Introduction
Allergic asthma is a common chronic inflammatory
airway disease. It affects >300 million individuals worldwide
with increasing prevalence and incidence (1,2).
It poses a serious global health concern, resulting in heavy
economic burdens on societies worldwide (3). The main pathological characteristics
of asthma are airway hyper-responsiveness (AHR), the infiltration
of inflammatory cells and mucus overproduction. In addition,
long-term airway inflammation leads to structural alterations in
the airways.
Type 2 inflammation is one of the major molecular
mechanisms underlying susceptibility to asthma. Type 2 immune
responses are characteristic of allergic asthma in the lower
airways. Airway type 2 immune responses are mainly mediated by
eosinophils, basophils, mast cells, Th2 cells, group 2 innate
lymphoid cells (ILC2s) and IgE-producing B cells (4,5).
Type 2 inflammation is an inflammatory pathway involving a
subpopulation of CD4+ T cells. Imbalances between
T-helper type 1 (Th1) and T-helper type 2 (Th2) immune reactions
are well known to be an underlying factor. In particular, Th2 cells
and their cytokines play essential roles in the development of
allergic airway inflammation in asthma (6). Briefly, following exposure to
allergens, the airway epithelium initiates inflammatory reactions
involving major pro-inflammatory factors, such as IL-33, IL-25 and
thymic stromal lymphopoietin (TSLP), which are cytokines that
regulate maturation of CD4+ T cells into Th2 cells
(7,8). Moreover, Th2 cells release large
amounts of Th2 cytokines, such as IL-4, IL-5 and IL-13, while
decreasing the production of Th1 cytokines. Type 2 cytokines drive
the cellular events in response to the allergen stimulation of the
epithelium. These events include activation of the airway
epithelial cells, chemoattraction of effector cells (mast cells,
eosinophils, and basophils), and remodeling of the epithelium and
subepithelial stroma. Such inflammatory responses result in
maladaptive changes in the airways, causing pathological reactions
to inhaled irritants (9).
IL-33 is a nuclear cytokine belonging to the IL-1
family and is primarily expressed in epithelial and endothelial
cells. IL-33 acts as an initiating signal for the inflammatory
responses and induces the release of Th2-type cytokines, triggering
Th2-cell-mediated immune responses. Recent studies have indicated
that the IL-33 level is elevated in asthmatic patients (10). In addition to this clinical
observation, IL-33 overproduction has been observed in the lungs of
mice in a model of severe influenza-induced asthma exacerbation.
This animal model recapitulates all the key features of asthma
observed in humans, including the physiological response to
treatment with corticosteroids (11).
Allergic asthma immune responses are initiated by
the interactions of allergens with epithelial cells, resulting in
the release of the cytokines, TSLP, IL-25 and IL-33. IL-25 and
IL-33 induce the release of IL-13 and IL-5 from IL-25R+
natural helper cells. IL-33 additionally induces the release of
IL-4 from basophils. These responses initiate a pathogenic cascade
that leads to the development of asthma in susceptible patients.
IL-33 initiates the inflammatory signal transduction pathways via
its receptor, ST2, which is mainly expressed in ILC2s, Th2 cells,
mast cells, eosinophils, and NK cells (12-14). Recent studies have reported that
IL-33 represents a potential link between the airway epithelium and
Th2-type inflammatory responses (15-17). Thus, IL-33 appears to be closely
involved in the development of asthma. A clinical study
demonstrated that the IL-33 level was significantly elevated in the
bronchial epithelia of asthmatic patients (18). Accordingly, it has been considered
one of the indices of asthma severity (19,20).
The present study investigated the effects of
osthole against asthma in mice and it was determined that it
ameliorates asthma through the IL-33/ST2 pathway. Currently, the
main therapeutic asthma medications include inhaled corticosteroids
(ICS), β2-agonists and leukotriene modifiers among several others
(21). ICS can effectively manage
the symptoms of mild to moderate asthma. However, a subgroup of
asthma patients chronically treated with ICS gradually become less
sensitive, or even resistant, to glucocorticoid. Consequently, the
patient becomes susceptible to exaggerated inflammatory
responses.
Chinese medicine is currently a major interest in
the identification of candidate targets for combinatorial therapy
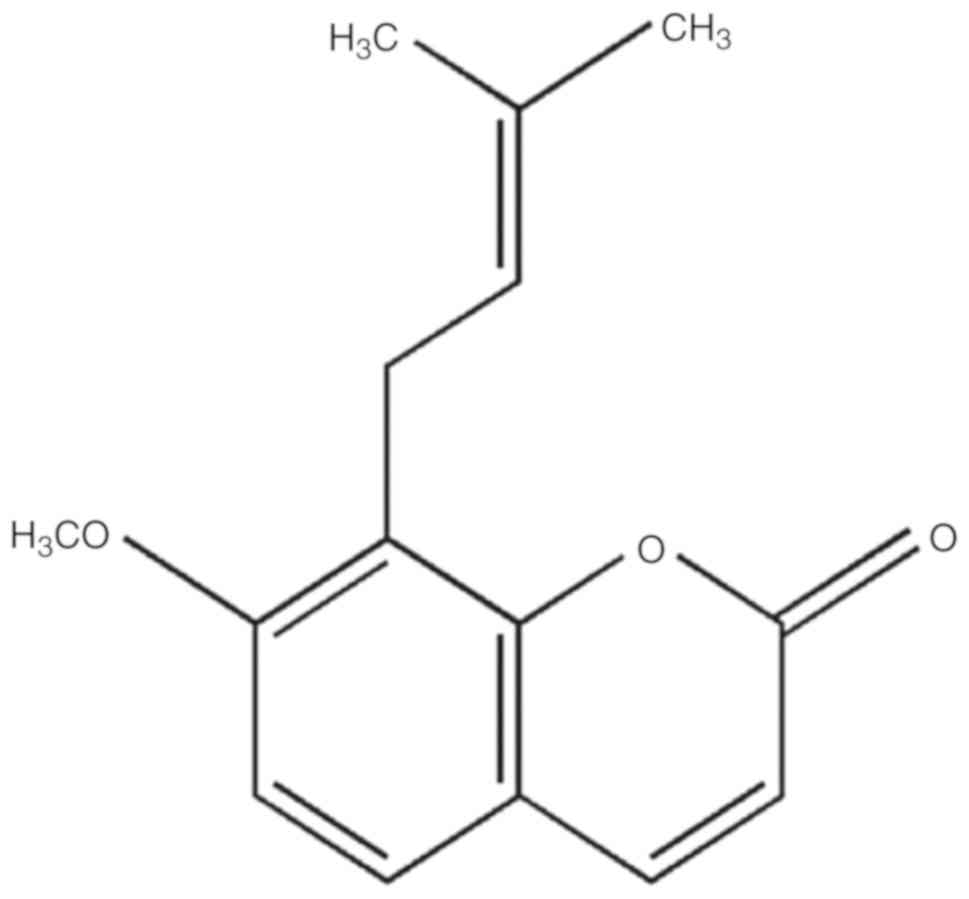
against the pathogenesis of allergic reactions (22,23). Osthole (chemical structure shown
in Fig. 1) is a natural compound
found in the Chinese herb Cnidii Fructus (She Chuang Zi). This herb
has a long history of use for asthma in China. Previous studies
have indicated that osthole has a variety of pharmacological and
biological activities, including anti-allergic, anti-inflammatory,
anti-apoptotic, anti-bacterial and antioxidant stress activi-ties
(24-26). Additionally, animal studies have
demonstrated that osthole can alleviate immune-mediated
inflammatory diseases, such as autoimmune encephalomyelitis, IgA
nephropathy, and contact dermatitis (27-29). In line with these physiological
responses, osthole has been found to inhibit expression of eotaxin,
an IL-4 induced eosinophil-specific C-C chemokine, in bronchial
epithelial cells (30).
Therefore, it is suggested that osthole affects
IL-33 expression, whereby it regulates the immune response cascade
downstream of IL-33, thus providing a possible new treatment option
for symptomatic relief medication for allergic asthma. In the
present study, the anti-asthmatic effects of osthole in asthmatic
mice were investigated.
Materials and methods
Reagents
Osthole (C15H16O3;
molecular weight, 244.29; purity, >98%, Fig. 1) was obtained from Winherb.
Ovalbumin (OVA; grade V), aluminum hydroxide [Al(OH)3],
dexamethasone (DEX), methacholine (Mch) and pentobarbital sodium
were purchased from Sigma-Aldrich; Merck KGaA. TRIzol reagent and
phosphate-buffered saline (PBS) were obtained from Invitrogen;
Thermo Fisher Scientific, Inc. SYBR-Green and the PrimeScript 1st
Strand cDNA Synthesis kit were supplied by Takara Biotechnology
Co., Ltd. The Luminex multiplex cytokine analysis kit was purchased
from Merck KGaA. The DAB kit was obtained from Boster Biological
Technology Co., Ltd.
Experimental animals and protocol
Female BALB/c mice (n=40; 6 weeks old, weighing
18-20 g) were purchased from Shanghai Xi Puer-Bikai Experimental
Animal Co., Ltd. They were housed in a barrier environment free of
specific pathogens, and maintained at 22±2°C with 12-h light/dark
cycles and free access to standard laboratory food and water. The
mice were used for the experiments after 7 days of acclimatization.
The experiments were carried out in accordance with the Guide for
the Care and Handling of Laboratory Animals of the Institutional
Animal Care, and the Ethics Committee of Shanghai Municipal
Hospital of TCM, Shanghai University of TCM.
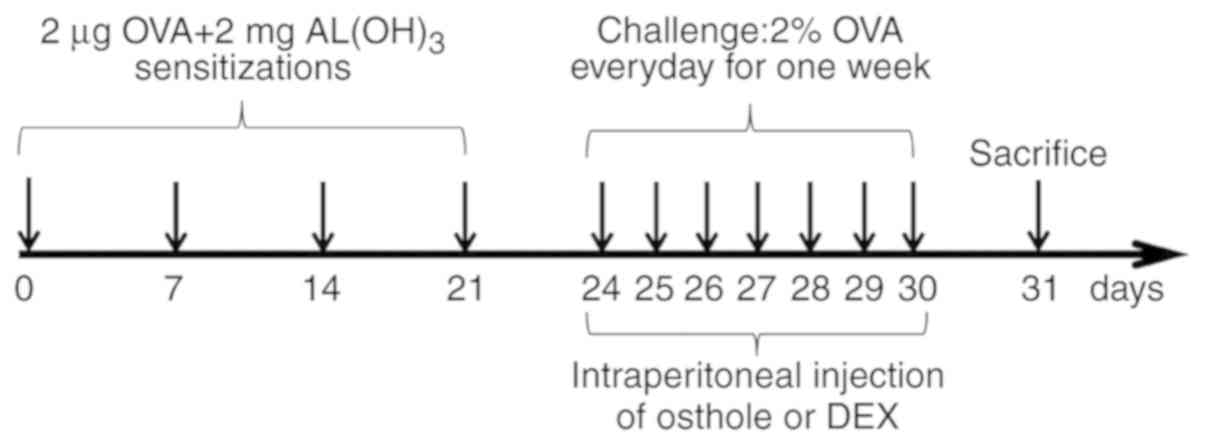
Sensitization and treatment protocol
The BALB/c mice were randomly divided into 5 groups
of 8 mice in each as follows: A normal control (NC) group, an
OVA-sensitized and challenged asthma model (asthma) group, a
dexamethasone (1 mg/kg) treatment (DEX) group, and 2 osthole
treatment groups at doses of 25 and 50 mg/kg (OS-25 and OS-50).
Apart from the mice in the NC group, the mice in all the other
groups were sensitized by an intraperitoneal injection of 20
µg OVA together with 2 mg of Al(OH)3 in 200
µl of PBS on days 1, 7, 14 and 21. The mice were then
sprayed with 2% OVA (w/v in PBS) for 30 min each day from day 24 to
day 30, as previously described (31). Briefly, the mice were placed in a
transparent box that was connected to the atomization tube of an
ultrasonic nebulizer, and atomized with OVA (402AI, Jiangsu Yuyue
Medical Instrument Co., Ltd.). PBS was used instead of OVA for the
NC group. From days 24 to 30, the mice in the drug intervention
groups were intraperitoneally administered osthole (25 or 50 mg/kg)
or dexamethasone (1 mg/kg) 1 h prior to the OVA atomization.
Osthole and DEX treatments were administered daily where indicated.
The mice in the NC and asthma groups were administered PBS only at
the same volume used for drug administration. Subsequently, the
animals were anesthetized by an intraperitoneal injection of
pentobarbital sodium (50 mg/kg) at 24 h after the final challenge
for AHR detection. A schematic diagram of the airway inflammation
induction is presented in Fig.
2.
AHR detection
Bronchial hyperreactivity was measured 24 h after
the final challenge. AHR was detected using an invasive, mouse
pulmonary function testing system (Buxco Electronics Inc.). The
entire procedure was performed on anesthetized mice. First, the
mice were anesthetized with pentobarbital sodium (50 mg/kg,
intraperitoneal injection), followed by tracheotomy, after which a
suitable cannula was inserted. The mice were then placed in the
plethysmograph chamber, and the ventilator-assisted mouse breathing
tube was opened while increasing the airway pressure baseline with
<5% change every 2.5 min until it tended to stabilize.
Subsequently, PBS and 3 concentrations of Mch (3.125, 6.25 and 12.5
mg/ml) were introduced into the head chamber via nebulization. The
airway responses were displayed through changes in airway
resistance (RL) and lung dynamic compliance (Cdyn) (32). The dose-dependent effect of
osthole (25 and 50 mg/kg) on airway resistance in asthmatic mice
was evaluated by statistical analysis. If the inhibitory effect
increased with increasing osthole concentration, the inhibitory
effect of osthole on airway resistance was considered to be
dose-dependent (33-35).
Collection of bronchoalveolar lavage
fluid (BALF)
After the airway reactivity test was completed, the
mice were euthanized by cervical dislocation, and death was
confirmed on the basis of lack of breathing, pulse, pupillary
reflex, corneal reflex and response to firm toe pinching. The lungs
were lavaged through the tracheal cannula 3 times with 0.8 ml cold
PBS. The BALF was then collected and centrifuged at 500 × g for 10
min at 4°C. The supernatants were transferred into new tubes and
stored at -80°C. The cell pellets were resuspended in PBS to
evaluate the abundance of inflammatory cells, including
lymphocytes, eosinophils, basophils and neutrophils. The cytokine
(INF-γ, IL-4, IL-5, IL-13 and IL-33) levels in BALF were analyzed
by Luminex assay following the manufacturer's instructions.
Histopathology of lung tissue assay
The right middle lung lobes were removed from the
mice and fixed with 4% paraformaldehyde. First, the lungs were
dehydrated with various concentrations of alcohol. They were then
embedded in paraffin and sectioned at a thickness of 3 µm.
Subsequently, the sections were stained with hematoxylin and eosin
(H&E; cat. no. AR1180; Boster Biological Technology, Ltd.) for
5 min, periodic acid-Schiff reagent (PAS; cat. no. BA4080B; BaSO
Biotech) for 10 min, and Masson's trichrome (M-T; cat. no. BA4079B;
BaSO Biotech) for 10 min at room temperature. The lung tissue
sections were examined at ×200 magnification using an optical
microscope (ECLIPSE 80i, Nikon Corp.). Finally, lung inflammation
was assessed by the degree of eosinophil infiltration, the amount
of mucus secretion and the area of collagen deposition.
qPCR assay
Reverse transcription-quantitative PCR (RT-qPCR) was
performed to determine the levels of the following Th1/Th2-related
cytokines in the lungs: IFN-γ, IL-4, IL-5, IL-13, IL-33 and ST2.
Total RNA was extracted from the lungs using TRIzol reagent. A
reverse transcriptase kit was used to perform first-strand cDNA
synthesis according to the manufacturer's protocol. The cDNA
samples were then amplified using an ABI 7500 sequence detector
with thermal cycling conditions of 30 sec at 95°C (hot-start step),
followed by 40 cycles of 5 sec at 95°C (denaturation step), 60 sec
at 60°C (annealing step) and 60 sec at 72°C (extension step).
Relative mRNA expression was analyzed using the 2−ΔΔCq
method (36). β-actin was used as
the internal control gene. The sequences of the primers used for
mouse β-actin, IL-4, IL-5, IL-13, IFN-γ, IL-33 and ST2 are
presented in Table I.
 | Table IPrimer sequences of the genes
detected by quantitative polymerase chain reaction. |
Table I
Primer sequences of the genes
detected by quantitative polymerase chain reaction.
| Gene | Primer sequences
(5′-3′) |
|---|
| β-actin | F:
5′-CCTCTATGCCAACACAGT-3′ |
| R:
5′-AGCCACCAATCCACACAG-3′ |
| IL-4 | F:
5′-TAGTTGTCATCCTGCTCTTCTT-3′ |
| R:
5′-CTCACTCTCTGTGGTGTTCTTC-3′ |
| IL-5 | F:
5′-CCATTGCCCACTCTGTAC-3′ |
| R:
5′-AGGCTTCCTGTCCCTACT-3′ |
| IL-13 | F:
5′-CAGCCTCCCCGATACCAAAAT-3′ |
| R:
5′-CCCCAGCAAAGTCTGATGTGA-3′ |
| IFN-γ | F:
5′-CAGCGACCGTGTCTGTAT-3′ |
| R:
5′-GAGGAGCGTCTGGAAATA-3′ |
| IL-33 | F:
5′-GTCAACAGACGCAGCAAA-3′ |
| R:
5′-TTAGGAAAGAACCCACGAA-3′ |
| ST2 | F:
5′-CTGGCACTGCATTTCCT-3′ |
| R:
5′-GCCTACGAGCAGGAGATT-3′ |
Immunohistochemistry (IHC)
ST2 expression in the lungs was evaluated by IHC
according to the manufacturer's instructions (ProteinTech Group,
Inc.). First, the lung sections were deparaffinized and washed 3
times with PBS. They were then incubated in 3% hydrogen peroxide
for 30 min. Subsequently, they were washed with PBS, and incubated
with rabbit polyclonal anti-ST2 antibody (1:100; cat. no.
11920-1-AP, ProteinTech Group, Inc.) for 12 h at 4°C in a humid
chamber. The sections were washed with PBS, covered with
biotinylated goat anti-rabbit (1:300; cat. no. SA1022, Wuhan Boster
Biological Technology, Ltd.), and incubated for 60 min at room
temperature. Once the incubation was completed, they were washed
again with PBS, incubated in a peroxidase solution for 20 min at
room temperature, and then washed again with PBS. Finally, the
sections were covered with 3,3-diaminobenzidine tetrahydrochloride
(DAB) chromogen and incubated for 3-10 min at room temperature
before being observed under a light microscope (ECLIPSE 80i, Nikon
Corp.) at ×200 magnification.
Statistical analysis
All data are presented as the means ± standard
deviation. Data analysis was performed using SPSS version 20.0 (IBM
SPSS, Inc.). The statistical graphs were created using GraphPad
Prism 6.0 software (GraphPad Software, Inc.). Results were
evaluated statistically using one-way analysis of variance (ANOVA)
and the Bonferroni post hoc test. Values of P<0.05 and P<0.01
were considered to indicate statistically significant and highly
statistically significant differences, respectively.
Results
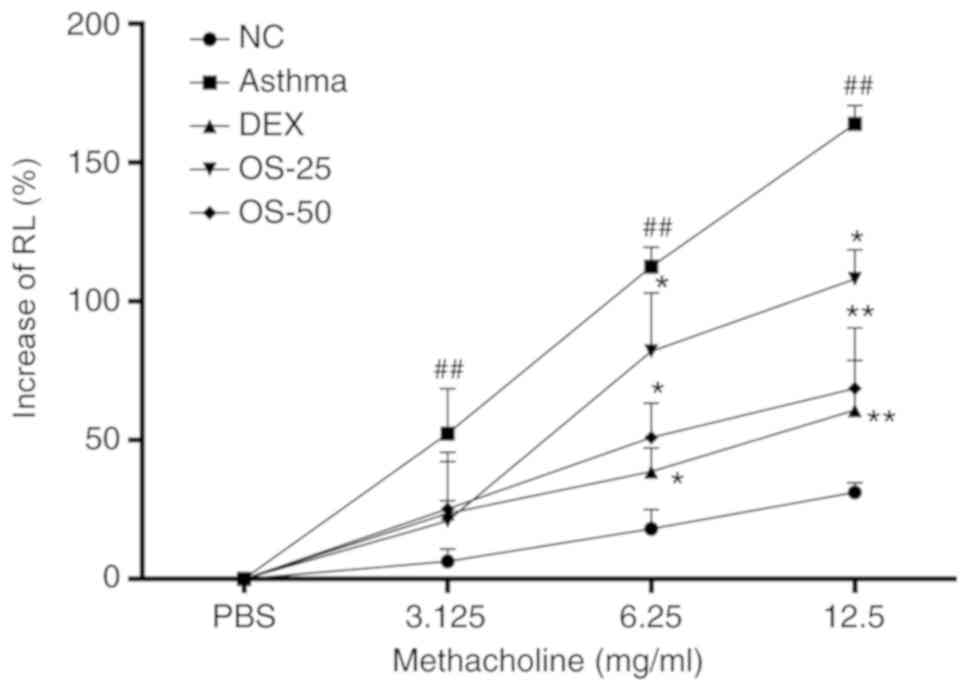
Osthole reduces AHR in asthmatic
mice
The effects of osthole on airway responses in a
mouse model of asthma was determined by nebulization with Mch. A
higher level of airway resistance was observed in the mice with
OVA-induced asthma compared with that observed in the normal
control group (P<0.05), whereas DEX and osthole treatment
significantly downregulated the OVA-induced AHR (P<0.05;
Fig. 3). In particular, osthole
exerted a more prominent effect at 50 mg/kg than that observed with
25 mg/kg.
Osthole attenuates airway inflammation in
asthmatic mice
Inflammatory cell infiltration is one of the most
prominent features of asthma. The total and differential cell
counts of BALF were estimated with an automated cell counter. The
numbers of total leukocytes and differential cells, including
neutrophils, eosinophils, lymphocytes, basophils and monocytes were
markedly increased in the asthma group compared with those of the
NC group (P<0.05; Fig. 4).
After the mice were treated with osthole or DEX, the numbers of
leukocytes and differential cells markedly decreased (P<0.05).
In addition, osthole treatment decreased the number of eosinophils
in a dose-dependent manner. It was clearly indicated that the
optimal dose of osthole with which to reduce the number of
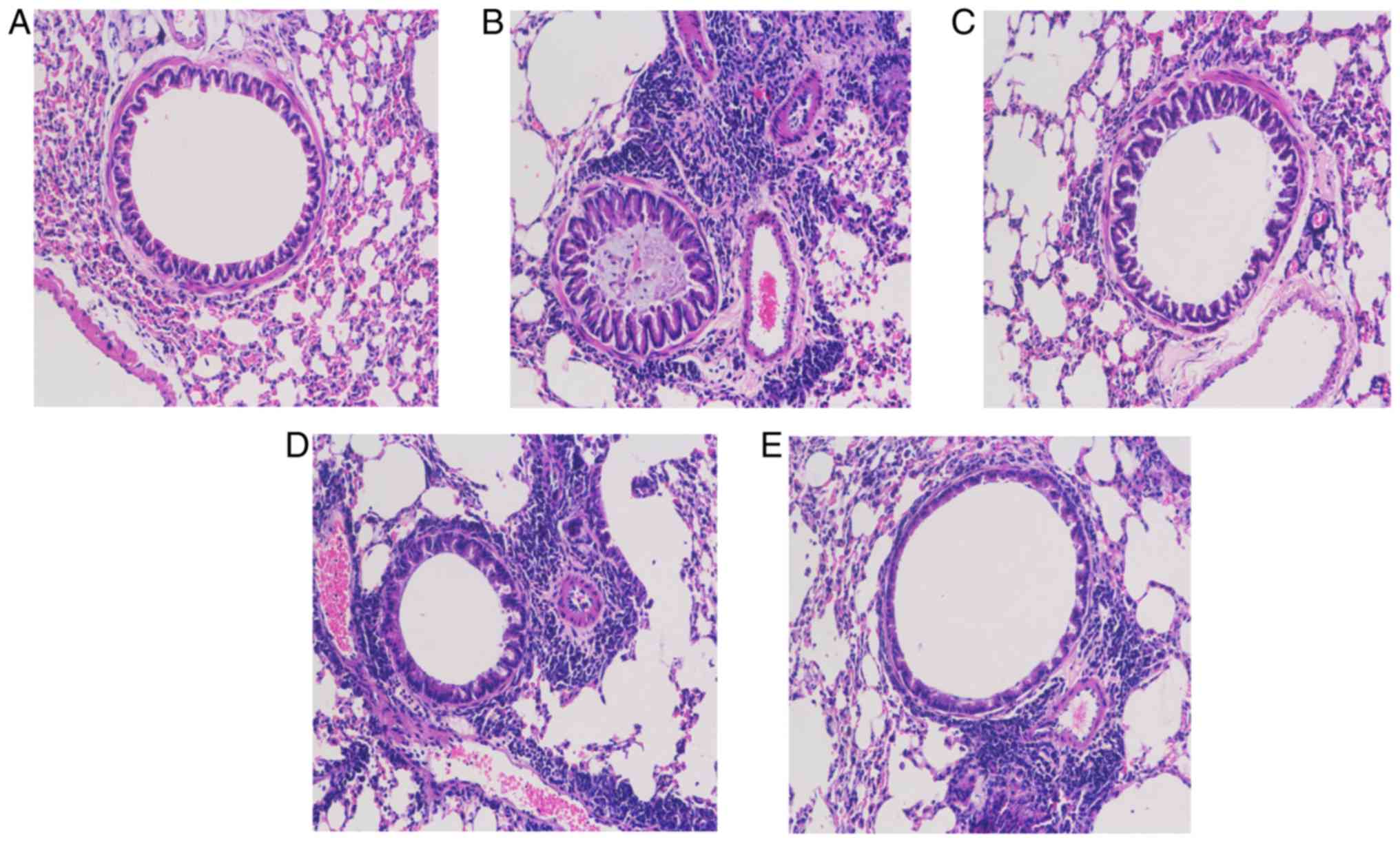
inflammatory cells was 50 mg/kg (P<0.05). H&E staining also
revealed that there were more inflammatory cells infiltrated in the
lungs of mice in the asthmatic group (Fig. 5B) compared with those in the lungs
of mice in the NC group (Fig.
5A). However, airway inflammation was alleviated in the OS and
DEX groups (Fig. 5C-E).
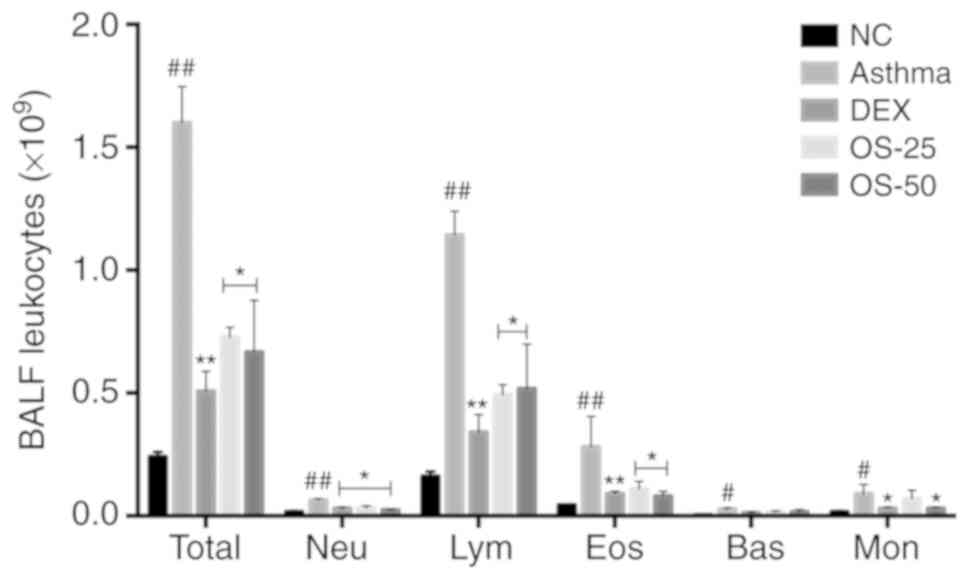 | Figure 4Osthole reduces the number of
inflammatory cells in the alveolar lavage fluid of asthmatic mice.
Total, leucocytes; eos, eosnophils; lym, lymphocytes; neu,
neutrophils; bas, basophils; mon, monocytes. The normal control
group (NC), OVA-sensitized and challenged asthma group (asthma),
dexamethasone (1 mg/kg) treatment group (DEX), and osthole
treatment groups at doses of 25 and 50 mg/kg (OS-25, OS-50). The
data are presented as the means ± SD. #P<0.05,
##P<0.01 vs. the normal control group;
*P<0.05, **P<0.01 vs. the asthma model
group. OVA, ovalbumin; DEX, dexamethasone. |
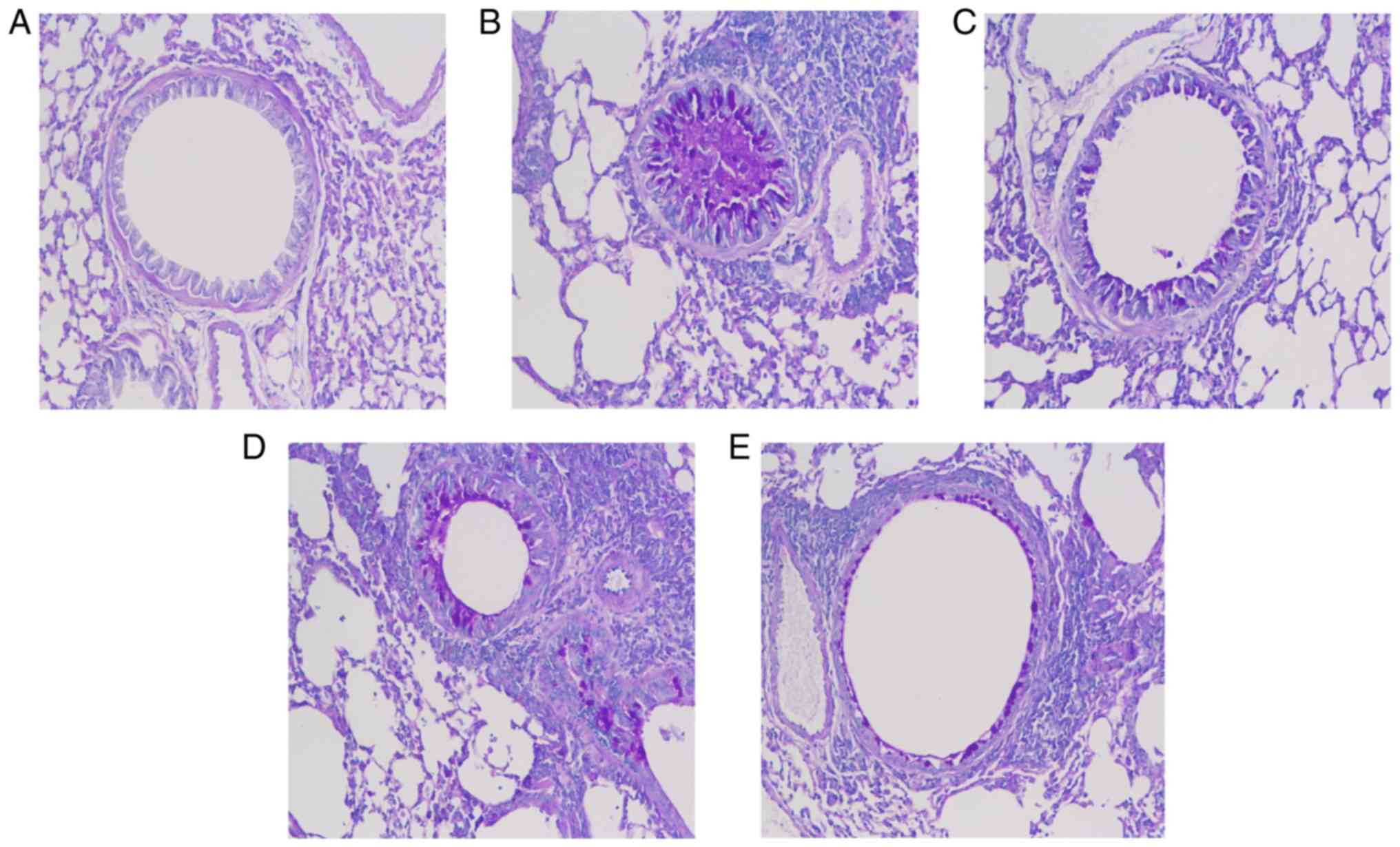
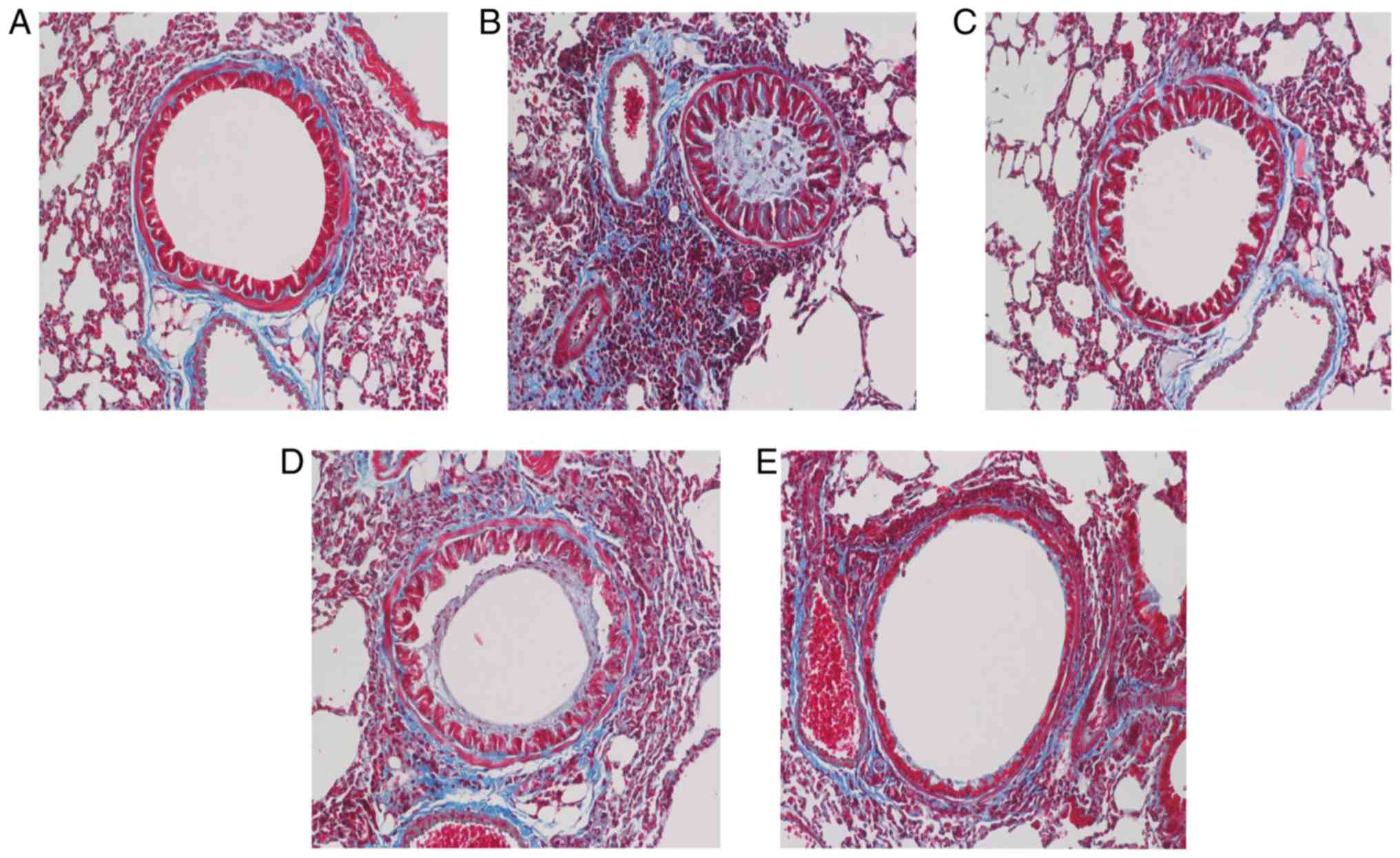
Osthole alleviates airway remodeling in
asthmatic mice
The degree of airway remodeling is associated with
the severity of asthma. In the present study, PAS and Masson's
trichrome staining were used to assess mucus secretion and collagen
deposition in addition to the presence of goblet cell hyperplasia
(Figs. 6 and 7). Compared with the NC group, the mice
in the asthma group exhibited increased mucus production, excessive
collagen deposition and goblet cell hyperplasia in the lungs
(Figs. 6A and B, and 7A and B), and these maladaptive changes
were suppressed in the DEX and OS groups (Figs. 6C-E and 7C-E). Of note, no marked differences
were observed between the 2 drug treatment groups (Fig. 7C-E).
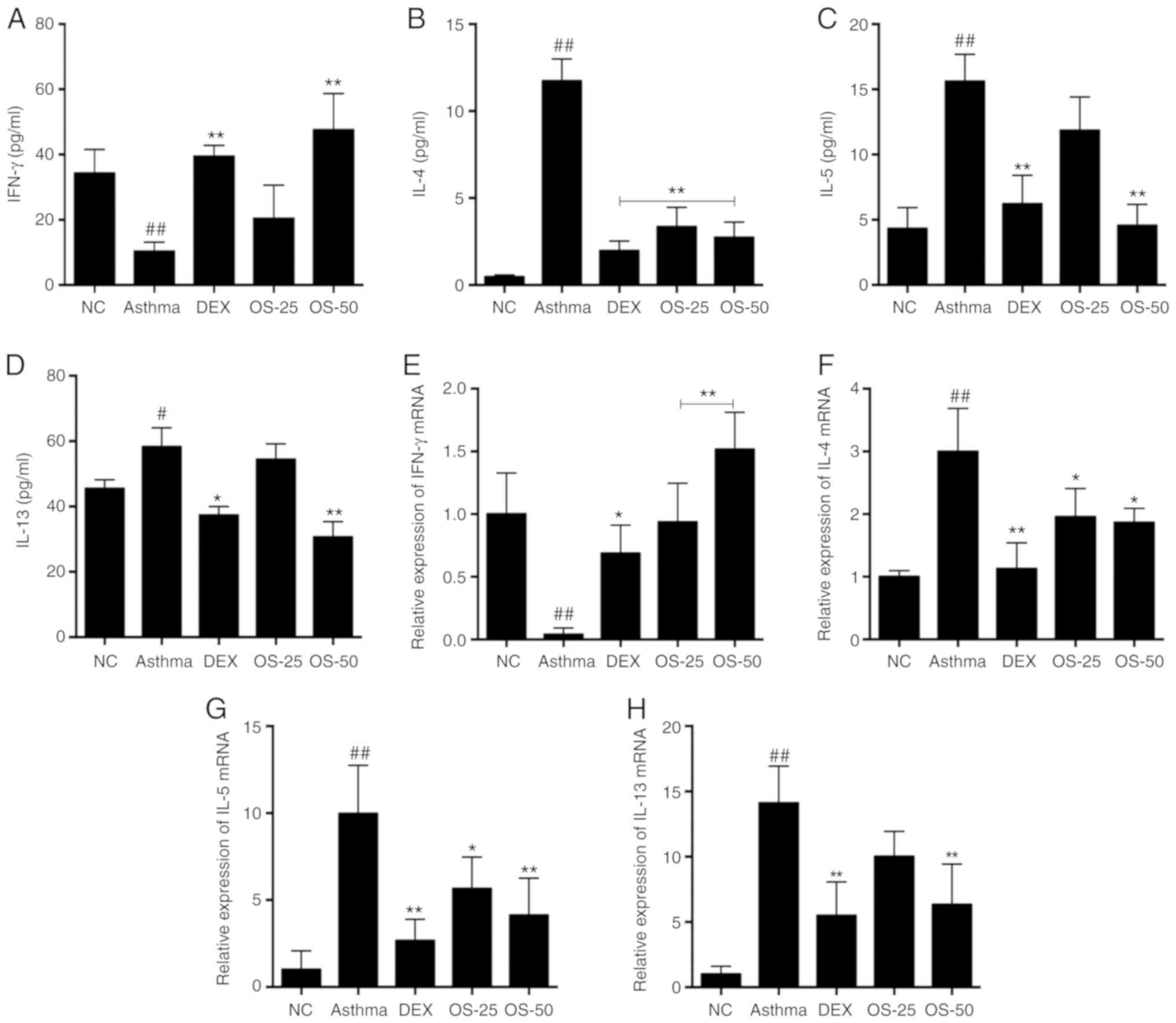
Osthole rebalances Th1/Th2-related
cytokines in the BALF and lungs of asthmatic mice
Subsequently, the present study evaluated the levels
of the Th1/Th2-related cytokines, IFN-γ, IL-4, IL-5 and IL-13, in
BALF using a Luminex multiplex cytokine analysis kit. The levels of
the Th2 cytokines, IL-4, IL-5 and IL-13, were significantly
elevated in the asthma group (P<0.05); however, these elevations
were markedly downregulated in the OS-50 and DEX groups (P<0.05;
Fig. 8B-D). At the same time, the
level of the Th1 cytokine, IFN-γ, was decreased in the asthma group
(P<0.05), while the DEX and OS groups exhibited a relatively
higher level of IFN-γ (P<0.05; Fig. 8A). Notably, the level of IFN-γ in
the OS-50 group was much higher than that in the DEX group
(Fig. 8A). Furthermore, the mRNA
levels of these cytokines were also evaluated in the lungs by
RT-qPCR. The IFN-γ mRNA levels in the OS and DEX groups were higher
than those in the asthma group (P<0.05), while the IL-4, IL-5
and IL-13 mRNA levels were lower in the OS and DEX groups
(P<0.05). Notably, the effect of osthole (50 mg/kg) on IFN-γ
mRNA expression was much more prominent than that of DEX (Fig. 8E).
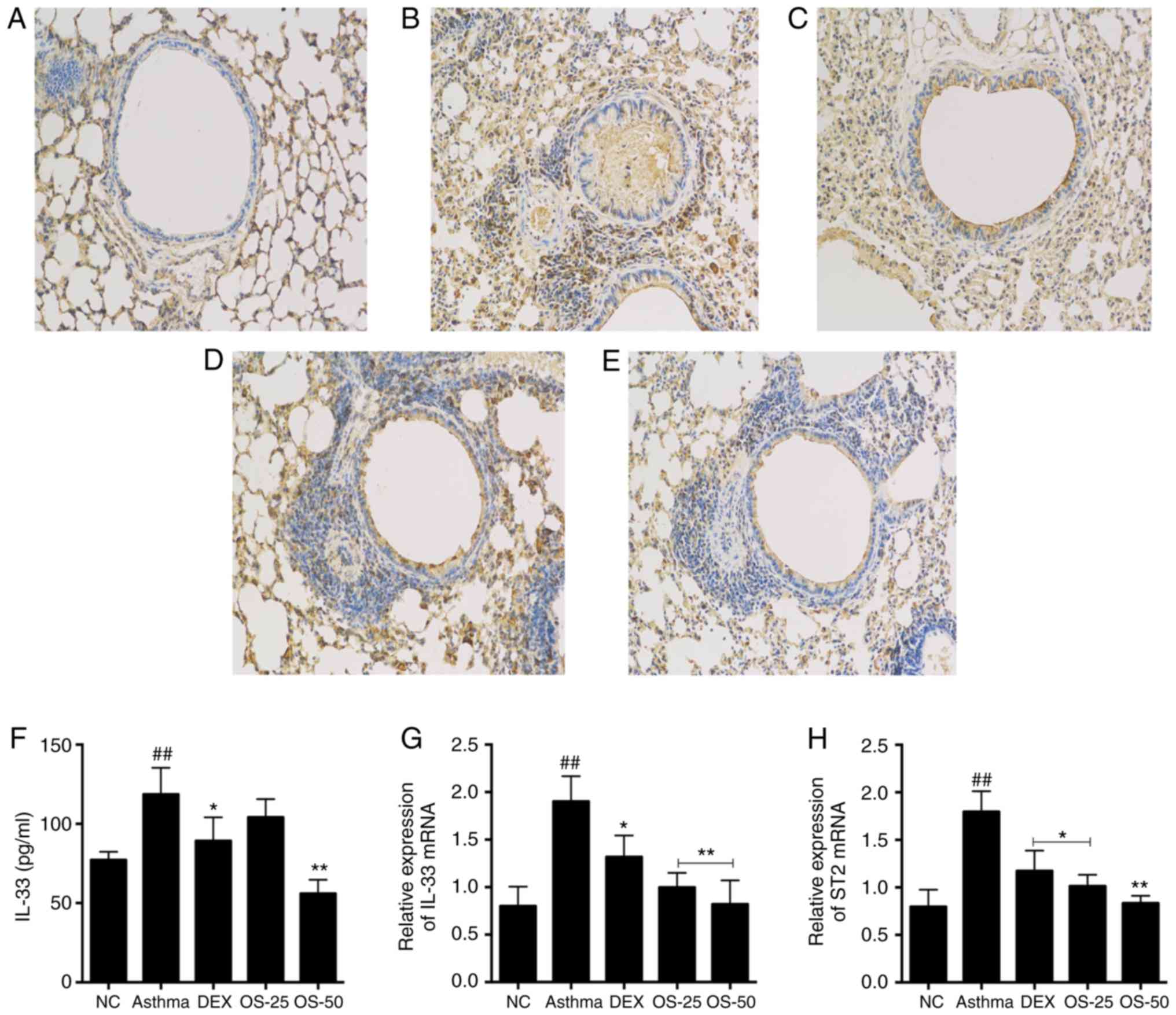
Osthole inhibits IL-33/ST2 signaling in
asthmatic mice
IL-33/ST2 signaling is involved in airway
inflammation in asthma. The level of IL-33 in BALF was assessed
using the Luminex multiplex cytokine analysis kit. Moreover, the
protein level of ST2 was evaluated by IHC. Furthermore, the mRNA
levels of IL-33 and ST2 in the lungs were quantified by RT-qPCR.
The levels of IL-33 in BALF of in the asthma group were higher than
those in the control group (P<0.05; Fig. 9F). On the other hand, the OS-50
and DEX groups exhibited significantly lower levels of IL-33 than
the asthma group.
In terms of gene expression, there was a significant
upregulation in the IL-33 and ST2 mRNA levels in the asthma group
compared with the NC group (P<0.05). Of note, the DEX and OS
groups exhibited significantly lower mRNA levels of IL-33 and ST2
compared with the asthma group (P<0.05; Fig. 9G and H). Moreover, the effect of
osthole treatment appeared to be dose-dependent, since the IL-33
and ST2 mRNA levels were markedly lower in the OS-50 group relative
to those in the OS-25 group. Furthermore, the IHC analysis of the
lung sections demonstrated that the highest ST2 protein expression
was observed in the asthma group. By constast, the DEX and OS-50
groups exhibited lower amounts of ST2 protein (Fig. 9A-E).
Discussion
AHR is a representative hallmark of allergic asthma.
It can be induced by exposure to allergens, such as OVA and house
dust mite (HDM) (37-39). Osthole is a natural compound
isolated from the Chinese herb, Cnidii Fructus, which has been
widely used for centuries in China for the treatment of allergic
disorders. In the present study, it was demonstrated for the first
time, at least to the best of our knowledge, that osthole treatment
can markedly decrease AHR in a mouse model of asthma.
Allergic asthma is a chronic airway inflammatory
disorder. It is considered to manifest from the immune responses of
Th2 cells, including innate lymphoid cells (ILC2s), CD4+
Th2 cells, eosinophils, neutrophils and mast cells. These
maladaptive immune responses are executed via the overproduction of
the IL-4, IL-5 and IL-13 cytokines. IL-4 and IL-5 promote the
recruitment of inflammatory cells. For example, IL-5 stimulates the
generation and activation of eosinophils. In the present study, the
types of inflammatory cells in BALF were analyzed. It was observed
that the number of leucocytes, particularly eosinophils, was
markedly elevated in the asthmatic mice as was expected, confirming
that the asthmatic model in the present study recapitulated the
major features of asthma observed in humans. Notably, the
osthole-treated asthmatic mice exhibited lower amounts of
leucocytes, including eosinophils, indicating that osthole
treatment reduced the number of inflammatory cells. Furthermore, it
was found that the levels of IL-4, IL-5 and IL-13 were increased in
the BALF of asthmatic mice, while the level of IFN-γ was decreased.
Osthole treatment partially suppressed the imbalance between these
Th1- and Th2-related cytokines. IL-13 has been reported to induce
smooth muscle cell proliferation and hypertrophy, goblet cell
metaplasia, the overproduction of mucus and excessive collagen
deposition (40). The present
study also observed that the asthmatic mice had significantly less
mucus and goblet cells when treated with osthole.
Epithelial cells (ECs) are the first line of defense
against pathogens. They also interact with a various types of
immune cells that trigger immune responses against foreign
pathogens. Most importantly, ECs have been reported to affect Th2
responses by secreting the pro-inflammatory cytokines, IL-33, IL-25
and TSLP (41). IL-33 is a newly
discovered member of the IL-1 family (42). It is mainly located in the cell
nucleus in addition to its extra-cellular role as a cytokine
(43,44). Genome-wide association studies
have indicated that IL-33 is one of the genes involved in asthma
development (45,46). IL-33 is constitutively expressed
in epithelial, endothelial, and mast cells (47,48). IL-33 is rapidly released form
epithelial cells after allergen exposure or tissue injury (13,49). IL-33 can function as an alarm to
warn the immune system. IL-33 signals activated via its receptor
ST2, which is expressed on natural helper cells, ILC2s,
eosinophils, basophils, Th2 cells, mast cells and NK cells
(50,51). Previous studies have revealed that
IL-33/ST2 signaling is the fundamental Th2-associated immune
response (12,52,53). The inhibition of IL-33 expression
reduces the symptoms of asthma, such as inflammation, in asthmatic
mice (54). In the present study,
it was identified that IL-33 and ST2 levels were significantly
elevated in the lungs of asthmatic mice; however, this inflammatory
response was significantly suppressed by osthole treatment. Further
studies are required to determine exactly which cells mediate
IL-33/ST2 signaling and to elucidate the mechanisms through which
they interact with other cells.
Osthole has a long history of clinical application
in the treatment of dermatitis and eczema. Although it has a
variety of biological activities, data on its efficacy on allergic
asthma is limited. In the present study, the immunomodulatory
effects of osthole in asthmatic mice were investigated. Osthole did
not exert an adverse side-effects when administered at 25 or 50
mg/kg in mice. The results indicated that exposure to OVA induced
allergic asthma in mice and that osthole significantly reduced
airway resistance, and the infiltration of the above-mentioned
inflammatory cells. In summary, the data suggest that osthole can
alleviate allergen-induced airway inflammation in mice.
Th2-type cytokines play a key role in the
progression of allergic asthma and can promote the development of
inflammatory phenotypes. IL-4 is an important inducer of IgE
production by B lymphocytes, and IL-5 promotes eosinophil
activation, maturation and recruitment. IL-13 is more effective in
promoting AHR and pathological changes. IL-33 stimulates excessive
secretion of Th2 cytokines. The treatment of asthmatic mice with
high doses of osthole (50 mg/kg) significantly reduced the
expression of the Th2 cytokines, IL-4, IL-5 and IL-13. Overall,
these data indicate that osthole can attenuate OVA-induced Th2
responses and inhibit the immunopathology of allergic asthma in
mice. Based on the aforementioned information, treatment strategies
for the pain suppression of Th2 immunity may improve the control of
allergic asthma. Osthole improves the severity of allergic asthma
in mice by inhibiting the Th2 immune response. The present study
further elucidated whether this inhibitory effect was in part due
to IL-33 downregulation in osthole-treated mice. The results
revealed that IL-33 production was significantly decreased in the
lungs and BALF. Likewise, it was observed that the IL-33 ligand,
ST2, was also significantly downregulated in the lungs.
In the present study, OVA was used to induce asthma
in mice, whereby it was observed that osthole reduced the degree of
inflammation in asthma. In the future, the authors aim to evaluate
the effects of osthole on allergic asthma induced by other
allergens, such as HDM.
IL-33 has been reported to trigger the activation of
inflammatory pathways through an IL-1-related receptor protein
complex of ST2 (ST2/IL-1RAcP) (20). Since dendritic cells (DCs)
function as antigen-presenting cells, they play a central role in
immune responses observed in asthma. The activation of Toll-like
receptor 4 (TLR4) by a fungal proteinase derived from
Aspergillus oryzae has been reported to be involved in the
epithelial overexpression of IL-33 in the airways, subsequently
activating DCs (55). Moreover,
HDM extracts can directly activate ECs to produce IL-25 and IL-33
and act on DCs. Thus, ECs appear to modulate DC activation through
secretion of IL-33 (56). Given
this link between ECs and DCs, further studies are required to
investigate the role of DCs in the osthole-induced alleviation of
airway inflammation in asthmatic mice.
There are several other limitations to the present
study. It is not clear which cell types were affected by Osthole,
resulting in reduced airway inflammatory responses. Osthole may
play a regulatory role in airway epithelial cells and dendritic
cells, which in turn affects the Th2 immune response process,
leading to a reduction in airway inflammation. The intervention of
osthole in airway epithelial cells and DCs is the content of
follow-up experiments by the authors.
In conclusion, the results of the present study
indicate that osthole administration reduces the severe symptoms of
Th2-mediated asthma in mice, in part as it reduces the expression
of IL-33. The data demonstrate that osthole treatment suppresses
Th2 immune responses by inhibiting the production and activation of
IL-33 in the early stages of antigen stimulation. Furthermore, it
was determined that Th1 and Th2 cells were rebalanced presumably
through the inhibition of IL-33/ST2 signaling. Taken together, the
results indicate that osthole may be a promising candidate for the
development of an asthma medication.
Funding
The present study was funded by a grant from the
National Natural Science Foundation of China (grant no.
81573937).
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
QY, LK, WH, NM, XL, GW and LW were involved in the
conceptualization of the study. QY and LK performed the
experiments, contributed to data analysis and wrote the manuscript.
GW and LW conceptualized the study design. QY, LK, WH, NM and XL
contributed to data analysis and provided experimental materials.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the
Committee on the Ethics of Animal Experiments of Shanghai Municipal
Hospital of TCM, Shanghai University of TCM.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Dr Jinfeng Wu
(Huashan Hospital, Fudan University)for his critical reading of the
manuscript. The authors also gratefully acknowledge the expert
technical assistance provided by Zhonghua Wu (Shanghai University
of Traditional Chinese Medicine) and Xiaoming Wang (Shanghai
University of Traditional Chinese Medicine).
References
|
1
|
Bostantzoglou C, Delimpoura V, Samitas K,
Zervas E, Kanniess F and Gaga M: Clinical asthma phenotypes in the
real world: Opportunities and challenges. Breathe (Sheff).
11:186–193. 2015. View Article : Google Scholar
|
|
2
|
Galli SJ, Tsai M and Pilponsky AM: The
development of allergic inflammation. Nature. 454:445–454. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bateman ED, Hurd SS, Barnes PJ, Bousquet
J, Drazen JM, FitzGerald M, Gibson P, Ohta K, O'Byrne P, Pedersen
SE, et al: Global strategy for asthma management and prevention:
GINA executive summary. Eur Respir J. 31:143–178. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oliphant CJ, Barlow JL and McKenzie AN:
Insights into the initiation of type 2 immune responses.
Immunology. 134:378–385. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fahy JV: Type 2 inflammation in
asthma-present in most, absent in many. Nat Rev Immunol. 15:57–65.
2015. View
Article : Google Scholar :
|
|
6
|
Martinez FD and Vercelli D: Asthma.
Lancet. 382:1360–1372. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hwang YH, Paik MJ and Yee ST: Diisononyl
phthalate induces asthma via modulation of Th1/Th2 equilibrium.
Toxicol Lett. 272:49–59. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Platts-Mills TA: The allergy epidemics:
1870-2010. J Allergy Clin Immunol. 136:3–13. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Locksley RM: Asthma and allergic
inflammation. Cell. 140:777–783. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Smith DE: IL-33: A tissue derived cytokine
pathway involved in allergic inflammation and asthma. Clin Exp
Allergy. 40:200–208. 2010. View Article : Google Scholar
|
|
11
|
Ravanetti L, Dijkhuis A, Sabogal Pineros
YS, Bal SM, Dierdorp BS, Dekker T, Logiantara A, Adcock IM, Rao NL,
Boon L, et al: An early innate response underlies severe
influenza-induced exacerbations of asthma in a novel
steroid-insensitive and anti-IL-5-responsive mouse model. Allergy.
72:737–753. 2017. View Article : Google Scholar
|
|
12
|
Saluja R, Ketelaar ME, Hawro T, Church MK,
Maurer M and Nawijn MC: The role of the IL-33/IL-1RL1 axis in mast
cell and basophil activation in allergic disorders. Mol Immunol.
63:80–85. 2015. View Article : Google Scholar
|
|
13
|
Kaur D, Gomez E, Doe C, Berair R, Woodman
L, Saunders R, Hollins F, Rose FR, Amrani Y, May R, et al: IL-33
drives airway hyper-responsiveness through IL-13-mediated mast
cell: Airway smooth muscle crosstalk. Allergy. 70:556–567. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ahmed A and Koma MK: Interleukin-33
triggers B1 cell expansion and its release of monocyte/macrophage
chemoattractants and growth factors. Scand J Immunol. 82:118–124.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Iijima K, Kobayashi T, Hara K, Kephart GM,
Ziegler SF, McKenzie AN and Kita H: IL-33 and thymic stromal
lymphopoietin mediate immune pathology in response to chronic
airborne allergen exposure. J Immunol. 193:1549–1559. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gordon ED, Simpson LJ, Rios CL, Ringel L,
Lachowicz-Scroggins ME, Peters MC, Wesolowska-Andersen A, Gonzalez
JR, MacLeod HJ, Christian LS, et al Alternative splicing of
interleukin-33 and type 2 inflammation in asthma. Proc Natl Acad
Sci USA. 113:8765–8770. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hristova M, Habibovic A, Veith C,
Janssen-Heininger YMW, Dixon AE, Geiszt M and van der Vliet A:
Airway epithelial dual oxidase 1 mediates allergen-induced IL-33
secretion and activation of type 2 immune responses. J Allergy Clin
Immunol. 137:1545–1556. 2016. View Article : Google Scholar
|
|
18
|
Préfontaine D, Nadigel J, Chouiali F,
Audusseau S, Semlali A, Chakir J, Martin JG and Hamid Q: Increased
IL-33 expression by epithelial cells in bronchial asthma. J Allergy
Clin Immunol. 125:752–754. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo Z, Wu J, Zhao J, Liu F, Chen Y, Bi L,
Liu S and Dong L: IL-33 promotes airway remodeling and is a marker
of asthma disease severity. J Asthma. 51:863–869. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Savenije OE, Mahachie John JM, Granell R,
Kerkhof M, Dijk FN, de Jongste JC, Smit HA, Brunekreef B, Postma
DS, Van Steen K, et al: Association of IL33-IL-1 receptor-like 1
(IL1RL1) pathway polymorphisms with wheezing phenotypes and asthma
in childhood. J Allergy Clin Immunol. 134:170–177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Keenan CR, Salem S, Fietz ER, Gualano RC
and Stewart AG: Glucocorticoid-resistant asthma and novel
anti-inflammatory drugs. Drug Discov Today. 17:1031–1038. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
He XF, Pan WD, Yao YL and Zhang HM: Recent
highlights of Chinese herbs in treatment of allergic disease:
Acting via mitogen-activated protein kinase signal pathway. Chin J
Integr Med. 23:570–573. 2017. View Article : Google Scholar
|
|
23
|
Shao YY, Zhou YM, Hu M, Li JZ, Chen CJ,
Wang YJ, Shi XY, Wang WJ and Zhang TT: The anti-allergic rhinitis
effect of traditional Chinese medicine of shenqi by regulating mast
cell degranulation and Th1/Th2 cytokine balance. Molecules.
22:E5042017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li YQ, Wang JY, Qian ZQ, Li YL, Li WN, Gao
Y and Yang DL: Osthole inhibits intimal hyperplasia by regulating
the NF-κB and TGF-β1/Smad2 signalling pathways in the rat carotid
artery after balloon injury. Eur J Pharmacol. 811:232–239. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gao Z, Wen Q, Xia Y, Yang J, Gao P, Zhang
N, Li H and Zou S: Osthole augments therapeutic efficiency of
neural stem cells-based therapy in experimental autoimmune
encephalomyelitis. J Pharmacol Sci. 124:54–65. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kordulewska NK, Kostyra E, Cieślińska A,
Fiedorowicz E and Jarmołowska B: Cytokine production by PBMC and
serum from allergic and non-allergic subjects following in vitro
histamine stimulation to test fexofenadine and osthole
anti-allergic proper-ties. Eur J Pharmacol. 791:763–772. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen X, Pi R, Zou Y, Liu M, Ma X, Jiang Y,
Mao X and Hu X: Attenuation of experimental autoimmune
encephalomyelitis in C57 BL/6 mice by osthole, a natural coumarin.
Eur J Pharmacol. 629:40–46. 2010. View Article : Google Scholar
|
|
28
|
Hua KF, Yang SM, Kao TY, Chang JM, Chen
HL, Tsai YJ, Chen A, Yang SS, Chao LK and Ka SM: Osthole mitigates
progressive IgA nephropathy by inhibiting reactive oxygen species
generation and NF-κB/NLRP3 pathway. PLoS One. 8:e777942013.
View Article : Google Scholar
|
|
29
|
Matsuda H, Tomohiro N, Ido Y and Kubo M:
Anti-allergic effects of cnidii monnieri fructus (dried fruits of
Cnidium monnieri) and its major component, osthol. Biol Pharm Bull.
25:809–812. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chiu PR, Lee WT, Chu YT, Lee MS, Jong YJ
and Hung CH: Effect of the Chinese herb extract osthol on IL-4
induced eotaxin expression in BEAS-2B cells. Pediatr Neonatol.
49:135–140. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sun J, Wu J, Xu C, Luo Q, Li B and Dong J:
Paeoniflorin attenuates allergic inflammation in asthmatic mice.
Int Immunopharmacol. 24:88–94. 2015. View Article : Google Scholar
|
|
32
|
Glaab T, Daser A, Braun A,
Neuhaus-Steinmetz U, Fabel H, Alarie Y and Renz H: Tidal
midexpiratory flow as a measure of airway hyperresponsiveness in
allergic mice. Am J Physiol Lung Cell Mol Physiol. 280:L565–L573.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Milstrey A, Wieskoetter B, Hinze D,
Grueneweller N, Stange R, Pap T, Raschke M and Garcia P:
Dose-dependent effect of parathyroid hormone on fracture healing
and bone formation in mice. J Surg Res. 220:327–335. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Qiao Y, Song L, Zhu C, Wang Q, Guo T, Yan
Y and LI Q: Dataset on preparation of the phosphorylated
counterparts of a momordica charantia protein for studying
antifungal activities against susceptible dose-dependent C.
albicans to antimycotics. Data Brief. 15:370–375. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Arancio AL, Cole KD, Dominguez AR,
Cohenour ER, Kadie J, Maloney WC, Cilliers C, Schuh SM, Bisphenol A
and Bisphenol AF: Di-N-Butyl phthalate, and 17β-estradiol have
shared and unique dose-dependent effects on early embryo cleavage
divisions and development in Xenopus laevis. Reprod Toxicol.
84:65–74. 2019. View Article : Google Scholar
|
|
36
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Virchow JC, Backer V, Kuna P, Prieto L,
Nolte H, Villesen HH, Ljørring C, Riis B and de Blay F: Efficacy of
a house dust mite sublingual allergen immunotherapy tablet in
adults with allergic asthma: A randomized clinical trial. JAMA.
315:1715–1725. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Qian J, Ma X, Xun Y and Pan L: Protective
effect of forsythiaside A on OVA-induced asthma in mice. Eur J
Pharmacol. 812:250–255. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wijerathne CUB, Seo CS, Song JW, Park HS,
Moon OS, Won YS, Kwon HJ and Son HY: Isoimperatorin attenuates
airway inflammation and mucus hypersecretion in an
ovalbumin-induced murine model of asthma. Int Immunopharmacol.
49:67–76. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Grotenboer NS, Ketelaar ME, Koppelman GH
and Nawijn MC: Decoding asthma: Translating genetic variation in
IL33 and IL1RL1 into disease pathophysiology. J Allergy Clin
Immunol. 131:856–865. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pichery M, Mirey E, Mercier P, Lefrancais
E, Dujardin A, Ortega N and Girard JP: Endogenous IL-33 is highly
expressed in mouse epithelial barrier tissues, lymphoid organs,
brain, embryos, and inflamed tissues: In situ analysis using a
novel Il-33-LacZ gene trap reporter strain. J Immunol.
188:3488–3495. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Schmitz J, Owyang A, Oldham E, Song Y,
Murphy E, McClanahan TK, Zurawski G, Moshrefi M, Qin J, Li X, et
al: IL-33, an interleukin-1-like cytokine that signals via the IL-1
receptor-related protein ST2 and induces T helper type
2-associ-ated cytokines. Immunity. 23:479–490. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liew FY, Pitman NI and McInnes IB:
Disease-associated functions of IL-33: The new kid in the IL-1
family. Nat Rev Immunol. 10:103–110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Griesenauer B and Paczesny S: The
ST2/IL-33 axis in immune cells during inflammatory diseases. Front
Immunol. 8:4752017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bønnelykke K, Sleiman P, Nielsen K,
Kreiner-Møller E, Mercader JM, Belgrave D, den Dekker HT, Husby A,
Sevelsted A, Faura-Tellez G, et al: A genome-wide association study
identifies CDHR3 as a susceptibility locus for early childhood
asthma with severe exacerbations. Nat Genet. 46:51–55. 2014.
View Article : Google Scholar
|
|
46
|
Wan Y, Shrine NR, Soler Artigas M, Wain
LV, Blakey JD, Moffatt MF, Bush A, Chung KF, Cookson WO, Strachan
DP, et al: Genome-wide association study to identify genetic
determinants of severe asthma. Thorax. 67:762–768. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Morita H, Arae K, Unno H, Miyauchi K,
Toyama S, Nambu A, Oboki K, Ohno T, Motomura K, Matsuda A, et al:
An inter-leukin-33-mast cell-interleukin-2 axis suppresses
papain-induced allergic inflammation by promoting regulatory T cell
numbers. Immunity. 43:175–186. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Martin NT and Martin MU: Interleukin 33 is
a guardian of barriers and a local alarmin. Nat Immunol.
17:122–131. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chackerian AA, Oldham ER, Murphy EE,
Schmitz J, Pflanz S and Kastelein RA: IL-1 receptor accessory
protein and ST2 comprise the IL-33 receptor complex. J Immunol.
179:2551–2555. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liew FY, Girard JP and Turnquist HR:
Interleukin-33 in health and disease. Nat Rev Immunol. 16:676–689.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Drake LY and Kita H: IL-33: Biological
properties, functions, and roles in airway disease. Immunol Rev.
278:173–184. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Jackson DJ, Makrinioti H, Rana BM, Shamji
BWH, Trujillo-Torralbo MB, Footitt J, Del-Rosario J, Telcian AG,
Nikonova A, Zhu J, et al: IL-33-dependent type 2 inflammation
during rhinovirus-induced asthma exacerbations in vivo. Am J Respir
Crit Care Med. 190:1373–1382. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Rank MA, Kobayashi T, Kozaki H, Bartemes
KR, Squillace DL and Kita H: IL-33-activated dendritic cells induce
an atypical TH2-type responses. J Allergy Clin Immunol.
123:1047–1054. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kurowska-Stolarska M, Stolarski B, Kewin
P, Murphy G, Corrigan CJ, Ying S, Pitman N, Mirchandani A, Rana B,
van Rooijen N, et al: IL-33 amplifies the polarization of
alternatively activated macrophages that contribute to airway
inflammation. J Immunol. 183:6469–6477. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Millien VO, Lu W, Shaw J, Yuan X, Mak G,
Roberts L, Song LZ, Knight JM, Creighton CJ, Luong A, et al:
Cleavage of brinogen by proteinases elicits allergic responses
through Toll-like receptor 4. Science. 341:792–796. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yanagawa Y, Suzuki M, Matsumoto M and
Togashi H: Prostaglandin E(2) enhances IL-33 production by
dendritic cells. Immunol Lett. 141:55–60. 2011. View Article : Google Scholar : PubMed/NCBI
|