Introduction
Osteoarthritis is the most common degenerative joint
disease, causing pain and dysfunction in numerous joints (1). Due to its complex pathogenesis,
there is currently no effective treatment for osteoarthritis (OA)
(1). Therefore, a better
understanding of the pathogenesis of OA would be of great
significance to aid in its early prevention and treatment.
The subchondral bone is located below the articular
cartilage, which plays a significant role in disease pathogenesis
(2). Changes in the
microstructure of the subchondral bone can be observed earlier than
damage to the articular cartilage (3,4).
Under physiological conditions, bone remodeling maintains bone
tissue integrity through the coupling of osteoclast-mediated
resorption and osteoblast-mediated formation of bone (5). Abnormal bone remodeling causes
microstructural destruction of the subchondral bone, altering its
mechanical properties (6).
Additionally, articular cartilage does not withstand shear stress
during joint movement, degenerating as a result (2,7).
Increased subchondral bone remodeling is characterized by increased
bone loss, decreased bone density, and deterioration of its
microstructure, which leads to articular cartilage degeneration
(8). Bone resorption due to
osteoclasts has been demonstrated to significantly increase in
anterior cruciate ligament transection (ACLT) and ovariectomized
(OVX) models of OA. A reduction in articular cartilage degeneration
can be observed when osteoclast-mediated bone resorption is
inhibited using alendronate (9,10).
In OVX mice, estrogen deficiency causes osteoclast-mediated bone
resorption, increased subchondral bone remodeling, and
deterioration of the subchondral bone microstructure, resulting in
degeneration of the articular cartilage, which establishes an
osteoporosis-associated OA mouse model induced by ovariectomy
(9,11,12). These studies suggest that abnormal
subchondral bone remodeling destroys subchondral bone
microstructure, inducing degeneration of the articular
cartilage.
Sclerostin, a potent inhibitor of the Wnt/β-catenin
pathway, is secreted by osteocytes and inhibits osteoblast-mediated
bone formation (13,14). As OA develops, concentrations of
sclerostin in the subchondral bone gradually decrease, activating
the Wnt/β-catenian pathway that promotes bone formation and
accelerates cartilage degradation (15). Previous studies have suggested
that mechanical stress leads to a decrease in sclerostin in
subchondral bone, however, the mechanism by which sclerostin levels
decline in the subchondral bone remains entirely unclear (16-18). LIF is secreted by osteoclasts and
acts between bone resorption and formation as a regulator of bone
remodelling (19). Studies on
osteoporosis have demonstrated that osteoclasts oversecrete LIF,
inhibiting the expression of sclerostin and, thus, promoting
abnormal bone formation and mediating abnormal bone remodelling
(20). Inhibition of
osteoclastogenesis can reduce the secretion of LIF, thereby
reducing abnormal bone remodeling in osteoporosis (20). However, the role of LIF in the
abnormal remodeling of the subchondral bone during OA remains
unclear.
Abnormal angiogenesis of the subchondral bone is an
important pathological feature that leads to abnormal bone
remodeling in OA (21). In a
mouse model of OA established by ACLT and OVX, the vascular
endothelial progenitor cell marker CD31 was significantly increased
in the subchondral bone (12,22-24) and the expression of the catabolic
factors, MMP-13 and VEGF were also significantly increased,
resulting in the articular cartilage being severely damaged.
Inhibition of the abnormal angiogenesis of the subchondral bone
alleviates articular cartilage degeneration (22-24).
Artemisinin, a well-known antimalarial drug, is a
sesquiterpene lactone isolated from Artemisia (25). Dihydroartemisinin (DHA) is a
semisynthetic derivative of artemisinin that has fewer side effects
(26). DHA reduces bone loss by
inhibiting receptor activator of nuclear factor κB ligand
(RANKL)-induced osteoclastogenesis in OVX mice and in a wear
particle-induced mouse osteolysis model (27,28). In addition, DHA has been revealed
to have an antiangiogenic effect (27,28). However, the role of DHA in the
abnormal bone remodeling and angiogenesis of OA remains unclear.
The aim of this study was to investigate the potential role of DHA
in alleviating abnormal subchondral bone remodeling and
angiogenesis in order to reduce articular cartilage degeneration in
a mouse model of OA.
Materials and methods
Ethics statement
All procedures and protocols were approved by the
Scientific Research Ethics Committee of the General Hospital of
Ningxia Medical University (protocol no. 2016-147). All experiments
were performed in accordance with the principles and guidelines of
the National Institutes of Health Guide for Care and Use of
Laboratory Animals.
Animals
A total of 120 female C57BL/6j mice (weight, 19-20
g), ten weeks old, were used in the present study. All mice were
housed at a constant temperature (25°C), at 55% humidity on a 12-h
light/dark cycle with free access to food and water. The mice were
anesthetized with pentobarbital sodium salt (60 mg/kg) by
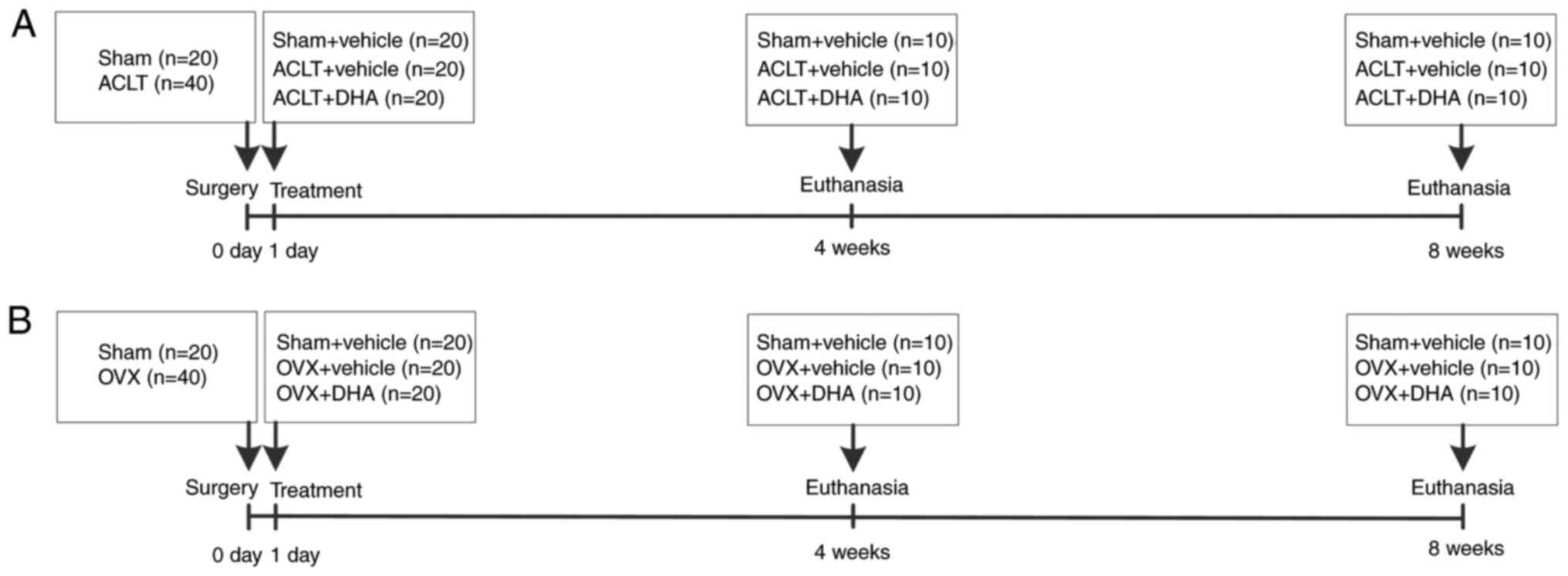
intraperitoneal injection. The schematic of the experimental
protocol is listed in Fig. 1.
Sixty mice were randomly divided into 3 groups: A
sham-operated group (n=20), ACLT+vehicle group (n=20), and ACLT+DHA
group (n=20). Surgery was performed on the right knees of each
mouse as previously described (29). In the two ACLT groups, the right
knee joint capsule was exposed using a medial parapatellar approach
and then the anterior cruciate ligament was transected with
micro-scissors, prior to closing the joint capsule and skin. The
same procedure was performed in the sham-operated group with the
exception of the transection of the ACL. The ACLT+DHA group was
treated with DHA (product no. D7439; Sigma-Aldrich; Merck KGaA) by
intraperitoneal injection (1 mg/kg/2 days) from the first
post-operative day until sacrifice (Fig. 1A). Flow displacement rate of
CO2 used for euthanasia was 30%. Mouse euthanasia was
confirmed, including lack of pulse, breathing, corneal reflex, and
response to firm toe pinch. DHA was prepared according to the
literature (30), and the method
was as follows: 25 mg of DHA was weighed in a precision electronic
balance, and dissolved in 1 ml of prepared DMSO, and then diluted
in 99 ml sterile PBS solution to prepare 1 mg/kg DHA. The sham and
vehicle treatment groups were administered solvent as a vehicle,
using the same dose, frequency and duration as the DHA group.
Sixty mice were randomly divided into 3 groups: A
sham-operated group (n=20), OVX+vehicle group (n=20), and OVX+DHA
group (n=20). According to the literature, the mice were operated
from the abdominal cavity through the back to remove the bilateral
ovaries (12,31). The same procedure was performed in
the sham-operated group, except that the bilateral ovaries were
only removed. The OVX+DHA group was treated with DHA from the first
day after operation (1 mg/kg/2 days) until sacrifice (Fig. 1B). Ovietectomy was performed under
anaesthesia. The mice were anesthetized with pentobarbital sodium
salt (60 mg/kg) by intraperitoneal injection. The method of
euthanasia and the configuration of DHA were the same as those of
the ACLT mice. The sham operation and carrier groups used the same
dose, frequency, and duration as the DHA group. Ten mice were
separately euthanized at 4 or 8 weeks after surgery in each group
by carbon dioxide inhalation, followed immediate harvesting of the
right knees for subsequent analysis.
Histology
Harvested knees were fixed in 4% paraformaldehyde
for 24 h at 4°C then decalcified in 10% EDTA (pH 7.4) for 3 weeks
prior to embedding in paraffin. The medial compartment of the knee
joints were cut into 4-µm-thick sections along the sagittal
plane and stained with safranin O-fast green at 25°C in the
laboratory. Deparaffinization of the slides was performed in xylene
two times for 10 min, followed by hydration in 100% alcohol twice
for 5 min, 95% alcohol for 2 min and 80% alcohol for 2 min.
Hematoxylin was added to slides for 3 min prior to hydrating the
slides gently in running water for 10 min. Slides were then stained
with 0.2% Fast Green (product no. F7252; Sigma-Aldrich; Merck KGaA)
for 3 min, and then subjected to 1% acetic acid for 5 sec, 0.1%
Safranin O (product no. S8884; Sigma-Aldrich; Merck KGaA) for 3
min. Slides were hydrated in 95% alcohol for 5 sec, 100% alcohol
twice for 15 sec, followed by 2 changes in xylene prior to
cover-slipping the slides. Regions from three slides per mouse were
imaged by light microscopy at a magnification of ×100 and three
fields per region were randomly selected per slide. The
Osteoarthritis Research Society International (OARSI)-modified
Mankin score was used to perform a histopathological grade
assessment of the cartilage.
Immunohistochemistry and
immunofluorescence
Standard immunostaining was conducted in the present
study. Sagittal sections that had been paraffin-embedded were
incubated overnight at 4°C with primary antibodies against matrix
metalloproteinase-13 (MMP-13) (1:100; product code ab39012; Abcam),
vascular endothelial growth factor A (VEGFA) (1:100; product code
ab52917; Abcam), leukemia inhibitory factor (LIF) (1:100; cat. no.
AB-449-NA; R&D Systems, Inc.), sclerostin (1:50; product code
ab63097; Abcam), β-catenin (1:100; cat. no. 51067-2-AP; ProteinTech
Group, Inc.), RANKL (1:100; product code ab216484; Abcam) and CD31
(1:25; product code ab28364; Abcam). For immunohistochemical
staining, sections were processed using a two-step IHC detection
reagent (ZSGB-Bio; OriGene Technologies, Inc.). Briefly, sections
were incubated with reaction enhancement solution (reagent 1) for
20 min at 37°C and then with enhanced enzyme-labeled goat
anti-rabbit IgG polymer (reagent 2) for 40 min at 37°C. The
sections were then developed using 3,3′-diaminobenzidine (DAB)
(ZSGB-Bio; OriGene Technologies, Inc.), followed by counterstaining
with hematoxylin (ZSGB-Bio; OriGene Technologies, Inc.). For
immunofluorescence staining, sections were incubated with Alexa
Fluor® 488 goat anti-rabbit secondary antibody (1:500;
product code ab150077; Abcam) for 1 h at 37°C in the dark. Regions
from three slides per mouse were imaged by fluorescence microscopy
at a magnification of ×400 and three fields per region were
randomly selected per slide. Image-Pro Plus 6.0 (Media Cybernetics,
Inc.) was used to count chondrocytes within the entire tibial
articular cartilage and all cells within the entire tibial
subchondral bone which exhibited positive staining for each
antibody.
Micro-computed tomography (CT)
analysis
Harvested knee joints were dissected free of soft
tissue, fixed overnight in 70% ethanol at 4°C and analyzed by
micro-CT (SkyScan 1176; Bruker micro-CT) at a resolution of 9
µm/pixel. The micro-CT scans were acquired over an exposure
time of 900 ms, a voltage of 50 kV and a current of 500 µA.
Images were reconstructed by NRecon version 1.1.11 (Bruker
micro-CT) and analyzed by CTAn, v1.15 (SkyScan1176 in vivo
micro-CT; Bruker). A sagittal view of the entire medial compartment
of the tibial subchondral bone was used for 3D histomorphometric
analysis, with bone volume/tissue volume (BV/TV, %), trabecular
separation (Tb.Sp), and BMD extracted for performing
comparisons.
Statistical analysis
Data were analyzed using one-way and two-way
factorial design analyses of the variations followed by
Student-Newman-Keuls and Bonferroni post hoc tests. Data are
presented as the means ± SD. For OARSI scores, data were analyzed
using Kruskal-Wallis test followed by Dunn's test. P<0.05 was
considered to indicate a statistically significant difference.
GraphPad Prism 5 software (GraphPad Software, Inc.) was used for
statistical analysis.
Results
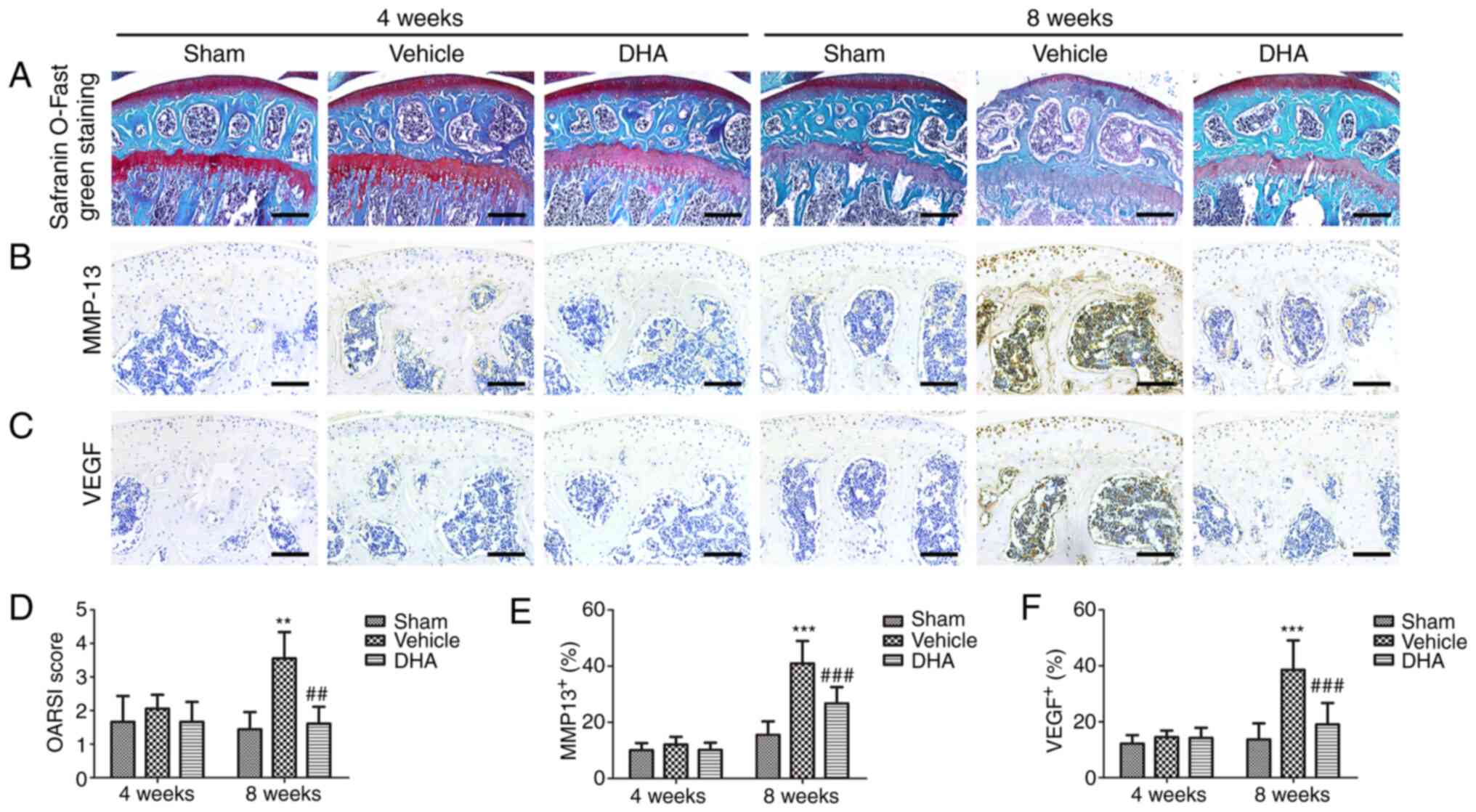
DHA preserves articular cartilage in ACLT
mice
Safranin O-fast green staining indicated that there
was significant loss of proteoglycans, with OARSI scores increasing
significantly in the ACLT+vehicle group relative to the
sham-operated group at 4- and 8-weeks following surgery (Fig. 2A and D). Similarly, results of
immunohistochemical staining demonstrated that the expression of
MMP-13 (Fig. 2B and E) and VEGF
(Fig. 2C and F) was significantly
increased after 4 and 8 weeks. Administration of DHA resulted in
retention of proteoglycans and OARSI scores that were improved in
the ACLT+DHA group compared with the ACLT+vehicle group after 4 and
8 weeks (Fig. 2A and D).
Aberrantly expressed MMP-13 (Fig. 2B
and E) and VEGF (Fig. 2C and
F) were recovered in the ACLT+DHA group compared with the
ACLT+vehicle group at both 4- and 8-weeks following surgery.
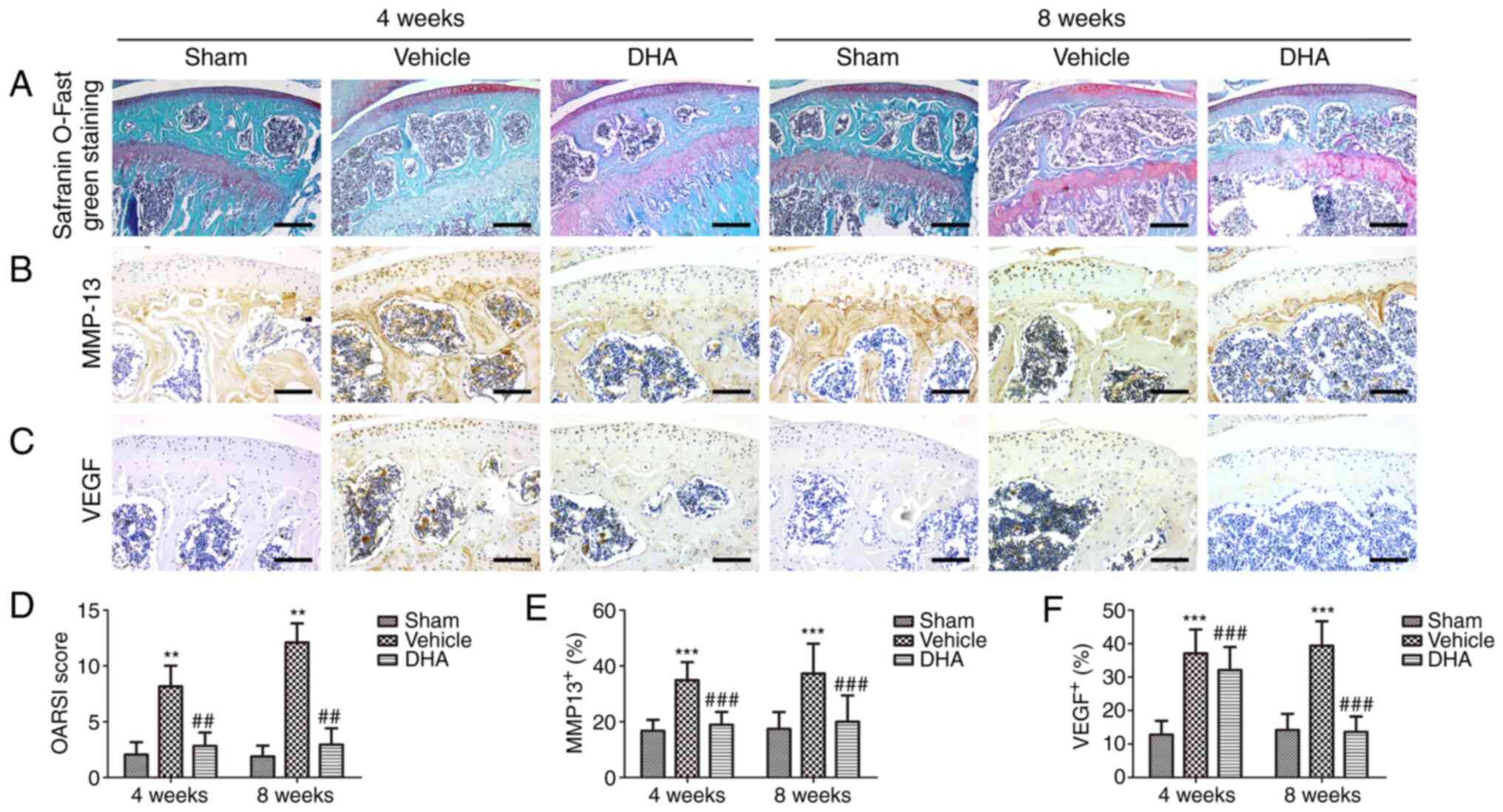 | Figure 2DHA preserves the articular cartilage
in ACLT mice. (A) Histological analysis of articular cartilage
using safranin O-fast green staining of sagittal sections of the
medial compartment of the tibia. Scale bar, 200 µm. (B, C, E
and F) Measurement of matrix (B and E) MMP-13 and (C and F) VEGF
expression by immunohistochemical staining and quantitative
analysis. Scale bar, 100 µm. (D) OARSI-modified Mankin score
of articular cartilage at various time-points. Sham, sham-operated
group; Vehicle, ACLT+vehicle group; DHA, ACLT+DHA group. n=10 per
group. **P<0.01 and ***P<0.001,
compared with the the sham-operated group. ##P<0.01
and ###P<0.001, compared with the ACLT + vehicle
group. DHA, dihydroartemisinin; ACLT, anterior cruciate ligament
transection; MMP-13, metalloproteinase-13; VEGF, vascular
endothelial growth factor; OARSI, Osteoarthritis Research Society
International. |
DHA restores the microarchitecture of the
subchondral bone in ACLT mice
Abnormal bone remodeling destroys the microstructure
of subchondral bone which results in articular cartilage
degeneration (21). It was
investigated whether DHA was able to protect articular cartilage by
preserving the microstructure of subchondral bone. The tibial
subchondral bone structure of the tibia was analyzed by micro-CT.
The results demonstrated that DHA improved the subchondral bone
microstructure (Fig. 3A). BV/TV
(Fig. 3B) and BMD (Fig. 3D) were significantly decreased in
the ACLT+vehicle group compared with the sham-operated group at
both 4- and 8-weeks but DHA abrogated these changes (ACLT+DHA
group) relative to the ACLT+vehicle group 4-weeks after surgery
(Fig. 3B and D). The Tb.Sp was
significantly increased in the ACLT+vehicle group relative to the
sham-operated group at both 4- and 8-weeks (Fig. 3C). DHA recovered these changes
(ACLT+DHA group) compared with the ACLT+vehicle group 4-weeks after
surgery (Fig. 3C). Collectively,
these data indicated that systemic administration of DHA inhibited
abnormal bone remodeling and restored the subchondral bone
microstructure.
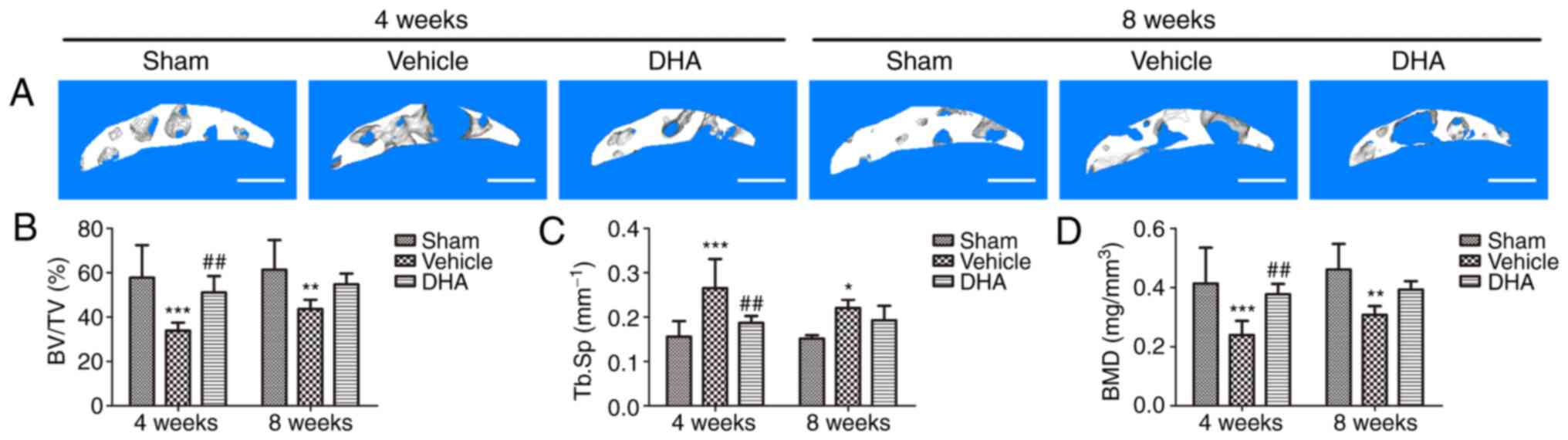 | Figure 3DHA restores the microarchitecture of
the subchondral bone of ACLT mice. (A) 3D micro-CT reconstruction
of sagittal views of the medial compartment of tibial subchondral
bone at different time-points after sham or ACLT surgery. Scale
bar, 500 µm. (B and D) Quantitative micro-CT analyses of the
microarchitecture of tibial subchondral bone: (B) BV/TV (%), (C)
Tb.Sp and (D) BMD. Sham, sham-operated group; Vehicle, ACLT+vehicle
group; DHA, ACLT+DHA group. n=6 per group. *P<0.05,
**P<0.01 and ***P<0.001, compared with
the sham-operated group. ##P<0.01, compared with the
vehicle group. DHA, dihydroartemisinin; ACLT, anterior cruciate
ligament transection; micro-CT, micro-computed tomography; BV/TV,
bone volume/tissue volume; Tb.Sp, trabecular separation; BMD, bone
mineral density. |
DHA inhibits abnormal bone remodeling in
the subchondral bone in ACLT mice
Immunohistochemistry and immunofluorescence were
performed to investigate whether systemic DHA was able to inhibit
abnormal bone remodeling and angiogenesis in subchondral bone
4-weeks after surgery. RANKL-immunostained sections demonstrated a
significant increase in the number of cells positive for RANKL in
the ACLT+vehicle group relative to the sham-operated group
(Fig. 4A and E). Administration
of DHA significantly reduced this increased number relative to the
ACLT+vehicle group (Fig. 4A and
E). To investigate whether osteoclasts altered the expression
of sclerostin in osteocytes, sclerostin expression was measured by
immunohistochemistry. The results demonstrated a significant
decrease in sclerostin expression in the ACLT+vehicle group
relative to the sham-operated group (Fig. 4B and F). Treatment with DHA
significantly increased sclerostin expression relative to the
ACLT+vehicle group (Fig. 4B and
F). The expression of β-catenin in osteoblasts was assessed by
immunohistochemistry and a significant increase was observed in the
ACLT+vehicle group compared with the sham-operated group (Fig. 4C and G). Administration of DHA
significantly reduced β-catenin expression compared with the
ACLT+vehicle group (Fig. 4C and
G).
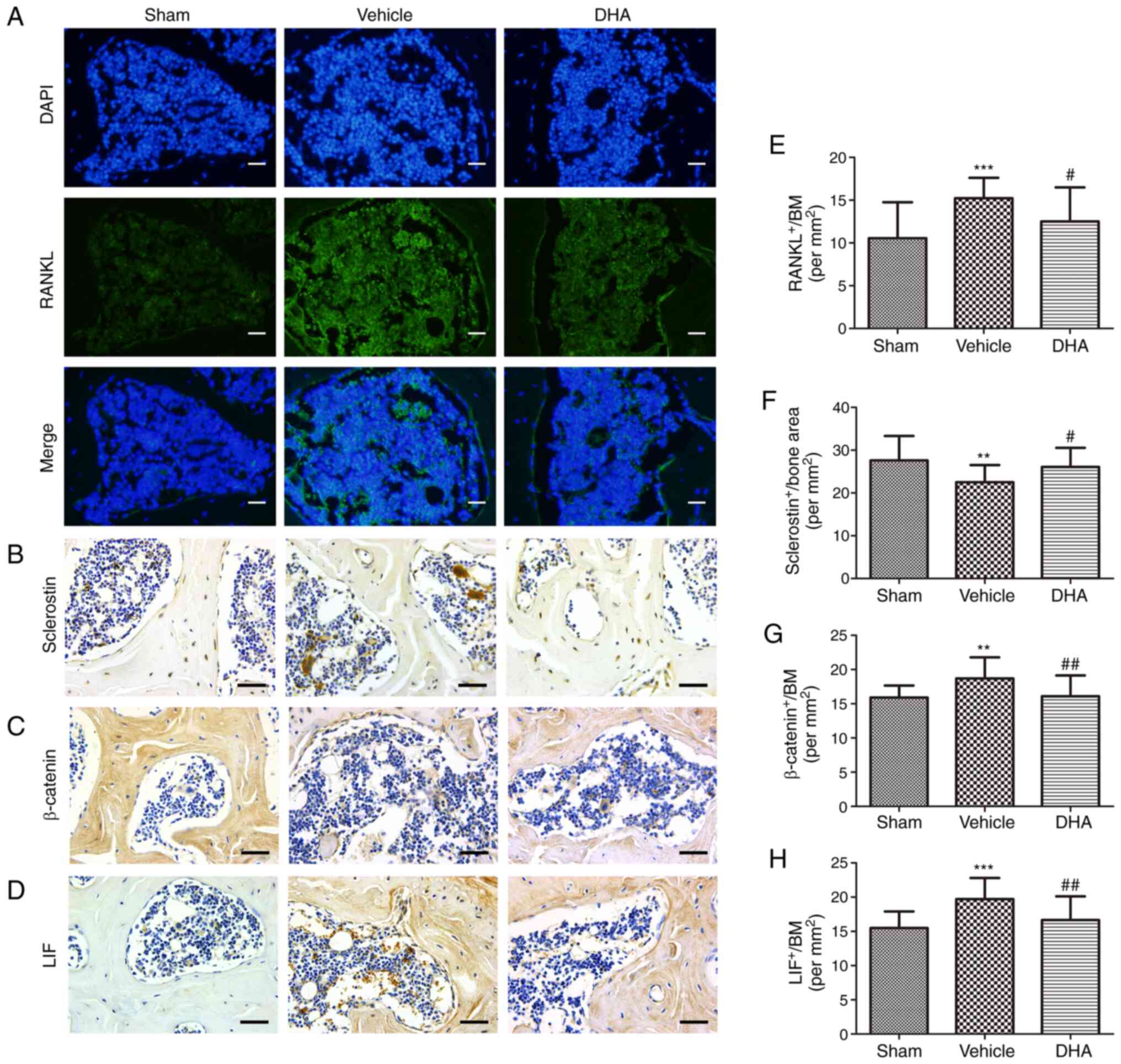 | Figure 4DHA inhibits abnormal bone remodeling
in the subchondral bone of ACLT mice. (A and E) Immunofluorescence
staining and quantification of the expression of RANKL (green) in
tibial subchondral bone. Cell nuclei were stained blue using DAPI.
Scale bar, 50 µm. (B-D and F-H) Immunohistochemical staining
and quantification of the expression of (B and F) sclerostin, (C
and G) β-catenin and (D and H) LIF in tibial subchondral bone.
Scale bar, 50 µm. Sham, sham-operated group; Vehicle,
ACLT+vehicle group; DHA, ACLT+DHA group. n=10 per group.
**P<0.01 and ***P<0.001, compared with
the sham-operated group. #P<0.05 and
##P<0.01, compared with the vehicle group. DHA,
dihydroartemisinin; ACLT, anterior cruciate ligament transection;
RANKL, receptor activator of nuclear factor κB ligand; LIF,
leukemia inhibitory factor. |
It was also investigated whether DHA inhibited the
excessive expression of LIF, which is secreted by osteoclasts in
the subchondral bone, using immunohistochemistry 4-weeks after
surgery. The results demonstrated that a significant increase in
LIF expression was observed in the ACLT+vehicle group compared with
the sham-operated group (Fig. 4D and
H). DHA significantly reduced this expression relative to the
ACLT+vehicle group (Fig. 4D and
H).
These results indicated that DHA inhibited the
excessive expression of LIF secreted by osteoclasts in ACLT mice,
thus reducing inhibition of sclerostin and preventing subchondral
bone remodeling, thereby restoring the subchondral bone
microstructure and inhibiting articular cartilage degeneration.
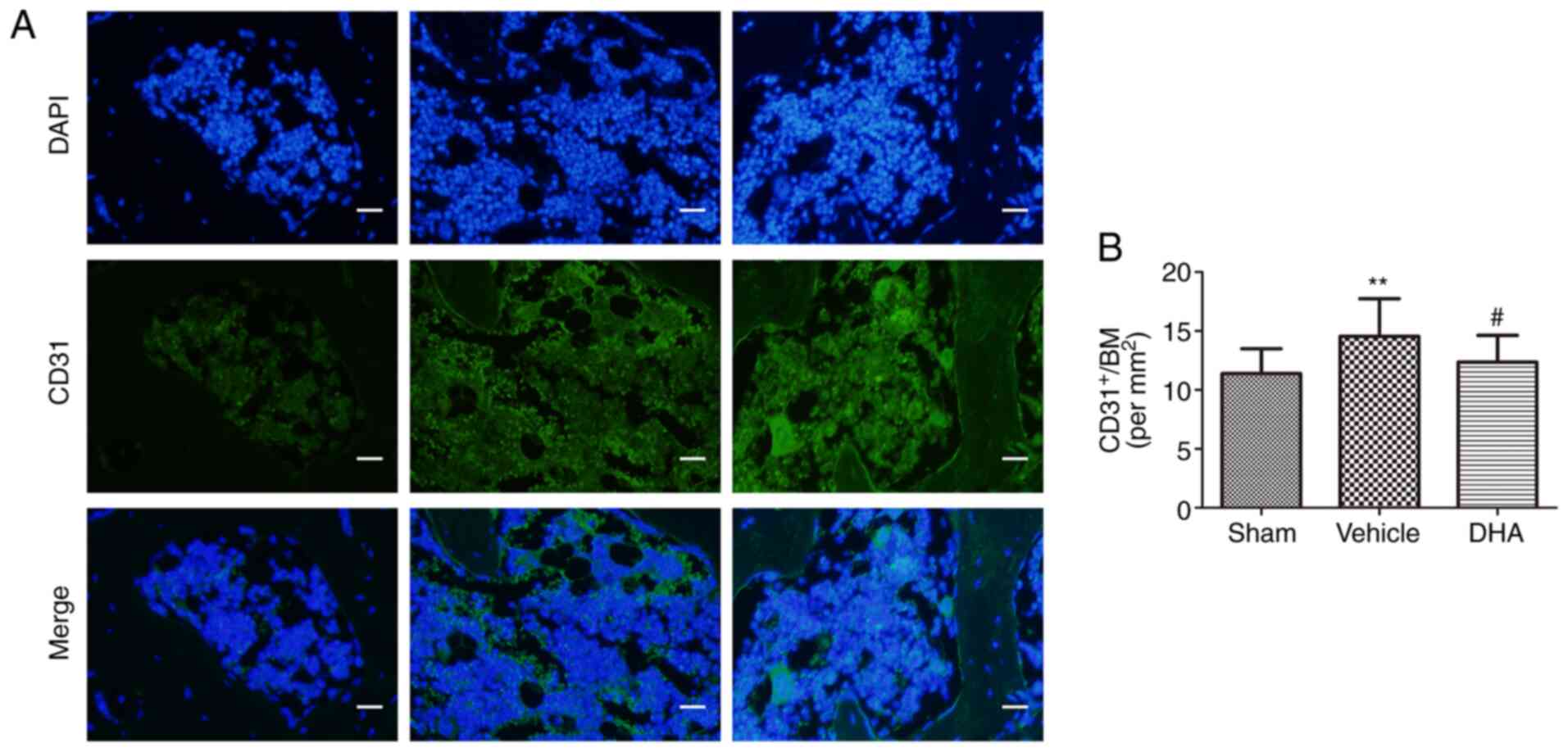
DHA inhibits abnormal angiogenesis in the
subchondral bone in ACLT mice
Abnormal angiogenesis in the subchondral bone is a
pathological feature of OA (32).
An angiogenic marker, CD31-positive endothelial progenitor cells
(33), was assessed using
immunofluorescence. The results indicated that its number
significantly increased in the ACLT+vehicle group relative to the
sham-operated group (Fig. 5) and
that the number was reduced in the ACLT+DHA group compared with the
ACLT+vehicle group (Fig. 5).
DHA preserves the articular cartilage in
OVX mice
Safranin O-fast green staining indicated that there
was significant loss of proteoglycans, with OARSI scores
significantly increasing in the OVX+vehicle group relative to the
sham-operated group at 8-weeks following surgery (Fig. 6A and D), but not 4-weeks.
Similarly, the results of the immunohistochemical staining
demonstrated that the expression of MMP-13 (Fig. 6B and E) and VEGF (Fig. 6C and F) significantly increased
after 8-weeks, but not 4-weeks. Administration of DHA resulted in
retention of proteoglycans and OARSI scores that were improved in
the OVX+DHA group compared with the OVX+vehicle group after 8-weeks
(Fig. 6A and D). Aberrantly
expressed MMP-13 (Fig. 6B and E)
and VEGF (Fig. 6C and F) were
recovered in the OVX+DHA group compared with the OVX+vehicle group
at 8-weeks following surgery.
 | Figure 6DHA preserves articular cartilage in
OVX mice. (A) Histological analysis of articular cartilage using
safranin O-fast green staining of sagittal sections of the medial
compartment of the tibia. Scale bar, 200 µm. (B, C, E and F)
Measurement of matrix (B and E) MMP-13 and (C and F) VEGF
expression by immunohistochemical staining and quantitative
analysis. Scale bar, 100 µm. (D) OARSI-modified Mankin score
of articular cartilage at various time-points. Sham, sham-operated
group; Vehicle, OVX+vehicle group; DHA, OVX+DHA group. n=10 per
group. **P<0.01 and ***P<0.001,
compared with the sham-operated group. ##P<0.01 and
###P<0.001, compared with the OVX+vehicle group. DHA,
dihydroartemisinin; OVX, ovariectomized; MMP-13,
metalloproteinase-13; VEGF, vascular endothelial growth factor;
OARSI, Osteoarthritis Research Society International. |
DHA restores the microarchitecture of the
subchondral bone in OVX mice
Abnormal bone remodeling destroys the microstructure
of the subchondral bone which results in articular cartilage
degeneration (21). It was
investigated whether DHA was able to protect the articular
cartilage by preserving the microstructure of the subchondral bone.
The tibial subchondral bone structure of the tibia was analyzed by
micro-CT. The mouse OA model established by OVX, was established
8-weeks after surgery, but not 4-weeks. The OVX mice were also
euthanized after 4 weeks but model establishment was not successful
at that time-point. Therefore, the micro-CT result only contains
8-week results. The results demonstrated that DHA improved the
subchondral bone microstructure (Fig.
7A). BV/TV (Fig. 7B) and BMD
(Fig. 7D) significantly decreased
in the OVX+vehicle group compared with the sham-operated group at
8-weeks, but DHA abrogated these changes (OVX+DHA group) relative
to the OVX+vehicle group 8-weeks after surgery (Fig. 7B and D). The Tb.Sp significantly
increased in the OVX+vehicle group relative to the sham-operated
group at 8-weeks (Fig. 7C). DHA
recovered these changes (OVX+DHA group) compared with the
OVX+vehicle group 8-weeks after surgery (Fig. 7C). Collectively, these data
indicated that systemic administration of DHA inhibited abnormal
bone remodeling and restored subchondral bone microstructure.
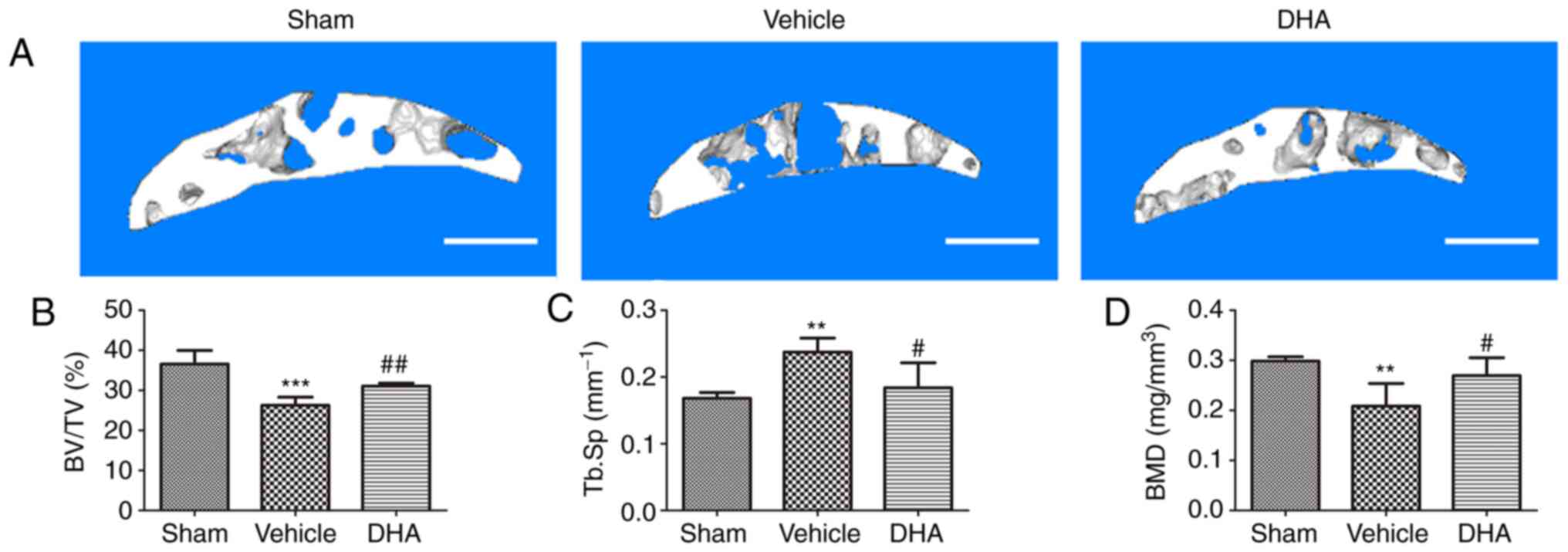 | Figure 7DHA restores the microarchitecture of
the subchondral bone in OVX mice. (A) 3D micro-CT reconstruction of
sagittal views of the medial compartment of tibial subchondral bone
at different time-points after sham or OVX surgery. Scale bar, 500
µm. (B-D) Quantitative micro-CT analyses of the
microarchitecture of tibial subchondral bone: (B) BV/TV (%), (C)
Tb.Sp and (D) BMD. Sham, sham-operated group; Vehicle, OVX+vehicle
group; DHA, OVX+DHA group. n=6 per group. **P<0.01
and ***P<0.001, compared with the sham-operated
group. #P<0.05 and ##P<0.01, compared
with the vehicle group. DHA, dihydroartemisinin; OVX,
ovariectomized; micro-CT, micro-computed tomography; BV/TV, bone
volume/tissue volume; Tb.Sp, trabecular separation; BMD, bone
mineral density. |
DHA inhibits abnormal bone remodeling in
subchondral bone in OVX mice
Immunohistochemistry and immunofluorescence were
performed to investigate whether systemic DHA was able to inhibit
abnormal bone remodeling and angiogenesis in the subchondral bone
8-weeks after surgery. RANKL-immunostained sections demonstrated a
significant increase in the number of cells positive for RANKL in
the OVX+vehicle group relative to the sham-operated group (Fig. 8A and E). Administration of DHA
significantly reduced that increased number relative to the
OVX+vehicle group (Fig. 8A and
E). To investigate whether osteoclasts altered the expression
of sclerostin in osteocytes, sclerostin expression was assessed by
immunohistochemistry. The results demonstrated a significant
decrease in sclerostin expression in the OVX+vehicle group relative
to the sham-operated group (Fig. 8B
and F). Treatment with DHA significantly increased sclerostin
expression relative to the OVX+vehicle group (Fig. 8B and F). The expression of
β-catenin in osteoblasts was assessed by immunohistochemistry and a
significant increase was observed in the OVX+vehicle group compared
with the sham-operated group (Fig. 8C
and G). Administration of DHA significantly reduced β-catenin
expression compared with the OVX+vehicle group (Fig. 8C and G).
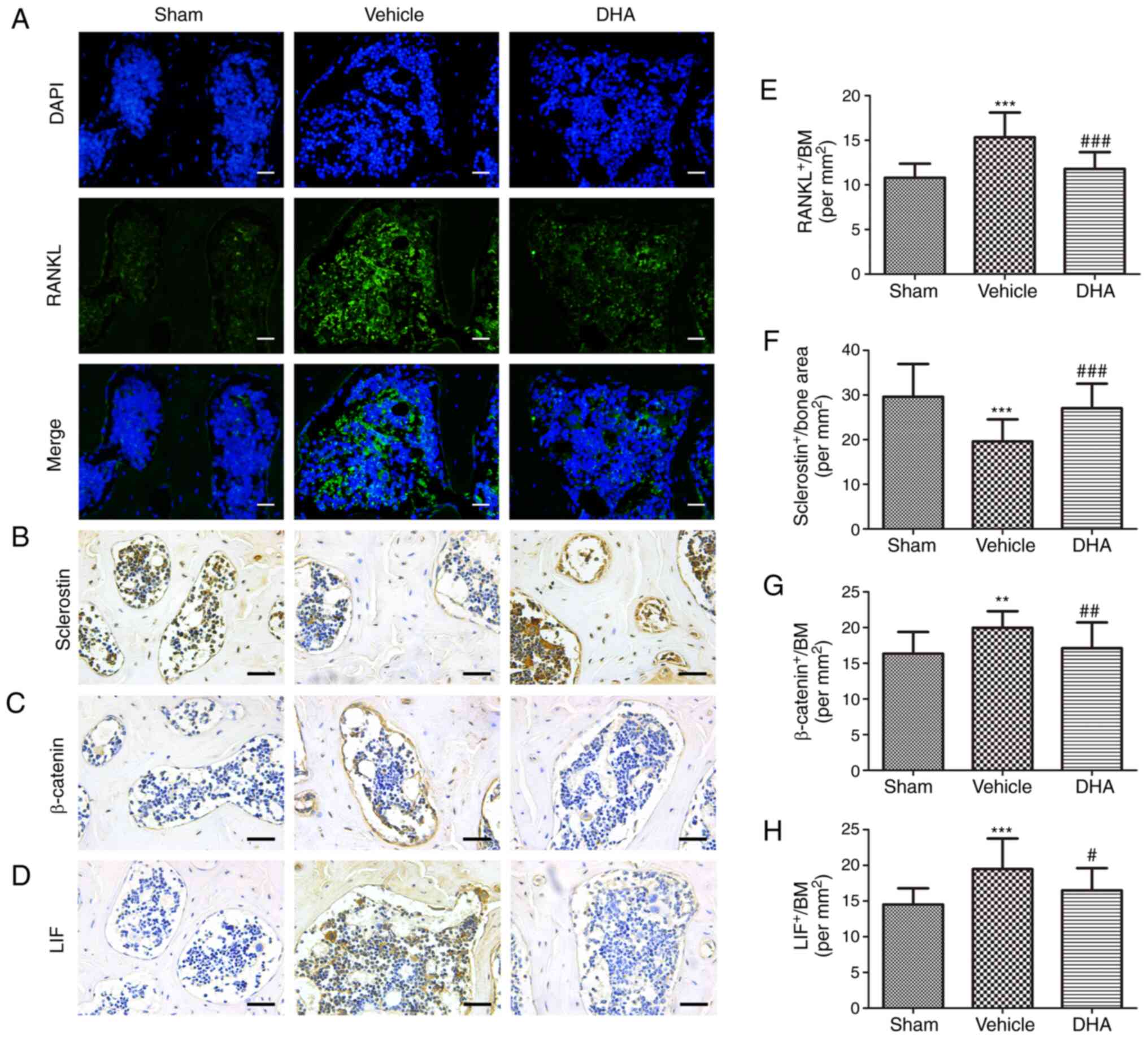 | Figure 8DHA inhibits abnormal bone remodeling
in the subchondral bone of OVX mice. (A and E) Immunofluorescence
staining and quantification of the expression of RANKL (green) in
tibial subchondral bone. Cell nuclei were stained blue using DAPI.
Scale bar, 50 µm. (B-D and F-H) Immunohistochemical staining
and quantification of the expression of (B and F) sclerostin, (C
and G) β-catenin and (D and H) LIF in tibial subchondral bone.
Scale bar, 50 µm. Sham, sham-operated group. Vehicle,
OVX+vehicle group. DHA, OVX+DHA group. n=10 per group.
**P<0.01 and ***P<0.001, compared with
the sham-operated group. #P<0.05 and
##P<0.01 and ###P<0.001 compared with
the vehicle group. DHA, dihydroartemisinin; OVX, ovariectomized;
RANKL, receptor activator of nuclear factor κB ligand; LIF,
leukemia inhibitory factor. |
It was investigated whether DHA inhibited excessive
expression of LIF secreted by osteoclasts in the subchondral bone
using immunohistochemistry 8-weeks after surgery. The results
demonstrated that a significant increase in LIF expression was
observed in the OVX+vehicle group compared with sham-operated group
(Fig. 8D and H). DHA
significantly reduced this expression relative to the OVX+vehicle
group (Fig. 8D and H).
The results indicated that DHA inhibited excessive
expression of LIF secreted by osteoclasts in OVX mice, thus
reducing inhibition of sclerostin and preventing subchondral bone
remodeling, thereby restoring the subchondral bone microstructure
and inhibiting articular cartilage degeneration.
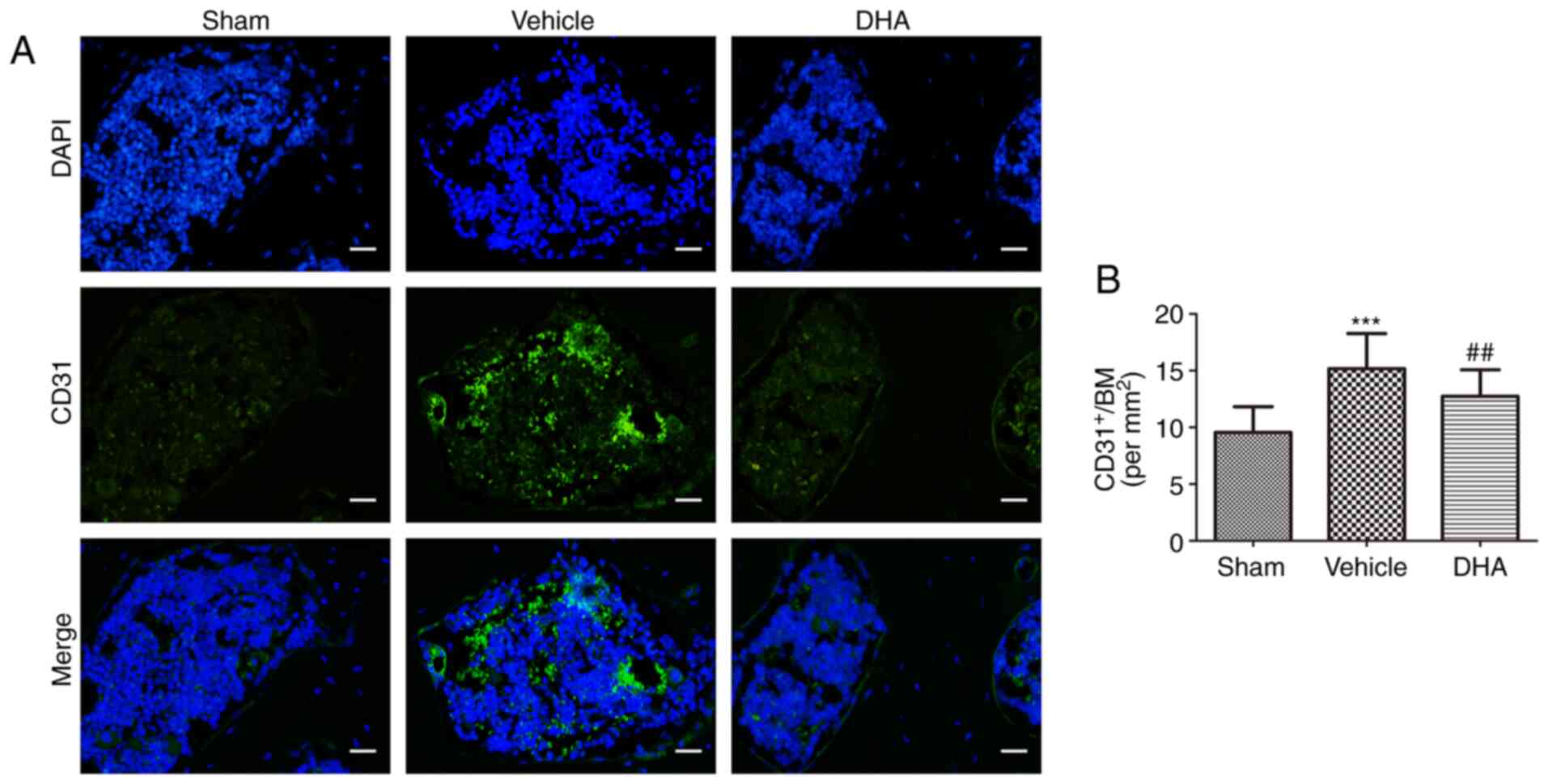
DHA inhibits abnormal angiogenesis in
subchondral bone in OVX mice
Abnormal angiogenesis in subchondral bone is a
pathological feature of OA (32).
The number of CD31-positive endothelial progenitor cells were
assessed using immunofluorescence. The results indicated that
compared with the sham-operated group, their numbers significantly
increased in the OVX+vehicle group (Fig. 9) and that compared with the
OVX+vehicle group, their numbers were decreased in the OVX+DHA
group (Fig. 9).
Discussion
OA is the most common degenerative joint disease and
possesses a complex pathogenesis. Abnormal bone remodeling and
angiogenesis in the subchondral bone destroys the subchondral bone
microstructure and leads to articular cartilage degeneration
(22,23). The present study revealed that DHA
decreased the inhibition of sclerostin by reducing the LIF
secretion of osteoclasts and, hence, attenuated aberrant bone
remodeling and inhibited angiogenesis in subchondral bone, thereby
preserving the subchondral bone microstructure and attenuating
articular cartilage degeneration.
The microstructural integrity of the subchondral
bone is especially important for the protection of articular
cartilage (2). Studies have
revealed that microstructural changes in subchondral bone may
precede the degeneration in articular cartilage in OA (3,4).
The integrity of the bone microstructure is maintained through the
coupling of bone remodeling, which includes the temporal and
spatial balance of bone resorption and formation. This in turn
involves resorption of the bone matrix by osteoclasts, normally
replaced by new bone matrix by osteoblasts (34). Abnormal bone remodeling can
disrupt the homeostatic balance of the bone and destroy the
integrity of the bone microstructure (35). At present, the mechanisms that
balance bone resorption and formation resulting in bone remodeling
remain unclear.
The most commonly used surgical method of inducing
OA is ACLT. In this method, the ACL is transected with
micro-scissors, which causes joint destabilization. The anterior
drawer test with the joint flexed is used to confirm that
transection of the ligament has occurred. The ACLT leads to the
increase of joint mechanical stress. The increased load on the
posterior tibial plateau leads to the destruction of cartilage and
subchondral bone, which leads to OA (29,36). Osteoclasts are the sole cells that
resorb bone matrix, through dissolution by the secretion of acids
and proteases. RANKL is a key factor in osteoclastogenesis
(37). The secretion of RANKL by
osteoblast precursors following abnormal mechanical stimulation
results in its binding to RANK on osteoclast precursors, inducing
osteoclastogenesis that mediates bone resorption (22). The prevalence of OA in
postmenopausal women is higher than that in men. Ovariectomy
significantly reduced the production of estrogen in mice, resulting
in increased bone resorption mediated by subchondral bone
osteoclasts, resulting in increased subchondral bone loss and bone
microstructure deterioration, thereby promoting articular cartilage
degeneration and inducing osteoarthritis (31,38). Osteoporosis (OP) is characterized
by increased bone loss, deterioration of bone microarchitecture and
an increased risk of fragility fractures (39). The pathogenesis of OA has not been
fully elucidated. OA is considered a whole-joint disease in which
all components are involved, with the subchondral bone, which is
located below the articular cartilage, playing a significant role
in disease pathogenesis (40). In
the subchondral bone of OA, osteoclast-mediated bone resorption
increases bone loss, and its microstructure deteriorates, which
leads to articular cartilage degeneration (41). Excessive bone resorption, which is
a hallmark of postmenopausal OP, also occurs at the early stage in
the development of OA (42). They
both have increased bone remodeling, increased osteoclast-mediated
bone resorption, and deterioration of bone microstructure in OVX
mice. The present study demonstrated that the expression of RANKL
significantly increased in the ACLT+vehicle and OVX+vehicle group
compared with the sham-operated group. This indicates that
RANKL-induced osteoclastogenesis is significantly enhanced in ACLT
mice. Consistent with previous studies, the present results
revealed that DHA reduced bone resorption by inhibiting
RANKL-mediated osteoclastogenesis.
The Wnt/β-catenin signaling pathway is considered
the most important pathway that regulates bone homeostasis
(43). Activation of the
Wnt/β-catenin pathway promotes differentiation of bone marrow
mesenchymal stem cells into osteoblasts, promoting osteoblast
proliferation and maturation, inhibiting osteoblast apoptosis, and
thereby promoting osteoblast-mediated bone formation (44). Sclerostin, encoded by the SOST
gene, is secreted by osteocytes and binds to LDL receptor-related
protein (LRP) 5/6 (13,45). It is an antagonist of the
Wnt/β-catenin signalling (13).
Sclerostin inhibits osteoblast-mediated bone formation by binding
to LRP5/6 on osteoblasts (45).
Previous studies have revealed that SOST knockout (KO) mice and
postmenopausal osteoporotic rats treated with sclerostin antibodies
exhibited a significant increase in bone volume, bone formation,
and the number of osteoblasts on the surface of their bones
(46). As OA progresses,
sclerostin expression significantly decreases, causing β-catenin
expression to significantly increase in the subchondral bone
leading to gradually worsening articular cartilage degeneration in
humans (47). The present study
demonstrated that compared with the sham-operated group, sclerostin
expression in osteocytes significantly decreased and β-catenin
expression in osteoblasts significantly increased in the
subchondral bone in the ACLT+vehicle group. This indicated that
decreased expression of sclerostin in the subchondral bone caused
abnormal remodeling that destroyed its microstructural integrity in
ACLT mice. After OVX, OP caused by oestrogen revealed an increase
in bone remodeling, specifically osteoclast-mediated enhancement of
bone resorption, and osteoblast-mediated bone formation was
relatively weakened (9). In
OVX-induced OP-related osteoarthritis, subchondral bone remodeling
increases, characterized by increased bone loss, decreased bone
mass, decreased bone density, and deteriorated bone
microarchitecture (9,12,31). The present study also demonstrated
that compared with the sham-operated group, sclerostin expression
significantly decreased in osteocytes, and β-catenin expression
also significantly increased in osteoblasts in the subchondral bone
in the OVX+vehicle group. This indicated that the decreased
expression of sclerostin in the subchondral bone of OP-induced
mouse OA caused abnormal bone formation and abnormality of the
subchondral bone remodeling, which destroyed the microstructural
integrity of the subchondral bone.
LIF is a bone remodeling regulatory protein that is
secreted by osteoclasts and binds to a receptor complex of
glycoprotein 130 (gp130)/LIF receptor (LIFR) on osteocytes, which
inhibits sclerostin (48,49). Thus, the inhibitory effect of
sclerostin on Wnt/β-catenin in osteoblasts is reduced, promoting
osteoblast-mediated bone formation in RANKL transgenic mice and
OPG-KO mice compared with wild-type mice (20). Treatment of OPG-KO mice with
anti-RANKL antibody, an anti-bone resorption agent, was revealed to
suppress the expression of LIF and increase the expression of
sclerostin, thereby reducing bone formation via inhibition of the
Wnt/β-catenin pathway (20).
Specific knockout of gp130 in osteocytes was revealed to increase
the expression of sclerostin, and, thus, inhibit bone formation in
mice (50). The aforementioned
studies demonstrated that the osteoclast-derived factor LIF
inhibited the expression of sclerostin in osteocytes and, hence,
promoted bone formation. The present study demonstrated that the
expression of LIF in osteoclasts significantly increased in the
ACLT+vehicle group relative to the sham-operated group. We
inhibited osteoclastogenesis by DHA which reduced the expression of
the osteoclast-derived factor LIF, thereby increasing the
expression of sclerostin that inhibits abnormal bone formation and
remodeling of the subchondral bone. Sclerostin is an important
factor in bone remodeling, and is affected by numerous factors
(51,52). Previous studies have found that
mechanical stimulation inhibit the expression of sclerostin
(16-18). Studies on osteoporosis have
revealed that osteoclasts oversecrete LIF and inhibit the
expression of sclerostin (20);
however, in the subchondral bone of OA, its mechanism is unclear.
We first established a model of OA in mice with ACLT mechanical
injury and it was revealed that LIF secreted by osteoclasts
decreased the expression of sclerostin. However, the influence of
the increased mechanical stress in the ACLT model cannot be
completely eliminated. We then used the OVX-induced OP-related
mouse OA model to further investigate the effect of LIF secreted by
osteoclasts on sclerostin. The present study revealed a significant
increase in the expression of LIF in osteoclasts in the OVX+vehicle
group compared with that in the sham-operated group. After
treatment with DHA, the expression of LIF in OVX+DHA group was
significantly lower than that in OVX+vehicle group. This indicated
that DHA inhibited osteoclastogenesis to reduce the expression of
osteoclast-derived factor LIF, and, therefore, the expression of
sclerostin was significantly increased. Compared with the
OVX+vehicle group, DHA significantly decreased Tb.Sp and
significantly increased BV/TV (%) and BMD in the subchondral bone
of the OVX+DHA group. Similarly, the expression of MMP-13 and VEGF
and the OARSI score in the articular cartilage were significantly
decreased. This indicated that DHA reduced the expression of
osteoclast-derived factor LIF by inhibiting osteoclastogenesis,
thereby reducing the inhibition of bone sclerotin by LIF, reducing
abnormal bone formation, inhibiting the increase of subchondral
bone remodeling, and retaining the microstructure of the
subchondral bone to slow the degeneration of articular
cartilage.
Angiogenesis provides oxygen and nutrients for bone
formation, which is associated with bone remodeling for bone
homeostasis (50). However,
abnormal angiogenesis is a key pathological feature of OA in the
subchondral bone. Abnormal angiogenesis in subchondral bone
increases with increased bone remodeling, which leads to abnormal
bone formation (22,23). CD31, encoded by the platelet
endothelial cell adhesion molecule (PECAM1) gene, is a specific
marker of endothelial progenitor cells and is used to assess
angiogenesis (32). The present
study demonstrated that CD31-positive endothelial progenitor cells
in the subchondral bone significantly increased in number in the
ACLT+vehicle group. Previous studies have also revealed that DHA
has anti-angiogenic effects (27,53,54). After treatment with DHA,
CD31-positive staining significantly decreased in the ACLT+DHA
group relative to the ACLT+vehicle group. Previous tudies have
revealed that in the OA model of OVX rats, the expression of CD31
in the subchondral bone of the model group was significantly
increased, and the microstructural destruction of the subchondral
bone similarly with the expression of MMP-13 in the articular
cartilage was increased, and the loss of proteoglycan was severe
(9,12,55). The present study revealed a
significant increase in the number of CD31-positively labeled
endothelial progenitor cells in the subchondral bone of the
OVX+vehicle group compared with sham-operated group. However, the
role of DHA in OVX-induced abnormal angiogenesis of the subchondral
bone in OP-associated OA remains unclear. The present experiments
revealed that CD31 expression was significantly reduced in the
OVX+DHA group compared with the OVX+vehicle group. These results
indicated that DHA inhibited abnormal angiogenesis of subchondral
bone and slowed abnormal bone remodeling.
DHA is an effective drug against malaria, and a
semi-synthetic derivative of artemisinin with fewer side effects.
It performs an important role in inhibiting inflammation, is
anti-angiogenic, suppresses cancer and is anti-osteoclastogenic
(26-28). DHA suppresses the expression of
osteoclast marker genes, such as cathepsin K, calcitonin receptor
and tartrate resistant acid phosphatase (TRACP); DHA also inhibits
RANKL-induced osteoclast formation and bone resorption, thus
reversing the ovariectomized bone loss in oestrogen
deficient-induced osteoporosis of OVX mice (27). In addition, DHA effectively
inhibited osteoclastogenesis and prevented breast cancer-induced
osteolysis (28). Abnormal bone
remodeling and angiogenesis in the subchondral bone destroys its
microstructure and causes articular cartilage degeneration
(22,23). The present study revealed that DHA
reduced the expression of the osteoclast-derived factor LIF by
inhibiting osteoclastogenesis, thereby reducing inhibition of
sclerostin and thus suppressing abnormal subchondral bone
remodeling. In addition, DHA inhibited angiogenesis resulting in
less abnormal bone remodeling. Therefore, DHA attenuated articular
cartilage degeneration by inhibiting abnormal bone remodeling and
angiogenesis in OA mice.
In conclusion, DHA decreased the inhibition of
sclerostin by reducing LIF secretion by osteoclasts and thus
attenuated aberrant bone remodeling and inhibited angiogenesis in
the subchondral bone, thereby preserving the subchondral bone
microstructure and attenuating articular cartilage degeneration.
Future studies are required to further verify the underlying
mechanism of DHA decreasing the inhibition of sclerostin by
reducing LIF secretion by osteoclasts, such as anti-LIF
antibodies.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81660373).
Availability of data and materials
The datasets used during the current study are
available from the corresponding author on reasonable request.
Authors' contributions
QJ designed the experiment. LM, XZ, YL, JW and XY
conducted the main experiments. LM analyzed the data, drafted the
manuscript, and revised it. All the authors read and approved the
final submitted manuscript.
Ethics approval and consent to
participate
The experimental procedures were carried out in
accordance with the National Institutes of Health Guide for the
Care and Use of Laboratory Animals, and were approved by the
Scientific Research Ethics Committee of the General Hospital of
Ningxia Medical University (Yinchuan, China) (protocol no.
2016-147).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Martel-Pelletier J, Barr AJ, Cicuttini FM,
Conaghan PG, Cooper C, Goldring MB, Goldring SR, Jones G, Teichtahl
AJ and Pelletier JP: Osteoarthritis. Nat Rev Dis Primers.
2:160722016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Goldring SR and Goldring MB: Changes in
the osteochondral unit during osteoarthritis: Structure, function
and cartilage-bone crosstalk. Nat Rev Rheumatol. 12:632–644. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hugle T and Geurts J: What drives
osteoarthritis?-synovial versus subchondral bone pathology.
Rheumatology (Oxford). 56:1461–1471. 2017.
|
|
4
|
Huebner JL, Hanes MA, Beekman B, TeKoppele
JM and Kraus VB: A comparative analysis of bone and cartilage
metabolism in two strains of guinea-pig with varying degrees of
naturally occurring osteoarthritis. Osteoarthritis Cartilage.
10:758–767. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Eriksen EF: Cellular mechanisms of bone
remodeling. Rev Endocr Metab Disord. 11:219–227. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bailey AJ, Mansell JP, Sims TJ and Banse
X: Biochemical and mechanical properties of subchondral bone in
osteoarthritis. Biorheology. 41:349–358. 2004.PubMed/NCBI
|
|
7
|
Radin EL and Rose RM: Role of subchondral
bone in the initiation and progression of cartilage damage. Clin
Orthop Relat Res. 34–40. 1986.PubMed/NCBI
|
|
8
|
Bellido M, Lugo L, Roman-Blas JA,
Castañeda S, Caeiro JR, Dapia S, Calvo E, Largo R and
Herrero-Beaumont G: Subchondral bone microstructural damage by
increased remodelling aggravates experimental osteoarthritis
preceded by osteoporosis. Arthritis Res Ther. 12:R1522010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu S, Chen K, Lan Y, Zhang N, Jiang R and
Hu J: Alendronate protects against articular cartilage erosion by
inhibiting subchondral bone loss in ovariectomized rats. Bone.
53:340–349. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Khorasani MS, Diko S, Hsia AW, Anderson
MJ, Genetos DC, Haudenschild DR and Christiansen BA: Effect of
alendronate on post-traumatic osteoarthritis induced by anterior
cruciate ligament rupture in mice. Arthritis Res Ther. 17:302015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Miyauchi Y, Sato Y, Kobayashi T, Yoshida
S, Mori T, Kanagawa H, Katsuyama E, Fujie A, Hao W, Miyamoto K, et
al: HIF1alpha is required for osteoclast activation by estrogen
deficiency in postmenopausal osteoporosis. Proc Natl Acad Sci USA.
110:16568–16573. 2013. View Article : Google Scholar
|
|
12
|
Cui Z, Xu C, Li X, Song J and Yu B:
Treatment with recombinant lubricin attenuates osteoarthritis by
positive feedback loop between articular cartilage and subchondral
bone in ovariectomized rats. Bone. 74:37–47. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li X, Zhang Y, Kang H, Liu W, Liu P, Zhang
J, Harris SE and Wu D: Sclerostin binds to LRP5/6 and antagonizes
canonical Wnt signaling. J Biol Chem. 280:19883–19887. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
van Bezooijen RL, ten Dijke P, Papapoulos
SE and Löwik CW: SOST/sclerostin, an osteocyte-derived negative
regulator of bone formation. Cytokine Growth Factor Rev.
16:319–327. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jia H, Ma X, Wei Y, Tong W, Tower RJ,
Chandra A, Wang L, Sun Z, Yang Z, Badar F, et al: Loading-induced
reduction in sclerostin as a mechanism of subchondral bone plate
sclerosis in mouse knee joints during late-stage osteoarthritis.
Arthritis Rheumatol. 70:230–241. 2018. View Article : Google Scholar
|
|
16
|
Robling AG, Niziolek PJ, Baldridge LA,
Condon KW, Allen MR, Alam I, Mantila SM, Gluhak-Heinrich J, Bellido
TM, Harris SE and Turner CH: Mechanical stimulation of bone in vivo
reduces osteocyte expression of Sost/sclerostin. J Biol Chem.
283:5866–5875. 2008. View Article : Google Scholar
|
|
17
|
Lin C, Jiang X, Dai Z, Guo X, Weng T, Wang
J, Li Y, Feng G, Gao X and He L: Sclerostin mediates bone response
to mechanical unloading through antagonizing Wnt/beta-catenin
signaling. J Bone Miner Res. 24:1651–1661. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tu X, Rhee Y, Condon KW, Bivi N, Allen MR,
Dwyer D, Stolina M, Turner CH, Robling AG, Plotkin LI and Bellido
T: Sost downregulation and local Wnt signaling are required for the
osteogenic response to mechanical loading. Bone. 50:209–217. 2012.
View Article : Google Scholar
|
|
19
|
Sims NA and Johnson RW: Leukemia
inhibitory factor: A paracrine mediator of bone metabolism. Growth
Factors. 30:76–87. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Koide M, Kobayashi Y, Yamashita T, Uehara
S, Nakamura M, Hiraoka BY, Ozaki Y, Iimura T, Yasuda H, Takahashi N
and Udagawa N: Bone formation is coupled to resorption via
suppression of sclerostin expression by osteoclasts. J Bone Miner
Res. 32:2074–2086. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Burr DB and Gallant MA: Bone remodelling
in osteoarthritis. Nat Rev Rheumatol. 8:665–673. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cui Z, Crane J, Xie H, Jin X, Zhen G, Li
C, Xie L, Wang L, Bian Q, Qiu T, et al: Halofuginone attenuates
osteoarthritis by inhibition of TGF-beta activity and H-type vessel
formation in subchondral bone. Ann Rheum Dis. 75:1714–1721. 2016.
View Article : Google Scholar
|
|
23
|
Zhen G, Wen C, Jia X, Li Y, Crane JL,
Mears SC, Askin FB, Frassica FJ, Chang W, Yao J, et al: Inhibition
of TGF-beta signaling in mesenchymal stem cells of subchondral bone
attenuates osteoarthritis. Nat Med. 19:704–712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ji B, Zhang Z, Guo W, Ma H, Xu B, Mu W,
Amat A and Cao L: Isoliquiritigenin blunts osteoarthritis by
inhibition of bone resorption and angiogenesis in subchondral bone.
Sci Rep. 8:17212018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Klayman DL: Qinghaosu (artemisinin): An
antimalarial drug from China. Science. 228:1049–1055. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tu Y: The development of new antimalarial
drugs: Qinghaosu and dihydro-qinghaosu. Chin Med J (Engl).
112:976–977. 1999.
|
|
27
|
Zhou L, Liu Q, Yang M, Wang T, Yao J,
Cheng J, Yuan J, Lin X, Zhao J, Tickner J and Xu J:
Dihydroartemisinin, an anti-malaria drug, suppresses estrogen
deficiency-induced osteoporosis, osteoclast formation, and
RANKL-induced signaling pathways. J Bone Miner Res. 31:964–974.
2016. View Article : Google Scholar
|
|
28
|
Feng MX, Hong JX, Wang Q, Fan YY, Yuan CT,
Lei XH, Zhu M, Qin A, Chen HX and Hong D: Dihydroartemisinin
prevents breast cancer-induced osteolysis via inhibiting both
breast caner cells and osteoclasts. Sci Rep. 6:190742016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Glasson SS, Blanchet TJ and Morris EA: The
surgical destabilization of the medial meniscus (DMM) model of
osteoarthritis in the 129/SvEv mouse. Osteoarthritis Cartilage.
15:1061–1069. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao YG, Wang Y, Guo Z, Gu AD, Dan HC,
Baldwin AS, Hao W and Wan YY: Dihydroartemisinin ameliorates
inflammatory disease by its reciprocal effects on Th and regulatory
T cell function via modulating the mammalian target of rapamycin
pathway. J Immunol. 189:4417–4425. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou S, Wang G, Qiao L, Ge Q, Chen D, Xu
Z, Shi D, Dai J, Qin J, Teng H and Jiang Q: Age-dependent
variations of cancellous bone in response to ovariectomy in
C57BL/6J mice. Exp Ther Med. 15:3623–3632. 2018.PubMed/NCBI
|
|
32
|
Mapp PI and Walsh DA: Mechanisms and
targets of angiogenesis and nerve growth in osteoarthritis. Nat Rev
Rheumatol. 8:390–398. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xie H, Cui Z, Wang L, Xia Z, Hu Y, Xian L,
Li C, Xie L, Crane J, Wan M, et al: PDGF-BB secreted by
preosteoclasts induces angiogenesis during coupling with
osteogenesis. Nat Med. 20:1270–1278. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Matsuo K and Irie N: Osteoclast-osteoblast
communication. Arch Biochem Biophys. 473:201–209. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sims NA and Martin TJ: Coupling the
activities of bone formation and resorption: A multitude of signals
within the basic multicellular unit. Bonekey Rep. 3:4812014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kamekura S, Hoshi K, Shimoaka T, Chung U,
Chikuda H, Yamada T, Uchida M, Ogata N, Seichi A, Nakamura K and
Kawaguchi H: Osteoarthritis development in novel experimental mouse
models induced by knee joint instability. Osteoarthritis Cartilage.
13:632–641. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Honma M, Ikebuchi Y, Kariya Y and Suzuki
H: Regulatory mechanisms of RANKL presentation to osteoclast
precursors. Curr Osteoporos Rep. 12:115–120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Miyatake K, Muneta T, Ojima M, Yamada J,
Matsukura Y, Abula K, Sekiya I and Tsuji K: Coordinate and
synergistic effects of extensive treadmill exercise and ovariectomy
on articular cartilage degeneration. BMC Musculoskelet Disord.
17:2382016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sniekers YH, Weinans H, Bierma-Zeinstra
SM, van Leeuwen JP and van Osch GJ: Animal models for
osteoarthritis: The effect of ovariectomy and estrogen treatment-a
systematic approach. Osteoarthritis Cartilage. 16:533–541. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bijlsma JW, Berenbaum F and Lafeber FP:
Osteoarthritis: An update with relevance for clinical practice.
Lancet. 377:2115–2126. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Goldring MB and Goldring SR: Articular
cartilage and subchondral bone in the pathogenesis of
osteoarthritis. Ann N Y Acad Sci. 1192:230–237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bultink IE and Lems WF: Osteoarthritis and
osteoporosis: What is the overlap? Curr Rheumatol Rep. 15:3282013.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Baron R and Kneissel M: WNT signaling in
bone homeostasis and disease: From human mutations to treatments.
Nat Med. 19:179–192. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Song L, Liu M, Ono N, Bringhurst FR,
Kronenberg HM and Guo J: Loss of wnt/β-catenin signaling causes
cell fate shift of preosteoblasts from osteoblasts to adipocytes. J
Bone Miner Res. 27:2344–2358. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
van Bezooijen RL, Roelen BA, Visser A, van
der Wee-Pals L, de Wilt E, Karperien M, Hamersma H, Papapoulos SE,
ten Dijke P and Löwik CW: Sclerostin is an osteocyte-expressed
negative regulator of bone formation, but not a classical BMP
antagonist. J Exp Med. 199:805–814. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li X, Ominsky MS, Warmington KS, Morony S,
Gong J, Cao J, Gao Y, Shalhoub V, Tipton B, Haldankar R, et al:
Sclerostin anti-body treatment increases bone formation, bone mass,
and bone strength in a rat model of postmenopausal osteoporosis. J
Bone Miner Res. 24:578–588. 2009. View Article : Google Scholar
|
|
47
|
Wu L, Guo H, Sun K, Zhao X, Ma T and Jin
Q: Sclerostin expression in the subchondral bone of patients with
knee osteoarthritis. Int J Mol Med. 38:1395–1402. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Walker EC, McGregor NE, Poulton IJ, Solano
M, Pompolo S, Fernandes TJ, Constable MJ, Nicholson GC, Zhang JG,
Nicola NA, et al: Oncostatin M promotes bone formation
independently of resorption when signaling through leukemia
inhibitory factor receptor in mice. J Clin Invest. 120:582–592.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Poulton IJ, McGregor NE, Pompolo S, Walker
EC and Sims NA: Contrasting roles of leukemia inhibitory factor in
murine bone development and remodeling involve region-specific
changes in vascularization. J Bone Miner Res. 27:586–595. 2012.
View Article : Google Scholar
|
|
50
|
Johnson RW, Brennan HJ, Vrahnas C, Poulton
IJ, McGregor NE, Standal T, Walker EC, Koh TT, Nguyen H, Walsh NC,
et al: The primary function of gp130 signaling in osteoblasts is to
maintain bone formation and strength, rather than promote
osteoclast formation. J Bone Miner Res. 29:1492–1505. 2014.
View Article : Google Scholar
|
|
51
|
Li X, Ominsky MS, Niu QT, Sun N, Daugherty
B, D'Agostin D, Kurahara C, Gao Y, Cao J, Gong J, et al: Targeted
deletion of the sclerostin gene in mice results in increased bone
formation and bone strength. J Bone Miner Res. 23:860–869. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
de Vries TJ and Huesa C: The osteocyte as
a novel key player in understanding periodontitis through its
expression of RANKL and sclerostin: A review. Curr Osteoporos Rep.
17:116–121. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Dong F, Zhou X, Li C, Yan S, Deng X, Cao
Z, Li L, Tang B, Allen TD and Liu J: Dihydroartemisinin targets
VEGFR2 via the NF-kB pathway in endothelial cells to inhibit
angiogenesis. Cancer Biol Ther. 15:1479–1488. 2014. View Article : Google Scholar
|
|
54
|
Ho WE, Peh HY, Chan TK and Wong WS:
Artemisinins: Pharmacological actions beyond anti-malarial.
Pharmacol Ther. 142:126–139. 2014. View Article : Google Scholar
|
|
55
|
Bei MJ, Tian FM, Xiao YP, Cao XH, Liu N,
Zheng ZY, Dai MW, Wang WY, Song HP and Zhang L: Raloxifene retards
cartilage degradation and improves subchondral bone
micro-architecture in ovariectomized rats with patella
baja-induced-patellofemoral joint osteoarthritis. Osteoarthritis
Cartilage. 28:344–355. 2020. View Article : Google Scholar
|