Introduction
Fibrosis is a protective reaction that is activated
in response to hepatic injury, causing a variety of diseases that
result in hepatocellular death. Therefore, liver fibrosis is
observed in patients with chronic viral hepatitis, non-alcoholic
fatty liver disease, alcoholic liver disease, obesity, cholestatic
and autoimmune liver diseases (1,2).
Previous studies have indicated that the inflammatory response in
the liver plays a crucial role in hepatic fibrogenesis during
hepatic fibrosis (HF) (3).
Hepatic macrophages, or Kupffer cells (KCs), are important immune
cells that are associated with the pathogenesis of chronic liver
injury, and have been recognized as potential targets for use in
the treatment of fibrosis (4).
Liver fibrosis has been widely demonstrated to be a reversible
process. Hepatic macrophages can serve dual functions in the
progression of experimental hepatic fibrosis, and can reverse
mechanisms that are associated with the degradation of excessive
extracellular matrix deposition in the liver (5). The investigation into novel
compounds that can be used to regulate macrophage function is
required for the identification of a therapeutic strategy for
HF.
Carbon tetrachloride (CCl4)-induced HF is
characterized by the activation of KCs and the relevant immune
response, which results in the secretion of cytokines, chemokines
and other pro-inflammatory factors (6). The mechanisms underlying the role of
macrophages in HF have not been fully elucidated. Macrophages are
classified into two phenotypes, those with M1 type pro-inflammatory
function and those with M2 type-immunoregulatory function (7,8).
Different phenotypes of macrophages have been identified to carry
out different functions in the progression of HF. M1 macrophages
are considered to induce pro-inflammatory cytokines and
fibrogenesis, while M2 macrophages can be subdivided into at least
5 subtypes; however, their function in inflammation and fibrosis
remains undetermined (9).
Previous studies have demonstrated that a variety of M2 subtypes
may exhibit pro- or anti-fibrotic activity (10-13). The mechanisms of macrophages as
regards the regulation of liver fibrosis are associated with
macrophage polarization. A variety of macrophage subtypes secrete a
number of different cytokines, and this can result in a dual
function response during the progression of HF.
The current study aimed to determine the effects of
margatoxin (MgTX) on the polarization of macrophages in RAW264.7
cells, and to detect the serum levels of inflammatory cytokines
following MgTX treatment in a model of HF. Hepatic macrophages in
the liver, which are also known as KCs, when activated during an
inflammatory condition, result in the release of a number of
pro-inflammatory cytokines and chemokines, and an increase the
activation of hepatic stellate cells (HSCs) (14). M1 macrophages are classical
macrophages that exhibit antiviral activity and release
pro-inflammatory cytokines, including tumor necrosis factor-α
(TNF-α), interleukin (IL)-1β and reactive oxygen species, while M2
macrophages are activated macrophages that promote the defense
against parasitic infections and are associated with tissue
remodeling, and secrete immune-modulatory cytokines, including
IL-10, transforming growth factor-β (TGF-β), IL-4 and IL-13
(15). Increasing evidence has
indicated that a number of pro-inflammatory cytokines, including
TNF-α and TGF-β, play a key role in the initiation of stellate cell
activation. In other liver diseases, fibrosis has been indicated to
be associated with the production of M2-associated cytokines and
TGF-β (15). However, whether
these cytokines originate from M2 macrophages during the modulation
of extracellular matrix remodeling remains undetermined. Previous
studies had revealed that M2 macrophage-derived cytokines,
including IL-4, IL-13 and IL-10, regulate the chronic inflammatory
response and ameliorate wound healing in the liver (16-19). On the whole, different macrophage
subtypes secrete a number of different cytokines, which play key
roles in the progression of HF.
Kv1.3 is a voltage-gated potassium channel subtype,
which has 6 transmembrane domains, including a voltage sensor and a
pore-forming region (20). MgTX
is a 39 amino-acid-long peptide that is stabilized by 3 disulfide
bridges, has a molecular weight of 4,185 and is isolated from the
venom of the scorpion Centruroidesn margaritatus. MgTX
exhibits a high affinity for Kv1.3 and is a selective inhibitor of
the Kv1.3 channel. Kv1.3-selective blockers have been indicated to
be effective in reducing disease severity in a number of animal
models of autoimmunity (21-24). Previous studies have demonstrated
that the selective Kv1.3-channel inhibition can significantly
suppress the number of leukocytes, and the progression of renal
fibrosis exhibits a significant decrease in cortical cell cycle
marker expression. Active Kv1.3 channels can promote cardiac
fibrosis by secreting TGF-β. The blockade of voltage-dependent
potassium channels (Kv) by specific antagonists decreases
macrophage cytokine production and inhibits proliferation (25-27). However, whether MgTX is associated
with HF has not, to the best of our knowledge, yet been determined.
The current study aimed to assess whether MgTX can ameliorate HF
in vivo, and to explore the underlying mechanisms that
govern the association between Kv1.3 and macrophages polarization
and cytokine secretion.
Materials and methods
Mouse model of HF
All animal protocols were approved by the Animal
Care and Use Committee of Anhui Medical University, China. Male
C57BL/6 mice (age, 4-6 weeks) were obtained from the Experimental
Animal Center of Anhui Medical University for use in creating the
model of CCl4-induced liver fibrosis. Early HF was generated by a
bi-weekly intraperitoneal injection of CCl4 (20% in olive oil) at a
dose of 0.01 ml/g/mouse for 4 weeks. The control mice were treated
intraperitoneally with the same volume of olive oil at the same
time intervals. MgTX was diluted in PBS and injected at 1 ml by
intraperitoneal injection at 3 different doses 1 h prior to the
CCl4 (olive oil) injection, namely the doses of MgTX used were
0.4179 mg/kg (high, H), 0.139 mg/kg (moderate, M), 0.0418 mg/kg
(low, L).
Collection of mouse blood and
tissues
Blood samples were collected from orbital sinus by
rapidly removing the eyeball after the mice were anesthetized with
chloral hydrate (4%, 400 mg/kg) by intraperitoneal injection.
Approximately 1 ml blood was collected in an EP tube for each
mouse. Following blood sample collection, the mice were sacrificed
by cervical dislocation. Liver tissues were obtained by a midline
laparotomy. Liver tissues were either fixed in 4% formalin for
immunohistochemistry, or stored at -80°C for use in western blot
analysis. All animal studies were conducted following the approval
from the Institutional Animal Care and Use Committee.
Histopathology and immunohistochemical
staining
The central left hepatic lobe was sectioned and
fixed in 4% paraformaldehyde for 48 h. Tissues were then embedded
in paraffin, and 5-µm-thick slices were stained using
hematoxylin (BA-4041, Baso) and eosin (BA-4022, Baso) for
morphological analysis, stained with hematoxylin for 20 min and
eosin for 1 min at room temperature. Masson's trichrome and Sirius
red staining were used for the evaluation of collagen expression.
Masson's trichrome kit (MST-8003/8004, Maixin-Bio) and Sirius red
staining kit (36324ES60, Yeasen Biotechnology (Shanghai) Co., Ltd.)
were carried out according to the manufacturer's instructions at
room temperature. Immunostaining was performed on paraffin-embedded
materials. The following primary antibodies were used: Matrix
metalloproteinase (MMP)12 rabbit polyclonal (diluted 1:100;
22989-1-AP, Proteintech), MMP13 rabbit polyclonal (diluted 1:200;
bs-10581R, Bioss), periostin (POSTN) rabbit polyclonal (diluted
1:200; bs-4994R, Bioss), C-C motif chemokine ligand 2 (CCL2) rabbit
polyclonal (diluted 1:100; bs-1101R, Bioss), Kv1.3 rabbit
polyclonal (diluted 1:100; bs-10229R, Bioss), δ-catenin rabbit
polyclonal (diluted 1:200; bs-7000R, Bioss), α-SMA rabbit
polyclonal (diluted 1:200; ab5694, Abcam), collagen I rabbit
polyclonal (diluted 1:150; bs-10423R, Bioss) were used; the primary
antibody reactions were incubated for 30 min at room temperature. A
secondary antibody kit (PV-6000, Zsbio) was used for
immunohistochemical staining. The average optical density for the
semi-quantitative analysis of immunohistochemical staining was
performed by CaseViewer (pananoramic MIDI, 3DHISTECH) and
JD801.
ELISA
Serum was extracted from blood by centrifugation
(-1,000 × g, 10 min) at 4°C and stored at -80°C for subsequent use.
The levels of IL-1β, IL-6, IL-10, IL-20, TNF-α and TGF-β were
determined using mouse serum using ELISA kits according to the
manufacturer's protocol. A mouse IL-1β ELISA kit (cat. no.
RK00006), mouse IL-6 ELISA kit (cat. no. RK00008), mouse IL-10
ELISA kit (cat. no. RK00016), mouse IL-20 ELISA kit (cat. no.
RK00110), mouse TNF-α ELISA kit (cat. no. RK00027) and a mouse
TGF-β1 ELISA kit (cat. no. RK00057) were used and were all
purchased from ABclonal, Inc.
Cells and cell culture
RAW264.7 cells were purchased from the Type Culture
Collection of the Chinese Academy of Sciences. The cells were
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with FBS (10%; Gibco; Thermo Fisher Scientific, Inc.),
penicillin and streptomycin (1%), and cultured at 37°C in a 5%
CO2 incubator. The RAW264.7 cells were treated with
lipopolysaccharide LPS (1 µg/ml) combined with interferon
(IFN)-γ (10 ng/ml) for 24 h to polarize the M1 macrophage
phenotype, or were treated with IL-4 (15 ng/ml) for 24 h to induce
the M2 macrophage phenotype. The RAW264.7 cells were also treated
with MgTX (10 nM) for 24 h at 37°C, as previously described
(27).
Total RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from the RAW264.7 cell lines
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). A ThermoScript RT-qPCR synthesis kit (Fermentas;
Thermo Fisher Scientific, Inc.) was used to synthesize the cDNA
according to the manufacturer's protocol. RT-qPCR-was performed
using ThermoScript RT-qPCR kits (Fermentas; Thermo Fisher
Scientific, Inc.). β-actin was used for normalization. Relative RNA
expression was calculated using the standard 2−ΔΔCq
method (28). Each experiment was
performed in triplicate and repeated at least 3 times. The
sequences of the primers (5′-3′) are presented in Table I.
 | Table ISequences of primers used for
RT-qPCR. |
Table I
Sequences of primers used for
RT-qPCR.
| Gene | Amplicon size
(bp) | Forward primer
(5′→3′) | Reverse primer
(5′→3′) |
|---|
| Mouse | β-actin | 120 |
AGTGTGACGTTGACATCCGT |
TGCTAGGAGCCAGAGCAGTA |
| CCL-2 | 128 |
AACTGCATCTGCCCTAAGGT |
CTGTCACACTGGTCACTCCT |
| Mrc2 | 83 |
ACGACTGTGAGACCTTCTGG |
CCTCCAGGACAGTGTGGATT |
| IL-10 | 88 |
TGCACTACCAAAGCCACAAG |
TCAGTAAGAGCAGGCAGCAT |
| Arg-1 | 135 |
GCAGTTGGAAGCATCTCTGG |
GAGAAAGGACACAGGTTGCC |
| TNF-α | 133 |
GACAGTGACCTGGACTGTGG |
TGAGACAGAGGCAACCTGAC |
| IL-1β | 98 |
GAAGAAGAGCCCATCCTCTG |
TCATCTCGGAGCCTGTAGTG |
| CD163 | 94 |
ATGGGTGGACACAGAATGGT |
AGCTCACAGCCACAACAAAG |
| iNOS | 143 |
CCTTGTTCAGCTACGCCTTC |
CTTCAGAGTCTGCCCATTGC |
Western blot analysis
The RAW264.7 cells were lysed using protein
extraction solution (Beyotime Institute of Biotechnology). The
protein concentration of each sample was measured using a NanoDrop
2000 Spectrophotometer (Thermo Fisher Scientific, Inc.). Samples
(20 µg/lane) underwent SDS-PAGE (10%) and were transferred
onto PVDF membranes (EMD Millipore). The PVDF membranes were
blocked with 5% non-fat milk for 2 h, and subsequently incubated
with primary antibodies at 4°C overnight. Primary antibodies
against δ-catenin (1:300, cat. no. bs-7000R, Bioss), α-SMA (1:300,
cat. no. bs-0189R, Bioss), collagen I (1:300, cat. no. bs-10423R,
Bioss), MMP12 (1:600, cat. no. 22989-1-AP, Proteintech), MMP13
(1:300, cat. no. bs-10581R, Bioss), POSTN (1:300, cat. no.
bs-4994R, Bioss), CCL2 (1:500, cat. no. bs1101R, Bioss), Kv1.3
(1:300, cat. no. bs-10229R, Bioss), iNOS (1:1,000, cat. no. ab4999,
Abcam), TNF-α (1:500, cat. no. BS1857, Bioworld), IL-β (1:300, cat.
no. bs0812R, Bioss), IL-10 (1:1,000, cat. no. bs0698R, Bioss),
CD163 (1:300, cat. no. bs2527R, Bioss), Arg-1 (1:200, cat. no.
sc-47715, Santa Cruz Biotechnology), Mrc2 (1:500, cat. no. ab70132,
Abcam) and β-actin (1:500, cat. no. TA-09, Zs-BIO) were used.
Subsequently, the PVDF membranes were washed 3 times with TBST,
followed by incubation with secondary antibodies [HRP-goat
anti-rabbit IgG (1:1,000; cat. no. ZB-2305; Zs-BIO)] at room
temperature for 1.5 h. After washing 3 times with TBST, the protein
blots were observed using an ECL-chemiluminescent kit (ECL-plus;
Thermo Fisher Scientific, Inc.),and the gray values of the bands
were measured using ImageJ software (version 1.4.3.67).
Statistical analysis
Statistical analysis was performed using the
Statistical Package for Social Sciences v.13.0 (SPSS, Inc.). Data
are summarized from at least 3 experiments and are presented as the
means ± standard deviation. The statistical evaluation of the
results was performed using a one-way ANOVA. Multiple comparisons
between the groups were performed using the SNK method. A value of
P<0.05 was considered to indicate a statistically significant
difference.
Results
Kv1.3 inhibition alleviates CCl4-induced
liver fibrosis in mice
To investigate the degree of HF in C57BL/6J mice
with CCl4-induced HF, histopathological analysis was performed.
Hematoxylin and eosin staining, Masson's staining and Sirius Red
staining demonstrated that the CCl4 (20%) injection caused
significant liver fibrosis, including serious steatosis,
inflammatory cell infiltration and fibrosis in the main block of
mouse livers compared with the control group. In comparison, no
evidence of liver injury was observed in the control group
with/without MgTX (0.4179 mg/kg). MgTX was therefore indicated to
attenuate liver fibrosis in mice with CCl4-induced HF (Fig. 1A). Immunohistochemistry from the
liver fibrosis tissues revealed extensive α-SMA- and collagen
I-positive staining, whereas the liver tissues in the control group
with/without MgTX indicated decreased staining. Additionally, the
results of western blot analysis for α-SMA and collagen I protein
expression were consistent with the results from
immunohistochemistry (Fig. 1B and
C). These results indicated that the mouse model of
CCL4-induced liver fibrosis was successfully established and that
MgTX protected the mice from CCl4-induced HF.
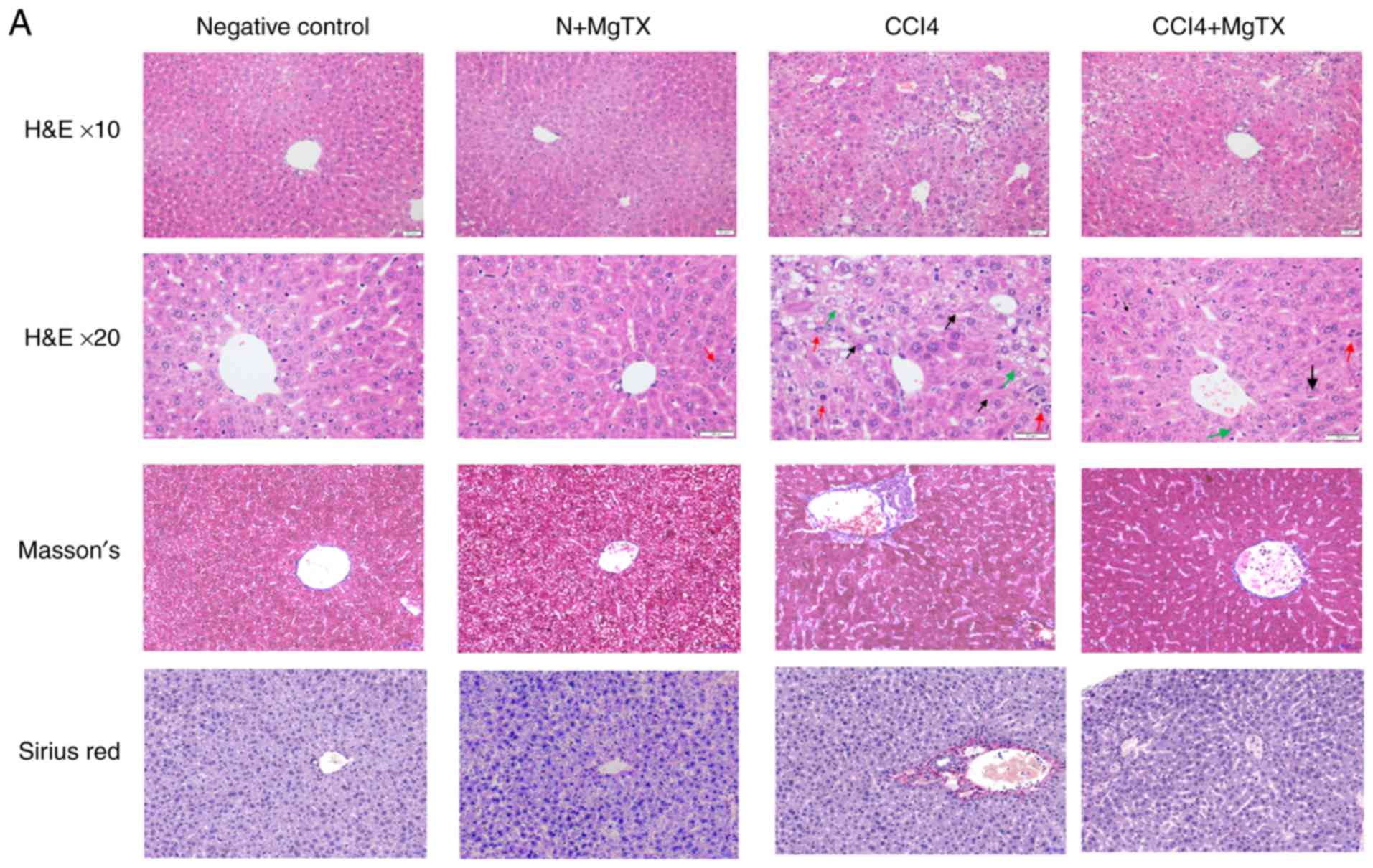 | Figure 1MgTX (0.4179 mg/kg) ameliorates
CCl4-induced HF in C57BL/6J mice. (A) Representative hematoxylin
and eosin staining, Masson's staining and Sirius Red staining of
liver tissues indicated an incomplete hepatic lobule structure and
irregularly arranged hepatic plates. Scattered degeneration,
necrosis and numerous inflammatory cells immersed in liver tissue
were identified compared with the HF model group treated with MgTX
(magnification, ×200; black arrows, fibrosis; green arrows,
steatosis; red arrows, inflammatory cell infiltration). (B)
Immunohistochemistry and (C) western blot analysis of hepatic
fibrosis tissues revealed increased α-SMA and collagen I expression
compared with the CCl4-exposed group treated with MgTX.
*P<0.05, ***P<0.001 vs. CCl4;
#P<0.05, ###P<0.001 vs. negative
control. Representative views from each group are presented
(original magnification, ×20; n=6). MgTX, margatoxin; CCl4, carbon
tetrachloride; HF, hepatic fibrosis. |
MgTX decreases the expression of
HF-associated proteins in mice
To investigate the expression levels of proteins
that are associated with HF in mice following the MgTX injection,
immunohistochemistry and western blot analysis were performed to
determine the expression of MMP12, MMP13, POSTN, δ-catenin, Kv1.3
and CCL2. Immunohistochemical analysis of the liver tissues
revealed that all proteins in the MgTX-treated mice with HF were
decreased compared with the tissues from the mice with CCl4-induced
HF not treated with MgTX (Fig.
2).
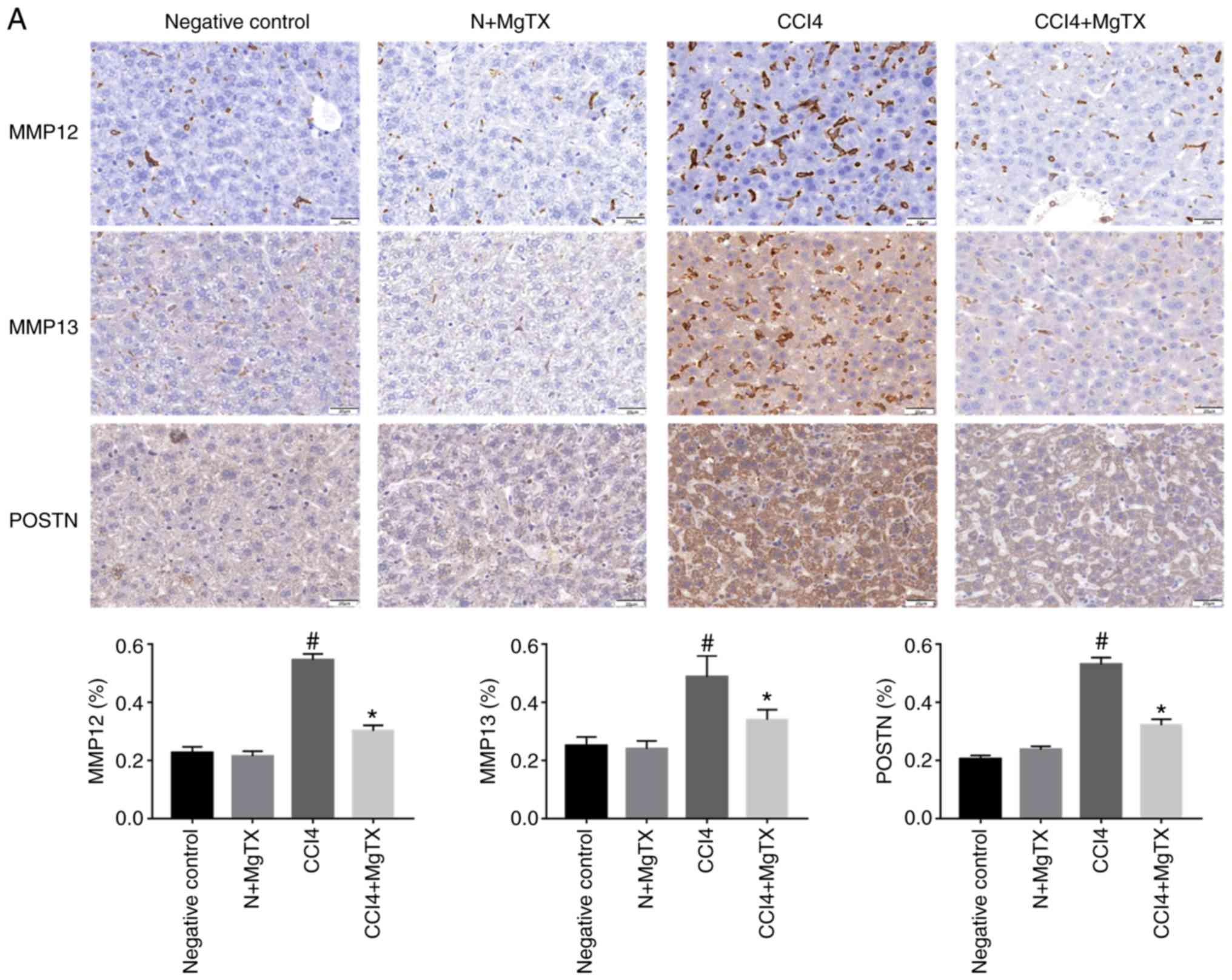 | Figure 2MgTX (0.4179 mg/kg) decreases the
expression of HF-associated proteins in C57BL/6J mice with
CCl4-induced HF (×200 field). (A) MMP12, MM13 and POSTN were
associated with the amount of ECM and collagen.
Immunohistochemistry of liver fibrosis tissues revealed MMP12, MM13
and POSTN exhibited increased staining compared with the HF group
treated with MgTX. *P<0.05, CCl4 vs. CCl4 + MgTX
group; #P<0.05, CCl4 vs. N group. (B) CCL2, Kv1.3 and
δ-catenin are associated with the migration of macrophages.
Immunohistochemistry of liver fibrosis tissues revealed increased
staining for CCL2 and Kv1.3 compared with the HF group treated with
MgTX. *P<0.05, CCL4 vs. CCL4 + MgTX group;
#P<0.05, CCL4 vs. N group. n=6. MgTX, margatoxin; HF,
hepatic fibrosis; CCl4, carbon tetrachloride; CCL2, C-C motif
chemokine ligand 2; MMP, matrix metalloproteinase; POSTN,
periostin; ECM, extracellular matrix. |
MgTX decreases the expression of
HF-associated proteins in RAW264.7 cells
The results of immunohistochemistry demonstrated
that the aforementioned proteins were expressed in the liver
tissue. Therefore, the current study aimed to determine protein
expression in macrophages. The proteins associated with HF were
determined in RAW264.7 cells treated with MgTX (10 nM; Control; M1,
M1 + MgTX and M2, MgTX and M2 + MgTX), to examine whether
macrophages are associated proteins that have been indicated to
play a role in HF. This analysis was also performed to determine
whether macrophages play a critical role in regulating the
expression of proteins associated with HF. The levels of MMP12,
MMP13, POSTN, CCL2, Kv1.3 and δ-catenin were determined by western
blot analysis. The results indicated that the levels of proteins
associated with HF in the M1 phenotype macrophages treated with
MgTX decreased compared with the other M1 phenotype groups.
However, the levels of proteins associated with HF in the M2
phenotype macrophages treated with MgTX were downregulated or
upregulated compared with the other M2 phenotype groups (Fig. 3).
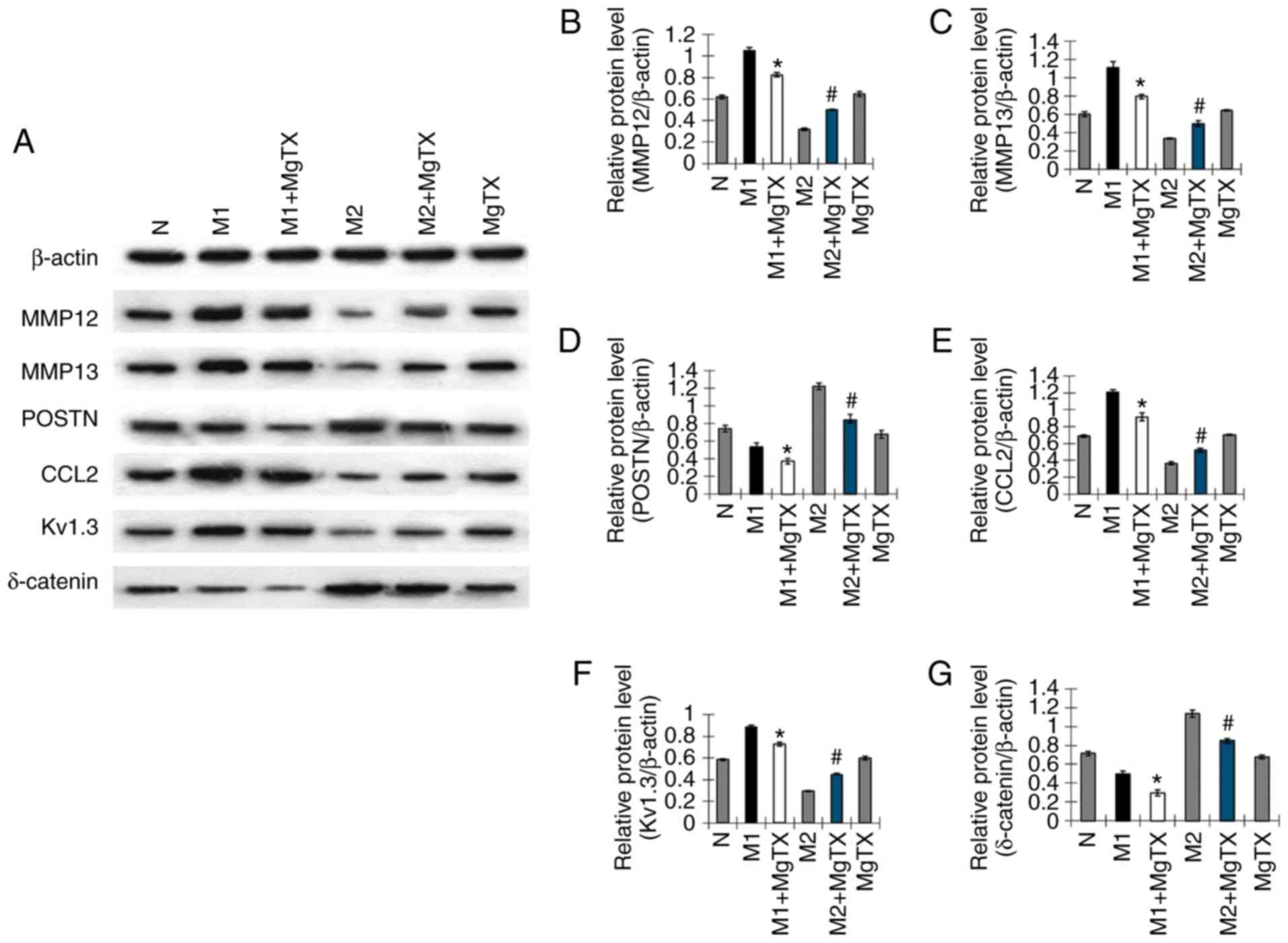 | Figure 3MgTX decreases the expression of
HF-associated proteins in RAW264.7 cells. To confirm the results of
HF-associated proteins from immunohisto-chemistry in RAW264.7
cells, (A) western blot analysis was performed to examine the
expression of MMP12, MM13, POSTN, CCL2, Kv1.3 and δ-catenin, which
were downregulated by MgTX in the M1 phenotype groups. (B-G) MMP12,
MM13, CCL2 and Kv1.3 were upregulated by MgTX in the M2 phenotype
groups; however, POSTN and δ-catenin were downregulated by MgTX in
the M2 phenotype groups. No significant difference was observed
between the control (N) and MgTX groups. *P<0.05 vs.
M1 group; #P<0.05 vs. M2 group. n=3. MgTX,
margatoxin; HF, hepatic fibrosis; MMP, matrix metalloproteinase;
POSTN, periostin; CCL2, C-C motif chemokine ligand 2. |
MgTX regulates the secretion of cytokines
in vivo
To explore the functional role of Kv1.3 in the
regulation of cytokine secretion in CCl4-induced HF, ELISAs were
performed to evaluate the circulation levels of inflammatory
cytokines in the serum of mice with CCl4-induced HF. The results
revealed that the serum levels of pro-inflammatory cytokines,
including IL-1β, IL-6, IL-20 and TNF-α were significantly decreased
with MgTX treatment (0.4179 mg/kg), while the levels of
anti-inflammatory cytokines, including IL-10, were increased in the
mice with CCl4-induced HF treated with MgTX (0.4179 mg/kg).
However, TGF-β expression was not significantly decreased in the
CCL4 + MgTX treatment group compared with the model groups
(Fig. 4).
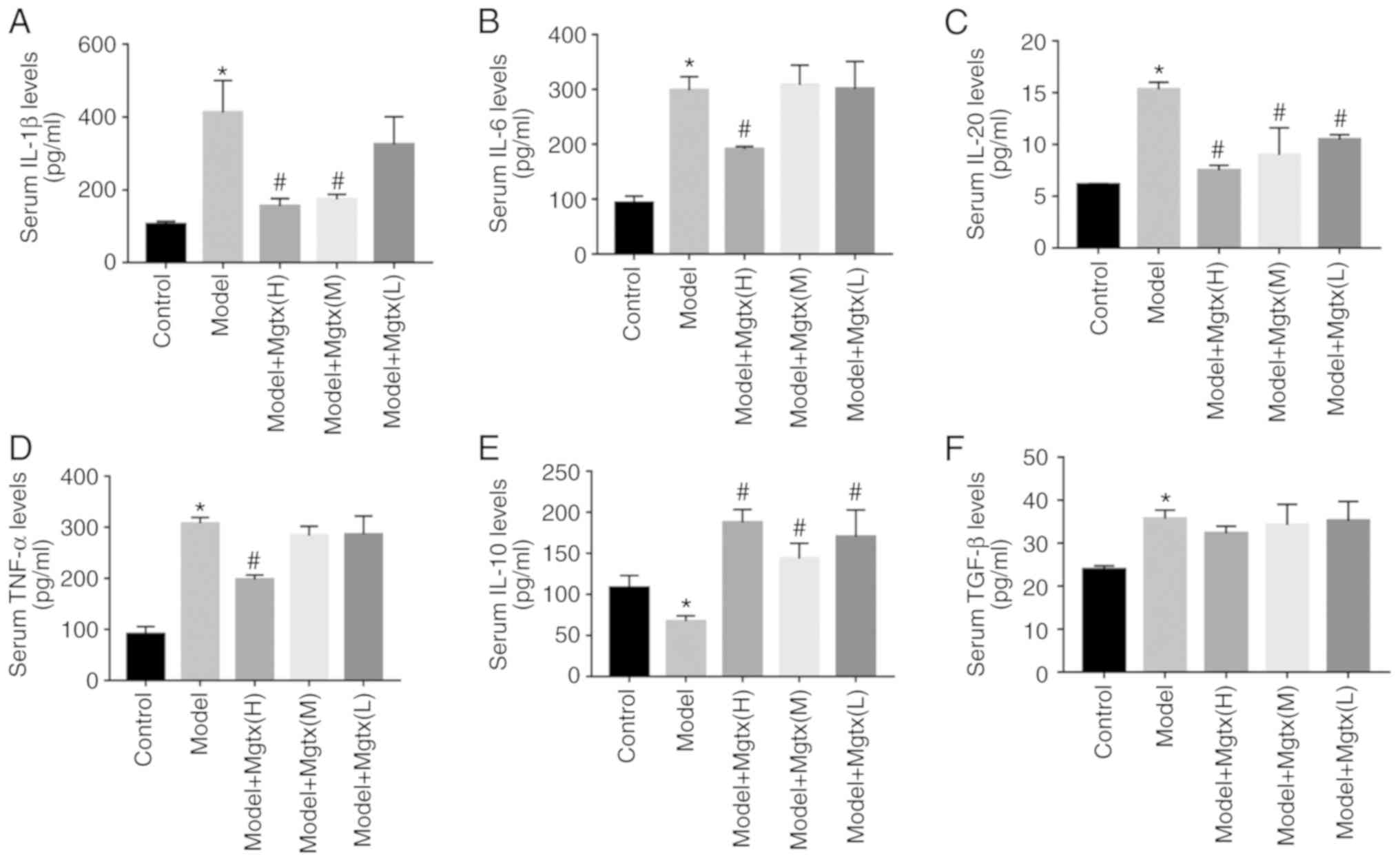 | Figure 4MgTX regulates cytokine secretion
in vivo. A total of 3 different MgTX concentrations were
injected into mice via intraperitoneal injection. The cytokines in
mouse serum were measured using ELISA kits, MgTX(H):0.41 mg/Kg,
MgTX(M):0.14 mg/Kg, MgTX(L):0.04 mg/Kg. (A-F) ELISAs to detect the
levels of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6 and IL-20)
in-mouse serum. Cytokine levels of IL-10 were inhibited compared
with the HF model group, whereas these levels were increased
compared with the HF model group treated with MgTX. The cytokine
levels of TGF-β group (n=6) increased in the model group, while
there was no significant difference observed in the HF model group
treated with MgTX. *P<0.05 vs. control group;
#P<0.05 vs. model group. n=6. MgTX, margatoxin;
TNF-α, tumor necrosis factor-α; IL, interleukin; TGF-β,
transforming growth factor-β; HF, hepatic fibrosis. |
MgTX regulates macrophage polarization in
vitro
To examine whether MgTX can regulate macrophage
polarization, M1 macrophages were induced using LPS (1
µg/ml) and IFN-γ (10 ng/ml) for 24 h, and M2 macrophages
were induced using IL-4 (15 ng/ml) for 24 h. RT-qPCR indicated that
treatment of M1 macrophages with MgTX (10 nM) decreased the mRNA
expression of M1-related markers (iNOS, CCL2, TNF-α and IL-1β),
while the mRNA expression levels of M2-related markers (IL-10,
CD163, Arg-1 and Mrc2) were upregulated by MgTX in M2 macrophages
(Fig. 5A-H). The primer sequences
are presented in Table I (5′-3′).
The results of western blot analysis were consistent with the
results of RT-qPCR (Fig. 5I-Q).
The results revealed that Kv1.3 contributed to the regulation of
macrophage polarization.
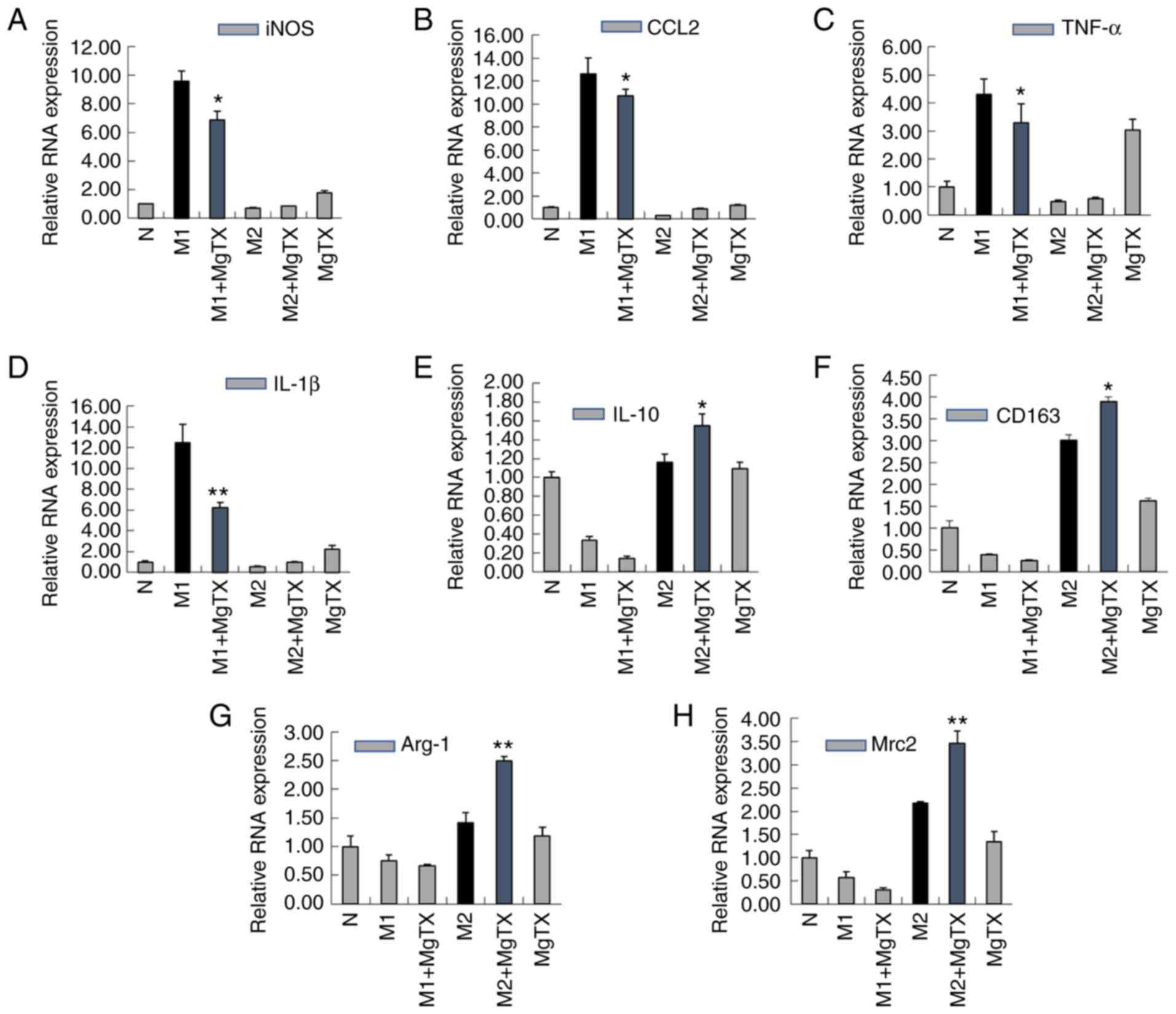 | Figure 5MgTX regulates macrophage
polarization in vitro. Reverse transcription-quantitative
PCR and western blot analysis were performed to detect the
expression of M1 markers and M2 markers in RAW264.7 cells. (A-H)
mRNA expression levels of iNOS, CCL2, TNF-α and IL-1β were
downregulated by MgTX in M1 phenotype macrophages. mRNA expression
levels of IL-10, CD163, Arg-1, MAC-2 were upregulated in M2
phenotype macrophages treated with MgTX compared with M2 phenotype
macrophages. (I-Q) protein expression levels of M1 markers (iNOS,
CCL2, TNF-α and IL-1β) and protein expression levels of M2 markers
(IL-10, CD163, Arg-1, Mrc2) were measured by western blot analysis.
*P<0.05, **P<0.01, M1 vs. M1 + MgTX
group, M2 vs. M2 + MgTX group. control group (N), n=3. MgTX,
margatoxin; CCL2, C-C motif chemokine ligand 2; TNF-α, tumor
necrosis factor-α; IL, interleukin; CD, cluster of
differentiation. |
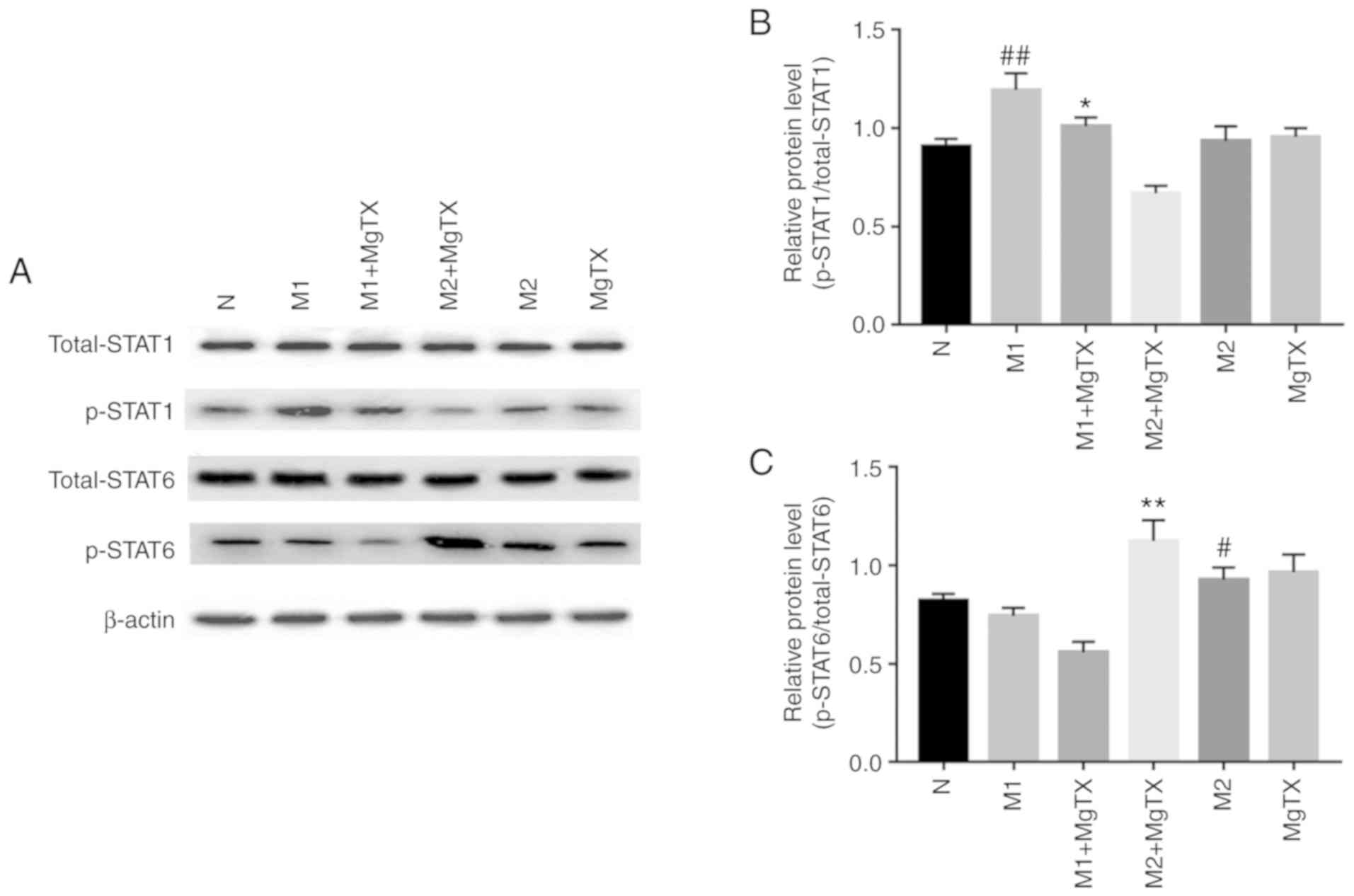
Kv1.3 regulates macrophage polarization
through the STAT1/STAT6 pathway
The underlying mechanisms through which MgTX
contributes to macrophage polarization was subsequently
investigated. Previous reports have indicated that the activation
of STAT1 signaling pathways, by IFNs and/or TLR signaling, can
switch macrophage toward the M1 phenotype, additionally, the
activation of STAT6 by IL-4 and/or IL-13 has been demonstrated to
switch macrophage function toward the M2 phenotype (29). In this study, the results of
western blot analysis revealed that the phosphorylation of STAT1
was significantly increased in M1 macrophages, and was
downregulated following MgTX (10 nM) treatment. Additionally, the
phosphorylation of STAT6 was significantly increased in M2
macrophages, and was upregulated by MgTX treatment (Fig. 6). These results indicated that
MgTX may regulate macrophage polarization through the STAT1/STAT6
signaling pathway.
Discussion
Fibrogenesis is a biochemical process that
represents damage in the majority of chronic diseases of the liver.
The activation of HSCs is an important pathophysiological mechanism
in the initiation, progression and regression of liver fibrosis. A
number of studies have identified crosstalk, which is at the level
of hepatic microcirculation, between the HSCs, sinusoidal
endothelial cells and KCs (30).
Studies on animal models and human studies have indicated that HF
is a reversible disease. The regression of HF can be possible in
the early stages of the condition, and partial and prolonged
recovery occurs in the late or advanced stages. The success of HF
treatment can increase survival and decrease the occurrence of
liver cirrhosis and hepatocellular carcinoma (31-36). Macrophages play a crucial role in
the occurrence and resolution of liver fibrosis. It has been well
established that infiltrating macrophages derived from monocytes
and KCs play a key role in the initiation and resolution of HF,
and, in mouse models of chronic liver injury, it has been
demonstrated that hepatic macrophages can promote the activation
and survival of HSCs (37,38).
Furthermore, a subtype of restorative macrophages has been
indicated to promote the resolution of fibrosis in chronic liver
injury (5,39). The current study demonstrated that
MgTX can protect liver cells from CCl4-induced HF injury. The
present study also explored whether MgTX attenuates liver fibrosis
through the regulation of cytokine secretion and the polarization
of macrophages.
The results of the current study indicated that MgTX
decreased the expression of HF associated proteins in mice.
Proteins of the MMP family are associated with the breakdown of the
extracellular matrix (ECM) during physiological processes. MMP12
and MMP13 are important proteins that are associated with the ECM.
Macrophages exhibit numerous functions in fibrogenesis, and
specific subtypes promote fibrosis resolution through enhanced ECM
degradation and by increasing MMP12 and MMP13 expression (40,41). In the current study, the
expression levels of MMP12 and MMP13 were increased in the HF
model, and this was due to an excess of ECM that requires
degradation. The results of the current study demonstrated that
MMP12 and MMP13 expression decreased in CCl4-exposed mice treated
with MgTX. POSTN is a 90-kDa secretory matricellular protein that
can regulate a number of effects during inflammation, wound healing
and tissue injury, and regulates key molecules of the fibrotic
matrix, including fibronectin and type I collagen via its EMILIN
domain, to promote fibrosis degradation (42,43). The results of the present study
indicated that POSTN was significantly downregulated by MgTX in the
HF model compared with the model group. MMP12, MM13 and POSTN are
associated with the progression of ECM and collagen. The results
revealed that MgTX decreased the expression of MMP12, MMP13 and
POSTN, in the CCl4-exposed group, attenuating HF. Exposure to CCl4
led to the elevation of liver enzymes, such as ALT and AST;
however, there was no significant change in the liver fibrosis
model with MgTX treatment. Additionally, a number of studies have
indicated that the M1 phenotype cells play an important role in
promoting HF (29,44). In this study, in RAW264.7 cell
lines, the expression of proteins associated with HF were decreased
in M1 phenotype macrophages treated with MgTX, which exerts an
anti-fibrotic effect in the progressive stage.
A therapeutic strategy for the treatment of HF is to
inhibit inflammatory monocyte infiltration. Kv1.3 plays a critical
role in monocyte migration, and the inhibition of Kv1.3 can inhibit
monocyte chemotaxis and monocyte infiltration into injury brain
(45). In the current study,
Kv1.3 was demonstrated to be associated with macrophage migration
in acute liver injury; Kv1.3 was indicated to downregulate the
expression of δ-catenin to inhibit macrophage migration. According
to another study by our group (unpublished data), RNA-seq profiling
analysis revealed that MgTX treatment downregulated δ-catenin.
According to wound healing and Transwell assays, δ-catenin
overexpression in RAW264.7 cells promoted migration, which was
suppressed upon silencing of δ-catenin. Another study revealed that
monocyte recruitment following light-induced cell death was
associated with the spatial-temporal expression of CCL2, and the
suppression of CCL2 in Müller cells decreased the recruitment of
monocytes/microglia (46).
Additionally, previous studies have revealed that macrophages may
promote HSC migration via CCL2 secretion, which can regulate the
crosstalk between cholangiocytes and portal fibroblasts (47,48). The inhibition of macrophage
migration infilter into the liver can reduce the immunological
reaction; thus, in this study a decrease in the expression of
Kv1.3, δ-catenin and CCL2 in the mice with CCl4-induced injury
treated with MgTX was observed. On the whole, the current study
indicated that MgTX inhibited macrophage migration to attenuate HF
in mice.
HF typically results from an inflammatory process
that affects hepatocytes or biliary cells. Inflammation leads to
the activation of effector cells, which results in the deposition
of ECM. Cytokines are released from inflammatory cells and play an
important role in HF as a key effector. The TGF-β cascade plays a
major role in fibrogenesis due to TGF-β being a potent stimulator
of the synthesis of extracellular matrix proteins (49). Pro-inflammatory cytokines,
including IL-6, IL-1β and TNF-α, are released from innate immune
cells and promote fibrogenesis via active HSCs. A recent study
demonstrated that the treatment of mice with recombinant IL-20
increased the expression of the pro-fibrotic cytokines, TGF-β and
TNF-α, and enhanced collagen synthesis in the liver; treatment with
neutralizing antibodies against IL-20 or IL20RA was revealed to
ameliorate the CCl4-induced HF in mice (50). Cytokines of the IL-10 family exert
protective effects, and IL-10 has been identified to exhibit a
suppressive effect, which inhibits the innate and adaptive immune
responses and prevents increased exacerbations. Therefore, IL-10
plays a protective role in the prevention of tissue injury in
chronic fibroproliferative diseases. Additionally, this has been
demonstrated in wound healing assays of scar formation in IL-10
knockout mice (51). The results
of the current study demonstrated that MgTX decreased the serum
levels of IL-6, IL-1β, IL-20 and TNF-α, increased the IL-10 serum
levels, but did not affect the TGF-β levels significantly. These
results indicated that MgTX regulated cytokine secretion in
macrophages; however, this effect was decreased in HSCs. Thus, HF
can be attenuated by MgTX through the regulation of the serum
levels of a number of cytokines.
A variety of macrophage subtypes release a number of
cytokines and exhibit functions in the pathophysiological process.
The M1 and M2 phenotype polarization of macrophages may directly
influence the outcome in chronic hepatic injury (52). In this study, RT-qPCR and western
blot analysis indicated that the levels of M1-related markers were
downregulated by MgTX, and that the levels of M2-related markers
were upregulated by MgTX. A number of studies have indicated that
STATs are crucial factors in M1 and M2 macrophage polarization
(53,54). p-STAT1 is required to promote M1
type macrophage activation in the presence of IFN-γ from innate
lymphocytes (55). By contrast,
p-STAT6 is an important regulator of M2 type macrophage
polarization in the presence of IL-13 or IL-4 during the TH2
cell-mediated immune responses (56,57). Therefore, the current study
examined the effects of MgTX on STAT signaling during macrophage
polarization. The phosphorylation and protein expression of p-STAT1
was markedly decreased in M1 macrophages treated with MgTX, and the
phosphorylation levels of p-STAT6 were increased in M2 macrophages
treated with MgTX. Therefore, MgTX was indicated to regulate
macrophage polarization via the STAT signaling pathway.
Furthermore, the phenotype of macrophages can be switched by MgTX;
thus, it would be of interest to examine the migratory ability of
macrophages in future studies.
HF, which is caused by a number of etiologies, is a
common pathological characteristic of end-stage hepatic diseases.
Advanced fibrosis can lead to cirrhosis, and currently, there is no
effective anti-fibrotic treatment that has been approved for human
use, at least to the best of our knowledge. Thus, the
identification of novel therapeutic options with which to prevent
the progression of liver fibrosis is urgently required. The current
study demonstrated that MgTX can ameliorate HF by regulating
macrophage polarization and cytokine secretion. These results
provide novel information on treatment options for the prevention
and reversion of liver fibrosis. However, further studies are
required to examine whether MgTX may be an effective candidate for
the treatment of advanced-stage fibrosis or cirrhosis and whether
MgTX may be a novel candidate for the treatment of HF or other
immunological diseases.
Acknowledgments
Not applicable.
Funding
The current study was supported by a grant from the
National Natural Science Foundation of China (grant no.
81500473).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
JL and YHL conceived and designed the experiments.
BMW and JDL performed the experiments. BMW analyzed the data and
wrote the paper. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal protocols were approved by the Animal
Care and Use Committee of Anhui Medical University, China.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mederacke I, Hsu CC, Troeger JS, Huebener
P, Mu X, Dapito DH, Pradere JP and Schwabe RF: Fate tracing reveals
hepatic stellate cells as dominant contributors to liver fibrosis
independent of its aetiology. Nat Commun. 4:28232013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zheng Z, Xu X, Zhang X, Wang A, Zhang C,
Hüttemann M, Grossman LI, Chen LC, Rajagopalan S, Sun Q and Zhang
K: Exposure to ambient particulate matter induces a NASH-like
phenotype and impairs hepatic glucose metabolism in an animal
model. J Hepatol. 58:148–154. 2013. View Article : Google Scholar
|
|
3
|
Czaja AJ and Carpenter HA: Progressive
fibrosis during corticosteroid therapy of autoimmune hepatitis.
Hepatology. 39:1631–1638. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schuppan D and Kim YO: Evolving therapies
for liver fibrosis. J Clin Invest. 123:1887–1901. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Duffield JS, Forbes SJ, Constandinou CM,
Clay S, Partolina M, Vuthoori S, Wu S, Lang R and Iredale JP:
Selective depletion of macrophages reveals distinct, opposing roles
during liver injury and repair. J Clin Invest. 115:56–65. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ramachandran P and Iredale JP:
Macrophages: Central regulators of hepatic fibrogenesis and
fibrosis resolution. J Hepatol. 56:1417–1419. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chawla A, Nguyen KD and Goh YP:
Macrophage-mediated inflammation in metabolic disease. Nat Rev
Immunol. 11:738–749. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mosser DM and Edwards JP: Exploring the
full spectrum of macrophage activation. Nat Rev Immunol. 8:958–969.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gundra UM, Girgis NM, Gonzalez MA, San
Tang M, Van Der Zande HJP, Lin JD, Ouimet M, Ma LJ, Poles J,
Vozhilla N, et al: Vitamin A mediates conversion of
monocyte-derived macrophages into tissueresident macrophages during
alternative activation. Nat Immunol. 18:642–653. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Borthwick LA, Suwara MI, Carnell SC, Green
NJ, Mahida R, Dixon D, Gillespie CS, Cartwright TN, Horabin J,
Walker A, et al: Pseudomonas aeruginosa induced airway epithelial
injury drives fibroblast activation: A mechanism in chronic lung
allograft dysfunction. Am J Transplant. 16:1751–1765. 2016.
View Article : Google Scholar :
|
|
11
|
Gordon S and Martinez FO: Alternative
activation of macrophages: Mechanism and functions. Immunity.
32:593–604. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Murray PJ, Allen JE, Biswas SK, Fisher EA,
Gilroy DW, Goerdt S, Gordon S, Hamilton JA, Ivashkiv LB, Lawrence
T, et al: Macrophage activation and polarization: Nomenclature and
experimental guidelines. Immunity. 41:14–20. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sica A, Invernizzi P and Mantovani A:
Macrophage plasticity and polarization in liver homeostasis and
pathology. Hepatology. 59:2034–2042. 2014. View Article : Google Scholar
|
|
14
|
Heymann F, Trautwein C and Tacke F:
Monocytes and macrophages as cellular targets in liver fibrosis.
Inflamm Allergy Drug Targets. 8:307–318. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Murray PJ and Wynn TA: Protective and
pathogenic functions of macrophage subsets. Nat Rev Immunol.
11:723–737. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chiaramonte MG, Donaldson DD, Cheever AW
and Wynn TA: An IL-13 inhibitor blocks the development of hepatic
fibrosis during a T-helper type 2-dominated inflammatory response.
J Clin Invest. 104:777–785. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kaviratne M, Hesse M, Leusink M, Cheever
AW, Davies SJ, McKerrow JH, Wakefield LM, Letterio JJ and Wynn TA:
IL-13 activates a mechanism of tissue fibrosis that is completely
TGF-beta independent. J Immunol. 173:4020–4029. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
McKenzie GJ, Bancroft A, Grencis RK and
McKenzie AN: A distinct role for interleukin-13 in
Th2-cell-mediated immune responses. Curr Biol. 8:339–342. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
McKenzie GJ, Fallon PG, Emson CL, Grencis
RK and McKenzie AN: Simultaneous disruption of interleukin (IL)-4
and IL-13 defines individual roles in T helper cell type 2-mediated
responses. J Exp Med. 189:1565–1572. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mackenzie AB, Chirakkal H and North RA:
Kv1.3 potassium channels in human alveolar macrophages. Am J
Physiol Lung Cell Mol Physiol. 285:L862–L868. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kazama I, Maruyama Y, Murata Y and Sano M:
Voltage-dependent biphasic effects of chloroquine on delayed
rectifier K(+)-channel currents in murine thymocytes. J Physiol
Sci. 62:267–274. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Toldi G, Bajnok A, Dobi D, Kaposi A,
Kovács L, Vásárhelyi B and Balog A: The effects of Kv1.3 and IKCa1
potassium channel inhibition on calcium influx of human peripheral
T lymphocytes in rheumatoid arthritis. Immunobiology. 218:311–316.
2013. View Article : Google Scholar
|
|
23
|
Beeton C, Wulff H, Standifer NE, Azam P,
Mullen KM, Pennington MW, Kolski-Andreaco A, Wei E, Grino A, Counts
DR, et al: Kv1.3 channels are a therapeutic target for T
cell-mediated autoimmune diseases. Proc Natl Acad Sci USA.
103:17414–17419. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chi V, Pennington MW, Norton RS, Tarcha
EJ, Londono LM, Sims-Fahey B, Upadhyay SK, Lakey JT, Iadonato S,
Wulff H, et al: Development of a sea anemone toxin as an
immu-nomodulator for therapy of autoimmune diseases. Toxicon.
59:529–546. 2012. View Article : Google Scholar
|
|
25
|
Kazama I, Maruyama Y, Endo Y, Toyama H,
Ejima Y, Matsubara M and Kurosawa S: Overexpression of delayed
rectifier K(+) channels promotes in situ proliferation of
leukocytes in rat kidneys with advanced chronic renal failure. Int
J Nephrol. 2012:5815812012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shao PP, Liu CJ, Xu Q, Zhang B, Li SH, Wu
Y, Sun Z and Cheng LF: Eplerenone reverses cardiac fibrosis via the
suppression of tregs by inhibition of Kv1.3 channel. Front Physiol.
9:8992018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Moreno C, Prieto P, Macías Á,
Pimentel-Santillana M, de la Cruz A, Través PG, Boscá L and
Valenzuela C: Modulation of voltage-dependent and inward rectifier
potassium channels by 15-epi-lipoxin-A4 in activated murine
macrophages: Implications in innate immunity. J Immunol.
191:6136–6146. 2003. View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
29
|
Yang Y, Wu XQ, Li WX, Huang HM, Li HD, Pan
XY, Li XF, Huang C, Meng XM, Zhang L, et al: PSTPIP2 connects DNA
methylation to macrophage polarization in CCL4-induced mouse model
of hepatic fibrosis. Oncogene. 37:6119–6135. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bitto N, Liguori E and La Mura V:
Coagulation, microenvironment and liver fibrosis. Cells. 7:pii:
E85. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang KK and Czaja AJ: Hepatocellular
carcinoma in corticosteroid-treated severe autoimmune chronic
active hepatitis. Hepatology. 8:1679–1683. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nishiguchi S, Kuroki T, Nakatani S,
Morimoto H, Takeda T, Nakajima S, Shiomi S, Seki S, Kobayashi K and
Otani S: Randomised trial of effects of interferon-alpha on
incidence of hepatocellular carcinoma in chronic active hepatitis C
with cirrhosis. Lancet. 346:1051–1055. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Roberts SK, Therneau TM and Czaja AJ:
Prognosis of histological cirrhosis in type 1 autoimmune hepatitis.
Gastroenterology. 110:848–857. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Singal AG, Volk ML, Jensen D, Di Bisceglie
AM and Schoenfeld PS: A sustained viral response is associated with
reduced liver-related morbidity and mortality in patients with
hepatitis C virus. Clin Gastroenterol Hepatol. 8:280–288.
288.e12010. View Article : Google Scholar
|
|
35
|
Lok AS, Everhart JE, Wright EC, Di
Bisceglie AM, Kim HY, Sterling RK, Everson GT, Lindsay KL, Lee WM,
Bonkovsky HL, et al: Maintenance peginterferon therapy and other
factors associated with hepatocellular carcinoma in patients with
advanced hepatitis C. Gastroenterology. 140:840–849. 2011.
View Article : Google Scholar :
|
|
36
|
Bruix J, Poynard T, Colombo M, Schiff E,
Burak K, Heathcote EJ, Berg T, Poo JL, Mello CB, Guenther R, et al:
Maintenance therapy with peginterferon alfa-2b does not prevent
hepatocellular carcinoma in cirrhotic patients with chronic
hepatitis C. Gastroenterology. 140:1990–1999. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Krenkel O and Tacke F: Liver macrophages
in tissue homeostasis and disease. Nat Rev Immunol. 17:306–321.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Seki E and Schwabe RF: Hepatic
inflammation and fibrosis: Functional links and key pathways.
Hepatology. 61:1066–1079. 2015. View Article : Google Scholar :
|
|
39
|
Baeck C, Wei X, Bartneck M, Fech V,
Heymann F, Gassler N, Hittatiya K, Eulberg D, Luedde T, Trautwein C
and Tacke F: Pharmacological inhibition of the chemokine C-C motif
chemo-kine ligand 2 (monocyte chemoattractant protein 1)
accelerates liver fibrosis regression by suppressing Ly-6C(+)
macrophage infiltration in mice. Hepatology. 59:1060–1072. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ramachandran P, Pellicoro A, Vernon MA,
Boulter L, Aucott RL, Ali A, Hartland SN, Snowdon VK, Cappon A,
Gordon-Walker TT, et al: Differential Ly-6C expression identifies
the recruited macrophage phenotype, which orchestrates the
regression of murine liver fibrosis. Proc Natl Acad Sci USA.
109:E3186–E3195. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Madala SK, Pesce JT, Ramalingam TR, Wilson
MS, Minnicozzi S, Cheever AW, Thompson RW, Mentink-Kane MM and Wynn
TA: Matrix metalloproteinase 12-deficiency augments extracellular
matrix degrading metalloproteinases and attenuates IL-13-dependent
fibrosis. J Immunol. 184:3955–3963. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Conway SJ, Izuhara K, Kudo Y, Litvin J,
Markwald R, Ouyang G, Arron JR, Holweg CT and Kudo A: The role of
periostin in tissue remodeling across health and disease. Cell Mol
Life Sci. 71:1279–1288. 2014. View Article : Google Scholar :
|
|
43
|
Mosher DF, Johansson MW, Gillis ME and
Annis DS: Periostin and TGF-β-induced protein: Two peas in a pod?
Crit Rev Biochem Mol Biol. 50:427–439. 2015.
|
|
44
|
Zhao XA, Chen G, Liu Y, Chen Y, Wu H,
Xiong Y, Wang G, Jia B, Li Y, Xia J, et al: Curcumin reduces Ly6Chi
monocyte infiltration to protect against liver fibrosis by
inhibiting Kupffer cells activation to reduce chemokines secretion.
Biomed Pharmacother. 106:868–878. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Eder C: Ion channels in monocytes and
microglia/brain macrophages: Promising therapeutic targets for
neurological diseases. J Neuroimmunol. 224:51–55. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Rutar M, Natoli R and Provis JM: Small
interfering RNA-mediated suppression of Ccl2 in Müller cells
attenuates microglial recruitment and photoreceptor death following
retinal degeneration. J Neuroinflammation. 9:2212012. View Article : Google Scholar
|
|
47
|
Pellicoro A, Ramachandran P, Iredale JP
and Fallowfield JA: Liver fibrosis and repair: Immune regulation of
wound healing in a solid organ. Nat Rev Immunol. 14:181–194. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kruglov EA, Nathanson RA, Nguyen T and
Dranoff JA: Secretion of MCP-1/CCL2 by bile duct epithelia induces
myofibroblastic transdifferentiation of portal fibroblasts. Am J
Physiol Gastrointest Liver Physiol. 290:G765–G771. 2006. View Article : Google Scholar
|
|
49
|
Rockey DC, Bell PD and Hill JA: Fibrosis-a
common pathway to organ injury and failure. N Engl J Med.
372:1138–1149. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chiu YS, Wei CC, Lin YJ, Hsu YH and Chang
MS: IL-20 and IL-20R1 antibodies protect against liver fibrosis.
Hepatology. 60:1003–1014. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Sziksz E, Pap D, Lippai R, Béres NJ,
Fekete A, Szabó AJ and Vannay Á: Fibrosis related inflammatory
mediators: Role of the IL-10 cytokine family. Mediators Inflamm.
2015:7646412015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Galastri S, Zamara E, Milani S, Novo E,
Provenzano A, Delogu W, Vizzutti F, Sutti S, Locatelli I, Navari N,
et al: Lack of CC chemokine ligand 2 differentially affects
inflammation and fibrosis according to the genetic background in a
murine model of steatohepatitis. Clin Sci (Lond). 123:459–471.
2012. View Article : Google Scholar
|
|
53
|
Khan Z, Cao DY, Giani JF, Bernstein EA,
Veiras LC, Fuchs S, Wang Y, Peng Z, Kalkum M, Liu GY and Bernstein
KE: Overexpression of the C-domain of angiotensin-converting enzyme
reduces melanoma growth by stimulating M1 macrophage polarization.
J Biol Chem. 294:4368–4380. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ding N, Wang Y, Dou C, Liu F, Guan G, Wei
K, Yang J, Yang M, Tan J, Zeng W and Zhu C: Physalin D regulates
macrophage M1/M2 polarization via the STAT1/6 pathway. J Cell
Physiol. 234:8788–8796. 2019. View Article : Google Scholar
|
|
55
|
Darnell JE Jr, Kerr IM and Stark GR:
Jak-STAT pathways and transcriptional activation in response to
IFNs and other extracellular signaling proteins. Science.
264:1415–1121. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Herbert DR, Hölscher C, Mohrs M, Arendse
B, Schwegmann A, Radwanska M, Leeto M, Kirsch R, Hall P, Mossmann
H, et al: Alternative macrophage activation is essential for
survival during schistosomiasis and downmodulates T helper 1
responses and immunopathology. Immunity. 20:623–635. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Brombacher F, Arendse B, Peterson R,
Hölscher A and Hölscher C: Analyzing classical and alternative
macrophage activation in macrophage/neutrophil-specific IL-4
receptor-alpha-deficient mice. Methods Mol Biol. 531:225–252. 2009.
View Article : Google Scholar : PubMed/NCBI
|