Introduction
Age-related cataract (ARC) is a common eye disease
that manifests as the lens changes from transparent to opaque with
age. ARC is the leading cause of visual impairment and blindness
worldwide among the aged population (1-3).
Studies have suggested that the oxidation of macromolecules
triggered by ultraviolet radiation, metabolic abnormalities,
smoking, drugs and other toxic factors contributes to the onset of
ARC (4-6). Oxidative stress can cause DNA
strand-breaks in lens cells, and the failure of the damage repair
may aggravate the irreversible lesion (7,8).
Oxidative stress stimulates the generation of
8-oxo-2′-de-oxyguanosine (8-oxo-dG) in lens epithelial cells
(LECs). If 8-oxo-dG:C cannot be repaired, a mismatch of 8-oxo-dG
with A will be formed during DNA replication, thus causing
functional alterations in related genes (9). 8-Oxoguanine DNA glycosylase (OGG1)
can recognize and excise 8-oxo-dG in DNA double-strands, thereby
restoring normal G:C pairing in the genome and preventing the
mutagenic effects of 8-oxo-dG (10). Studies have indicated that the
downregulation of the OGG1 gene may lead to the impairment
of its DNA repair function, which is related to the occurrence and
development of ARC (11,12).
Long non-coding RNAs (lncRNAs) refer to RNAs with
>200 nucleotides in length, without protein-coding capacity or
with limited protein-coding capacity. lncRNAs have important
regulatory functions and can participate in the proliferation,
apoptosis and migration of cells (13-15). Recently, a variety of lncRNAs
playing key roles in ARC have been identified (16-19). However, the regulation of
OGG1 by lncRNAs has rarely been reported, at least to the
best of our knowledge.
In the present study, CRISPR/Cas9 gene editing was
used to knock out the ogg1 gene in zebrafish. The model
demonstrated that the disruption of ogg1 aggravated
cataract, and the lncRNAs targeting OGG1 were then screened
out from the human lens epithelial cell line (SRA01/04) and patient
lens capsule samples in order to investigate their role in the
pathogenesis of ARC.
Materials and methods
Clinical samples
According to the classification standard of Lens
Opacities Classification III (LOCS III), 10 patients with cortical
ARC, nuclear ARC and posterior subcapsular ARC were randomly
selected, respectively (20). For
the controls, 10 age-matched patients with transparent lens removed
from vitreoretinal disease were selected. Patients with complex
cataract due to ocular trauma, high myopia, glaucoma or uveitis, as
well as those with systemic diseases, such as hypertension or
diabetes, were excluded. Continuous curvilinear capsulorhexis was
performed during cataract surgery to obtain the lens capsule
specimens. The present study followed the principles of the
Helsinki Declaration and was approved by the Ethics Committee of
the Affiliated Hospital of Nantong University. The study objectives
and procedures were explained to all the participants, and related
informed consent was signed from all the participants.
Zebrafish
In the present study, zebrafish of the AB strain was
raised and maintained in a circulating water system at 28.5°C with
a light time of 14 h and a dark time of 10 h per day. The embryos
were fed with E3 medium in an incubator at 28.5°C, and the medium
was changed every day. The experimental protocol was approved by
the Animals Care and Use Committee of Nantong University and was
conducted in conformity with National Institutes of Health
Guidelines for the Care and Use of Laboratory Animals.
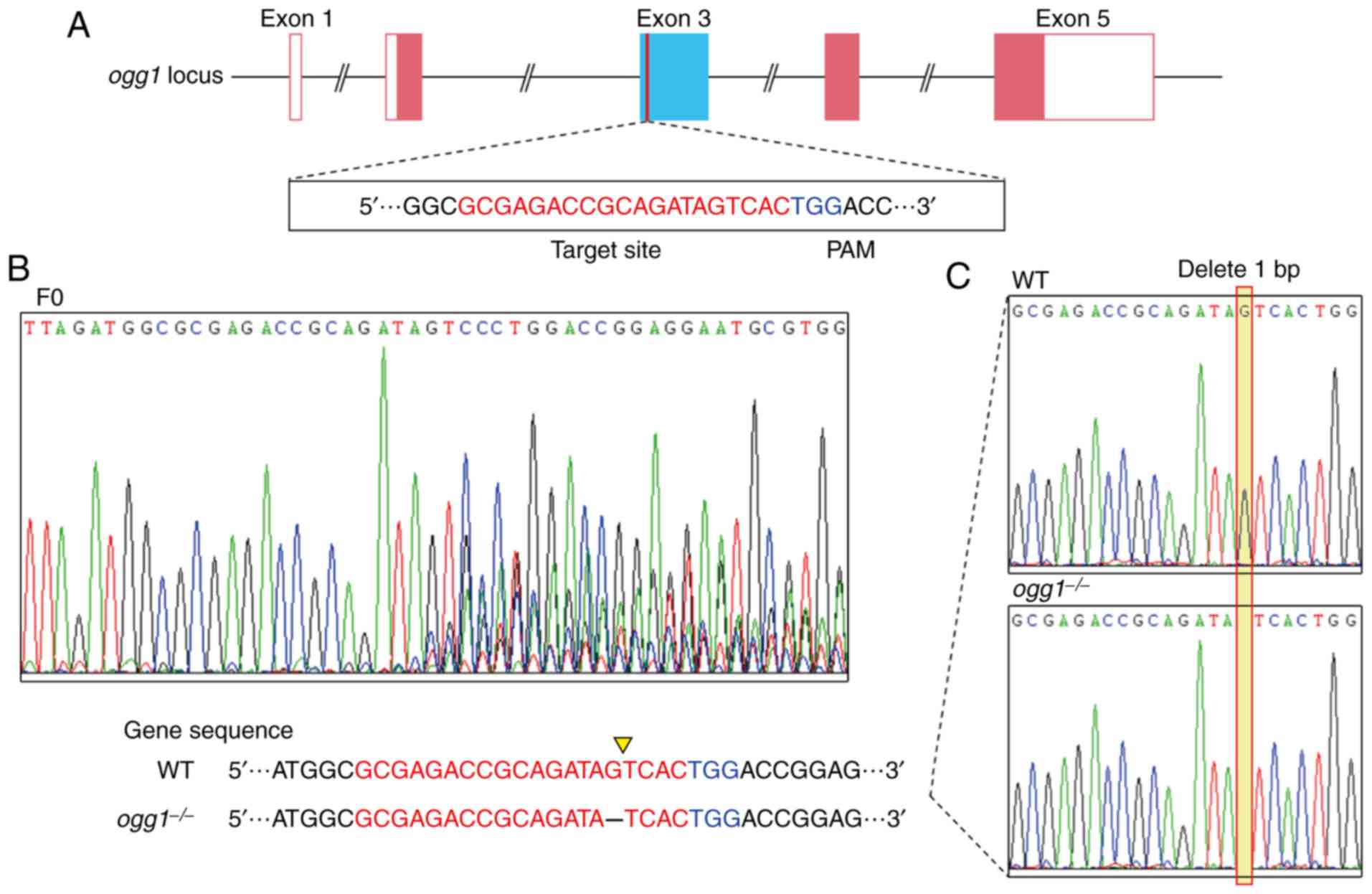
Generation of ogg1 mutants
The gRNA sequence was designed based on the
information obtained from the CHOPCHOP website (http://chopchop.cbu.uib.no/). The target site located
in exon 3 of the zebrafish ogg1 gene
(5′-GCGAGACCGCAGATAGTCAC-3′) has 20 bases upstream of the
protospacer adjacent motif (PAM) (TGG). The gRNA and the Cas9 mRNA
were synthesized by in vitro transcription using the
mMESSAGE mMACHINE T7 kit and the MAXIscript kit (both from Ambion;
Thermo Fisher Scientific, Inc.), respectively, which were mixed in
proportion (gRNA 50 ng/µl and Cas9 mRNA 150 ng/µl)
and injected into one cell-phase zebrafish embryo (F0) (21). A total of 10-15 embryos were
selected at 2 days post-fertilization (dpf) to extract the genomic
DNA for sequencing and identification. Mature F0 zebrafish were
then hybridized with wild-type AB zebrafish to produce the
offspring (F1) containing heterozygous mutants, and the
heterozygous F1 generations were then genotyped by tail fin-cutting
sequencing to determine the presence of frameshift mutation. The F1
male and female zebrafish with the same mutant type were mated to
produce the homozygous F2 mutant. The cataract model was
constructed monocularly when the F2 mutant zebrafish grew to 3
months of age. The Cas9 template plasmid and gRNA template plasmid
were donated by the Jiangsu Key Laboratory of
Neurodegeneration.
Construction of zebrafish cataract
model
Oxidative damage to the lens was performed by
injecting 0.5 µl of 2% hydrogen peroxide into the anterior
chamber of adult zebrafish with 32G needles (22).
H&E staining
The zebrafish were euthanized by an overdose of
MS222 (250 mg/l, Sigma-Aldrich; Merck KGaA) prior to obtaining the
samples. After immersing the zebrafish in this solution for
approximately 60 sec, opercular movement ceased. Immersion was
continued for 15 min following the cessation of opercular movement
to ensure that the zebrafish had died. The paraffin-embedded
sections were soaked with hematoxylin solution (Beijing Solarbio
Science & Technology Co., Ltd.) for 5 min, and then soaked with
1% hydrochloric acid ethanol for 3 sec, followed by rinsing with
distilled water. Eosin solution (Beijing Solarbio Science &
Technology Co., Ltd.) was then used for the 3-min staining of the
paraffin sections, followed by dehydration with 80, 95 and 100%
ethanol in gradient, hyalinization with xylene, sealing with
neutral resin, and observation with a microscope (Zeiss AG).
Cell culture and oxidative damage
The human LEC cell line, SRA01/04, was purchased
from the Cell Bank of the Chinese Academy of Sciences, which was
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.) containing
1% penicillin/streptomycin and 10% fetal bovine serum (FBS) (Gibco;
Thermo Fisher Scientific, Inc.) at 37°C and 5% CO2 in a
humid environment.
The cells were divided into the control group, the
UVB group and the H2O2 group. The UVB group
was further subdivided into 3 subgroups, which were irradiated with
ultra-violet rays for 5, 10 and 30 min, respectively. The cells
were maintained in culture following irradiation and harvested 24 h
later. One XX-15B UVB lamp (Spectroline; Spectronics Corporation)
was used with a spectrum of 280-320 nm and a maximum emission peak
of 312 nm. The H2O2 group was further
subdivided into 3 subgroups, and the cells are soaked in medium
containing 10, 20 and 50 µmol/l H2O2,
respectively, for 24 h and then harvested. In total, 2 subgroups
with a similar apoptotic rate were selected from the UVB group and
H2O2 group, respectively, for lncRNA
high-throughput sequencing.
Flow cytometry
Cells were incubated with FITC and PI for 15 min in
darkness at room temperature using the Annexin V-FITC/PI Apoptosis
Detection kit (BD Biosciences) and then detected by flow cytometry
(BD Biosciences). Annexin V-FITC-positive cells were defined as
early apoptotic cells, and Annexin V-FITC and PI dual-positive
cells were defined as late apoptotic cells.
High-throughput lncRNA sequencing
Total RNA was extracted from the cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. RNA integrity was measured by 1.5%
agarose gel electrophoresis with an OD260/280 between 1.8 and 2.0.
High-throughput lncRNA sequencing was performed by Sinotech
Genomics. First, the quality of the original data was evaluated to
filter out unqualified sequences. Genome mapping was then performed
to classify the sequences according to their location. The mRNAs
and lncRNAs were quantified, the association between the samples
was analyzed, and the differentially expressed mRNAs and lncRNAs
were screened out.
RT-qPCR
Total RNA was reverse transcribed using the
PrimeScript RT kit (Takara Bio, Inc.) and RT-qPCR was performed
using the Step One Plus Real-Time PCR System (Applied Biosystems;
Thermo Fisher Scientific, Inc.) using SYBR-Green (Takara Bio, Inc.)
with GADPH being used as the reference for lncRNA and OGG1
mRNA, and U6 for miRNA. The sequences of the forward and reverse
primers were as follows, respectively: OGG1,
5′-TTGATGATGTCACCTACCATGG-3′ and 5′-CATATGAGGACTCTCGTAGCTG-3′;
miR-4728-5p, 5′-TGGGAGGGGAGAGGCAG-3′ and
5′-AGTGCAGGGTCCGAGGTATT-3′; NONHSAT143692.2,
5′-TGGTCCTTCCTCCACAGACTTCAG-3′ and 5′-TCACCACCAGCCTGAGCAGAC-3′;
GAPDH, 5′-TGAAGGTCGGAGTCAACGGATTTGGT-3′ and
5′-CATGTGGGCCATGAGGTCCACCAC-3′; U6, 5′-CTCGCTTCGGCAGCACA-3′ and
5′-AACGCTTCACGAATTTGCGT-3′. Real-time cycling conditions for lncRNA
and OGG1 mRNA were 95°C for 10 min for initial denaturation,
followed by 45 cycles of 95°C within 10 sec for denaturation, 60°C
for 10 sec for annealing and 72°C for 10 sec for the extension
step. Real-time cycling conditions for miRNA were 95°C for 10 min
for initial denaturation, followed by 40 cycles of 95°C within 2
sec for denaturation, 60°C for 20 sec for annealing and 70°C for 10
sec for the extension step. Relative gene expression levels were
calculated by comparing the GAPDH or U6 levels using the
2−ΔΔCq method (23).
Bioinformatics prediction and luciferase
activity assay
RegRNA 2.0 (http://regrna2.mbc.nctu.edu.tw/) and TargetScan
(http://www.targetscan.org/) were used to
predict the miRNAs that bind both the differentially expressed
lncRNA and the OGG1 mRNA 3′UTR.
The 293T cells were purchased from the Cell Bank of
the Chinese Academy of Sciences. The wild-type or mutant-type
sequence fragment of differentially expressed lncRNA
(NONHSAT143692.2) and OGG1 mRNA 3′UTR that can bind to the
same miRNA (miR-4728-5p) was cloned into the pmirGLO dual
luciferase vector (Promega Corporation). The vector and miRNA
mimics or NC were co-transfected into 293T cells. After 48 h, the
luciferase activity was evaluated using the Dual Luciferase
Reporter Assay System (Promega Corporation) according to the
manufacturer's instructions. These experiments were repeated at
least 3 times in triplicate. The relative luciferase activities
were determined by calculating the ratio of Renilla
luciferase activities over Firefly luciferase activities.
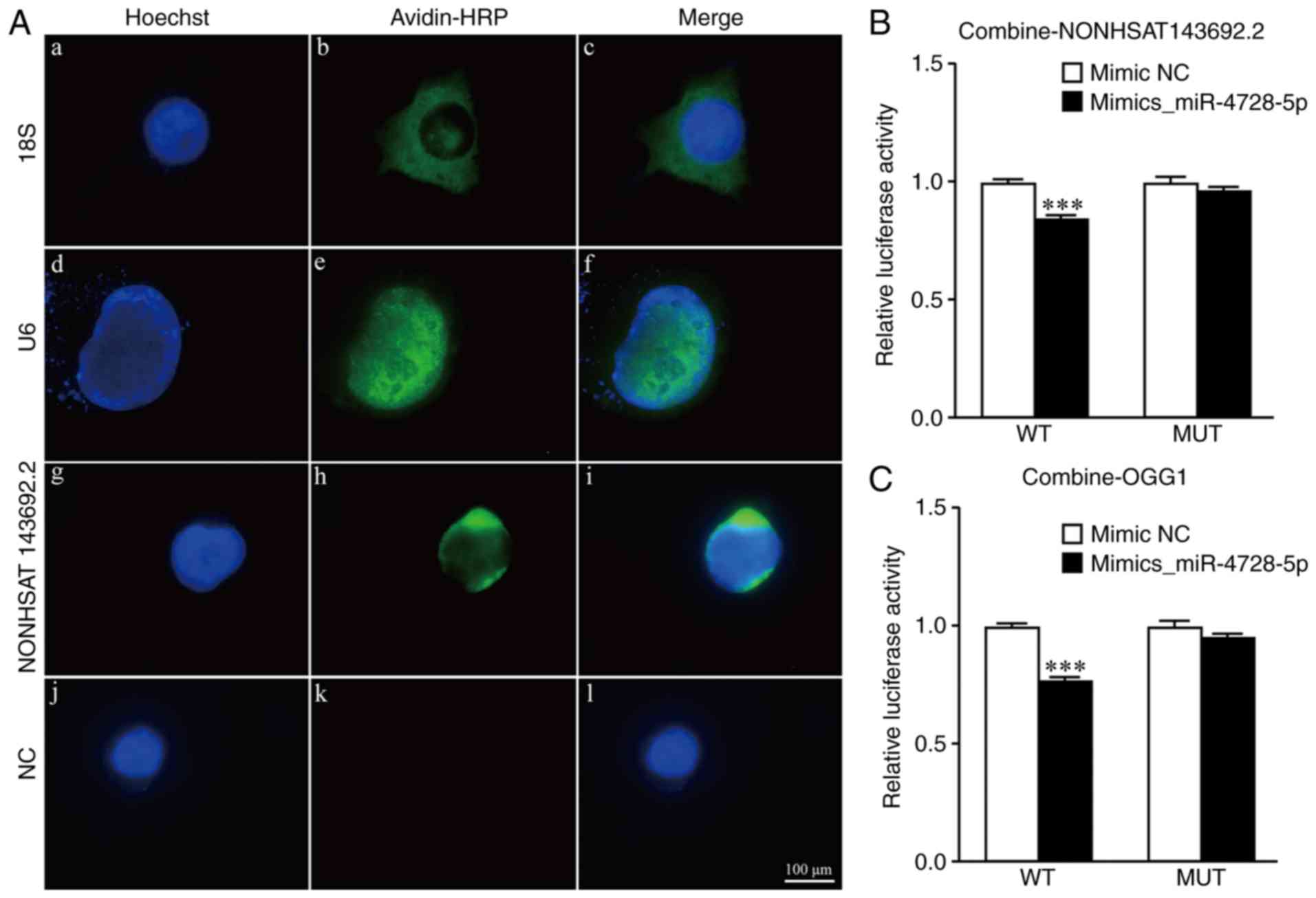
RNA fluorescence in situ
hybridization
Digoxin-labeled probe (Exon Biotechnology Inc.,
Guangzhou, China) was used to identify the intracellular
localization of lncRNA. Cells were treated with 0.5% Triton X-100,
20 µg/ml proteinase K, and 4% PFA for 5 min, and then soaked
with 3% H2O2 for 30 min. The following
conditions were used: Probe: Hybridization solution=1:50 dilution,
5-min denaturation at 88°C, 10-min equilibration at 37°C, and 24-h
hybridization. Cells were incubated with 3% BSA for 30 min at 37°C,
followed by 1-h culture with Avidin-HRP: 1% BSA=1:100 dilution,
15-min culture with TSA (green): 0.15% H2O2:
TSA dilution=1:1:100, and final counterstaining with Hoechst
(Guangzhou RiboBio Co., Ltd.) for 15 min at room temperature. The
cells were then observed under a confocal microscope (Leica
Microsystems GmbH).
Lentivirus infection and cell
transfection
The NONHSAT143692.2 lentiviral shRNAs were
synthesized and sub-cloned into the pGLV3/H1/GFP&Puro vector
(GenePharma) to knockdown the expression of NONHSAT143692.2.
NONHSAT143692.2-overexpressing lentiviral constructs were generated
by subcloning sh-NONHSAT143692.2 into pGLV5/EF-1aF/GFP&Puro
vector (GenePharma). Empty vectors were used as negative controls,
respectively. All the constructed plasmids were confirmed by
sequencing (Invitrogen; Thermo Fisher Scientific, Inc.). They were
added to SRA01/04 cells at approximately 70% confluency. The
lentivirus was diluted in a 1:5 mixture with the medium and the
total volume was approximately 1 ml. Stable SRA01/04 cells were
then selected by puromycin (2.5 µg/ml, Sigma-Aldrich; Merck
KGaA) for a further 48 h.
Human miR-4728-5p mimics/control and miR-4728-5p
inhibitor/control were purchased from GenePharma. The transfection
of SRA01/04 cells was carried out using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The final miR-4728-5p mimics/control
concentration was 50 nM, and the final miR-4728-5p
inhibitor/control concentration was 100 nM for transfection.
EDU assay
Cell proliferation was measured using the BeyoClick
EdU Cell Proliferation kit (Beyotime Institute of Biotechnology,
Inc.). EdU working solution (20 µM) pre-warmed at 37°C and
cell culture medium were added to 24-well plates with equal volumes
for a 4-h incubation at 37°C. After rinsing, the cells were fixed
with 4% PFA for 15 min at room temperature, followed by incubation
with 0.3% Triton X-100 for 15 min at room temperature. The Click
reaction solution (Beyotime Institute of Biotechnology, Inc.) was
prepared according to the instructions (Click Reaction Buffer:
CuSO4: Azide 555: Click Additive Solution=430:20:1:50)
for incubating the cells for 30 min in darkness at room
temperature, the nucleus of which were stained with Hoechst and
observed under a fluorescence microscope (Zeiss AG).
TUNEL assay
Apoptosis was detected using the One Step TUNEL
Apoptosis Assay kit (Beyotime Institute of Biotechnology, Inc.).
Cells were fixed with 4% PFA for 30 min at room temperature and
incubated with 0.3% Triton X-100 for 5 min at room temperature. The
TUNEL detection solution was then prepared according to the
instructions for incubating the cells at 37°C in darkness for 60
min, the nucleus of which were stained with Hoechst and observed
under a fluorescence microscope (Zeiss AG).
Western blot analysis
The cell pellets were homogenized in a buffer
containing 50 mM Tris (pH 7.4), 150 mM NaCl, 1% NP-40, 0.25% sodium
deoxycholate and 1% protease inhibitor. The lysate was centrifuged
at 13,000 × g for 10 min at 4°C. Protein concentrations were
determined using a BCA kit (Pierce; Thermo Fisher Scientific,
Inc.). The protein samples with equal amounts (300 µg) were
then separated by 10% SDS-PAGE gel electrophoresis and then
transferred to a PVDF membrane (Thermo Fisher Scientific, Inc.).
The membrane was blocked with 5% skim milk at 37°C and incubated
with rabbit anti-human OGG1 primary antibody (1:1,000, ab124741;
Abcam) and GADPH (1:2,000, ab181602; Abcam), overnight at 4°C.
Peroxidase-conjugated goat anti-rabbit IgG secondary antibody
(Santa Cruz Biotechnology, Inc., sc-2004; 1:10,000) was then added
for a further 2-h incubation at room temperature, and the cells
were then visualized using an ECL kit (Pierce; Thermo Fisher
Scientific, Inc.) for protein band quantification by densitometry
using ImageJ software 1.52a (NIH).
Statistical analysis
Each experiment was repeated 3 times, and the data
are expressed as the means ± standard deviation. The results were
analyzed using SPSS 25.0 and GraphPad Prism 8.0, using
single-factor analysis of variance (one-way ANOVA), carried out
between the mean pairwise comparisons using the LSD test and
Tukey's test, with P<0.05 being considered to indicate a
statistically significant difference.
Results
Ogg1 mutation and lens lesions in
zebrafish
Ogg1 mutant zebrafish were successfully
generated by CRISPR/Cas9 technology. Genotyping revealed a deletion
of a base G in ogg1 exon 3, which resulted in the frameshift
mutation (Fig. 1).
Adult (>3 months old) wild-type and mutant
zebrafish lenses remained transparent (Fig. 2A). However, following the
injection of the low dose of H2O2 into the
anterior chamber, the wild-type adult zebrafish only developed mild
cataract, whereas the mutant adult zebrafish developed severe
cataract. H&E staining revealed that the lens cortex of the
mutant fish exhibited a clear vacuolar-like structure, which was
similar to human cortical cataract, while no similar structure was
found in the wild-type fish (Fig.
2B).
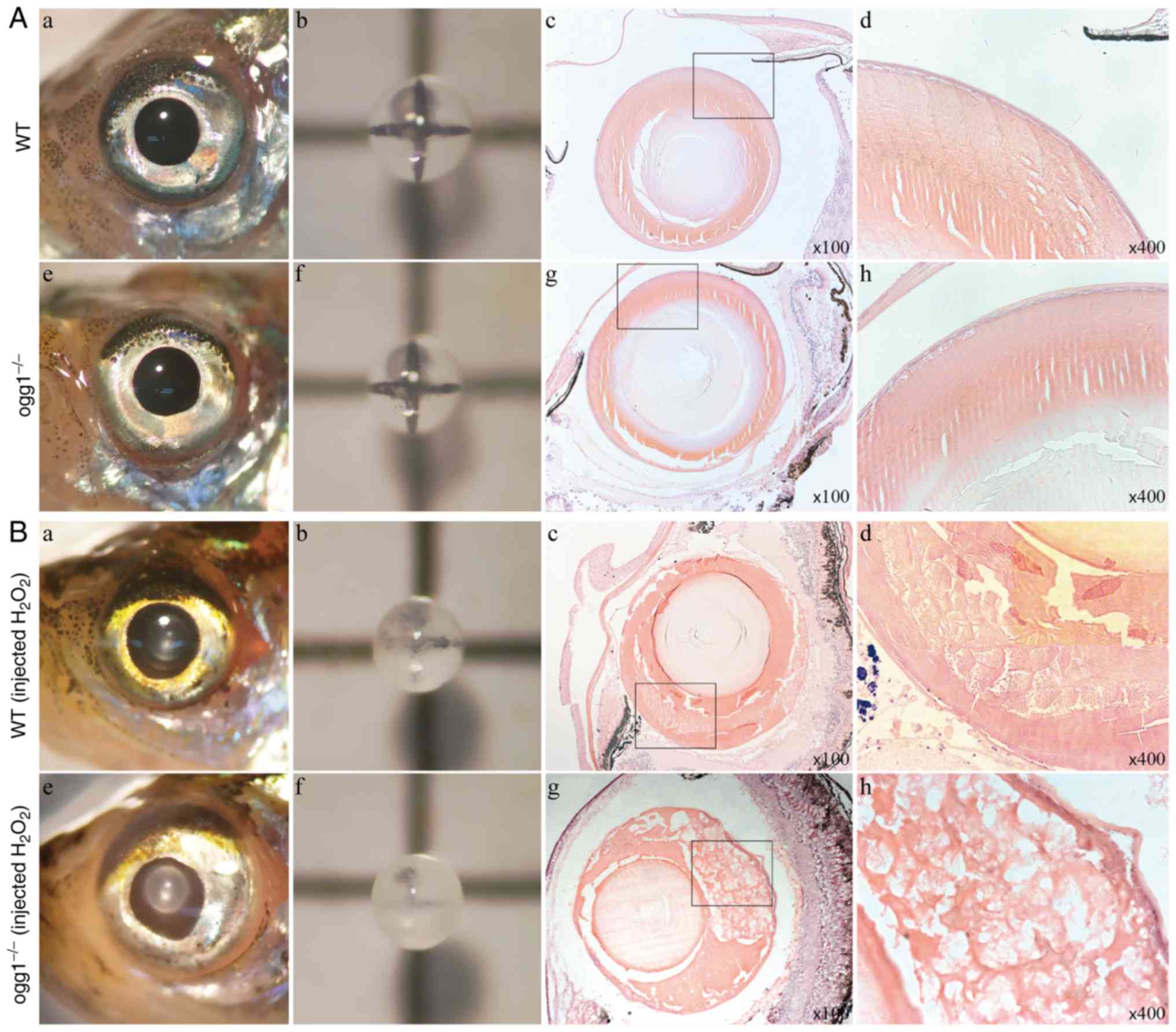 | Figure 2Phenotypic analysis of ogg1
mutant zebrafish. (A) The lenses of wild-type and
ogg1−/− adult zebrafish were all transparent, and
the lens cortex was uniformly dense with H&E staining. (B)
Following the injection of 5 µl of 2%
H2O2 in the anterior chamber, compared with
the wild-type fish, the lens opacity of ogg1−/−
adult zebrafish was more evident. H&E-staining revealed a
slightly looser lens cortex structure in the wild-type fish, as
well as a large number of vacuole-like structures appearing in the
mutant lens cortex. (A and B) (a-d) Present images of the anterior
segment, lens, H&E-stained sections (×100 magnification) and
H&E-stained sections (×400 magnification) of adult zebrafish in
the wild-type group, respectively. (e-h) Present images of the
anterior segment, lens, H&E-stained sections (×100
magnification) and H&E-stained sections (×400 magnification) of
adult zebrafish in the mutant group, respectively. WT, wild-type;
ogg1−/−, homozygous mutant of ogg1. |
Distinct expression of lncRNAs with
target gene as OGG1 following UVB exposure
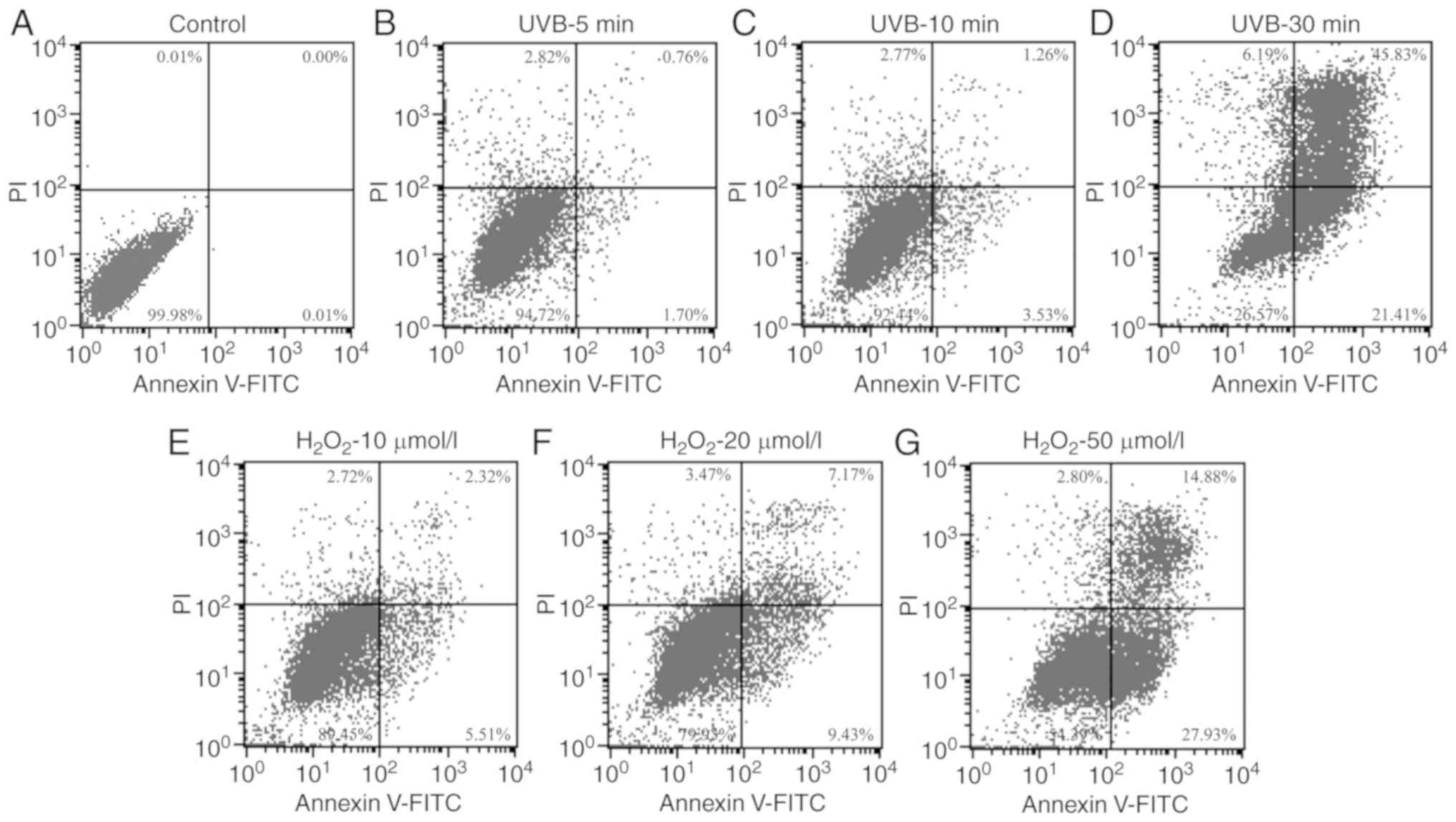
Flow cytometry revealed similar apoptotic rates in
the SRA01/04 cells irradiated with UVB for 10 min or exposed to 10
µmol/l H2O2 (Fig. 3).
High-throughput sequencing revealed that compared
with the control group, in the UVB group, there were 1,410
upregulated mRNAs, and 3,270 downregulated mRNAs; 295 lncRNAs were
upregulated, while 2,177 lncRNAs were downregulated. In the
H2O2 group, 1,211 mRNAs were upregulated,
while 3,756 mRNAs were downregulated; in addition, 44 lncRNAs were
upregulated, while 344 lncRNAs were downregulated (Fig. 4A).
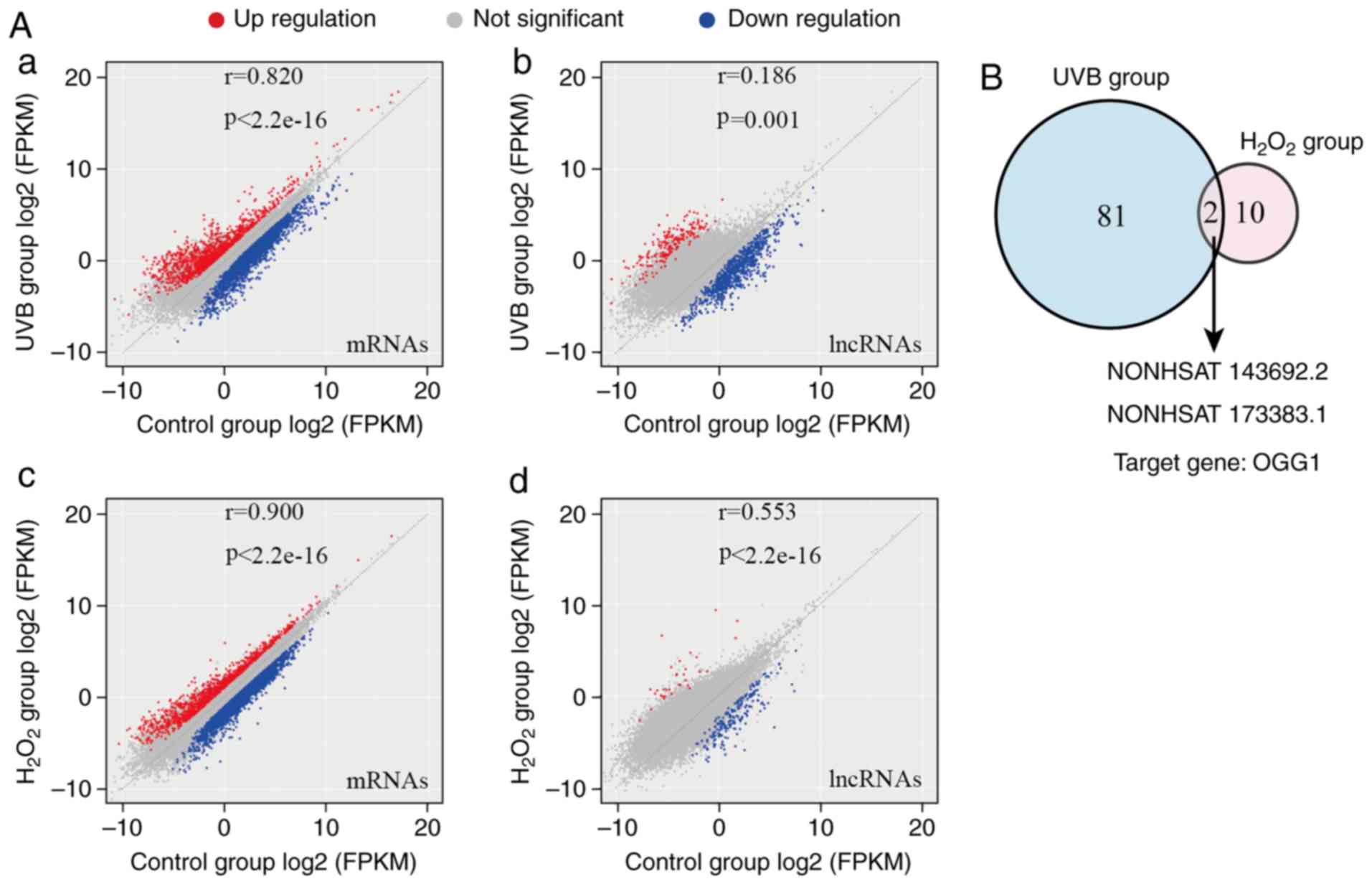 | Figure 4Results of high-throughput lncRNA
sequencing. (A) Scatter plot of sequencing results. (a) In the UVB
group, there were 1,410 upregulated mRNAs, and 3,270 downregulated
mRNAs; (b) 295 lncRNAs were upregulated, while 2,177 lncRNAs were
downregulated. (c) In the H2O2 group, 1,211
mRNAs were upregulated, while 3,756 mRNAs were downregulated; (d)
in addition, 44 lncRNAs were upregulated, while 344 lncRNAs were
downregulated. (B) Expression levels of NONHSAT143692.2 and
NONHSAT173383.1, which exhibited the most significant differences
in the 2 groups. |
According to the bioinformatics prediction, the
lncRNAs with the target gene as OGG1 and exhibiting a
differential expression were selected. There were 81 lncRNAs in the
UVB group and 10 in the H2O2 group. The 2
groups had 2 identical lncRNAs exhibiting the most significant
differences: NONHSAT143692.2 and NONHSAT173383.1 (Fig. 4B).
The SRA01/04 cell line was irradiated with UVB again
to verify the screening results. In the early stages of injury (5
min of irradiation), the expression levels of OGG1 mRNA and
NONHSAT143692.2 increased significantly (P<0.01), while the
expression of NONHSAT173383.1 decreased significantly (P<0.001).
In the late stages of injury (30 min of continuous irradiation),
the expression levels of OGG1 mRNA and NONHSAT143692.2
decreased significantly (P<0.01, P<0.001), while the
expression of NONHSAT173383.1 increased significantly (P<0.001)
(Fig. 5A–C).
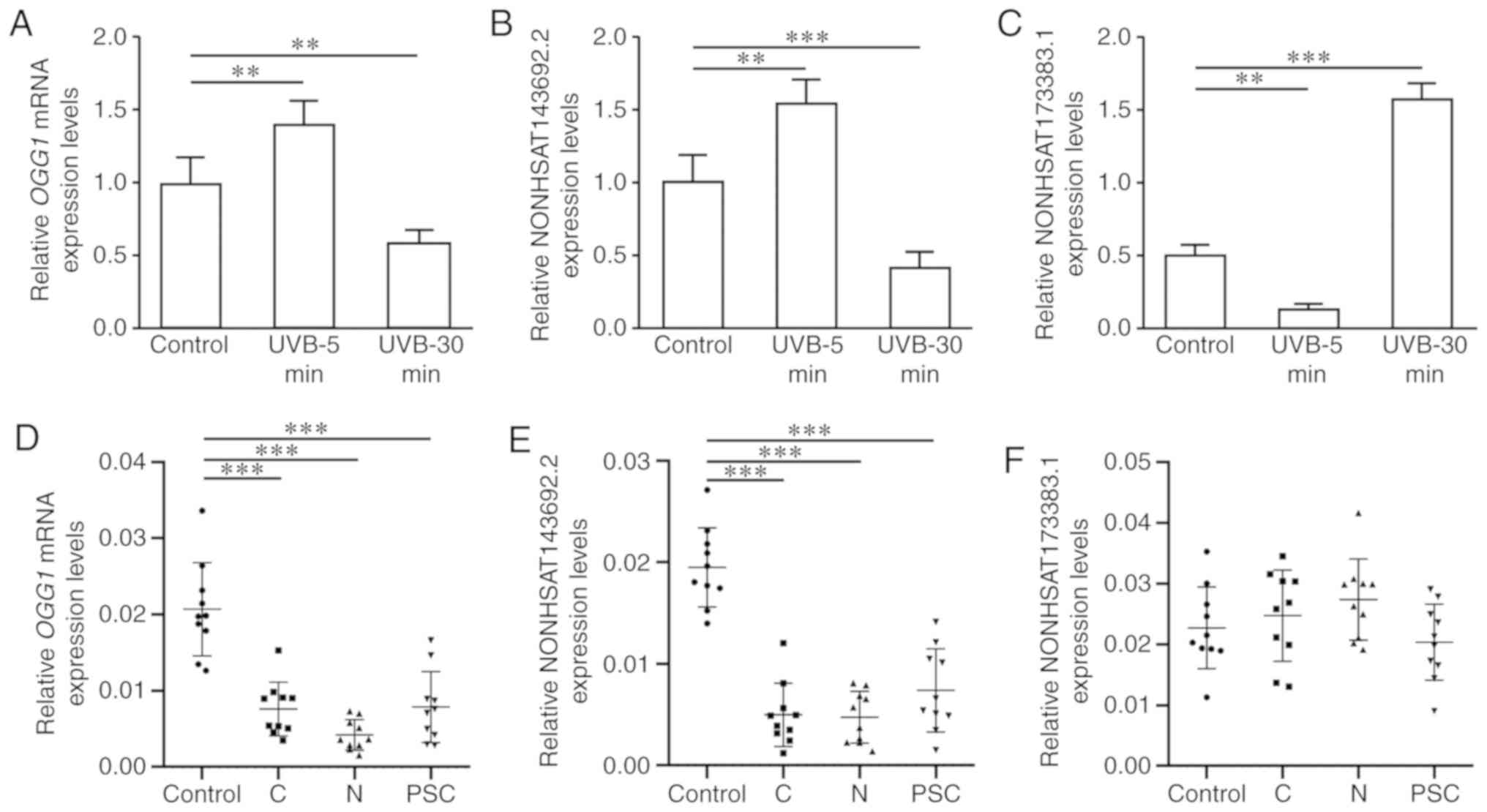 | Figure 5Expression levels of OGG1
mRNA, NONHSAT143692.2 and NONHSAT173383.1 in SRA01/04 cells and
lens capsule specimens from patients with cataract. (A) OGG1
mRNA expression was upregulated in the early stages, whereas it was
downregulated in the late stages of oxidative damage. (B)
NONHSAT143692.2 was upregulated in the early stages, whereas it was
downregulated in the late stages of oxidative damage. (C)
NONHSAT173383.1 was downregulated in the early stages, whereas it
was upregulated in the late stages of oxidative damage. (D)
Compared with the control group, OGG1 mRNA expression was
downregulated in patients with ARC. (E) Compared with the control
group, NONHSAT143692.2 expression was downregulated in patients
with ARC. (F) NONHSAT173383.1 did not exhibit any marked
differences in expression between patients with ARC and the control
group. ARC, age-related cataract; C, cortical ARC; N, nuclear ARC;
PSC, posterior subcapsular ARC. **P<0.01,
***P<0.001. |
In lens capsule samples from patients with cataract,
the expression levels of OGG1 mRNA and NONHSAT143692.2 in
the cortical, nuclear and posterior subcapsular ARC were all
significantly decreased (P<0.001); however, NONHSAT173383.1
expression exhibited no significant differences among groups
(Fig. 5D–F). Therefore,
NONHSAT143692.2 was selected for further research.
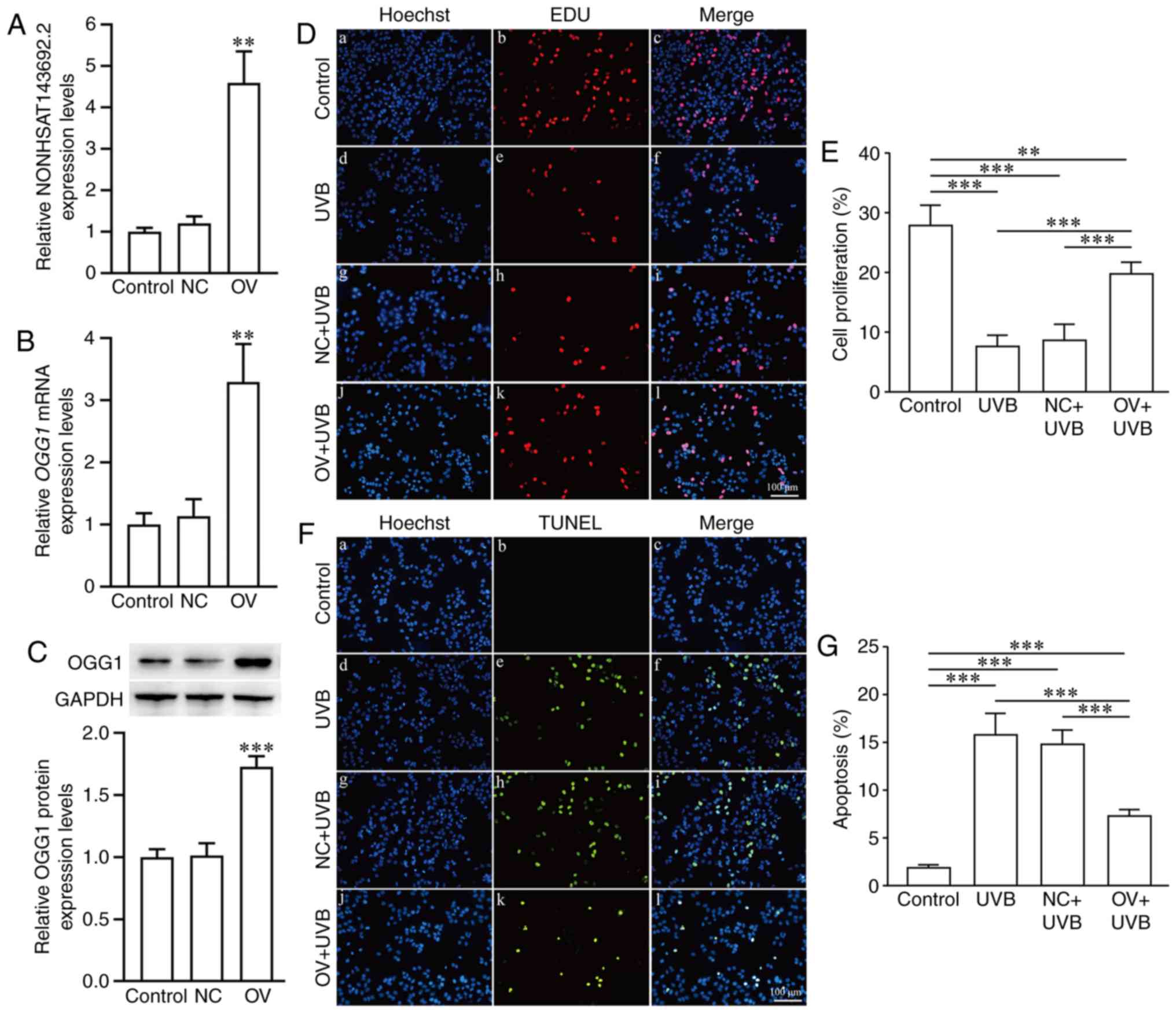
Regulatory effects of NONHSAT143692.2 on
the expression of OGG1 and cellular phenotypes of LECs
Through lentiviral invasion, SRA01/04 cells
overexpressing NONHSAT143692.2 (OV group) and an NC group were
established. Following the overexpression of NONHSAT143692.2, the
expression levels of OGG1 mRNA and OGG1 protein also
increased (P<0.01 and P<0.001) (Fig. 6A–C). EdU assay revealed that
following UVB irradiation, the number of cells proliferating in the
OV group was significantly higher than that in the NC group
(Fig. 6D and E). At the same
time, TUNEL assay revealed that the number of apoptotic cells in
the OV group was significantly lower than that in the NC group
(Fig. 6F and G).
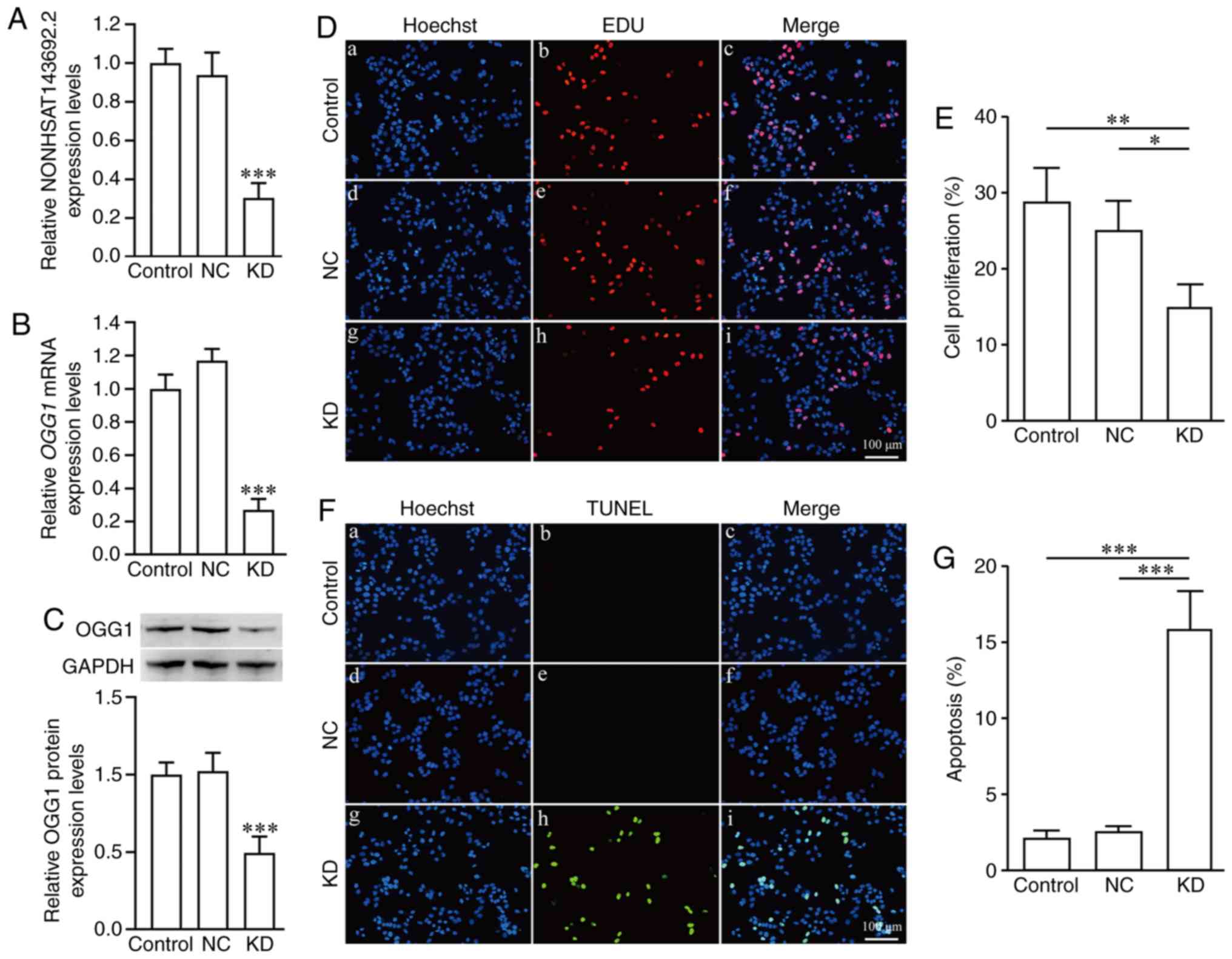
Using the same method of lentivirus invasion,
SRA01/04 cells (KD group) in which NONHSAT143692.2 was knocked down
and the NC group were established. Following the knockdown
NONHSAT143692.2, the expression levels of OGG1 mRNA and OGG1
protein also decreased (P<0.001 and P<0.001) (Fig. 7A–C). EdU assay revealed that the
cells in the KD group proliferated significantly less than those in
the NC group (Fig. 7D and E). At
the same time, TUNEL assay demonstrated that there were
significantly more apoptotic cells in the KD group than the NC
group (Fig. 7F and G). The cells
in the KD group were unable to tolerate UVB irradiation.
Binding of miR-4728-5p to NONHSAT143692.2
and OGG1 mRNA 3′UTR
RNA fluorescence in situ hybridization
revealed that NONHSAT143692.2 was expressed in the cytoplasm; thus,
which suggests the existence of a competing endogenous RNA (ceRNA)
mechanism (Fig. 8A). RegRNA 2.0
website analysis revealed that 6 miRNAs could bind to
NONHSAT143692.2: miR-1285-3p, miR-1273e, miR-3922-5p, miR-4695-5p,
miR-4728-5p and miR-1273g-3p. TargetScan website analysis revealed
that miR-4728-5p could be bind with OGG1 mRNA 3′UTR.
To verify the binding of m i R- 4728-5p to
NONHSAT143692.2, a luciferase reporter gene containing the expected
miR-4728-5p binding site (NONHSAT 143692.2_WT) and the
corresponding mutation site (NONHSAT143692.2_MUT) was constructed.
The results revealed that co-transfection with miR-4728-5p and
NONHSAT143692.2_WT significantly reduced the luciferase activity
(P<0.001); however, co-transfection with miR-4728-5p and
NONHSAT143692.2_MUT did not alter the luciferase activity (Fig. 8B).
To verify the binding of miR-4728-5p to the
OGG1 mRNA 3′UTR, a luciferase reporter gene containing the
expected miR-4728-5p binding site (OGG1_WT) and the corresponding
mutation site (OGG1_MUT) was constructed. The results revealed that
co-transfection with miR-4728-5p and OGG1_ WT significantly reduced
the luciferase activity (P<0.001), whereas co-transfection with
miR-4728-5p and OGG1_MUT did not markedly alter the luciferase
activity (Fig. 8C).
Changes in miR-4728-5p, OGG1 mRNA, OGG1
protein and NONHSAT143692.2 expression
miR-4728-5p expression in SRA01/04 cells following
UBV exposure presented a two-phase change, with a decrease at 5 min
following irradiation (P<0.001), and an increase after 30 min of
continuous irradiation (P<0.001) (Fig. 9A).
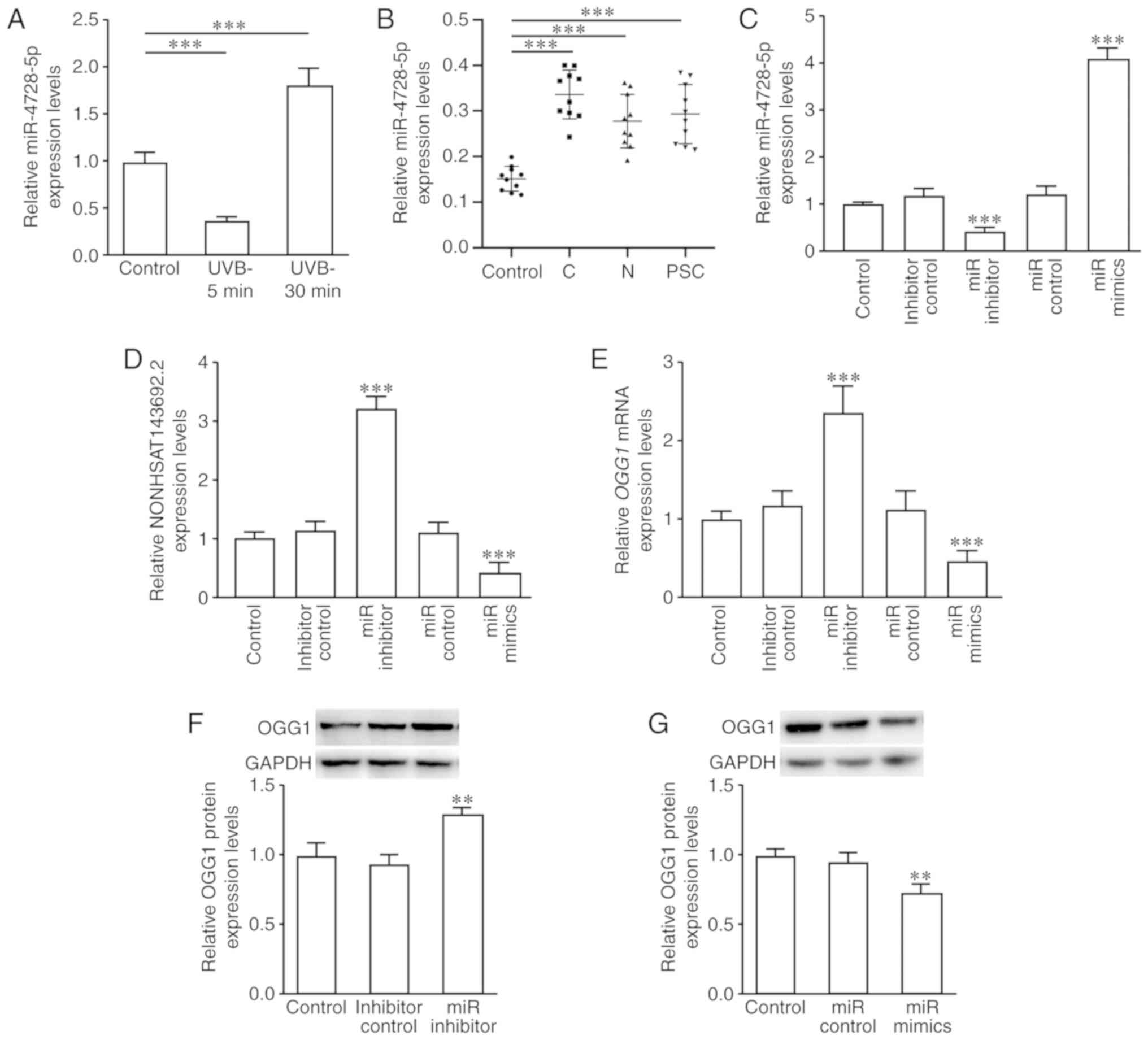 | Figure 9miR-4728-5p expression and effects on
NONHSAT143692.2, OGG1 mRNA, and OGG1 protein expression
following intervention. (A) miR-4728-5p expression was
downregulated in the early stages of oxidative damage and
upregulated in the late stages. (B) miR-4728-5p expression was
upregulated in patients with ARC compared to the control group.
(C-G) Following the knockdown of miR-4728-5p, the expression levels
of NONHSAT143692.2, OGG1 mRNA and OGG1 protein were
upregulated; following the overexpression of miR-4728-5p, these
results were reversed. ARC, age-related cataract; C, cortical ARC;
N, nuclear ARC; PSC, posterior subcapsular ARC.
**P<0.01, ***P<0.001. |
In the lens capsule samples of the patients with
cataract, miR-4728-5p expression was significantly increased in the
cortical, nuclear and posterior capsule ARC (P<0.001) (Fig. 9B). These changes were inversely
associated with the changes in OGG1 mRNA and NONHSAT143692.2
expression.
The SRA01/04 cells were then transfected with
miR-4728-5p control, miR-4728-5p inhibitor and miR-4728-5p mimics.
RT-qPCR revealed that transfection with miR-4728-5p inhibitor
upregulated the expression levels of NONHSAT143692.2 (P<0.001)
and OGG1 mRNA (P<0.001), while transfection with miR-4728-5p
mimics exhibited an opposite trend (Fig. 9C–E). The results of western blot
analysis were consistent with the results of RT-qPCR (Fig. 9F and G).
Discussion
Studies have demonstrated that oxidative
stress-induced DNA damage caused by ultraviolet rays and endogenous
reactive oxygen species can cause the apoptosis of lens epithelial
cells, resulting in lens opacity (24,25). The majority of DNA oxidative
damage can be repaired by the base excision repair pathway (BER),
whereas OGG1 plays an important role in the BER pathway,
particularly in removing 8-oxo-dG (26). Studies on human subjects and other
models have demonstrated that the loss of OGG1 function may
cause various eye diseases, such as ARC or age-related macular
degeneration (AMD), as well as cancers, such as breast cancer and
gastric cancer (11,12,27-30). In previous studies, it was found
that a decreased OGG1 function increased the apoptosis of
LECs and decreased the proliferation of LECs (31,32). In the present study, an
ogg1 mutant zebrafish model was constructed, and it was
demonstrated that when the eyes were placed under oxidative stress,
the mutant zebrafish with the disrupted OGG1 function
developed more severe lens lesions.
Studies in recent years have indicated that lncRNAs
and miRNAs are associated with certain eye diseases, such as
cataract, glaucoma, diabetic retinopathy, or eye tumors (33-36). LncRNAs and miRNAs play an
important role in the apoptosis and proliferation of LECs. They may
be involved in the occurrence and development of ARC via different
mechanisms; however, their association with OGG1 has not yet
been reported (16-19), at least to the best of our
knowledge. We hypothesized that after oxidative stress, lncRNA and
microRNA may increase the apoptosis of LECs by inhibiting the
function of OGG1, leading to lens opacity.
In the present study, through high-throughput
sequencing, it was found that among all lncRNAs that may act on
OGG1, the expression of NONHSAT143692.2 was altered most
substantially in both the UVB group and H2O2
group. NONHSAT143692.2 is located on chromosome 16 with a total
length of 2,466 nts. Its function has not yet been reported, at
least to the best of our knowledge. It was found that this lncRNA
can be detected in the cytoplasm. The majority of the currently
known lncRNAs are located in the nucleus, and a few are located in
the cytoplasm. lncRNAs located in the cytoplasm usually compete for
miRNAs as a type of ceRNA to communicate with mRNAs, affecting the
stability and translation rate of mRNAs. In the present study, it
was demonstrated that NONHSAT143692.2 can regulate the expression
of OGG1 by adsorbing miR-4728-5p; thus, it was hypothesized
that NONHSAT143692.2 may be a molecular sponge.
miR-4728-5p is encoded by the HER2 intron. It has
been reported that the expression of miR-4728 in patients with
breast cancer or gastric cancer is significantly increased, and it
can be used as a non-invasive biomarker for breast cancer and
gastric cancer (37). In the
absence of HER2 inhibitors, miR-4728 can promote tumorigenesis
(38). Breast cancer and gastric
cancer are closely related to DNA oxidative damage, and ARC is also
the result of long-term effects of oxidative stress on the lens and
continuous accumulation of DNA damage. A previous study
demonstrated that young women with early-onset cataracts due to
deficiency in antioxidant function are more likely to develop
breast cancer (39). In the
present study, it was found that miR-4728-5p expression was also
significantly increased in LECs exposed to UVB for a long period of
time and lens capsule samples from patients with ARC, which is
similar to breast cancer and gastric cancer. It is considered that
when a patient's cataract is severe and requires surgery, his/her
lens becomes opaque due to long-term oxidative stress. Therefore,
only SRA01/04 cells exposed to UVB for a long period of time
exhibited the same expression changes as those of patients with
ARC. However, in the early stages of LECs under oxidative stress,
miR-4728-5p was underexpressed. These changes were negatively
associated with changes in NONHSAT143692.2 and
OGG1expression. miR-4728-5p only exists in Homo
sapiens, which also suggests that the pathogenesis of cataract
in human lens with age may greatly differ from that of other
species. In the future, the authors aim to design and perform
experiments to verify the above-mentioned findings in
vivo.
In conclusion, the present study demonstrates that
the loss of OGG1 function may be one of the mechanisms
responsible for the development of ARC. NONHSAT143692.2 regulates
the expression of OGG1 in LECs through the ceRNA mechanism
in combination with miR-4728-5p. The
NONHSAT143692.2/miR-4728-5p/OGG1 axis may thus play an
important role in the development of ARC. This novel concept may
provide new insight into the molecular diagnosis and treatment of
ARC.
Funding
The present study supported by the National Natural
Science Foundation of China (grant nos. 81770906 and 81873676) and
Science and Technology Project of Nantong, China (grant no.
JC2019078).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HG designed the study paper and performed the
critical revision of the manuscript. TZ, JZ, BQ, HX and SZ
performed data collection and analysis. TZ and JZ wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
This study followed the principles of the Helsinki
Declaration and was approved by the Ethics Committee of the
Affiliated Hospital of Nantong University. The study objectives and
procedures were explained to all the participants, and related
informed consent was signed from all the participants. The
zebrafish experimental protocol was approved by the Animals Care
and Use Committee of Nantong University and was conducted in
conformity with National Institutes of Health Guidelines for the
Care and Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
References
|
1
|
Pascolini D and Mariotti SP: Global
estimates of visual impairment: 2010. Br J Ophthalmol. 96:614–618.
2012. View Article : Google Scholar
|
|
2
|
Wei M, Xing KY, Fan YC, Libondi T and Lou
MF: Loss of thiol repair systems in human cataractous lenses.
Invest Ophthalmol Vis Sci. 56:598–605. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Khairallah M, Kahloun R, Bourne R, Limburg
H, Flaxman SR, Jonas JB, Keeffe J, Leasher J, Naidoo K, Pesudovs K,
et al: Number of people blind or visually impaired by cataract
world-wide and in world regions, 1990 to 2010. Invest Ophthalmol
Vis Sci. 56:6762–6769. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chang JR, Koo E, Agrón E, Hallak J,
Clemons T, Azar D, Sperduto RD, Ferris FL III, Chew EY, Age-Related
Eye Disease and Study Group: Risk factors associated with incident
cataracts and cataract surgery in the Age-related Eye Disease Study
(AREDS): AREDS report number 32. Ophthalmology. 118:2113–2119.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hejtmancik JF and Kantorow M: Molecular
genetics of age-related cataract. Exp Eye Res. 79:3–9. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zheng LR, Ma JJ, Zhou DX, An LF and Zhang
YQ: Association between DNA repair genes (XPD and XRCC1)
polymorphisms and susceptibility to age-related cataract (ARC): A
meta-analysis. Graefes Arch Clin Exp Ophthalmol. 252:1259–1266.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Erol Tinaztepe Ö, Ay M and Eser E: Nuclear
and mitochondrial DNA of age-related cataract patients are
susceptible to oxidative damage. Curr Eye Res. 42:583–588. 2017.
View Article : Google Scholar
|
|
8
|
Sorte K, Sune P, Bhake A, Shivkumar VB,
Gangane N and Basak A: Quantitative assessment of DNA damage
directly in lens epithelial cells from senile cataract patients.
Mol Vis. 17:1–6. 2011.PubMed/NCBI
|
|
9
|
Cooke MS, Evans MD, Dizdaroglu M and Lunec
J: Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB
J. 17:1195–1214. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bruner SD, Norman DP and Verdine GL:
Structural basis for recognition and repair of the endogenous
mutagen 8-oxoguanine in DNA. Nature. 403:859–866. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu X, Lai W, Lin H and Liu Y: Association
of OGG1 and MTHFR polymorphisms with age-related cataract: A
systematic review and meta-analysis. PLoS One. 12:e01720922017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu XC, Guo XH, Chen B, Li ZH and Liu XF:
Association between the 8-oxoguanine DNA glycosylase gene Ser326Cys
polymorphism and age-related cataract: A systematic review and
meta-analysis. Int Ophthalmol. 38:1451–1457. 2018. View Article : Google Scholar
|
|
13
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Quinn JJ and Chang HY: Unique features of
long non-coding RNA biogenesis and function. Nat Rev Genet.
17:47–62. 2016. View Article : Google Scholar
|
|
16
|
Shen Y, Dong LF, Zhou RM, Yao J, Song YC,
Yang H, Jiang Q and Yan B: Role of long non-coding RNA MIAT in
proliferation, apoptosis and migration of lens epithelial cells: A
clinical and in vitro study. J Cell Mol Med. 20:537–548. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li G, Song H, Chen L, Yang W, Nan K and Lu
P: TUG1 promotes lens epithelial cell apoptosis by regulating
miR-421/caspase-3 axis in age-related cataract. Exp Cell Res.
356:20–27. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheng T, Xu M, Qin B, Wu J, Tu Y, Kang L,
Wang Y and Guan H: lncRNA H19 contributes to oxidative damage
repair in the early age-related cataract by regulating miR-29a/TDG
axis. J Cell Mol Med. 23:6131–6139. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xiang J, Chen Q, Kang L, Zhang G, Wang Y,
Qin B, Wu J, Zhou T, Han Y and Guan H: LncRNA PLCD3-OT1 functions
as a CeRNA to prevent age-related cataract by sponging miR-224-5p
and regulating PLCD3 expression. Invest Ophthalmol Vis Sci.
60:4670–4680. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chylack LT Jr, Wolfe JK, Singer DM, Leske
MC, Bullimore MA, Bailey IL, Friend J, McCarthy D and Wu SY: The
lens opacities classification system III. The longitudinal study of
cataract study group. Arch Ophthalmol. 111:831–836. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jao LE, Wente SR and Chen W: Efficient
multiplex biallelic zebrafish genome editing using a CRISPR
nuclease system. Proc Natl Acad Sci USA. 110:13904–13909. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Prior HM, Letwin K, Tuininga A and Nguyen
M: A simple method of cataract induction in adult zebrafish.
Zebrafish. 15:211–212. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
24
|
Ji Y, Cai L, Zheng T, Ye H, Rong X, Rao J
and Lu Y: The mechanism of UVB irradiation induced-apoptosis in
cataract. Mol Cell Biochem. 401:87–95. 2015. View Article : Google Scholar
|
|
25
|
Jia Y, Qin Q, Fang CP, Shen W, Sun TT,
Huang YL, Li WJ and Deng AM: UVB induces apoptosis via
downregulation of CALML3-dependent JNK1/2 and ERK1/2 pathways in
cataract. Int J Mol Med. 41:3041–3050. 2018.PubMed/NCBI
|
|
26
|
Carter RJ and Parsons JL: Base excision
repair, a pathway regulated by posttranslational modifications. Mol
Cell Biol. 36:1426–1437. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang AL, Lukas TJ, Yuan M and Neufeld AH:
Increased mitochondrial DNA damage and downregulation of DNA repair
enzymes in aged rodent retinal pigment epithelium and choroid. Mol
Vis. 14:644–651. 2008.PubMed/NCBI
|
|
28
|
Synowiec E, Blasiak J, Zaras M, Szaflik J
and Szaflik JP: Association between polymorphisms of the DNA base
excision repair genes MUTYH and hOGG1 and age-related macular
degeneration. Exp Eye Res. 98:58–66. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peng Q, Lu Y, Lao X, Chen Z, Li R, Sui J,
Qin X and Li S: Association between OGG1 Ser326Cys and APEX1
Asp148Glu polymorphisms and breast cancer risk: A meta-analysis.
Diagn Pathol. 9:1082014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yousaf S, Khan AU, Akram Z, Kayani MA,
Nadeem I, Begum B and Mahjabeen I: Expression deregulation of DNA
repair pathway genes in gastric cancer. Cancer Genet. 237:39–50.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Y, Li F, Zhang G, Kang L, Qin B and
Guan H: Altered DNA methylation and expression profiles of
8-Oxoguanine DNA glycosylase 1 in lens tissue from age-related
cataract patients. Curr Eye Res. 40:815–821. 2015. View Article : Google Scholar
|
|
32
|
Kang L, Zhao W, Zhang G, Wu J and Guan H:
Acetylated 8-oxoguanine DNA Glycosylase 1 and its relationship with
p300 and SIRT1 in lens epithelium cells from age-related cataract.
Exp Eye Res. 135:102–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Congrains A, Kamide K, Ohishi M and Rakugi
H: ANRIL: Molecular mechanisms and implications in human health.
Int J Mol Sci. 14:1278–1292. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu JY, Yao J, Li XM, Song YC, Wang XQ, Li
YJ, Yan B and Jiang Q: Pathogenic role of lncRNA-MALAT1 in
endothelial cell dysfunction in diabetes mellitus. Cell Death Dis.
5:e15062014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lu L, Yu X, Zhang L, Ding X, Pan H, Wen X,
Xu S, Xing Y, Fan J, Ge S, et al: The long non-coding RNA RHPN1-AS1
promotes uveal melanoma progression. Int J Mol Sci. 18:2262017.
View Article : Google Scholar :
|
|
36
|
Shang W, Yang Y, Zhang J and Wu Q: Long
noncoding RNA BDNF-AS is a potential biomarker and regulates cancer
development in human retinoblastoma. Biochem Biophys Res Commun.
497:1142–1148. 2018. View Article : Google Scholar
|
|
37
|
Liu Z, Zhang J, Gao J and Li Y:
MicroRNA-4728 mediated regulation of MAPK oncogenic signaling in
papillary thyroid carcinoma. Saudi J Biol Sci. 25:986–990. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Floros KV, Lochmann TL, Hu B, Monterrubio
C, Hughes MT, Wells JD, Morales CB, Ghotra MS, Costa C, Souers AJ,
et al: Coamplification of miR-4728 protects HER2-amplified breast
cancers from targeted therapy. Proc Natl Acad Sci USA.
115:E2594–E2603. 2018. View Article : Google Scholar
|
|
39
|
Chiang CC, Lin CL, Peng CL, Sung FC and
Tsai YY: Increased risk of cancer in patient with early-onset
cataracts: A nationwide population-based study. Cancer Sci.
105:431–436. 2014. View Article : Google Scholar : PubMed/NCBI
|