is passive and is determined by osmotic gradients
and membrane permeability to water. During systolic depolarization,
Na+ enters the cardiomyocyte due to the large voltage
and concentration gradients, and repolarization is mainly achieved
via K+ efflux. Furthermore, the intracellular
Na+ concentration is dispersed by the effects of
Na+-K+-ATPase (an Na+ pump). Ionic
homeostasis is an energy consuming process and any energy
deficiency results in Na+ overload and cell edema
(9,10). Furthermore, Na+
concentration is also necessary for intracellular pH regulation.
H+ efflux is largely dependent on Na+ influx
via the Na+-H+ exchanger (NHE) and
Na+-HCO3- transporter. Therefore, the
accumulation of intracellular H+ also causes
Na+ overload and induces changes in cell volume
(11). During myocardial
ischemia, due to energy insufficiency and glycolytic activation
(lactate accumulation leads to a decrease in pH), inactivation of
the Na+ pump and activation of the NHE results in
Na+ influx and leads to cardiomyocyte edema.
Aquaporins (AQPs) are a large membrane protein
family, which usually oligomerize into tetramers, forming pores.
These pores are permeable to water and serve a vital role in
osmoregulation and water transport. AQPs have also been identified
in the human cardiomyocyte membrane (12). The protein and mRNA expression
levels of sarcolemmal AQP-4 in cardiomyocytes are enhanced during
I/R, and this increase has been connected to enlarged infarct sizes
(13). A decrease in the
expression of AQP-4 in mouse brains downregulates the cell edema
and necrosis induced by ischemia (14). Furthermore, the expression of
AQP-1 has been identified in rat cardiomyocytes and appears to be
associated with water movement (15). These results reveal that AQP may
have a negative effect on I/R injury via regulating water transport
and cardiomyocyte volume.
Pigment epithelium derived factor (PEDF) is an
endogenous multifunctional protein which is commonly expressed in
normal tissues (16), and serves
protective roles in cardiomyocytes during oxygen-glucose
deprivation (OGD) (17) and
recovery (18). Our recent study
suggested that PEDF protects cardiomyocytes undergoing OGD through
reducing glycolytic activation and the level of ATP (19). These results motivated the
hypothesis that PEDF may decrease cardiomyocyte edema during OGD by
reducing lactate accumulation. However, the low level of ATP
induced by PEDF may have a negative effect on cardiomyocyte edema
during OGD. Furthermore, the association between PEDF and AQP in
cardiomyocytes during OGD/recovery (I/R) remains to be fully
elucidated. Therefore, the aims of the present study were to
investigate the effect on PEDF on cardiomyocyte edema during I/R
and to elaborate the underlying mechanisms of PEDF in the
regulation of cardiomyocyte edema.
Materials and methods
Reagents
Cleaved caspase-3 antibody (cat. no. 9664, 1:1,000)
was purchased from Cell Signaling Technology, Inc. (Danvers, MA,
USA). Antibodies targeting protein kinase receptor-interacting
protein 3 (RIP3, cat. no. 17563-1-AP, 1:1,000), AQP1 (cat. no.
20333-1-AP, 1:1,000), AQP4 (cat. no. 16473-1-AP, 1:1,000) and
β-tubulin (cat. no. 66240-1-lg, 1:5,000) were purchased from
ProteinTech Group, Inc. (Rosemont, IL, USA). Antibody targeting
α-sarcomeric actin (α-SCA, cat. no. SAB4200689, 1:500) was
purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). ATP
disodium salt hydrate (cat. no. A1852), Na+-binding
benzofuran isophthalate (SBFI, cat. no. 129423-53-6), the Lactate
Assay kit II (cat. no. MAK065) and bromodeoxy-uridine (BrdU; cat.
no. B5002, 0.1 mM) were purchased from Sigma-Aldrich; Merck KGaA).
Calcein AM (UltraPure grade, AAT bioquest 22003) was obtained from
Shanghai Yubo Biological Technology Co., Ltd. (Shanghai, China).
Fructose-1, 6-diphosphate (FDP, 20 mM, cat. no. 81028-91-3) was
obtained from Shanghai Guangrui Biological Technology Co., Ltd.
(Shanghai, China). Arginine vasopressin (AVP; 0.1 nM; cat. no.
113-79-1) was purchased from Hangzhou Jinqi Biotechnology Co., Ltd.
(Hangzhou, China). The Sodium Potassium ATPase Activity kit (cat.
no. QS1700) was obtained from Shanghai Cablebridge Biotechnology
Co., Ltd. (Shanghai, China).
PEDF protein preparation
Recombinant rat PEDF (GenBank™ accession no.
NM_177927) was synthesized by Cusabio Biotech, Co., Ltd. (Wuhan,
China) as described previously (20). Our previous studies demonstrated
that 10 nmol/l is the optimal PEDF concentration for cardiomyocyte
protection (21).
Lentivirus (LVs) and plasmid
preparation
Recombinant lentivirus was prepared as described
previously (22). The PEDF
overexpression plasmid was constructed and packaged in 293T cells
(American Type Culture Collection, Manassas, VA, USA), and the
concentrated titer of the viral suspension was 2×1012
TU/l.
Animals experiments
Adult male Sprague-Dawley (SD) rats (8-12 weeks old,
200-250 g) were supplied by the Experimental Animal Center of
Xuzhou Medical University (Xuzhou, China) and housed in a
controlled environment (humidity, 50-60%). A total of 3 rats were
housed per cage and were maintained at room temperature under a 12
h light/dark cycle; rats were provided free access to food and
water. The animal experiments performed in the present study
conformed to the Guide for the Care and Use of Laboratory Animals
published by the National Institutes of Health (Publication, 8th
Edition, 2011, Bethesda, MD, USA) (23). The animal procedures were approved
by the Xuzhou Medical University Committee on Animal Care. The I/R
models were established as described previously (18). Intramyocardial gene delivery was
performed 1 week prior to the I/R experiment in the rats. PEDF-LV
(2×107 TU) with enhanced infection solution (Shanghai
GeneChem Co., Ltd., Shanghai, China) was delivered into the
infracted area of the myocardium. Sham-operated animals underwent
an identical surgical procedure without left-anterior descending
coronary artery ligation. For the rat I/R model, following 30 min
of ischemic treatment, reperfusion was allowed for 2 h by releasing
the ligatures. The animal models were randomly divided into eight
groups as follows: i) Sham group, surgical procedure without artery
ligation (n=10); ii) Sham + PEDF-LV group, surgical procedure
without artery ligation, PEDF-LV was transferred prior to surgery
(n=10); iii) I group, 0.5 h ischemia (n=10); iv) ischemia + PEDF-LV
group, 0.5 h ischemia, PEDF-LV was transferred prior to surgery
(n=10); v) ischemia + vehicle group, 0.5 h ischemia, vehicle was
transferred prior to surgery (n=10); vi) I/R group, 0.5 h ischemia
and 2 h reperfusion (n=10); vii) I/R + PEDF-LV group, 0.5 h
ischemia and 2 h reperfusion, PEDF-LV was transferred prior to
surgery (n=10); viii) I/R + vehicle group, 0.5 h ischemia and 2 h
reperfusion, vehicle was transferred prior to surgery (n=10).
Rat neonatal cardiomyocyte isolation and
culture
Neonatal rat cardiomyocytes were isolated from
neonatal male (1-3 days, 5-7 g, n=700) SD rats as previously
described (24); rats were
purchased from the Experimental Animal Centre of Xuzhou Medical
University. The neonatal male SD rats were housed in the
Experimental Animal Centre of Xuzhou Medical University and housed
in a controlled environment (temperature, 20-25°C; humidity,
50-60%). The neonatal rats and their mother were housed one cage
and were maintained at room temperature under a 12 h light/dark
cycle. The neonatal rats were provided breast milk and the mother
rat was provided free access to sterile food and water. The
neonatal rats were anesthetized with sodium pentobarbital and
sacrificed by decapitation. Their hearts were rapidly removed into
dishes of ice, and the vessels and atria were discarded. The
ventricles were then dissected and minced into 1 mm3
pieces, and washed in PBS. The minced tissue was digested in a PBS
with 1 mg/ml trypsin, 1 mg/ml collagenase type II, and 0.2 mg/ml
glucose for 5 min at 37°C and incubated with 0.1 mmol/l BrdU to
selectively enrich for cardiomyocytes by inhibiting the growth of
cardiac fibroblasts. The cardiomyocytes were then purified using
the differential adhesion method. The cardiomyocytes were evaluated
by indirect immunofluorescence staining with α-SCA antibody. The
isolated cardiomyocytes were cultured in Dulbecco's modified
Eagle's medium (Gibco; Thermo Fisher Scientific, Inc.) with 4.5 g/l
glucose and 10% fetal bovine serum at 37°C. OGD was completed by
culturing the cardiomyocytes in a glucose-free DMEM (Gibco; Thermo
Fisher Scientific, Inc.) without FBS and a tri-gas incubator (Heal
Force, Shanghai, China) with 1% O2/5% CO2/94%
N2 at 37°C. OGD/R was performed as previously described
(25). Following the OGD
procedure, the medium was replaced with DMEM with 4.5 g/l glucose,
and then transferred into a humidified normoxic atmosphere at 37°C.
The cardiomyocytes were treated with or without 10 nM PEDF 1 h
prior to ischemia or reperfusion. The following experimental groups
were included: Normal group, OGD group (control, OGD 0.5 h), OGD +
PEDF group, OGD/R group (control, OGD 0.5 h and recovery 6 h),
OGD/R + PEDF group.
Cell volume measurements
Cell volumes were measured by confocal laser
scanning microscopy (Olympus Corporation, Tokyo, Japan). Firstly,
the cardiomyocytes were stained with calcein AM (2 μM) to
stain the cellular cytosol. The cells were then cultured with
pre-hypoxic glucose deprivation medium, maintained in an incubator
containing 1% O2/5% CO2/94% N2 at
37°C for 15 min. Mineral oil was added to the upper layer of the
medium to prevent oxygen in the air from contacting the medium, to
ensure that the cardiomyocytes were always in an anoxic state
(26). Each culture plate with
stained cardiomyocytes was placed on an inverted microscope.
Furthermore, a single rod-shaped viable cardiomyocyte from each
plate was selected and then scanned using an Argon beam (488 nm).
Using ImagePro Plus 6.0 software (Media Cybernetics, Inc.,
Rockville, MD, USA) to calculate the area of each layer of the
cell, the sum of the area of each layer of the cell multiplied by a
plane distance of 3 μm was equal to the capacity of the
cell.
Cell viability assessment
The rat primary cardiomyocytes (1×104 per
well) were seeded into wells of a 96-well plate (Corning, New York,
USA). The cell viability was assessed using a CCK-8 kit (Dojindo
Molecular Technologies, Inc., Tokyo, Japan). The absorbance at 450
nm was measured using a microplate reader (BioTek Synergy2,
Instruments, Inc., Winooski, VT, USA). The mean optical density
(OD) value from at least three parallel groups was used to
calculate the cell viability.
Protein extraction
The rat heart tissue proteins were extracted from
the left ventricular myocardium using a lysis buffer (100 mmol/l
Tris-HCl, 4% SDS, 20% glycerine, 200 mmol/l DTT, phosphatase and
protease inhibitors, pH 6.8). The cardiomyocytes were lysed using
the Cell Total Protein Extraction kit (Sangon Biotech, Shanghai,
China) for whole cell lysates. The proteins were extracted using
the Nuclear and Cytoplasmic Protein Extraction kit (Beyotime
Institute of Biotechnology, Shanghai, China) for nuclear and
cytoplasmic lysates according to the manufacturer's protocol. The
protein concentrations were measured using a bicinchoninic acid
assay.
Western blotting
For western blotting analysis, the cells were
solubilized in lysis buffer (100 mmol/l Tris-Hcl, 4% SDS, 20%
glycerine, 200 mmol/l DTT, phosphatase and protease inhibitors, pH
6.8). Protein concentration from the supernatant was determined
using a BCA protein assay kit (Pierce; Thermo Fisher Scientific,
Inc.). Total cellular protein was denatured by boiling for 10 min
with an equal volume of 2X Tris-glycine SDS buffer. A total of 50
ng protein per lane was separated by 12% SDS-PAGE and transferred
to nitrocellulose membranes (EMD Millipore, Billerica, MA, USA).
Following blocking with 5% non-fat milk/PBS-T for 3 h at room
temperature, the membranes were incubated with primary antibodies
overnight at 4°C. Then, fluorescence-labeled secondary antibodies
was added for 2 h at room temperature. Cleaved caspase-3 (1:1,000),
RIP3 (1:1,000), AQP1(1:1,000), AQP4 (1:1,000) and β-tubulin
(1:5,000) primary antibodies were used, followed by relative
secondary antibodies (anti-rabbit IgG H+L DyLight™ 800 4X PEG
1:30,000; cat. no. 5151, and anti-mouse IgG H+L DyLight™ 680
1:15,000; cat. no. 5470, Cell Signaling Technology, Inc.) and
images were captured using the Odyssey infrared imaging system
(LI-COR Biosciences, Lincoln, NE, USA). Western blots were
quantified using ImageJ software (v1.50; National Institutes of
Health). The levels of protein were calculated by the ratio of
relative protein/β-tubulin.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
The total cardiomyocyte RNA was extracted using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
following the manufacturer's protocol. The RNA (1,000 nmol) was
then subjected to reverse transcription with the Prime Script RT
reagent kit and gDNA Eraser (Takara Biotechnology Co., Ltd.,
Dalian, China). PCR was conducted with a final volume of 20
μl containing 10 μl 2X SYBR-Green PCR Master mix, 0.1
μM of each primer and 100 μg genomic DNA. The mixture
was subjected to qPCR amplification (95°C for 10 min), 45 cycles
(95°C for 10 sec, 60°C for 10 sec, 72°C for 20 sec), and one cycle
(95°C for 1 min, 65°C for 1 min and 97°C with continuous) and then
cooled to 40°C for 30 sec using a Roche Light Cycler 480 (Roche
Diagnostics GmbH, Mannheim, Germany). Gene expression was
normalized to that of 18s RNA. Gene expression was quantified using
the 2-∆∆Cq method (27). The following primers, synthesized
by GenScript (Piscataway, NJ, USA), were used: APQ1 forward,
5'-ACCTGCTGGCC ATTGACTAC-3' and reverse, 5'-CCA GGG CAC TCC CAA TGA
AT-3'; APQ4 forward, 5'-CCC CAG AAG ACA GCA CCT G-3' and reverse,
5'-GAG GGA GGT CCA CAC TTA CAG C'; 18sRNA forward, 5'-CCT GGA TAC
CGC AGC TAG GA-3' and reverse, 5'-GCG GCG CAA TAC GAA TGC CCC-3'.
The levels of mRNA were quantified through 18S rRNA to normalize
AQP-1 and AQP-4.
High performance liquid chromatography
(HPLC) analysis
ATP levels was measured using an HPLC system
(SIL-20A, Shimadzu Corporation, Kyoto, Japan) with detection at 205
nm. Mobile phase A: 10 mmol/l KH2PO4, 2.5%
acetonitrile, 5.8 mmol/l tetrabutylammonium hydroxide, pH 6.0;
Mobile phase B: 10 mmol/l KH2PO4, 25%
acetonitrile, 5.8 mmol/l tetra-butylammonium hydroxide, pH 5.5;
Reversed-phase C18 5 μm column (250×4. 6 mm) was used. The
division of the peak area of samples by that of standards
represented the ATP concentrations.
Extracellular acidification rate (ECAR)
measurement
The ECAR was measured using a Seahorse XF96
extracellular flux analyzer (Seahorse Bioscience, North Billerica,
MA, USA) which reflects the activity of glycolysis. The
cardiomyocytes were seeded in 96-well assay plates at
8×103 cells/well and incubated at 37°C prior to
analysis. ECAR was recorded immediately following the completion of
OGD for the indicated time periods. The unit for ECAR was
mpH/min.
Morphological assessment
The left ventricle was fixed in 10% paraformaldehyde
and embedded in paraffin. Coronal sections (4-μm thick) of
the left ventricle from the middle section were stained to
determine the cross-sectional area (CSA) of cardiomyocytes as
described previously (28,29).
A total of 50 cross-sections of cardiac muscle fibers were counted
in at least 10 images of the left ventricle to assess the CSA. Each
cell was traced individually, and its CSA was measured directly.
The CSA measurements were performed on 10 rats, with 500 nuclei
counted using ImagePro Plus software. Ultrathin sections were
obtained using an EM UC7 (Leica Microsystems GmbH, Wetzlar,
Germany). Mitochondrial CSA was determined using a Tecnai G2 T12
(FEI; Thermo Fisher Scientific, Inc.) transmission electron
microscope to quantify the mitochondrial edema. For comparison of
mitochondrial edema, electron microscope micrographs of thin
sections were evaluated. In each of the 10 images of the left
ventricle, ~50 mitochondria were confirmed.
Measurement of SBFI AM, lactic acid and
NA+-K+-ATPase activity
The concentrations of SBFI AM, lactic acid and
NA+-K+-ATPase activity were measured using
the respective detection kits, according to the manufacturers'
protocols.
Statistical analysis
Data in the present study are expressed as the mean
± standard error of the mean. All statistical analyses were
conducted using a SPSS 19.0 software (IBM Corp., Armonk, NY, USA).
Data between two groups were compared using Student's
t-test, and multiple comparisons used one-way analysis of
variance and Student-Newman-Keuls test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Cell edema increases in cardiomyocytes
during OGD and OGD/R
In order to examine the effects of ischemia and
reperfusion on cardiomyocyte swelling, primary neonatal
cardiomyocytes exposed to OGD for 30 min were isolated to mimic
in vivo ischemia, and incubated in normal media with glucose
and fetal bovine serum under normoxia to mimic in vivo
recovery, thus simulating in vivo reperfusion. The volume of
cardiomyocytes was measured by confocal tomography (Fig. 1A). The results indicated that,
following OGD for 30 min, the diameter of the cardiomyocytes
increased from 8 to 10 layers, and after 6 h, the number of layers
increased further to 14. The cell volume at each time point
post-OGD/R was calculated by the aforementioned formula. The
cardiomyocytes were significantly swollen within 15 min of OGD and
continued to swell for 30 min, following which the volume tended to
stabilize within 2 h. The maximum cell volume during the whole
OGD/R process was ~1.4-fold higher than that of the normal cells
(Fig. 1B). Within 30 min of
recovery, the cell volume increased significantly and peaked at 6
h; the volume of these cells was ~1.75-fold higher than that of
normal cardiomyocytes (Fig. 1C).
The cell volumes stabilized within 12 h.
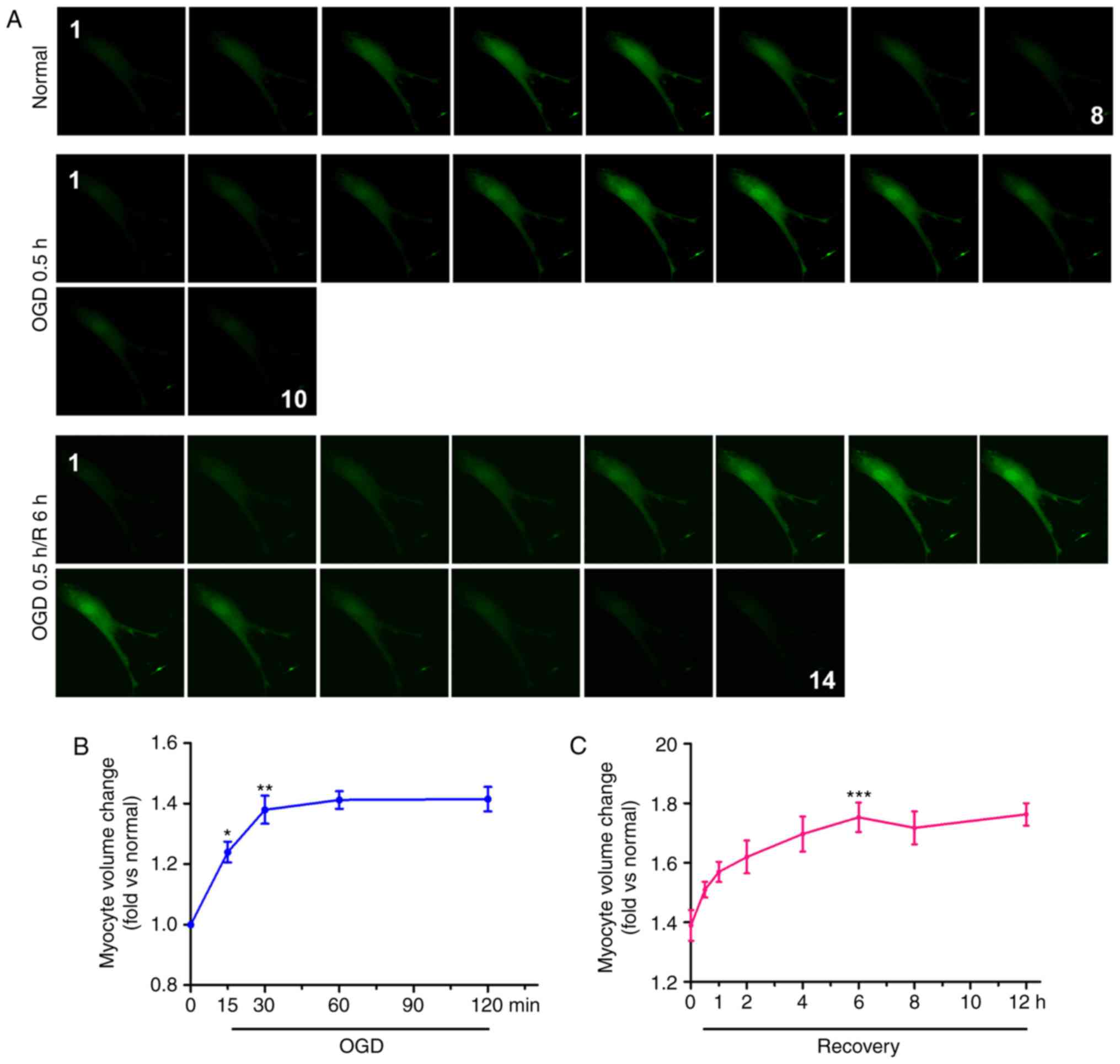 | Figure 1Cell edema increases in
cardiomyocytes during OGD and OGD/R. (A) Calcein-AM was used to
stain neonatal cardiomyocytes, and cell volume was measured with a
confocal microscope by stack scanning (original magnification,
×400). The time courses of cell volume changes in neonatal
cardiomyocytes following (B) OGD and (C) 30 min of OGD/R (n=6) are
shown. *P<0.05 and **P<0.01 and
***P<0.001, vs. relative normal group. (D)
Hematoxylin and eosin-stained micrographs of myocardial sections
from the left ventricle (original magnification, ×400). The graph
shows CSA values of the Sham, I 0.5 h and I 0.5 h/R 2 h groups; the
results are presented as the mean CSA of 50 random cells. The
experiment was repeated six times. **P<0.01 and
***P<0.001, vs. Sham group. (E) Transmission electron
microscopy in cardiomyocytes. Arrows indicate mitochondria.
(original magnification, ×11,000). OGD, oxygen-glucose deprivation;
OGD/R, OGD/recovery; I, ischemia; R, reperfusion; CSA,
cross-sectional area. |
In addition, the effects of ischemia and reperfusion
on cardiomyocyte edema were examined in vivo. The results
revealed that the cardiomyocyte CSA in the infarct zone of the
ischemia group was significantly higher than that in the sham group
(Fig. 1D). Compared with the
ischemia and sham groups, the cardiomyocyte CSA in the infarct zone
of the I/R group was significantly increased. As shown in Fig. 1E, the mitochondria were swollen
and the space between the organelles was increased in the ischemia
group compared with the sham group; these changes were more
prominent in the I/R group. These results suggested that OGD and
OGD/R insults can lead to cardiomyocyte edema.
PEDF protects against OGD-induced
cardiomyocyte edema and injury
To investigate the effect of PEDF on cardiomyocytes
exposed OGD for 30 min (ischemia), the cell viability, and the
levels of cleaved caspase-3 and RIP3 were evaluated. Cleaved
caspase-3 and RIP3 are required for cell apoptosis (30) and necrosis (31), respectively. It was observed that
PEDF was downregulated in the infarct region following AMI and I/R.
Subsequently, a lentivirus carrying PEDF was transferred using
intramyocardial injections to induce its overexpression in a rat
AMI and I/R model (Fig. S1A and
B). The results revealed that PEDF pre-treatment significantly
alleviated OGD (ischemia)-induced cardiomyocyte edema (Fig. 2A). In addition, the level of
cleaved caspase-3 was significantly increased and cell viability
was decreased in the OGD group compared with the normal group;
however, no significant change in the protein expression of RIP3
was observed (Fig. 2B and C).
Furthermore, PEDF significantly alleviated the OGD-induced
cardiomyocyte apoptosis and decrease of cell viability. In
addition, PEDF pre-treatment significantly alleviated OGD
(ischemia)-induced cardiomyocyte edema in vitro and in
vivo (Fig. 2D and E). These
results demonstrated that PEDF has protective effects on
cardiomyocytes undergoing OGD to inhibit cell edema and injury.
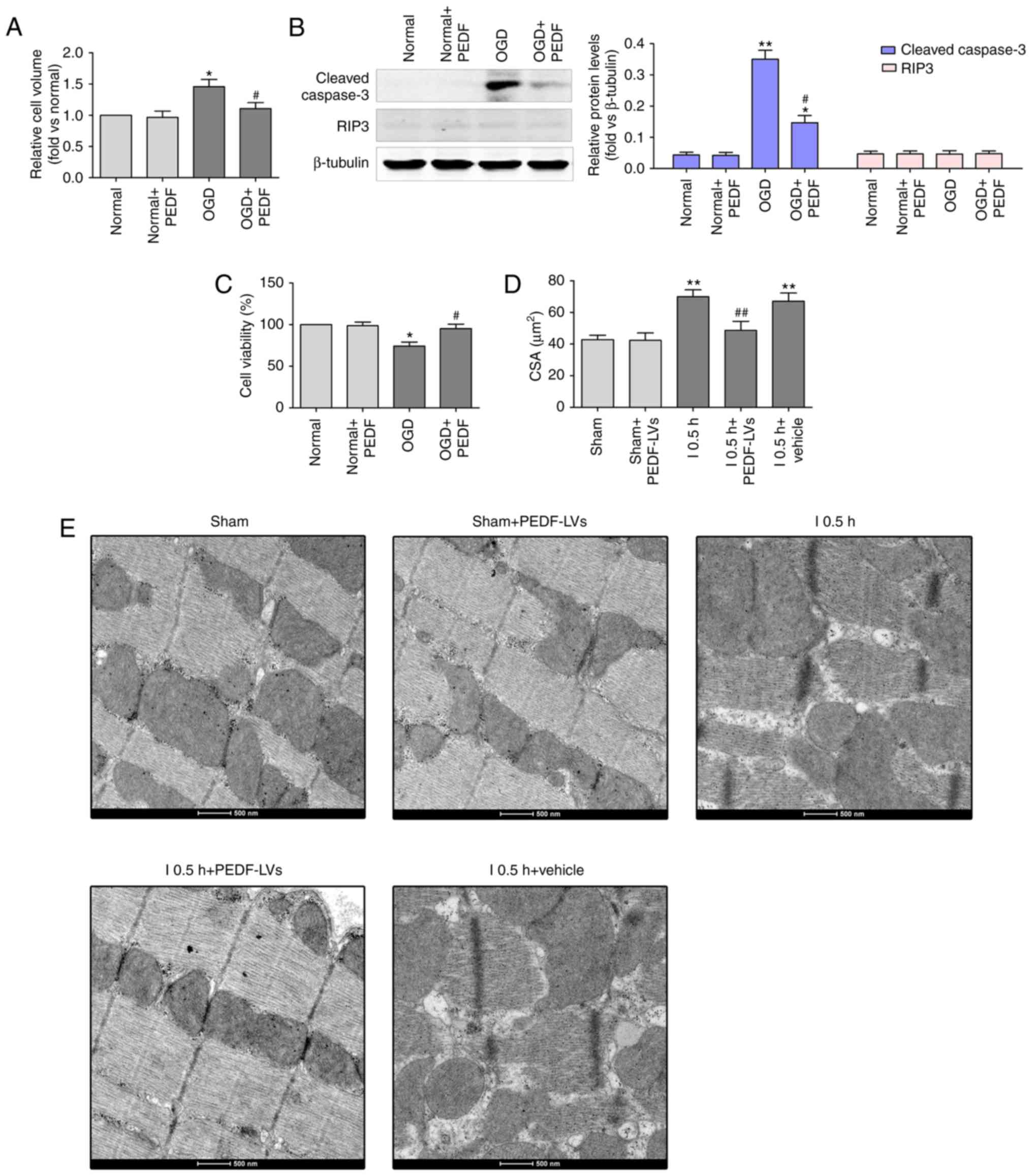 | Figure 2PEDF protects against OGD
(ischemia)-induced cardiomyocyte edema and injury. (A) Cell volumes
of 10 nmol/l PEDF-treated normal neonatal cardiomyocytes and
neonatal cardiomyocytes following 30 min of OGD were measured by
using confocal microscope stack scanning measurement (n=30).
*P<0.05, vs. relative normal group,
#P<0.05, vs. OGD group. (B) Protein levels of cleaved
caspase-3 and RIP3 in PEDF-treated normal neonatal cardiomyocytes
and neonatal cardiomyocytes following OGD were analyzed by western
blotting (n=6). (C) A Cell Counting kit-8 assay was performed to
assess cell viability in neonatal cardiomyocytes (n=6).
*P<0.05 and **P<0.01, vs. relative
normal controls; #P<0.05, vs. relative OGD controls.
(D) Cardiomyocyte CSAs were measured following ischemia. Rats were
divided into the Sham, Sham + PEDF-LVs, AMI, AMI + PEDF-LVs and AMI
+ vehicle groups (n=6). **P<0.01, vs. Sham group;
#P<0.05 and ##P<0.01, vs. AMI group.
(E) Transmission electron microscopy of cardiomyocytes (original
magnification, ×11,000). OGD, oxygen-glucose deprivation; PEDF,
pigment epithelium derived factor; LV, lentivirus; RIP3,
receptor-interacting protein 3; AMI, acute myocardial ischemia;
CSA, cross-sectional area. |
PEDF reduces the concentration of
Na+ and activation of glycolysis in cardiomyocytes
undergoing OGD
As shown in Fig.
3A, Na+ concentration was significantly increased in
cardiomyocytes during OGD compared with that in the normal group,
and was significantly reduced in the OGD + PEDF group. In addition,
the level of ATP was decreased in the OGD + PEDF group compared
with that in the OGD group (Fig.
3B). However, no significant difference was identified in the
activity of Na+-K+ ATPase between the OGD +
PEDF and normal groups (Fig. 3C).
Lactic acid content and ECAR were significantly increased in
cardiomyocytes during OGD compared with the normal group, and were
significantly reduced in the OGD + PEDF group (Fig. 3D and E). No significant
differences in the mRNA and protein expression levels of AQP1 were
observed in the OGD and OGD + PEDF groups compared with the normal
group (Fig. 3F and G). Notably,
no mRNA or protein expression of AQP4 was detected in
cardiomyocytes. These results demonstrated that PEDF treatment
decreased the concentration of Na+ and the activation of
glycolysis in cardiomyocytes during OGD.
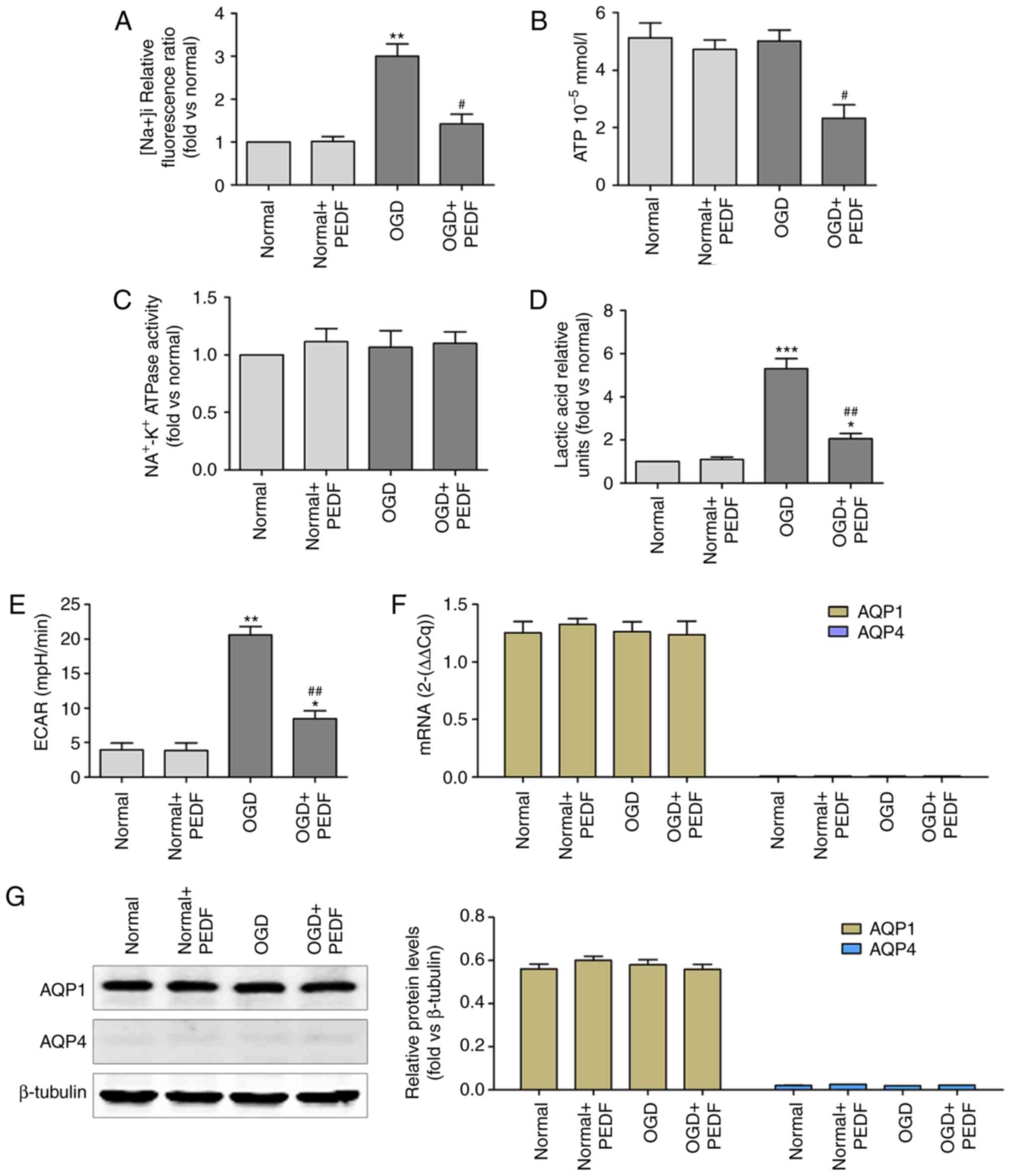 | Figure 3PEDF reduces Na+
concentration and glycolytic activation in cardiomyocyte undergoing
OGD. (A) Na+ concentration, (B) ATP concentration, (C)
Na+-K+ ATPase activity, (D) lactic acid
concentration, (E) extracellular acidification rate, and (F) mRNA
and (G) protein expression levels of aquaporin were measured in
neonatal cardiomyocytes. Cells were divided into the Normal, Normal
+ PEDF, OGD and OGD + PEDF groups (n=6). *P<0.05 and
**P<0.01 and ***P<0.001, vs. Normal
group; #P<0.05 and ##P<0.01, vs. OGD
group. OGD, oxygen-glucose deprivation; PEDF, pigment epithelium
derived factor; ECAR, extracellular acidification rate. |
Effects of PEDF on OGD-induced
cardiomyocyte edema and injury are associated with decreases in the
activation of glycolysis
The Na+ and lactic acid content were
increased in the PEDF + FDP group compared with the PEDF group
(Fig. 4A and B). Cardiomyocytes
that underwent OGD were treated with PEDF and the glycolytic
agonist, FDP, to increase ECAR, activating glycolysis (Fig. 4C). Furthermore, FDP significantly
inhibited the positive effects on the cardiomyocytes exposed to OGD
caused by PEDF (Fig. 4D and E),
and inhibited the inhibitory effects of PEDF on cardiomyocyte edema
(Fig. 4F). These results revealed
that glycolytic activation inhibited the PEDF-induced inhibition of
cardiomyocyte edema. Therefore, PEDF may inhibit OGD-induced
cardiomyocyte edema by decreasing the activation of glycolysis.
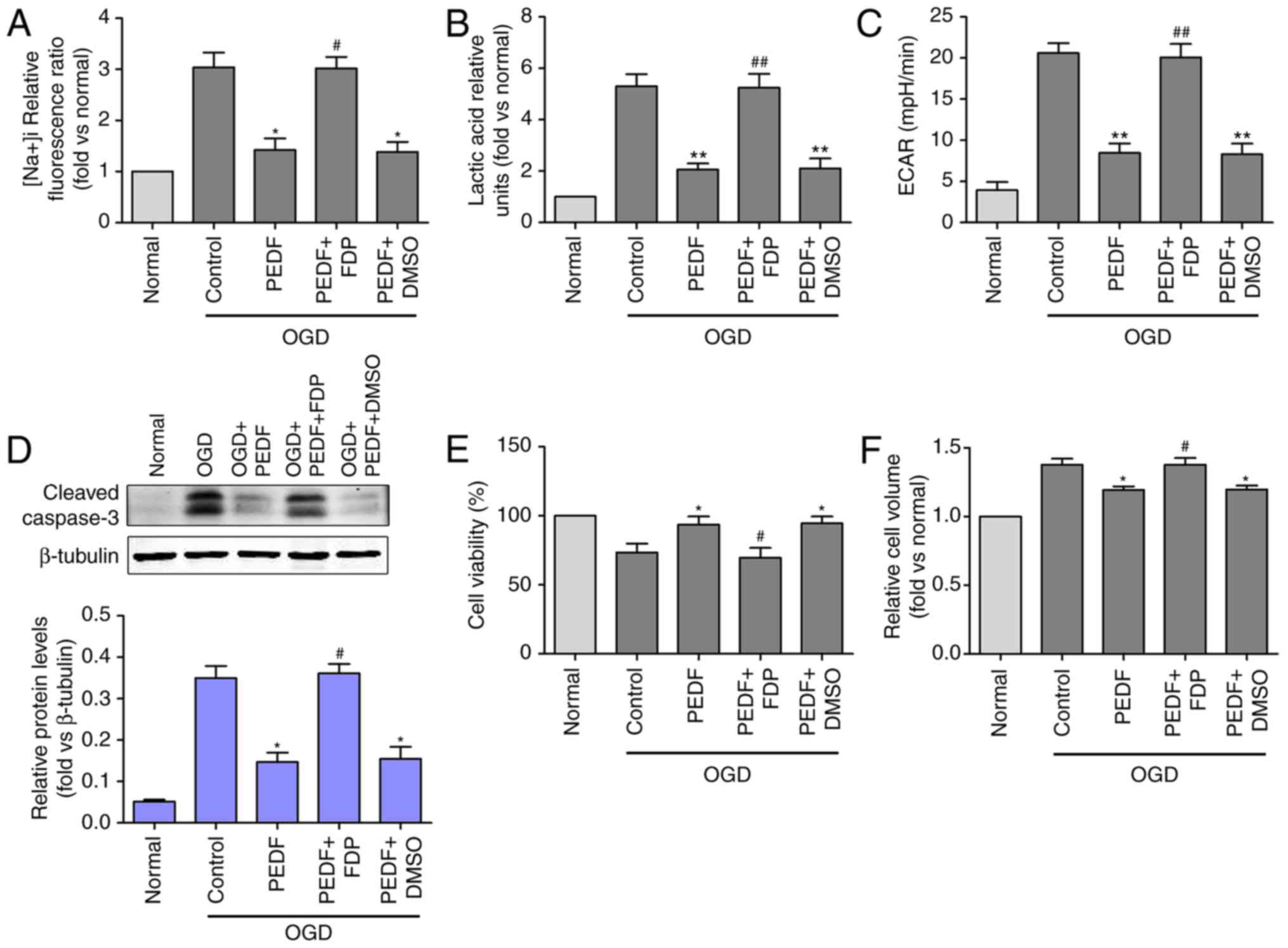 | Figure 4Effects of PEDF on OGD-induced
cardiomyocyte edema and injury are associated with decreases in
glycolytic activation. (A) Na+ concentration, (B) lactic
acid concentration, (C) extracellular acidification rate, (D)
protein expression of cleaved caspase-3, and (E) cell viability and
(F) cell volume were measured in neonatal cardiomyocytes. Cells
were divided into the OGD, OGD + PEDF, OGD + PEDF + FDP, OGD + PEDF
+ DMSO groups (n=6). *P<0.05 and
**P<0.01, vs. OGD group; #P<0.05 and
##P<0.01, vs. OGD + PEDF group. OGD, oxygen-glucose
deprivation; PEDF, pigment epithelium derived factor; FDP,
fructose-1, 6-diphosphate; RIP3, receptor-interacting protein 3;
ECAR, extracellular acidification rate. |
PEDF inhibits cardiomyocyte edema and
injury during OGD/R (I/R)
As PEDF decreased OGD-induced cardiomyocyte edema,
the present study investigated whether PEDF also reduced
cardiomyocyte edema during OGD/R. The results revealed that the
in vitro cell volume during OGD/R (I/R) was significantly
increased compared with that in the OGD group (Fig. 5A). The protein expression levels
of cleaved caspase-3 and RIP3 were increased, and cell viability
was decreased in the OGD/R group compared with the OGD group
(Fig. 5B and C). In addition, the
in vivo results demonstrated that the CSA, mitochondrial
edema and the space between organelles during OGD/R were
significantly increased compared with those in the OGD group
(Fig. 5D and E). Importantly,
PEDF treatment significantly inhibited cardiomyocyte edema during
OGD/R, PEDF significantly inhibited apoptosis and necrosis, and
increased cell viability in cardiomyocytes during OGD/R.
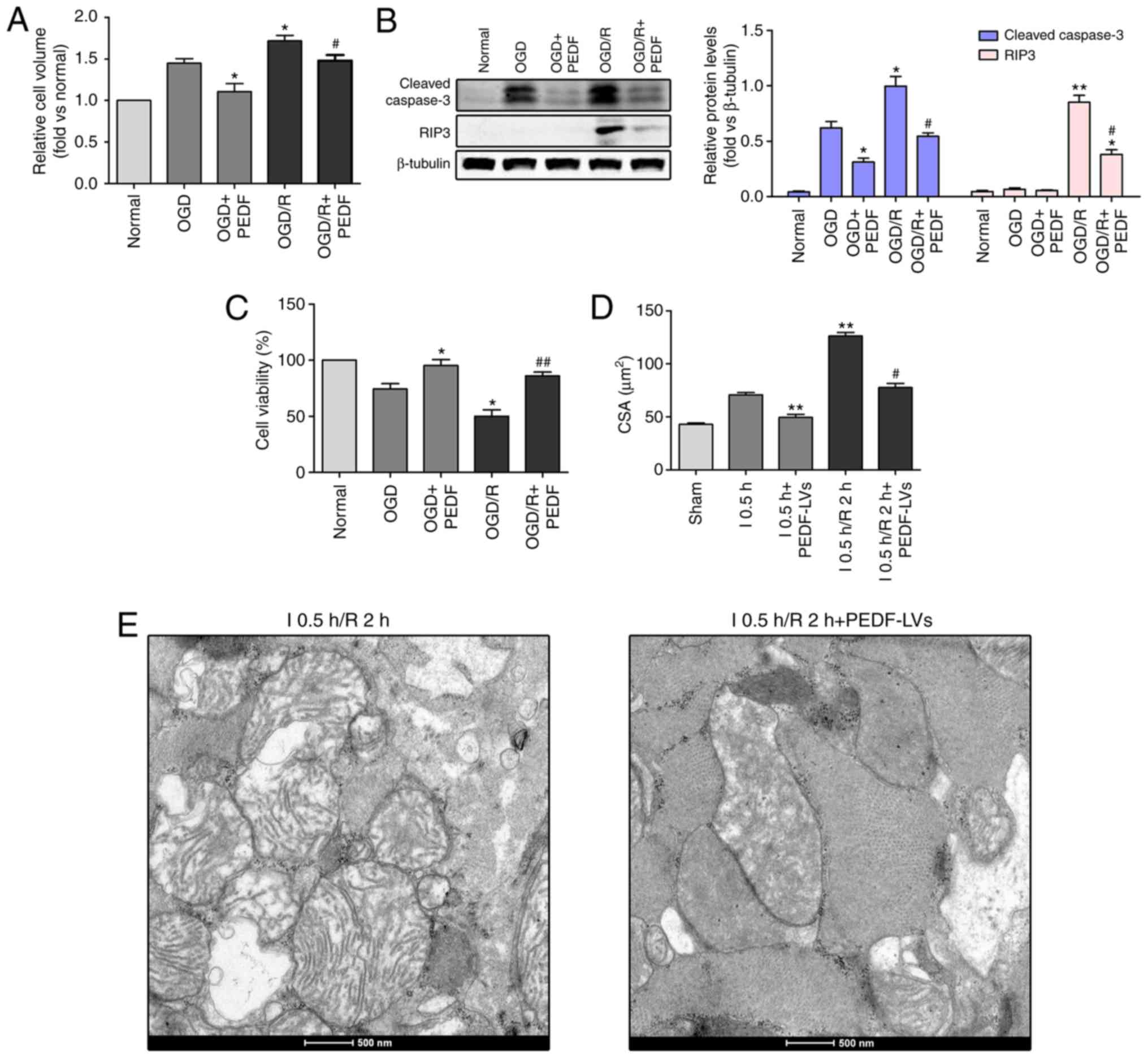 | Figure 5PEDF inhibits cardiomyocyte edema and
injury in OGD/R. (A) Cell volume, (B) protein expression of cleaved
caspase-3 and RIP3, and (C) cell viability were measured in
neonatal cardiomyocytes. Cells were divided into the OGD, OGD +
PEDF, OGD/R and OGD/R + PEDF groups (n=6). *P<0.05
and **P<0.01, vs. OGD control group;
#P<0.05 and ##P<0.01, vs. OGD/R control
group. (D) CSAs in the I 0.5 h, I0.5 h + PEDF-LVs, I 0.5 h/R2 h and
I 0.5 h/R 2 h + PEDF-LVs groups; the results are presented as the
mean CSA of 50 random cells (n=6). **P<0.01, vs. I
group; #P<0.05, vs. I 0.5 h/R 2 h group. (E)
Transmission electron microscopy of cardiomyocytes (original
magnification, ×11,000). OGD, oxygen-glucose deprivation; I,
ischemia; R, reperfusion; PEDF, pigment epithelium derived factor;
CSA, cross-sectional area; LV, lentivirus; RIP3,
receptor-interacting protein 3. |
Effects of PEDF on OGD/R cardiomyocyte
edema and injury are associated with a decrease of the expression
of AQP1
The mechanism of PEDF that allows it to reduce
OGD/R-induced cardiomyocyte edema was then investigated. Although
the mechanism of PEDF-induced reductions in edema in cardiomyocytes
undergoing OGD was demonstrated to be associated with a decrease of
glycolytic activity, no significant difference in Na+
concentration was identified between the OGD/R and OGD/R + PEDF
groups (Fig. 6A). No significant
differences in the ATP level and Na+-k+
ATPase activity were identified between the normal, OGD, OGD/R, and
OGD/R + PEDF groups (Fig. 6B and
C). Additionally, no significant differences in lactic acid
content or ECAR were identified between the OGD/R and OGD/R + PEDF
groups (Fig. 6D and E). Notably,
OGD/R significantly increased the mRNA and protein expression
levels of AQP1 in cardiomyocytes, whereas the expression of AQP1
decreased significantly following PEDF treatment (Fig. 6F and G). These results suggested
that PEDF decreases the expression of AQP1, rather than glycolytic
activity, in cardiomyocytes during OGD/R.
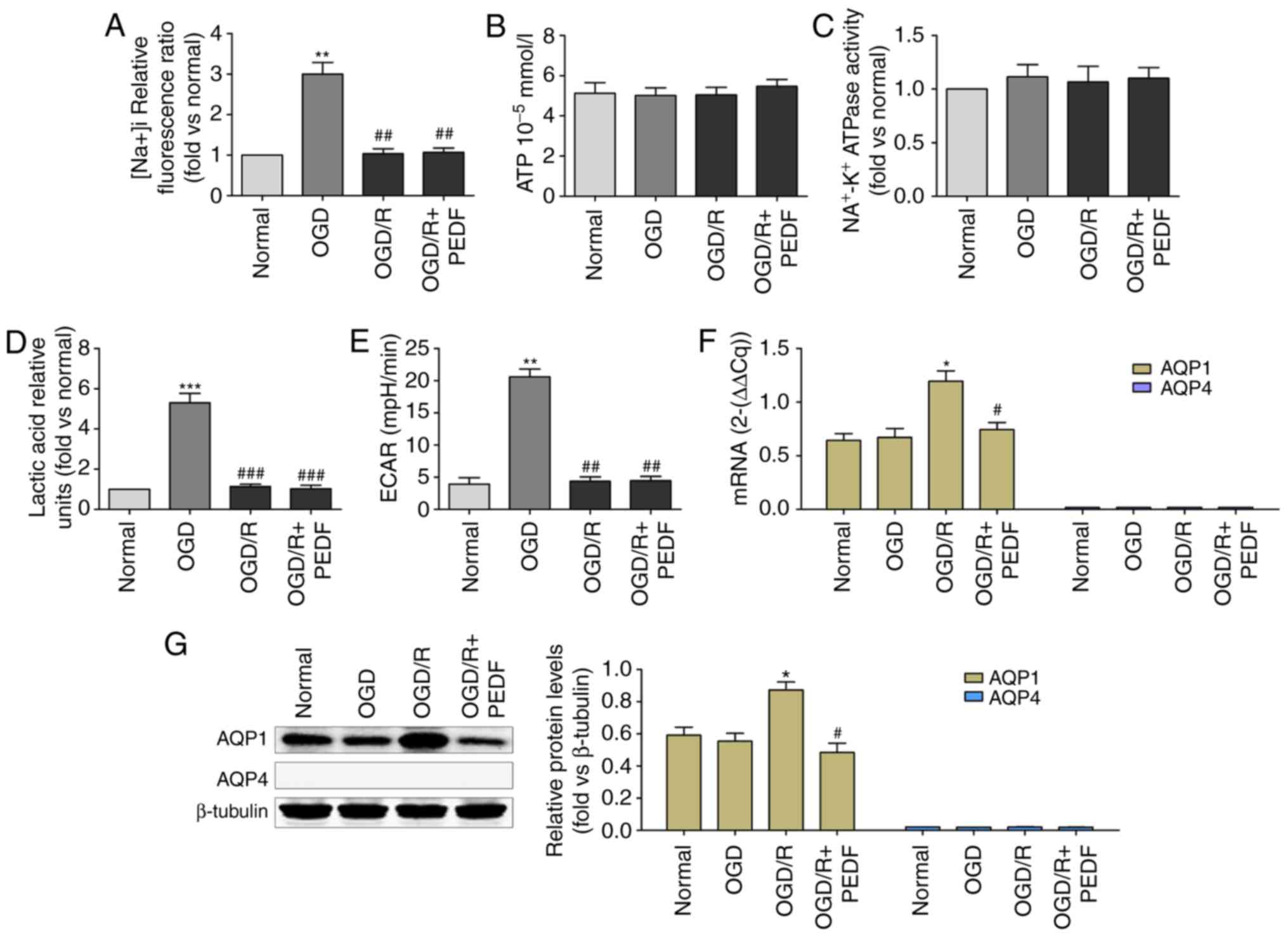 | Figure 6PEDF decreases the expression of AQP1
in cardiomyocytes during OGD/R. (A) Na+ concentration,
(B) ATP concentration, (C) Na+-K+ ATPase
activity, (D) lactic acid concentration, (E) extracellular
acidification rate, and (F) mRNA and (G) protein expression of AQP1
were measured in neonatal cardiomyocytes. Cells were divided into
the Normal, OGD, OGD/R and OGD/R + PEDF groups (n=6).
*P<0.05 and **P<0.01 and
***P<0.001, vs. Normal group; #P<0.05
and ##P<0.01, vs. OGD/R group. OGD, oxygen-glucose
deprivation; I, ischemia; R, reperfusion; PEDF, pigment epithelium
derived factor; AQP, aquaporin; ECAR, extracellular acidification
rate. |
Treatment with the AQP1 agonist AVP significantly
alleviated the inhibited expression of AQP1 induced by PEDF
(Fig. 7A-C). AVP treatment also
eliminated PEDF-induced protection against apoptosis and necrosis,
which prolonged cell viability in cardiomyocytes during OGD/R
(Fig. 7D and E). Notably,
OGD/R-induced cardiomyocyte edema was significantly increased in
the PEDF + AVP group compared with that in the PEDF group (Fig. 7F). These results demonstrated that
PEDF relieves OGD/R-induced cardiomyocyte edema via inhibiting the
mRNA and protein expression of AQP1.
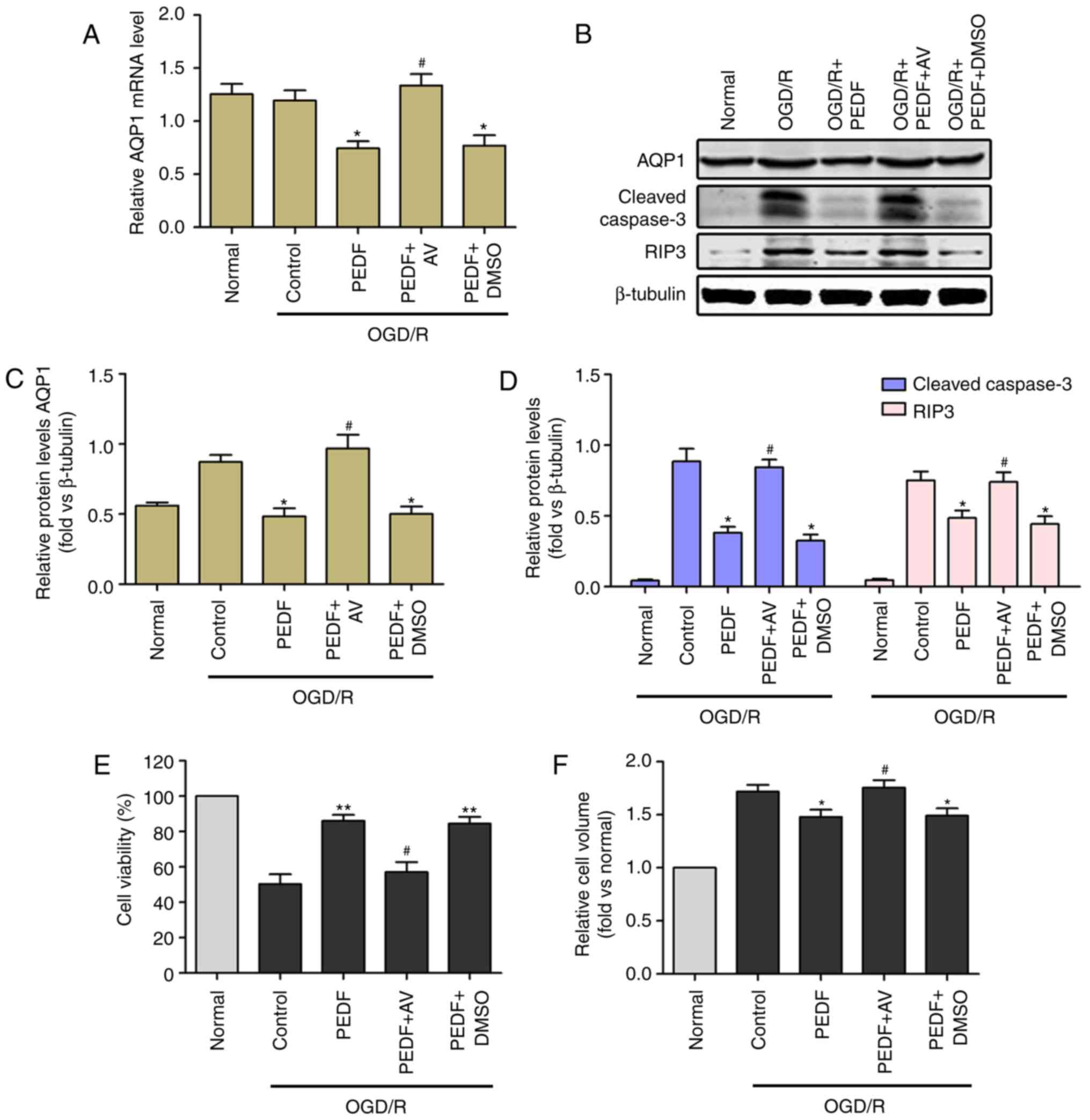 | Figure 7Reductions in cardiomyocyte edema and
damage induced by PEDF during OGD/R are associated with inhibition
of the expression of AQP1. (A) mRNA and (B) protein expression
levels of AQP1, protein expression of (C) cleaved caspase-3 and (D)
RIP3, and (E) cell viability and (F) cell volume were measured in
neonatal cardiomyocytes. Cells were divided into the OGD/R, OGD/R +
PEDF, OGD/R + PEDF + AV and OGD/R + PEDF + DMSO groups (n=6).
*P<0.05 and **P<0.01, vs. OGD/R group;
#P<0.05, vs. OGD/R + PEDF group. OGD, oxygen-glucose
deprivation; I, ischemia; R, reperfusion; PEDF, pigment epithelium
derived factor; AQP1, aquaporin 1; RIP3, receptor-interacting
protein 3; AVP, arginine vasopressin. |
Discussion
To the best of our knowledge, the present study is
the first to demonstrate that PEDF reduces OGD-induced
cardio-myocyte edema through decreasing lactate accumulation, and
inhibits the expression of AQP1 during the recovery period to
prevent further cell edema. In the present study, cardiomyocytes
exhibited significant edema following 30 min of OGD. Although the
edema of the cardiomyocytes was aggravated further following a
longer period of hypoxia, the changes were not significant. In
addition, although the present study did not examine changes in the
size of cellular edema in the myocardium during the I/R period,
significant cardiomyocyte edema following 30 min of ischemia and 30
min of ischemia/2 h of reperfusion were observed. PEDF inhibited
OGD-induced cardiomyocyte edema in vitro and ex
vivo.
It is noteworthy that PEDF reduced the ATP level in
cardiomyocytes during OGD. However, Na+ pump activation
was not inhibited in the PEDF group compared with that in the OGD
group, suggesting that the remaining ATP was sufficient to maintain
Na+ pump functionality and cell activity. The lactate
content and ECAR in PEDF-treated cardiomyocytes undergoing OGD was
significantly decreased compared with those in the OGD group,
indicating that PEDF inhibited H+ levels in
cardiomyocytes undergoing OGD. Inhibition of the activity of NHE
prevented Na+ influx, thereby reducing cellular osmotic
gradients and preventing the occurrence of OGD-induced
cardiomyocyte edema. Furthermore, no significant differences in the
Na+ pump, activity of NHE or lactate content were
identified between the OGD/R + PEDF and OGD + PEDF groups, which
indicated that PEDF was not able to alleviate cardiomyocyte edema
during the recovery period in terms of the further inhibition of
lactate accumulation.
Notably, no protein expression of AQP4 was detected
in rat cardiomyocytes, however, AQP4 mRNA was expressed. These data
are consistent with that of previous studies (12,32) and demonstrated that AQP4 is not
involved in the occurrence of cellular edema in rat cardiomyocytes
caused by OGD. The protein and mRNA expression levels of AQP1 were
detected in rat cardiomyocytes, and were significantly increased in
cardiomyocytes in the OGD/R group compared with those in the OGD
group, which demonstrates that PEDF has no significant effects on
the protein or mRNA levels of AQP1 in cardiomyocytes undergoing
OGD. PEDF reduced AQP1 protein content and thus downregulated
cardiomyocyte edema in the OGD/R period. The addition of the AQP1
agonist, AVP, inhibited the effects of PEDF on cardiomyocyte edema
during OGD/R. As PEDF also decreased the mRNA level of AQP1, it was
hypothesized that PEDF reduces the protein expression of AQP1 by
inhibiting AQP1 transcription. Previous studies have demonstrated
that the protein and mRNA expression of AQP1 may be downregulated
by microRNA (miR)-214 in the ischemic heart (33). Future investigations aim to
determine whether PEDF mediates the decreased protein and mRNA
levels of APQ1 via miRNA-214 and to examine the association between
PEDF and miR-214. Further investigations are required to elucidate
the mechanism.
A limitation of the present study was the
restriction of the experiments, as the effects of PEDF on
cardiomyocyte edema during OGD/R were only investigated in the
cells and interstitial space. In addition, although it was
determined that PEDF protected cardiomyocytes against edema when
they underwent 15 min-2 h of OGD and 30 min-12 h of recovery,
longer durations of OGD and recovery were been investigated.
In conclusion, the present study revealed that PEDF
protects cardiomyocytes during OGD/R (I/R) via decreasing lactate
accumulation and the expression of AQP1, respectively. These data
presents a novel mechanism by which PEDF inhibits cardiomyocyte
edema during OGD/R. The effect of PEDF on cellular edema may
contribute to maintaining cell viability and improve the prognosis
of patients with myocardial infarctions.
Supplementary Materials
Funding
The present study was supported by a grant from
National Nature Science Foundation of China (grant no. 81570242)
and the Natural Science Foundation of Jiangsu Province (grant no.
BK20150207).
Availability of data and materials
The authors declare that the materials described in
the manuscript, including all relevant raw data, are freely
available to any scientist wishing to use them for non-commercial
purposes, without breaching participant confidentiality.
Authors' contributions
BH, HM, FQ, HD and ZZ conceived and designed the
experiments. BH, HM, YY, ZL, XL, HZ, QZ and MW performed the
experiments. BH, HM, HD and ZZ analyzed the data. BH, HM, FQ and YY
acquired the reagents, materials and tools for analysis. BH and HM
produced the manuscript.
Ethics approval and consent to
participate
The experiments described here conform to the Guide
for the Care and Use of Laboratory Animals published by the
National Institutes of Health (34). All animal care and experimental
protocols were approved by the Animal Care and Use Committee of
Xuzhou Medical University (license no. SYXK 2002-0038; Jiangsu,
China) and international guidelines (European Council Directive
2010/63/EU) on the ethical use of animals (35).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to express thanks to Dr
Tianyun Wang (Laboratory of Pharmacology, Xuzhou Medical
University) for the HPLC analysis.
References
|
1
|
Garcia-Dorado D and Oliveras J: Myocardial
oedema: A preventable cause of reperfusion injury? Cardiovasc Res.
27:1555–1563. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Basuk WL, Reimer KA and Jennings RB:
Effect of repetitive brief episodes of ischemia on cell volume,
electrolytes and ultra-structure. J Am College Cardiol. 8(1 Suppl
A): 33A–41A. 1986. View Article : Google Scholar
|
|
3
|
Bragadeesh T, Jayaweera AR, Pascotto M,
Micari A, Le DE, Kramer CM, Epstein FH and Kaul S: Post-ischaemic
myocardial dysfunction (stunning) results from myofibrillar oedema.
Heart. 94:166–171. 2008. View Article : Google Scholar
|
|
4
|
Garcia-Dorado D, Oliveras J, Gili J, Sanz
E, Pérez-Villa F, Barrabés J, Carreras MJ, Solares J and
Soler-Soler J: Analysis of myocardial oedema by magnetic resonance
imaging early after coronary artery occlusion with or without
reperfusion. Cardiovasc Res. 27:1462–1469. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Butler TL, Egan JR, Graf FG, Au CG,
McMahon AC, North KN and Winlaw DS: Dysfunction induced by ischemia
versus edema: Does edema matter? J Thoracic Cardiovasc Surg.
138:141–147. 147 e1412009. View Article : Google Scholar
|
|
6
|
Sanz E, Garcia Dorado D, Oliveras J,
Barrabés JA, Gonzalez MA, Ruiz-Meana M, Solares J, Carreras MJ,
García-Lafuente A and Desco M: Dissociation between anti-infarct
effect and anti-edema effect of ischemic preconditioning. Am J
Physiol. 268:H233–H241. 1995.PubMed/NCBI
|
|
7
|
Friedrich MG, Abdel-Aty H, Taylor A,
Schulz-Menger J, Messroghli D and Dietz R: The salvaged area at
risk in reperfused acute myocardial infarction as visualized by
cardiovascular magnetic resonance. J Am Coll Cardiol. 51:1581–1587.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fernandez-Jimenez R, Sanchez-Gonzalez J,
Aguero J, García-Prieto J, López-Martín GJ, García-Ruiz JM,
Molina-Iracheta A, Rosselló X, Fernández-Friera L, Pizarro G, et
al: Myocardial edema after ischemia/reperfusion is not stable and
follows a bimodal pattern: Imaging and histological tissue
characterization. J Am Coll Cardiol. 65:315–323. 2015. View Article : Google Scholar
|
|
9
|
Leaf A: Maintenance of concentration
gradients and regulation of cell volume. Ann NY Acad Sci.
72:396–404. 1959. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vandenberg JI, Rees SA, Wright AR and
Powell T: Cell swelling and ion transport pathways in cardiac
myocytes. Cardiovasc Res. 32:85–97. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Inserte J, Garcia-Dorado D, Ruiz-Meana M,
Solares J and Soler J: The role of Na+-H+ exchange occurring during
hypoxia in the genesis of reoxygenation-induced myocardial oedema.
J Mol Cell Cardiol. 29:1167–1175. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Butler TL, Au CG, Yang B, Egan JR, Tan YM,
Hardeman EC, North KN, Verkman AS and Winlaw DS: Cardiac aquaporin
expression in humans, rats, and mice. Am J Physiol Heart Circ
Physiol. 291:H705–H713. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kruse E, Uehlein N and Kaldenhoff R: The
aquaporins. Genome Biol. 7:2062006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Manley GT, Fujimura M, Ma T, Noshita N,
Filiz F, Bollen AW, Chan P and Verkman AS: Aquaporin-4 deletion in
mice reduces brain edema after acute water intoxication and
ischemic stroke. Nat Med. 6:159–163. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Page E, Winterfield J, Goings G,
Bastawrous A and Upshaw-Earley J: Water channel proteins in rat
cardiac myocyte caveolae: Osmolarity-dependent reversible
internalization. Am J Physiol. 274:H1988–H2000. 1998.PubMed/NCBI
|
|
16
|
He X, Cheng R, Benyajati S and Ma JX: PEDF
and its roles in physiological and pathological conditions:
Implication in diabetic and hypoxia-induced angiogenic diseases.
Clin Sci (Lond). 128:805–823. 2015. View Article : Google Scholar
|
|
17
|
Gao X, Zhang H, Zhuang W, Yuan G, Sun T,
Jiang X, Zhou Z, Yuan H, Zhang Z and Dong H: PEDF and PEDF-derived
peptide 44mer protect cardiomyocytes against hypoxia-induced
apoptosis and necroptosis via anti-oxidative effect. Sci Rep.
4:56372014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao Q, Liu Z, Huang B, Yuan Y, Liu X,
Zhang H, Qiu F, Zhang Y, Li Y, Miao H, et al: PEDF improves cardiac
function in rats subjected to myocardial ischemia/reperfusion
injury by inhibiting ROS generation via PEDFR. Int J Mol Med.
41:3243–3252. 2018.PubMed/NCBI
|
|
19
|
Qiu F, Zhang H, Yuan Y, Liu Z, Huang B,
Miao H, Liu X, Zhao Q, Zhang H, Dong H and Zhang Z: A decrease of
ATP production steered by PEDF in cardiomyocytes with
oxygen-glucose deprivation is associated with an AMPK-dependent
degradation pathway. Int J Cardiol. 257:262–271. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang H, Hui H, Li Z, Pan J, Jiang X, Wei
T, Cui H, Li L, Yuan X, Sun T, et al: Pigment epithelium-derived
factor attenuates myocardial fibrosis via inhibiting endothelial-
to- mesenchymal transition in rats with acute myocardial
infarction. Sci Rep. 7:419322017. View Article : Google Scholar
|
|
21
|
Lu P, Zhang YQ, Zhang H, Li YF, Wang XY,
Xu H, Liu ZW, Li L, Dong HY and Zhang ZM: Pigment
epithelium-derived factor (PEDF) improves ischemic cardiac
functional reserve through decreasing hypoxic cardiomyocyte
contractility through PEDF receptor (PEDF-R). J Am Heart Assoc.
5:e0031792016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang H, Wang Z, Feng SJ, Xu L, Shi HX,
Chen LL, Yuan GD, Yan W, Zhuang W, Zhang YQ, et al: PEDF improves
cardiac function in rats with acute myocardial infarction via
inhibiting vascular permeability and cardiomyocyte apoptosis. Int J
Mol Sci. 16:5618–5634. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Maczewski M and Mackiewicz U: Effect of
metoprolol and ivabradine on left ventricular remodelling and Ca2+
handling in the post-infarction rat heart. Cardiovasc Res.
79:42–51. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun M, Ouzounian M, de Couto G, Chen M,
Yan R, Fukuoka M, Li G, Moon M, Liu Y, Gramolini A, et al:
Cathepsin-L ameliorates cardiac hypertrophy through activation of
the autophagylysosomal dependent protein processing pathways. J Am
Heart Assoc. 2:e0001912013. View Article : Google Scholar
|
|
25
|
Hao J, Li WW, Du H, Zhao ZF, Liu F, Lu JC,
Yang XC and Cui W: Role of vitamin C in cardioprotection of
ischemia/reperfusion injury by activation of mitochondrial KATP
channel. Chem Pharm Bull (Tokyo). 64:548–557. 2016. View Article : Google Scholar
|
|
26
|
Henry P, Popescu A, Puceat M, Hinescu ME
and Escande D: Acute simulated ischaemia produces both inhibition
and activation of K+ currents in isolated ventricular myocytes.
Cardiovasc Res. 32:930–939. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
28
|
Camilion de Hurtado MC, Portiansky EL,
Perez NG, Rebolledo OR and Cingolani HE: Regression of
cardiomyocyte hypertrophy in SHR following chronic inhibition of
the Na(+)/H(+) exchanger. Cardiovasc Res. 53:862–868. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li XD, Yang YJ, Geng YJ, Cheng YT, Zhang
HT, Zhao JL, Yuan JQ and Gao RL: The cardioprotection of
simvastatin in reperfused swine hearts relates to the inhibition of
myocardial edema by modulating aquaporins via the PKA pathway. Int
J Cardiol. 167:2657–2666. 2013. View Article : Google Scholar
|
|
30
|
Porter AG and Janicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang DW, Shao J, Lin J, Zhang N, Lu BJ,
Lin SC, Dong MQ and Han J: RIP3, an energy metabolism regulator
that switches TNF-induced cell death from apoptosis to necrosis.
Science. 325:332–336. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Frigeri A, Gropper MA, Umenishi F,
Kawashima M, Brown D and Verkman AS: Localization of MIWC and GLIP
water channel homologs in neuromuscular, epithelial and glandular
tissues. J Cell Sci. 108:2993–3002. 1995.PubMed/NCBI
|
|
33
|
Rutkovskiy A, Bliksoen M, Hillestad V,
Amin M, Czibik G, Valen G, Vaage J, Amiry-Moghaddam M and
Stensløkken KO: Aquaporin-1 in cardiac endothelial cells is
downregulated in ischemia, hypoxia and cardioplegia. J Mol Cell
Cardiol. 56:22–33. 2013. View Article : Google Scholar
|
|
34
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals. National
Academies Press; Washington, DC: pp. 2202011
|
|
35
|
European Commission: Directive 2010/63/EU
of the European Parliament and of the council of 22 September 2010
on the protection of animals used for scientific purposes. Off J
Eur Union. L276:33–79. 2010.
|














