Introduction
Ovarian cancer (OC) is one of the most common
malignancies of the female reproductive organs. For instance, its
morbidity is second only to cervical cancer and corpus carcinoma;
however, the mortality of OC ranks first (1). The 5-year survival rates of OC are
~90 and ~15% in the early and advanced phases, respectively
(2). OC can be divided into
epithelial, germ and stromal cell neoplasms (3). Epithelial ovarian carcinoma (EOC),
which accounts for >70% of the cases of OC, has a 30% 5-year
survival rate and the highest mortality rate among all
gynecological tumors worldwide, and therefore, poses a serious
threat to women (4,5). As ovary tumor tissues are small and
reside deeply in the pelvic cavity, and patients lack typical
symptoms, the diagnosis of EOC during the early stage is very
difficult. Moreover, <30% of patients with EOC are reported to
have tumors confined to the ovary during surgery; the majority of
tumors spread to the pelvic and abdominal organs, and thus, early
diagnosis is important (6).
However, EOC typically presents at an advanced stage, leading to an
elevated mortality rate (7).
Considering the difficulty in early diagnosis and the distinctive
metastasis of EOC, investigating the biological and molecular
mechanisms of advance metastasis is crucial for the development of
effective therapeutic approaches against EOC (8).
MicroRNAs (miRNAs/miRs) are short, single-stranded,
non-coding RNAs present in different tissues and bodily fluids that
can regulate target gene expression post-transcriptionally.
Previous studies have reported that miRNAs are abnormally expressed
in various tumors and can act as tumor suppressors or oncogenes,
with roles in tumor development via negative regulation of specific
genes (9-11). For example, miR-195 has been
observed to be downregulated in EOC, and was revealed to decrease
cell proliferation and induce apoptosis of human OVCAR-3 cells by
inhibiting VEGFR2 and AKT signaling (12). In addition, miR-195 is
downregulated in human laryngeal squamous cell carcinoma, while the
overexpression of miR-195 inhibits cell viability, migration and
invasion, as well as promotes apoptosis of AMC-HN-8 cells (13). However, the roles of miR-195 in
the cell cycle, migration and invasion of EOC cells are yet to be
fully elucidated.
Cell division cycle 42 (CDC42) has been identified
as a target gene of miR-195 (14). miR-195 expression is downregulated
after overexpression of CDC42 in esophageal squamous cell carcinoma
(ESCC), and the combined aberrant expression of miR-195 and CDC42
could predict malignant progression and unfavorable prognosis of
ESCC (15). Furthermore,
down-regulated expression of miR-195, but upregulated cyclin D1
(CCND1) expression has been observed in papillary thyroid carcinoma
(PTC) cells, and miR-195 inhibits tumor growth and metastasis in
PTC by targeting CCND1 and fibroblast growth factor 2 (16). CDC42/CCND1 signaling pathways are
also involved in endothelial cell migration and proliferation
(17). However, it remains to be
determined whether the CDC42/CCND1 pathway serves a role in EOC,
and whether miR-195 acts as a tumor inhibitor in EOC by targeting
the CDC42/CCND1 signaling pathway.
The present study hypothesized that miR-195 may
suppress cell proliferation, migration and invasion in EOC via
inhibition of the CDC42/CCND1 pathway.
Materials and methods
Patients and tumor tissues
Human EOC tumor tissues and adjacent non-tumor
tissues were obtained from 78 patients (mean age, 52.33±9.87 years;
age range, 42-60 years) diagnosed with EOC via pathology, and who
had undergone surgical excision with no prior anticancer treatments
at The Second Hospital of Shanxi Medical University between January
2018 and December 2018. After resection, tissues were immediately
snap-frozen in liquid nitrogen and stored at −80°C for the
following experiments. The clinical cases of the patients were
acquired to analyze the association between miR-195, CDC42 and
CCND1 expression levels and age, differentiation degree, TNM
staging, serum CA12-5 content and International Federation of
Gynecology and Obstetrics (FIGO) staging. The present work was
approved by the Clinical Ethical Committee of The Second Hospital
of Shanxi Medical University and written informed consent was
obtained from all patients before the start of the study.
Cell culture
The wild-type human ovarian epithelial cell line
HS832.Tc, and human EOC cell lines OVCA420, OVCAR-3, A2780 and
SKOV3, were purchased from the Cell Bank of Chinese Academy of
Sciences. All cells were cultured in DMEM (Sigma-Aldrich; Merck
KGaA) or RPMI 1640 (Invitrogen; Thermo Fisher Scientific, Inc.)
with 10% FBS (Sigma-Aldrich; Merck KGaA) and 0.5% penicillin or
streptomycin and maintained in a 5% CO2 humidified
chamber at 37°C.
Reverse transcription-quantitative PCR
(RT-qPCR)
RT-qPCR was conducted to determine miR-195
expression in tissues and cells, as well as to determine CDC42 and
CCND1 mRNA expression levels in OVCAR-3 cells, as previously
described (12). Total RNA was
isolated using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) at room temperature, according to the
manufacturer's instructions. Following cDNA synthesis using the
TaqMan™ miRNA Reverse Transcription kit (Applied Biosystems; Thermo
Fisher Scientific, Inc.) at 65°C for 1 h. PCR was performed using
TaqMan™ miRNA assay (Invitrogen; Thermo Fisher Scientific, Inc.)
under the following thermocycling conditions for 42 cycles: Initial
denaturation at 95°C for 5 min, followed by 40 cycles of 94°C for
30 sec, 65°C for 30 sec, 72°C for 30 sec, and a final extension
step at 72°C for 5 min. Relevant primer sequences: hsa-miR-195
forward, 5′-ACA CTC CAG CTG GGT AGC AGC ACA GAA AT-3′ and reverse,
5′-TGGTGTCGTGGAGTCG-3′; human U6 (used as an internal control of
miR-195) forward, 5′-CTC GCT TCG GCA GCA CA-3′ and reverse, 5′-AAC
GCT TCA CGA ATT TGC GT-3′; human CDC42 forward, 5′-GCC CGT GAC CTG
AAG GCT GTCA-3′ and reverse, 5′-TGC TTT TAG TAT GAT GCC GAC
ACCA-3′; human CCND1 forward, 5′-TCC TAC TAC CGC CTC ACA-3′ and
reverse, 5′-ACC TCC TCC TCC TCC TCT-3′; and human GAPDH (used as an
internal control of CDC42 and CCND1) forward, 5′-AAG GTG AAG GTC
GGA GTCA-3′ and reverse, 5′-GGA AGA TGG TGA TGG GAT TT-3′. Relative
quantification was performed using the 2−ΔΔCq method
(18).
Western blotting
Western blotting was performed to detect the protein
expression levels of CDC42 and CCND1 from tissues and cells, as
previously described (13). Total
proteins were extracted with RIPA buffer (Beyotime Institute of
Biotechnology). The BCA kit (Beyotime Institute of Biotechnology)
was used to determine the concentration. In total, 80 µg of
each sample were resolved using 10% SDS-PAGE at 75 V for 2 h. After
transfer to PVDF membranes at 350 mA for 2 h, membranes were
blocked with 5% skim milk diluted in TBS for 1 h at room
temperature. The membranes were probed overnight at 4°C with the
following primary antibodies: Monoclonal anti-CDC42 (rabbit;
1:1,000; cat. no. 2466S; Cell Signaling Technology, Inc.),
monoclonal anti-CCND1 (rabbit; 1:1,000; cat. no. 3300S; Cell
Signaling Technology, Inc.) and monoclonal anti-GAPDH (rabbit;
1:1,000; cat. no. 5174S; Cell Signaling Technology, Inc.; used as
an internal control). After probing with horseradish peroxidase
(HRP)-conjugated secondary antibody (goat anti-rabbit IgG; 1:5,000;
cat. no. TA130059; OriGene Technologies, Inc.) at room temperature
for 1 h, proteins were detected using an enhanced chemiluminescence
reagent (Bio-Rad Laboratories, Inc.), and an Alpha Innotech
instrument (Bio-Rad Laboratories, Inc.) was used to scan and
quantify the images. GAPDH was used to normalize the signals.
Immunohistochemistry
The human EOC tumor tissues and adjacent non-tumor
tissues were collected, fixed with 4% paraformaldehyde at 25°C for
24 h and dehydrated with a sucrose solution (gradient, 10, 20 and
30%) at room temperature. After the tissues were harvested and
dehydrated with 30% sucrose overnight, these were sectioned at
15-µm thickness using a freezing microtome (Leica CM1900;
Leica Microsystems GmbH). The sections were washed three times for
5 min each time in 0.01 M PBS. The endogenous oxidase activity was
blocked with 3% hydrogen peroxide at 25°C in a closed environment
for 15 min. Then, sections were washed three times (5 min/time)
with PBS and incubated with 2% goat serum (Sigma-Aldrich; Merck
KGaA) at 37°C in a wet box for 20 min. The antigen retrieval
protocol was performed at 95°C in a microwave for 15 min with 0.01
M citrate buffer (pH 6.0). The tissue sections were then incubated
for 30 min at 37°C in a water bath, followed by overnight
incubation at 4°C with a polyclonal CDC42 antibody (rabbit; 1:300;
cat. no. 2462S, Santa Cruz Biotechnology, Inc.) or a polyclonal
CCND1 antibody (rabbit; 1:200; cat. no. 12531S, Santa Cruz
Biotechnology, Inc.). After washing with 0.1 mol/l PBS three times
for 5 min, the sections were incubated for 30 min at 37°C with
HRP-conjugated goat anti-rabbit IgG secondary antibody (1:5,000;
cat. no. W10814; Thermo Fisher Scientific, Inc.), followed by three
washes for 5 min with 0.1 mol/l PBS and added liquid detergent. The
3,3′-diaminobenzidine staining reaction was performed at room
temperature for 30 sec, followed by the addition of hematoxylin at
room temperature for 15 min for counterstaining and differentiation
for 1 sec. The negative control was PBS. After the slides were
naturally dried, sealed the slides with neutral resin (Beyotime
Institute of Biotechnology). Observation was performed under light
microscope at ×40 magnification and analysis was performed using
ImageJ software (v1.8.0, National Institutes of Health).
RNA interference and transfection
assays
For overexpression or knockdown of miR-195, miR-195
mimic (5′-UAG CAG CAC AGA AAU AUU GGC-3′), miR-195 mimic NC (5′-UUC
UCC GAA CGU GUC ACG UTT-3′), miR-195 inhibitor (5′-GCC AAU AUU UCU
GUG CUG CUA-3′) and inhibitor-NC (5′-CAA UAU UUC UGU GCU GCU AUU-3′
all were from Shanghai GenePharma Co., Ltd. Transfection into
OVCAR-3 cells were performed using Lipofectamine® 2000
Transfection reagent (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's instructions.
To generate OVCAR-3 cells overexpressing CDC42 or
CCND1, pcDNA 3.1-CDC42 and pcDNA 3.1-CCND1 (Generay Biotech Co.,
Ltd.) were cloned into a pcDNA 3.1 vector (Promega Corporation) and
transfected into OVCAR-3 cells at 10 nM concentration.
Lipofectamine® 2000 Transfection reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) was used to mediate the
transfection. The empty vector was used as the control. To
knockdown CDC42 or CCND1 expression in OVCAR-3 cells, HS_CDC42
small interfering (si)RNA (5′-CAT CAG ATT TTG AAA ATA TTT AA-3′),
HS_CCND1 siRNA (5′-GCC ACA GAT GTG AAG TTC A-3′) and NC siRNA
(5′-AAT TCT CCG AAC GTG TCA CGT-3′), provided by GeneCopoeia, Inc.,
were transfected into OVCAR-3 cells using Lipofectamine RNAiMAX
(Invitrogen; Thermo Fisher Scientific, Inc.) at 50 nM, according to
the manufacturer's instructions.
Cell proliferation evaluation
Cell proliferation was evaluated using a
Bromodeoxyuridine (BrdU) incorporation assay and a colony formation
assay. The BrdU incorporation assay was performed at 24 h following
transfection of OVCAR-3 cells, as previous reported (19). Colony formation was performed at
48 h after transfection, as previous reported (20).
Cell cycle and cell apoptosis assay
After transfection for 48 h, cell cycle and
apoptosis were detected using flow cytometry (21). The cell cycle was detected using
20 µg/ml PI (Becton, Dickinson and Company) at 37°C for 30
min in dark. Cell apoptosis was detected using a BD apoptosis assay
kit (Becton, Dickinson and Company) according to the manufacturer's
instruction. A flow cytometer (Flow Sight; Merck KGaA) was used to
assess cell cycle distribution and cell apoptosis. Cell cycle was
analyzed using a FacsCalibur system (Cytek Development Inc.).
FlowJo software (v10.0.7; Becton, Dickinson and Company) was used
to analyze the percentage of apoptotic cells.
Cell migration and invasion assay
A total of 24 h after transfection, cell migration
was assessed using the wound healing assay. Cells were scrapped
vertically using pipette head tips. Then, cell were washed with PBS
and cultured with serum-free medium at 37°C for 24 h.
A total of 48 h after transfection, cell invasion
was measured using a Transwell invasion assay, as described in a
previous study (22). Wound
migration were observed under a light microscope at ×40
magnification and cell invasion were observed under a light
microscope at ×200 magnification. Migrated cell and invaded cell
were analyzed using ImageJ software (v1.8.0, National Institutes of
Health).
Statistical analysis
SPSS 15.0 software (SPSS, Inc.) was used for
statistical analyses. Each experiment was repeated three times.
Data are presented as the mean ± standard deviation. One-way ANOVA
followed by Tukey's multiple comparison test was used to compare
the data between groups. Correlation between miR-195 and CDC42 or
CCND1 was analyzed using Pearson's correlation coefficient.
Expression levels of miR-195, CDC42 and CCND1 in healthy tissues
and EOC tissues were compared using a paired t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression levels of miR-195, CDC42 and
CCND1 in EOC tissues and cells
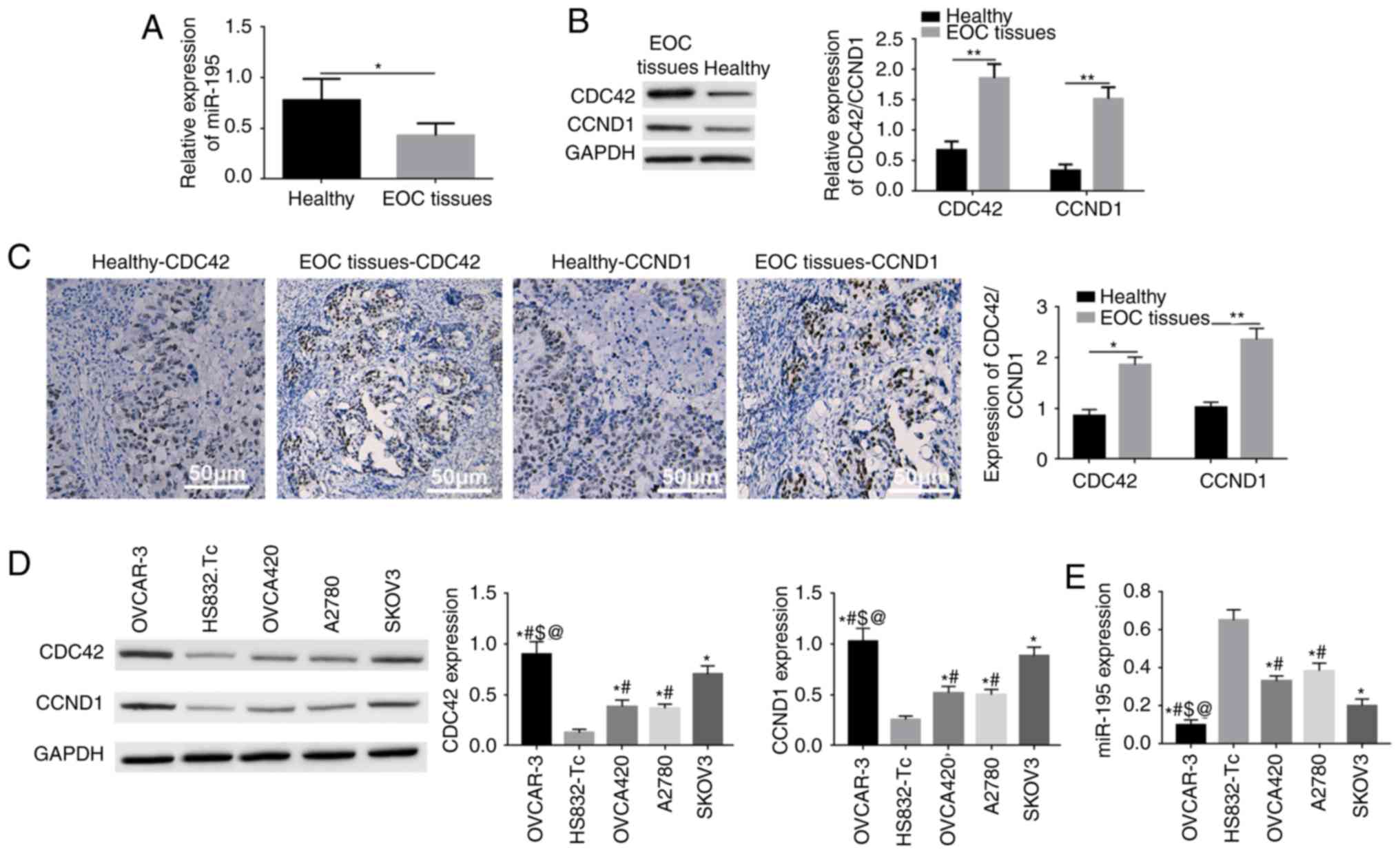
RT-qPCR results demonstrated that miR-195 was
significantly downregulated in EOC tissues (Fig. 1A). Moreover, western blotting and
immunohistochemistry results indicated that CDC42 and CCND1 were
significantly upregulated in EOC tissues (Fig. 1B and C). It was identified that
the downregulation of miR-195 or increased expression levels of
CDC42 and CCND1 were associated with a reduced degree of
differentiation, advanced clinical TNM stage, higher serum CA125
content and advanced FIGO staging (Table I).
 | Table IAssociations of miR-195, CDC42 and
CCND1 expression levels with the clinicopathological features in 78
patients with epithelial ovarian cancer. |
Table I
Associations of miR-195, CDC42 and
CCND1 expression levels with the clinicopathological features in 78
patients with epithelial ovarian cancer.
| Clinicopathologic
parameters | Number | miR-195
expression | P-value | CDC42
expression | P-value | CCND1
expression | P-value |
|---|
| Age, years | | | | | | | |
| 40-49 | 38 | 0.46±0.09 | 0.925 | 1.78±0.18 | 0.738 | 1.62±0.17 | 0.622 |
| 50-60 | 40 | 0.44±0.07 | | 1.81±0.24 | | 1.58±0.14 | |
| Differentiation
degree | | | | | | | |
| Low | 26 | 0.34±0.06 | <0.01 | 1.88±0.26 | <0.01 | 1.74±0.16 | <0.01 |
| Middle | 23 | 0.41±0.08 | | 1.79±0.18 | | 1.63±0.15 | |
| High | 29 | 0.55±0.10 | | 1.62±0.13 | | 1.52±0.13 | |
| TNM stage | | | | | | | |
| I | 23 | 0.61±0.12 | <0.01 | 1.63±0.14 | <0.01 | 1.53±0.13 | <0.01 |
| II | 36 | 0.54±0.10 | | 1.76±0.16 | | 1.66±0.16 | |
| III-IV | 19 | 0.38±0.07 | | 1.92±0.28 | | 1.78±0.17 | |
| Serum CA125 content
(U/ml) | | | | | | | |
| >83 | 53 | 0.37±0.07 | 0.007 | 1.86±0.25 | 0.003 | 1.77±0.15 | 0.004 |
| <83 | 25 | 0.63±0.13 | | 1.58±0.11 | | 1.52±0.12 | |
| FIGO staging | | | | | | | |
| I | 23 | 0.61±0.12 | <0.01 | 1.63±0.14 | <0.01 | 1.53±0.13 | <0.01 |
| II | 35 | 0.53±0.09 | | 1.77±0.15 | | 1.64±0.15 | |
| III | 15 | 0.42±0.08 | | 1.85±0.15 | | 1.76±0.16 | |
| IV | 5 | 0.37±0.08 | | 2.02±0.20 | | 1.95±0.18 | |
RT-qPCR results suggested that miR-195 was
downregulated in EOC tissues, while western blotting results
identified that CDC42 and CCND1 were upregulated in various EOC
cell lines, including OVCA420, OVCAR-3, A2780 and SKOV3 cells.
Additionally, the lowest expression of miR-195 and the highest
expression levels of CDC42 and CCND1 occurred in OVCAR-3 cells
(Fig. 1D and E). Thus, subsequent
experiments were performed in OVCAR-3 cells.
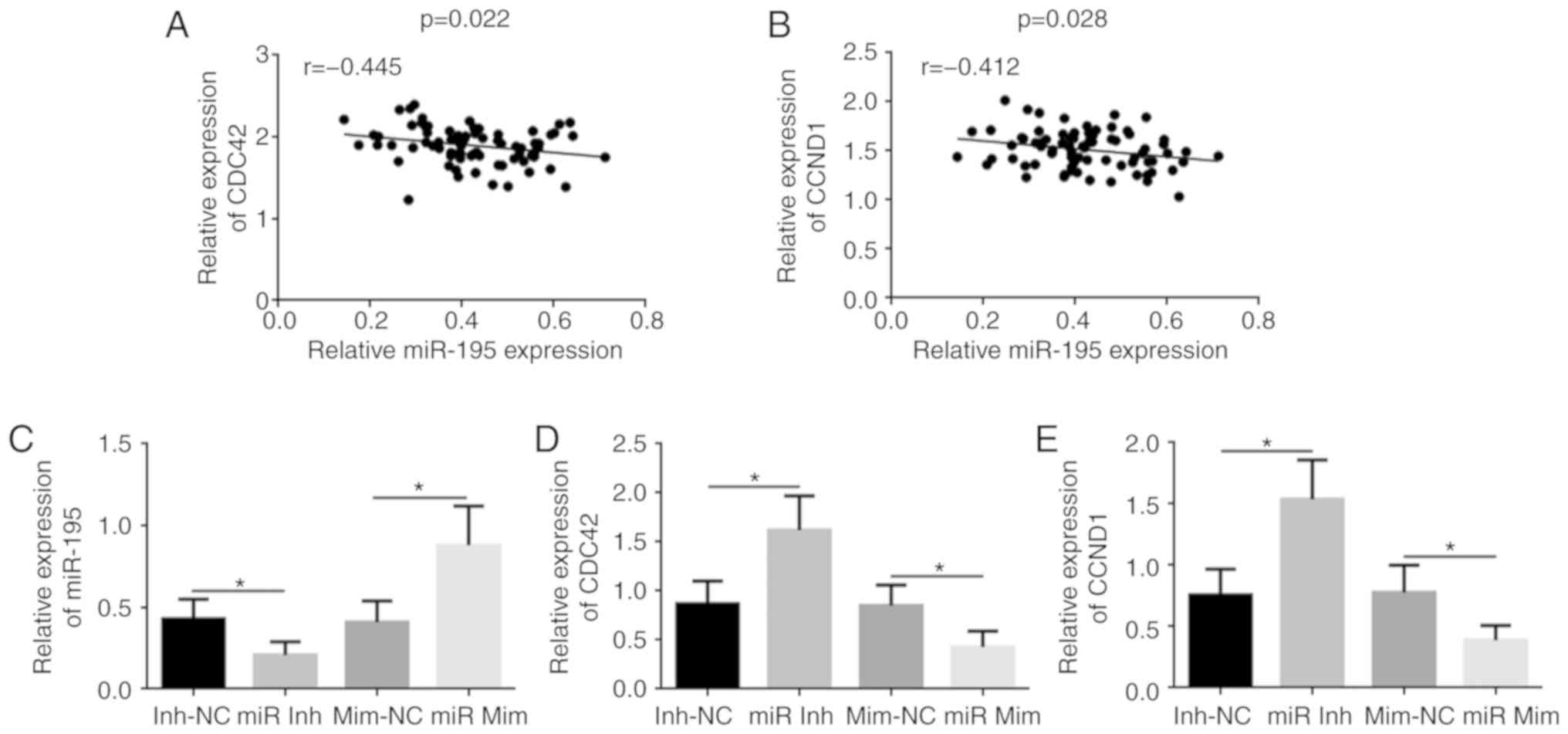
miR-195 could regulate the expression
levels of CDC42 and CCND1
It was found that the miR-195 mRNA expression was
negatively correlated with CDC42 and CCND1 protein expression
levels in human EOC tissues (Fig. 2A
and B). Transfection of the miR-195 mimic led to significantly
increased expression of miR-195 in OVCAR-3 cells compared with the
mimic-NC group, while the miR-195 inhibitor significantly decreased
miR-195 expression compared with the miR-195 inhibitor-NC group
(Fig. 2C). miR-195 knockdown
resulted in the upregulation of CDC42 and CCND1, while miR-195
overexpression caused downregulation of CDC42 and CCND1 in OVCAR-3
cells (Fig. 2D and E).
miR-195 represses cell proliferation by
inhibiting CDC42 and CCND1 expression levels in OVCAR-3 cells in
vitro
In order to investigate whether the effect of
miR-195 in cell proliferation and apoptosis was CDC42/CCND1
dependent, OVCAR-3 cells overexpressing CDC42 or CCND1 and knocking
down CDC42 or CCND1 in OVCAR-3 cells were generated (Fig. S1). RT-qPCR results indicated that
knockdown of miR-195 significantly promoted CDC42 and CCND1
expression levels, but transfection with CDC42 or CCND1 siRNA
reversed this trend, and induced downregulation of CDC42 or CCND1
in OVCAR-3 cells (Fig. 3A and B).
By contrast, overexpression of miR-195 significantly reduced CDC42
and CCND1 expression levels, but transfection with a CDC42 or CCND1
overexpression vector resulted in upregulation of CDC42 and CCND1
(Fig. 3C and D).
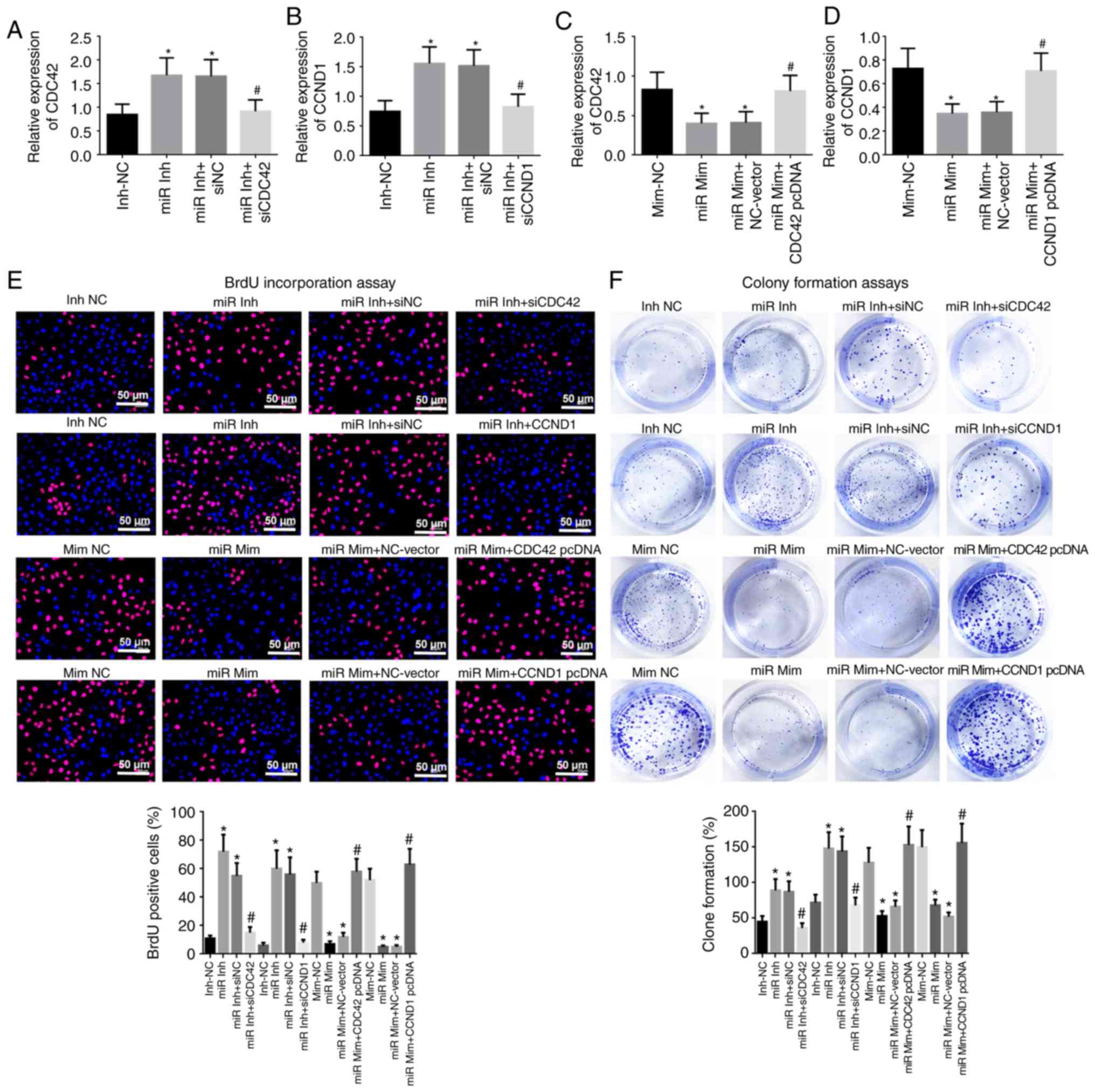 | Figure 3miR-195 inhibits cell proliferation by
reducing CDC42 and CCND1 expression levels in OVCAR-3 cells in
vitro. mRNA expression levels of (A) CDC42 and (B) CCND1 in
OVCAR-3 cells transfected with miR-195 inhibitor, inhibitor-NC,
miR-195 inhibitor + NC and miR-195 inhibitor + CDC42 or CCND1
siRNAs. mRNA expression levels of (C) CDC42 and (D) CCND1 in
OVCAR-3 cells transfected with miR-195 mimic, mimic-NC, miR-195
mimic + NC-vector and miR-195 mimic + CDC42 or CCND1 pcDNA. (E)
Cell proliferation was assessed using BrdU immunostaining at 24 h
after transfection. Red, OVCAR-3 cells labeled with BrdU; blue,
nuclei counterstained by BrdU. Scale bar, 50 µm. (F)
Evaluation of cell proliferation via colony formation assays at 48
h after transfection. Data were analyzed using one-way ANOVA.
*P<0.05 vs. Inh-NC or Mim-NC groups;
#P<0.05 vs. miR Inh-siNC or miR Mim + NC-Vector
groups. BrdU, 5-bromo-2-deoxyuridine; miR, miR-195; Inh, inhibitor;
NC, negative control, siRNA, short interfering RNA; Mim, mimic;
CDC42, Cell division cycle 42; CCND1, cyclin D1. |
Knockdown of miR-195 resulted in enhanced cell
proliferation, with significantly increased percentages of
BrdU-positive cells and clone formation; however, this enhanced
cell proliferative ability was significantly reversed after CDC42
or CCND1 siRNA co-transfection (Fig.
3E and F). Overexpression of miR-195 and co-transfection with
miR-195 mimic and the CDC42 or CCND1 overexpression vector
demonstrated opposite outcomes to treatment with miR-195 inhibitor
and CDC42 or CCND1 siRNAs (Fig. 3E
and F). These findings indicated that miR-195 repressed cell
proliferation by inhibiting CDC42 and CCND1 expression levels in
OVCAR-3 cells in vitro.
miR-195 inhibits cell cycle entry and
promotes apoptosis in OVCAR-3 cells by downregulating CDC42 and
CCND1 expression levels in vitro
Flow cytometry with PI staining identified that the
miR-195 inhibitor increased cell cycle entry, which arrested
OVCAR-3 cells at S and G2 phases, with significantly
fewer EOC cells appearing in G1 phase (Fig. 4). However, following
co-transfection with CDC42 or CCND1 siRNA, cell cycle entry was
inhibited, with reduced S and G2 phase cells and
increased G1 phase OVCAR-3 cells compared with cells
transfected with NC siRNA. Additionally, overexpression of miR-195
restrained OVCAR-3 cell cycle entry, while overexpression of CDC42
or CCND1 reversed this pattern and facilitated cell cycle entry
(Fig. 4).
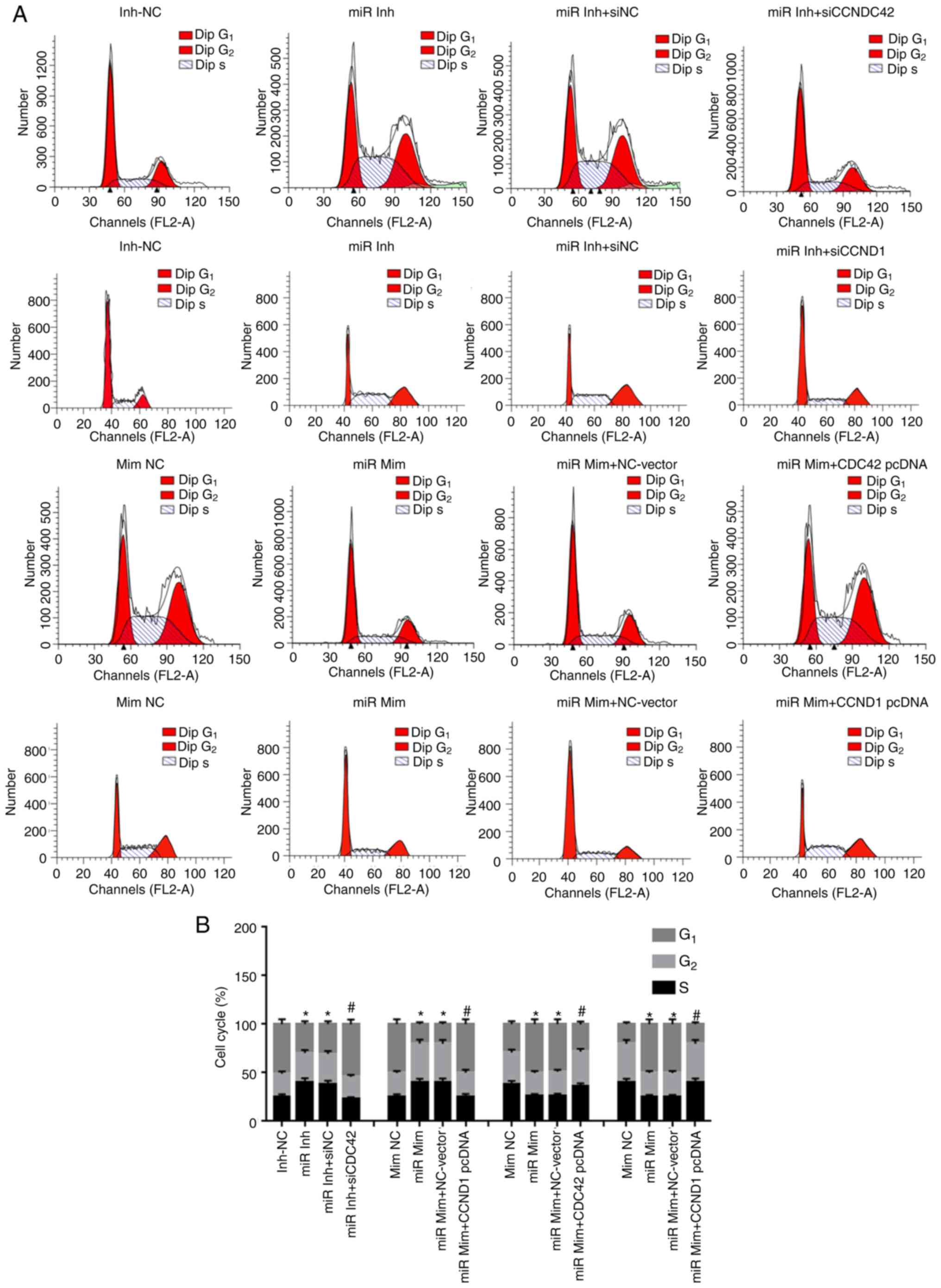 | Figure 4miR-195 inhibits cell cycle entry in
OVCAR-3 cells by downregulating CDC42 and CCND1 expression levels
in vitro. (A) Effects of miR-195, CDC42 and CCND1 on the
percentage of PI-stained OVCAR-3 cells at G1, S and
G2 phases. (B) Cell cycle of each group. Data were
analyzed via one-way ANOVA. *P<0.05 vs. Inh-NC or
Mim-NC groups; #P<0.05 vs. miR Inh-siNC or miR Mim +
NC-Vector groups. miR, miR-195; Inh, inhibitor; NC, negative
control, siRNA, short interfering RNA; Mim, mimic; CDC42, Cell
division cycle 42; CCND1, cyclin D1. |
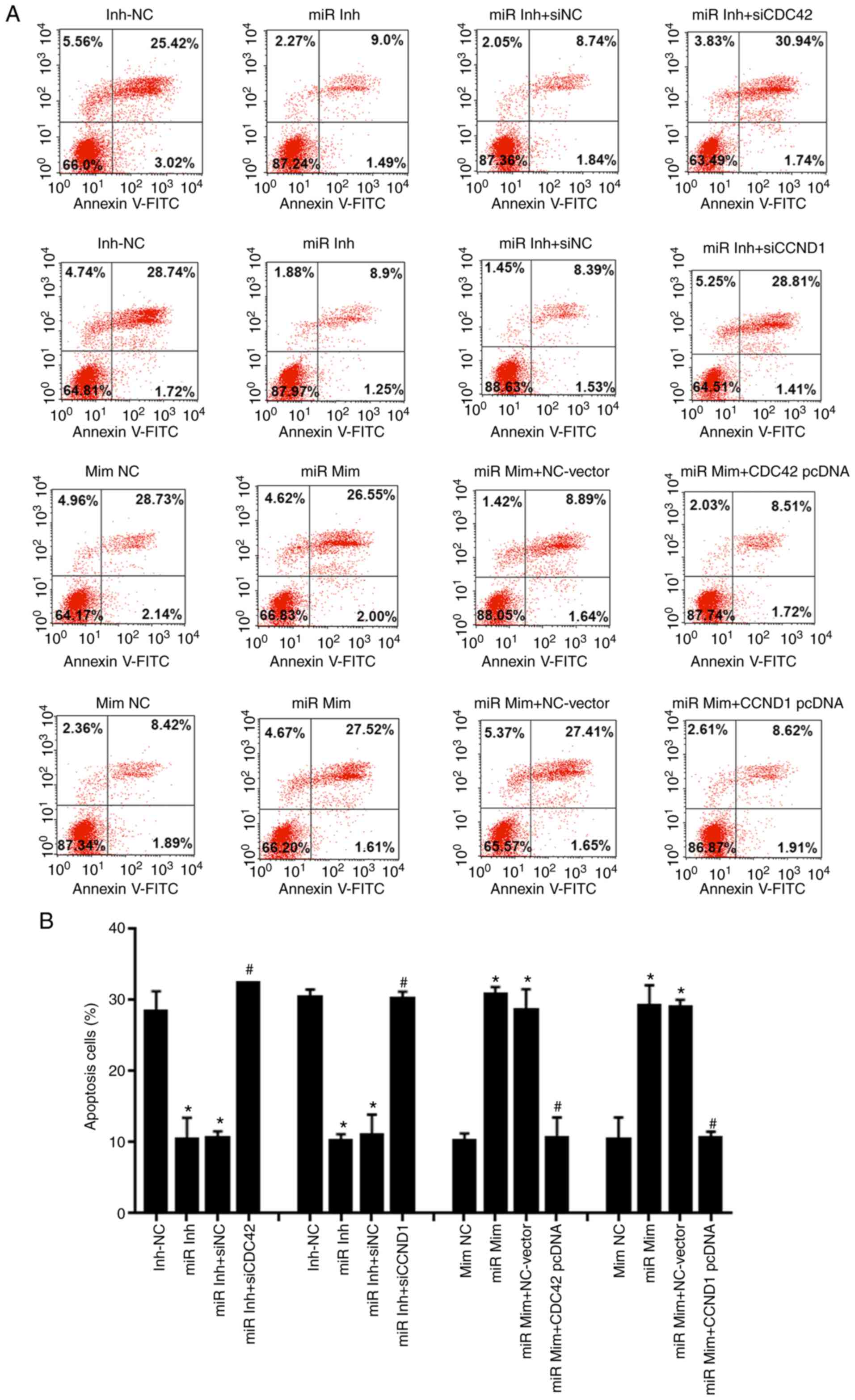
Flow cytometry with Annexin V-FITC/PI staining
suggested that knockdown of miR-195 reduced apoptosis of OVCAR-3
cells in vitro compared with the miR-195 inhibitor-NC group.
Conversely, following co-transfection with CDC42 or CCND1 siRNA,
the apoptotic rate was significantly increased compared with the NC
siRNA (Fig. 5). Moreover,
transfection with miR-195 mimic and overexpression vectors of CDC42
or CCND1 exerted the opposite effects to the miR-195 inhibitor and
CDC42 or CCND1 siRNAs (Fig.
5).
miR-195 suppresses migration and invasion
in OVCAR-3 cells by inhibiting CDC42 and CCND1 expression levels in
vitro
The wound healing assay results demonstrated that
knock-down of miR-195 caused increased OVCAR-3 cell migration,
compared with the miR-195 inhibitor-NC group. Furthermore, OVCAR-3
cells migrated toward the wound more slowly following
co-transfection with CDC42 siRNA compared with cells transfected
with the NC siRNA. The migration of OVCAR-3 cells was significantly
weakened after overexpression of miR-195 and reversed following
transfection with the CDC42 or CCND1 overexpression vector
(Fig. 6A and B).
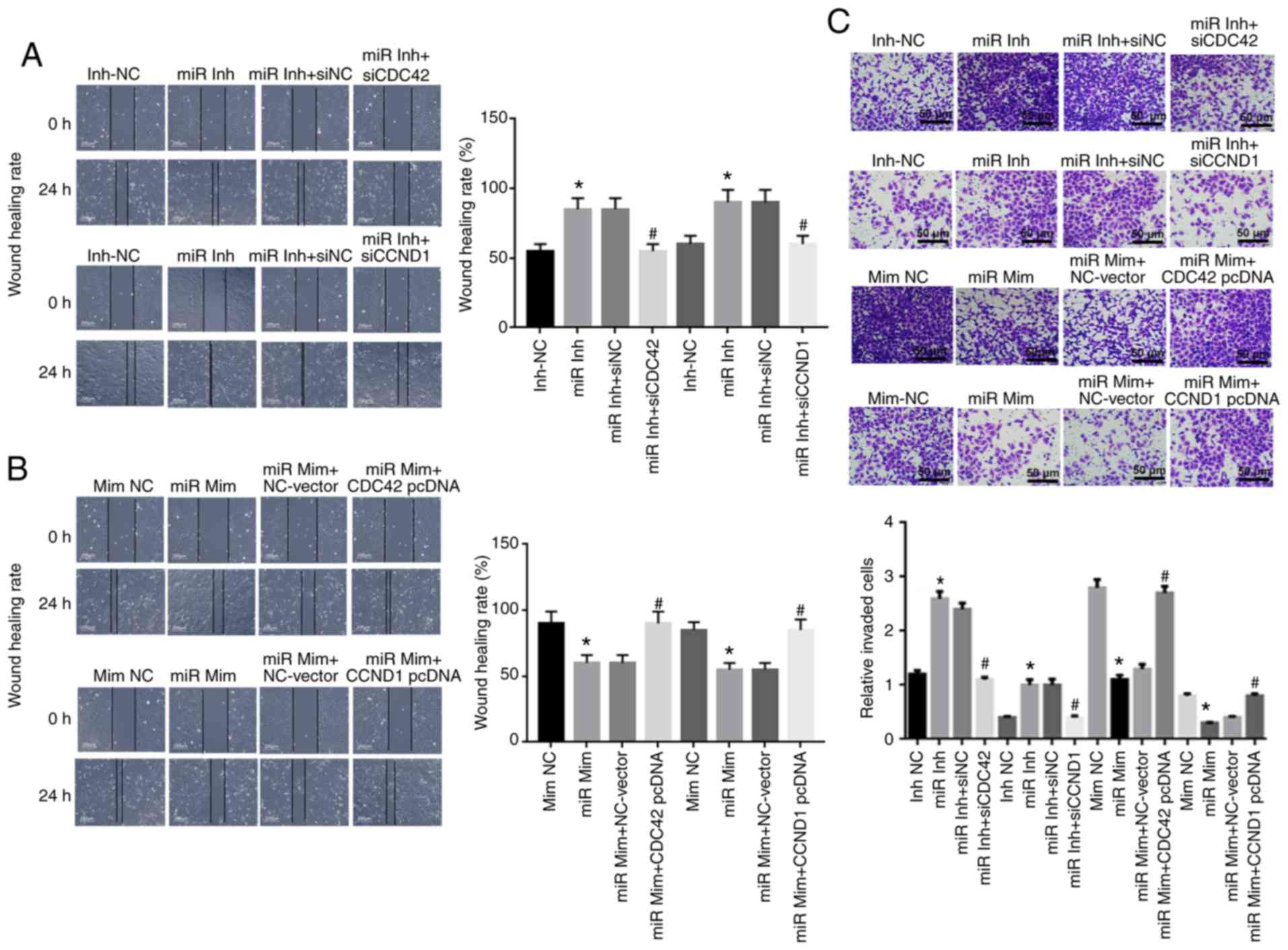 | Figure 6miR-195 suppresses cell migration and
invasion in OVCAR-3 cells by inhibiting CDC42 and CCND1 expression
levels in vitro. Effects of miR-195 (A) inhibitor or (B)
mimic, CDC42 and CCND1 on OVCAR-3 cell migration using wound
healing assay at 0 and 24 h after transfection. Scale bar, 200
µm. (C) Effects of miR-195, CDC42 and CCND1 on OVCAR-3 cell
invasion via Transwell invasion assay at 48 h after transfection.
Scale bar, 50 µm. Data were analyzed using one-way ANOVA.
*P<0.05 vs. Inh-NC or Mim-NC groups;
#P<0.05 vs. miR Inh-siNC or miR Mim + NC-Vector
groups. miR, miR-195; Inh, inhibitor; NC, negative control, siRNA,
short interfering RNA; Mim, mimic; CDC42, Cell division cycle 42;
CCND1, cyclin D1. |
The Transwell invasion assay identified a
significant increase in the number of invading OVCAR-3 cells in the
miR-195 inhibitor group compared with the miR-195 inhibitor-NC
group; however, the number of invading OVCAR-3 cells was reduced
following co-transfection with CDC42 or CCND1 siRNA compared with
NC siRNA (Fig. 6C). In addition,
the miR-195 mimic induced a significant decrease in invading cells
compared with the NC mimic, while a higher number of invading
OVCAR-3 cells was observed in the group co-transfected with the
CDC42 or CCND1 overexpression vector compared with the NC-vector
group (Fig. 6B).
Discussion
EOC is the most frequent histopathological type of
OC, with a 5-year survival rate of ~30% and the highest mortality
rate of all gynecological tumors worldwide (1). Thus, EOC is a serious threat to the
lives of women. As EOC is difficult to diagnose at an early stage,
there are limited treatment options and high rates of metastasis
and recurrence following operative therapy (7,8).
Therefore, the identification of biological targets for the
diagnosis and treatment of EOC is an urgent issue. In the present
study, miR-195 expression was found to be downregulated in human
EOC tissues and multifold cells, particularly in OVCAR-3 cells,
compared with the corresponding control groups. Moreover, this
downregulation of miR-195 was closely associated with reduced
differentiation, higher clinical TNM stage and increased serum EOC
marker/CA125 content. Thus, miR-195 may be a potential target of
EOC.
Previous studies have reported that aberrant
expression of miRNAs may be a marker of the development and
progression of various tumors or cancer types, as they are
associated with abnormal cell viability, proliferation, migration,
invasion, apoptosis and differentiation (23-25). The present results suggested that
miR-195 was downregulated in human EOC tissues and cells, which led
to an increase in cell proliferation, cell cycle entry, migration
and invasion, as well as a decrease in apoptosis of OVCAR-3 cells
in vitro. Another published report regarding the role of
miR-195 in OC indicated that miR-195 was downregulated in human OC
serum samples, which resulted in an increase in cell proliferation
and decreased apoptosis of human OVCAR-3 cells, which was in line
with the current findings that knockdown of miR-195 induced an
increase in cell proliferation and a decrease in cell apoptosis.
Previous studies have also observed downregulation of miR-195 in
other cancer types. For example, decreased miR-195 expression in
hepatocellular carcinoma tissues significantly promotes
proliferation, invasion and tumorigenesis (26). Moreover, miR-195 represses cell
proliferation, migration and invasion, and accelerates apoptosis in
glioma by decreasing spalt like transcription factor 4 protein
expression (27). Downregulation
of miR-195 in colon carcinoma tissues is also associated with
increased cell proliferation, migration and invasion (28). Furthermore, miR-195 is
downregulated in PTC tissues and cells, which facilitates cell
cycle progression by inducing apoptosis (29). These previous findings were in
accordance with the present results, and to the best of our
knowledge, the current study was the first to demonstrate that
miR-195 is downregulated in human EOC tissues and cells, and is
associated with the CDC42/CCND1 pathway.
The present results suggested that the molecular
mechanism of miR-195 in EOC was achieved via inhibition of the
CDC42/CCND1 pathway. Previous studies have revealed that CDC42 and
CCND1 are targets of miR-195 in ESCC and PTC, respectively
(15,16). Thus, the present study did not
perform a luciferase reporter assay. The current results indicated
that expression levels of CDC42 and CCND1 were negatively
correlated with miR-195 in EOC tissues. In addition, CDC42 and
CCND1 had a corresponding opposite effect following interference of
miR-195 expression, which indirectly suggested that CDC42 and CCND1
are components of a downstream pathway of miR-195. In the present
study, knock-down of miR-195 effectively increased cell
proliferation and migration, decreased cell apoptosis and elevated
CDC42 and CCND1 protein expression levels. A previous study
identified that inhibition of CDC42 expression may be a key
mechanism via which miR-18a impairs cancer cell proliferation
(30). In the current study,
overexpression of miR-195 significantly decreased cell
proliferation and migration, promoted cell apoptosis and reduced
CDC42 and CCND1 protein expression levels; these effects were
reversed by CDC42 and CCND1 overexpression. The CDC42/CCND1
signaling pathway has been reported to promote cell migration and
proliferation in endothelial cells. In addition, activation of the
CDC42/CCND1 pathway could induce cell proliferation, migration and
invasion to increase Ewing sarcoma metastasis (31). Overexpression of CDC42 and CCND1
has also been shown to cause increased cell migration, changes in
cell morphology, G1/S phase cell cycle progression and
decreased apoptosis in colorectal cancer cells (30). Collectively, these previous
findings are in line with the present results, which found that
overexpression of CDC42 and CCND1 was associated with enhanced cell
proliferation, cell cycle progression, migration and invasion, as
well as attenuated apoptosis in EOC cells. Therefore, miR-195 may
suppress cell proliferation, migration and invasion in EOC via
inhibition of the CDC42/CCND1 pathway.
There are limitations to the present study. First, a
luciferase assay was not performed to directly confirm the
relationship between miR-195 and CDC42. At present, evidence that
CCND1 can act as a target of miR-195 has been reported in ESCC
(15). The present results can
only indirectly demonstrate that miR-195 could target CDC42 via
assessing the reverse effect of CDC42 and CCND1 siRNA or CDC42 and
CCND1 overexpression. Secondly, western blotting and RT-qPCR were
not conducted to measure markers of migration, invasion and
apoptosis in cells. Finally, the present study did not investigate
AKT and its inhibitors on the effect of miR-195. A previous study
revealed that miR-195 inhibited VEGFR2 and AKT signaling (12); therefore, further evidence
regarding the under-lying molecular mechanism of miR-195 is
required.
In conclusion, the present study demonstrated that
miR-195 was downregulated, while CDC42 and CCND1 were upregulated,
in EOC tissues and cells, which led to increased malignancy of EOC.
miR-195 can repress human EOC cell cycle entry, migration and
invasion, and CDC42 and CCND1 serve as tumor promoters in EOC
progression. Furthermore, it was identified that inhibition of the
CDC42/CCND1 pathway via overexpression of miR-195 may be a possible
mechanism for suppression of EOC progression. Thus, miR-195 may be
an effective agent for EOC therapy and diagnosis via
anti-proliferative, migration-inhibiting and pro-apoptotic effects
via inhibition of the CDC42/CCND1 signaling pathway.
Supplementary Data
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XH made substantial contributions to the conception
and design of the study. QJ and JY performed the experiments. XS
and HG participated in this experiment, and analyzed the data. JG
contributed to acquisition of data. YG organized the figure and
interpreted the data. XH reviewed and edited the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Clinical
Ethical Committee of The Second Hospital of Shanxi Medical
University, and written informed consent was obtained from all
patients before the start of the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Li X, Du N, Zhang Q, Li J, Chen X, Liu X,
Hu Y, Qin W, Shen N, Xu C, et al: MicroRNA-30d regulates
cardiomyocyte pyroptosis by directly targeting foxo3a in diabetic
cardiomyopathy. Cell Death Dis. 5:e14792014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parizadeh SM, Jafarzadeh-Esfehani R,
Ghandehari M, Hasanzadeh M, Parizadeh SMR, Hassanian SM,
Rezaei-Kalat A, Aghabozorgi AS, Rahimi-Kakhki R, Zargaran B, et al:
Circulating and tissue microRNAs as biomarkers for ovarian cancer
prognosis. Curr Drug Targets. 20:1447–1460. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mülayim B, Gürakan H, Dagli V, Mülayim S,
Aydin O and Akkaya H: Unaware of a giant serous cyst adenoma: A
case report. Arch Gynecol Obstet. 273:381–383. 2006. View Article : Google Scholar
|
|
4
|
Menke K, Schwermer M, Falke K, Felenda J,
Beckmann C, Stintzing F, Voigt A, Schramm A and Zuzak TJ: Taraxacum
officinale extract induces antitumorigenic effects in ovarian
carcinoma cell lines. Eur J Gynaecol Oncol. 40:106–112. 2019.
|
|
5
|
Ferlay J, Steliarova-Foucher E,
Lortet-Tieulent J, Rosso S, Coebergh JW, Comber H, Forman D and
Bray F: Cancer incidence and mortality patterns in Europe Estimates
for 40 countries in 2012. Eur J Cancer. 49:1374–1403. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lim W and Song G: Discovery of prognostic
factors for diagnosis and treatment of epithelial-derived ovarian
cancer from laying hens. J Cancer Prev. 18:209–220. 2013.
View Article : Google Scholar
|
|
7
|
Lheureux S, Gourley C, Vergote I and Oza
AM: Epithelial ovarian cancer. Lancet. 393:1240–1253. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Motohara T and Katabuchi H: Ovarian cancer
stemness: Biological and clinical implications for metastasis and
chemotherapy resistance. Cancers (Basel). 11:9072019. View Article : Google Scholar
|
|
9
|
Kagiya T: MicroRNAs: Potential biomarkers
and therapeutic targets for alveolar bone loss in periodontal
disease. Int J Mol Sci. 17:13172016. View Article : Google Scholar :
|
|
10
|
Piletič K and Kunej T: MicroRNA epigenetic
signatures in human disease. Arch Toxicol. 90:2405–2419. 2016.
View Article : Google Scholar
|
|
11
|
Varamo C, Occelli M, Vivenza D, Merlano M
and Lo Nigro C: MicroRNAs role as potential biomarkers and key
regulators in melanoma. Genes Chromosomes Cancer. 56:3–10. 2017.
View Article : Google Scholar
|
|
12
|
Chen J: miRNA-195 suppresses cell
proliferation of ovarian cancer cell by regulating VEGFR2 and AKT
signaling pathways. Mol Med Rep. 18:1666–1673. 2018.PubMed/NCBI
|
|
13
|
Pang H, Xu X, Dai L, Wang K and Yao X:
MicroRNA-195 is associated with regulating the pathophysiologic
process of human laryngeal squamous cell carcinoma. Mol Med Rep.
17:5283–5291. 2018.PubMed/NCBI
|
|
14
|
Fu HL, Wu DP, Wang XF, Wang JG, Jiao F,
Song LL, Xie H, Wen XY, Shan HS, Du YX and Zhao YP: Altered miRNA
expression is associated with differentiation, invasion, and
metastasis of esophageal squamous cell carcinoma (ESCC) in patients
from Huaian, China. Cell Biochem Biophys. 67:657–668. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun N, Ye L, Chang T and Li X and Li X:
microRNA-195-Cdc42 axis acts as a prognostic factor of esophageal
squamous cell carcinoma. Int J Clin Exp Pathol. 7:6871–6879.
2014.PubMed/NCBI
|
|
16
|
Yin Y, Hong S, Yu S, Huang Y, Chen S, Liu
Y, Zhang Q, Li Y and Xiao H: MiR-195 inhibits tumor growth and
metastasis in papillary thyroid carcinoma cell lines by targeting
CCND1 and FGF2. Int J Endocrinol. 2017:61804252017. View Article : Google Scholar :
|
|
17
|
Pi J, Liu J, Zhuang T, Zhang L, Sun H,
Chen X, Zhao Q, Kuang Y, Peng S, Zhou X, et al: Elevated Expression
of miR302-367 in endothelial cells inhibits developmental
angiogenesis via CDC42/CCND1 mediated signaling pathways.
Theranostics. 8:1511–1526. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
19
|
Dong N, Xu B, Benya SR and Tang X:
MiRNA-26b inhibits the proliferation, migration, and
epithelial-mesenchymal transition of lens epithelial cells. Mol
Cell Biochem. 396:229–238. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dong M, Xie Y and Xu Y: miR-7-5p regulates
the proliferation and migration of colorectal cancer cells by
negatively regulating the expression of Krüppel-like factor 4.
Oncol Lett. 17:3241–3246. 2019.PubMed/NCBI
|
|
21
|
Fan X, Liu M, Tang H, Leng D, Hu S, Lu R,
Wan W and Yuan S: MicroRNA-7 exerts antiangiogenic effect on
colorectal cancer via ERK signaling. J Surg Res. 240:48–59. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cui J, Li W, Liu G, Chen X, Gao X, Lu H
and Lin D: A novel circular RNA, hsa_circ_0043278, acts as a
potential biomarker and promotes non-small cell lung cancer cell
proliferation and migration by regulating miR-520f. Artif Cells
Nanomed Biotechnol. 47:810–821. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Galoian KA, Guettouche T, Issac B, Qureshi
A and Temple HT: Regulation of onco and tumor suppressor MiRNAs by
mTORC1 inhibitor PRP-1 in human chondrosarcoma. Tumour Biol.
35:2335–2341. 2014. View Article : Google Scholar
|
|
24
|
Wang H: Predicting cancer-related MiRNAs
using expression profiles in tumor tissue. Curr Pharm Biotechnol.
15:438–444. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Thomas J, Ohtsuka M, Pichler M and Ling H:
MicroRNAs: Clinical relevance in colorectal cancer. Int J Mol Sci.
16:28063–28076. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xie X, Xu X, Sun C and Yu Z: Long
intergenic noncoding RNA SNHG16 interacts with miR-195 to promote
proliferation, invasion and tumorigenesis in hepatocellular
carcinoma. Exp Cell Res. 383:1115012019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen LP, Zhang NN, Ren XQ, He J and Li Y:
miR-103/miR-195/miR-15b regulate SALL4 and inhibit proliferation
and migration in Glioma. Molecules. 23:29382018. View Article : Google Scholar :
|
|
28
|
Li B and Wang S and Wang S: MiR-195
suppresses colon cancer proliferation and metastasis by targeting
WNT3A. Mol Genet Genomics. 293:1245–1253. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Maroof H, Irani S, Arianna A, Vider J,
Gopalan V and Lam AK: Interactions of vascular endothelial growth
factor and p53 with miR-195 in thyroid carcinoma: Possible
therapeutic targets in aggressive thyroid cancers. Curr Cancer Drug
Targets. 19:561–570. 2019. View Article : Google Scholar
|
|
30
|
Humphreys KJ, McKinnon RA and Michael MZ:
miR-18a inhibits CDC42 and plays a tumour suppressor role in
colorectal cancer cells. PLoS One. 9:e1122882014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Satterfield L, Shuck R, Kurenbekova L,
Allen-Rhoades W, Edwards D, Huang S, Rajapakshe K, Coarfa C,
Donehower LA and Yustein JT: miR-130b directly targets ARHGAP1 to
drive activation of a metastatic CDC42-PAK1-AP1 positive feedback
loop in Ewing sarcoma. Int J Cancer. 141:2062–2075. 2017.
View Article : Google Scholar : PubMed/NCBI
|