Introduction
Gastric cancer (GC) is the fifth most frequently
diagnosed malignancy and the third leading cause of
cancer-associated mortality worldwide (1). According to cancer statistics in
China in 2015, GC is the second and the third most commonly
diagnosed type of cancer among males and females, respectively, in
the Chinese population (2).
Despite improvements in the clinical treatment of GC in recent
years, the survival rate of patients with GC remains <30%
(2). As researchers spare no
efforts to study cancer progression, views on tumor development
have markedly changed, and the malignant cells themselves been
shown to make up a complex area, termed the tumor microenvironment
(TME) (3). Recent evidence has
indicated that the crosstalk between tumor cells and the TME is
essential for tumor progression, invasion and metastasis (4).
Inflammation is regarded as one of the hallmarks of
cancer; inflammatory cells and associated factors play an essential
role in almost every stage of tumor progression, including
metastasis (5). Neutrophils, the
most abundant type of white blood cells in the peripheral
bloodstream, are important components of tumor-associated
infiltrating inflammatory and immune cells in GC (6). Neutrophils used to be considered
inert bystander cells in cancer progression and development,
however this view has changed (7,8).
Advances in neutrophil biology have revealed that neutrophils can
release their decondensed chromatin and form large extracellular
DNA networks, termed neutrophil extracellular traps (NETs).
Intensive investigations on NETs have demonstrated a potential
involvement of NETs in neoplastic disease. The pro-tumor and
antitumor effects of NETs have been described (9). It has been reported that tumor
cells and cancer cell-primed platelets can promote the release of
NETs by host neutrophils (10,11). In patients with Ewing's sarcoma,
the presence of tumor-associated neutrophils and NETs is only
detected in patients with metastasis, indicating that NETs may
promote tumor progression (12).
In a study comparing human triple-negative breast cancer with
luminal and HER-2+ breast cancer, it was identified that
the presence of NETs was associated with the metastatic burden, and
the triple-negative breast cancer cases exhibited the highest
number of NETs (13).
Furthermore, it has been observed that circulating NETs, produced
upon stimulation in infectious and non-infectious diseases, capture
circulating tumor cells, which then promotes tumor progression and
metastasis in vitro and in vivo (14). The degradation or inhibition of
NETs by DNAse-1 or neutrophil elastase (NE) inhibitor has been
demonstrated to prevent metastasis in a murine cecal ligation and
puncture model (15).
Citrullinated histone H3 (cit-h3; a NETs activation marker) is
detectable in the plasma of patients with advanced cancer, and can
serve as a distinct prognostic marker that is associated with a
poor clinical outcome in numerous types of tumor (16). After deposition of NETs was found
to be significant in malignant tumors, it was also identified that
NETs increase the risk of thrombosis in patients with cancer, which
may be associated with a poor prognosis (17,18). Conversely, the antitumor effects
of NETs have also been reported in colon cancer and melanoma.
Following the stimulation of tumor cells with NETs, the ability to
metastasize and proliferate is decreased (19,20). In addition, NETs may act as a
double-edged sword in cancer biology and the effects of NETs on
tumor cells may vary in different types of tumors. To the best of
our knowledge, there is no clear conclusion on the interaction
between NETs and tumors or whether NETs function as a pro-tumor or
antitumor factor. The present study aimed to provide evidence to
uncover the role of NETs in GC progression.
Metastatic progression is regulated by alterations
in the TME, such as inflammation, angiogenesis, and cancerized
stroma and intravasation, which is termed epithelial-mesenchymal
transition (EMT) (21). EMT in
tumor progression allows a polarized epithelial cell to acquire a
mesenchymal cell phenotype, and it facilitates the intravasation of
tumor cells into blood or lymph vessels and the subsequent
formation of distant metastasis (22). Neutrophils have been observed to
promote the EMT process (23);
however, the underlying mechanisms remain unclear. Related factors
released by neutrophils, such as interleukin (IL)-17, can promote
the EMT process of tumor cells via the JAK2/STAT3 pathway (24). NETs have been reported to be able
to promote tumor cell adhesion and thus promote tumor cell
progression and metastasis (25). In the present study, a large
amount of NETs formation was found in the plasma of patients with
advanced GC and in GC tumor tissues; however, the interaction
between tumor and NETs needs to be further investigated. The aim of
the present study was to reveal the role of NETs in the progression
and metastasis of GC.
Materials and methods
Patients
Patients with gastric cancer and healthy control
subjects were recruited at the Department of Gastrointestinal
Surgery in the Second Affiliated Hospital of Harbin Medical
University of China, between July, 2017 to December, 2019. Patients
were diagnosed with GC by a histological diagnosis which involved
post-operative pathology and tumor-node-metastasis (TNM) staging
was assessed according to the 8th American Joint Committee on
Cancer (AJCC) Staging Classification Guidelines (26). The exclusion criteria were as
follows: An age <18 years, pregnancy, cardiovascular disease,
diabetes, active or chronic infection, liver or renal dysfunction,
other coexisting cancers, platelets and/or blood coagulation
disorders, and the administration of anticoagulant and/or
anti-platelet treatment. The main characteristics of the patients
and healthy controls are presented in Table I. The present study was approved
by the Research Ethics Committee of Second Affiliated Hospital of
Harbin Medical University, all subjects manually signed the
informed consent.
 | Table IClinicopathological characteristics
between the GC group and normal group. |
Table I
Clinicopathological characteristics
between the GC group and normal group.
| Variables | Control (n=16) | Stage I (n=12) | Stage II
(n=12) | Stage III
(n=14) | Stage IV
(n=13) |
|---|
| Sex
(male/female) | 12/4 | 8/4 | 8/4 | 10/4 | 10/3 |
| Age (years) | 55.67±9.39 | 63.83±6.99 | 60.83±10.63 | 58.67±14.14 | 61.00±9.95 |
| Leukocyte
(109/l) | 5.28±1.09 | 5.09±1.14 | 6.25±1.37 | 5.89±1.65 | 6.13±1.57 |
| Neutrophil
(109/l) | 3.50±0.99 | 3.36±0.96 | 4.26±1.77 | 4.89±2.29 | 4.19±1.46 |
| Neutrophils
(%) | 60.57±8.85 | 60.88±9.74 | 57.11±10.77 | 65.44±12.18 | 65.82±9.00 |
| CEA (0-5
ng/ml) | 2.71
(0.39-4.07) | 3.08
(0.46-11.55) | 2.51
(0.59-5.96) | 5.05a–c(0.73-25.33) | 5.92a–c (1.68-26.39) |
| CA724 (0-6.9
U/ml) | 2.67
(0.86-5.89) | 2.69
(0.78-5.54) | 3.19
(0.98-11.35) | 12.65a–c (0.63-48.43) | 50.48a–c (0.81-302.17) |
| CA199 (0-37
U/ml) | 6.86
(2.58-18.56) | 7.16
(0.61-14.08) | 78.92
(0.59-832.5) | 154.31a–c (3.24-1009.36) | 281.74a–c(5.71-1127.25) |
| MPO-DNA (405
nm) | 0.16±0.05 | 0.22±0.06 | 0.22±0.08 | 0.55±0.17a,b,c | 0.60±0.20a–c |
| NE (ng/ml) | 0.49±0.31 | 0.55±0.33 | 0.72±0.44 | 1.99±0.53a,b,c | 1.96±0.61a–c |
| cf-DNA
(µg/ml) | 0.11±0.05 | 0.12±0.07 | 0.16±0.07 | 0.55±0.15a,b,c | 0.57±0.16a–c |
|
Differentiation | | | | | |
| Well | / | 4 | 5 | 5 | / |
| Moderate | / | 4 | 4 | 6 | / |
| Poor | / | 4 | 3 | 3 | / |
| Lymphovascular | / | 2 | 3 | 9 | 9 |
| Invasion | | | | | |
| Liver
metastasis | / | 0 | 0 | 0 | 5 |
| Peritoneal
metastasis | / | 0 | 0 | 0 | 6 |
| Lung
metastasis | / | 0 | 0 | 0 | 2 |
| Bone
metastasis | / | 0 | 0 | 0 | 1 |
Cells and cell culture
The human GC cell line, AGS, was obtained from the
Cell Bank of Chinese Academy of Science. Cells were cultured in
RPMI-1640 medium (#31870082, Gibco; Thermo Fisher Scientific, Inc.)
containing 10% fetal bovine serum (FBS, #04-001-1ACS, BI)
supplemented with penicillin (100 U/ml) and streptomycin (#ST488,
Beyotime Institute of Biotechnology, Inc.; 100 U/ml) at 37°C with
5% CO2 in a humidified atmosphere.
Isolation of neutrophils
Fresh peripheral blood was obtained from patients
with GC and healthy controls using the commercial EDTA-k2
anticoagulant tubes routinely used at the Second Affiliated
Hospital of Harbin Medical University. Blood samples were obtained
within 2 h prior to isolation. Human neutrophils were isolated
using polymorphprep™ (27).
Briefly, 5 ml of fresh whole blood were layered on top of 5 ml of
PolymorphPrep (#1114683, Axis-Shield) in a 15 ml tube and
centrifuged at 500 × g for 30 min at room temperature. The lower
leukocyte band containing neutrophils was collected, washed and
resuspended into 5 ml of ACK lysis buffer (#C3702, Beyotime
Institute of Biotechnology, Inc.), washed twice in HBSS without
Ca2+/Mg2+, and finally resuspended in
serum-free RPMI-1640 (#31870082, Gibco; Thermo Fisher Scientific,
Inc.).
Stimulation and inhibition of NET
formation
Purified neutrophils (500,000) isolated from GC
patients or healthy controls were subsequently incubated at 37°C in
5% CO2 then activated overnight with 25 nM PMA (#P1585,
phorbol 12-myristate 13-acetate, Sigma-Aldrich; Merck KGaA). In the
inhibition assay, PMA and the inhibitors were used in combination.
NETs inhibitors were added 30 min prior to neutrophil activation to
inhibit NETs formation. The PAD4 inhibitor, GSK484 (#17488, Cayman
Chemical Company) was used at 10 µM to inhibit NETs
formation, and 1.5 units/ml of DNase I (#04536282001,
Sigma-Aldrich; Merck KGaA) was used to digest NETs scaffolds. The
supernatant containing NETs was collected and centrifugation at 250
× g for 10 min at room temperature. Isolated NETs were stored at
−80°C until further use as previously reported (28). To investigate the effect of the
TME on NETs formation, neutrophils were treated using 20% plasma
from the same patients with GC (n=20) or the controls (n=10) as
above, following a previously described protocol (29).
Immunofluorescence in neutrophils
To further assess NETs formation in vitro,
neutrophils were seeded on poly-L-lysine-coated coverslips
(#354085, Corning, Inc.) and stimulated with PMA for 3 h and then
fixed with 4% paraformaldehyde (PFA, #BL539A, Biosharp) for 20 min
at room temperature, rinsed twice in PBS, incubated in 50 mM of
NH4Cl for 10 min at room temperature and permeabilized
with 0.5% Triton X-100 (#BB151-500, Thermo Fisher Scientific, Inc.)
for 1 min at room temperature. The cells were then blocked in PBS
containing 1% bovine serum albumin (BSA, #A3294, Sigma-Aldrich;
Merck KGaA) for 30 min at 37°C and incubated with neutrophil
elastase (NE) antibody (#sc-55549, Santa Cruz Biotechnology, Inc.)
at 1:400 dilution in blocking buffer for 1 h at room temperature.
Following washes 3 times in PBS, the cells were incubated in the
presence of fluorochrome-conjugated secondary antibodies (#A11001,
Alexa Fluor 488, Invitrogen; Thermo Fisher Scientific, Inc.) at
1:200 dilution at room temperature for 1 h, rinsed twice in PBS,
stained with Hoechst 33342 (#C1022, Beyotime Institute of
Biotechnology) at 1:5 dilution at room temperature for 5 min,
rinsed in PBS, and the coverslips were mounted onto glass slides
using a fluorescence microscope (DM400B, Leica Microsystems GmbH).
The percentage of NETs was evaluated by counting the number of
NET-releasing neutrophils out of the total number of
neutrophils.
Measurement of the formation of NETs in
patients with GC
Circulating cell-free DNA (cfDNA), NE and MPO-DNA
complex, were all measured as a presence for NETs in the subjects'
plasma (30,31). The Quant-iT PicoGreen dsDNA Assay
kit (#P11496, Invitrogen; Thermo Fisher Scientific, Inc.) was used
to quantify the GC patient cfDNA levels in plasma as per the
provided directions. Human plasma NE was quantified using selective
ELISA (#MM-1433H1, Jiangsu Meimian Industrial Co., Ltd.) based on
provided instruction. To quantify the MPO-DNA complex in the
plasma, a capture ELISA was used as previously described (32). In brief, 5 µg/ml anti-MPO
antibody (#sc-52707, Santa Cruz Biotechnology, Inc.) was coated
onto 96-well plates (dilution 1:500 in 50 µl) at 4°C
overnight. After washing with PBS 3 times (300 µl each), 20
µl of samples were added to the wells with 80 µl
incubation buffer containing a peroxidase-labeled anti DNA mAb
(cat. no. 11774425001, Cell Death ELISAplus, Roche
Diagnostics; dilution, 1:25). The plate was incubated for 2 h, and
shaking at 300 rpm at room temperature. Following 3 washes with PBS
(300 µl each), 100 µl peroxidase substrate (ABST) of
the kit was added. The absorbance at 405 nm wavelength was measured
following 20 min of incubation at room temperature in the dark.
Cell cycle analysis
Following treatment with NETs or NETs inhibitors for
24 h, the AGS cells were washed with cold PBS and then fixed
overnight with 70% cold ethanol at −20°C. The cells were then
washed again with cold PBS and incubated in the dark with propidium
iodide (PI, #ST511, Beyotime Institute of Biotechnology) staining
solution containing RNase A for 30 min at room temperature. The
cell cycle was measured using a FACSCanto II flow cytometer (BD
Biosciences). The proportion of cells in the growth 0/growth
1(G0/G1), synthesis (S) and growth 2/mitosis (G2/M) phase were
calculated using FlowJo software programs (version 10), and are
represented as DNA histograms.
Migration assay
For the migration assay, 2×105cancer
cells were loaded into the upper chamber of a cell culture insert
with 8 µM pore size (#3422, Corning Inc.) with NETs culture
medium (CM) with or without NETs inhibitors and the lower chamber
was supplemented with cell culture medium containing 10% FBS.
Following 24 h of incubation at 37°C, the migrated cells were fixed
and stained with crystal violet staining solution (#G1063, Beijing
Solarbio Science & Technology Co., Ltd.) at room temperature
and were counted under a microscope (Olympus Corp.).
MTT assay
The AGS cells were plated in 96-well plates at a
density of 3×103per well. After the cells had reached
approximately 80% confluency, they were starved overnight. The
cells were then exposed to NETs CM with or without inhibitors for
24 and 48 h, respectively. Following incubation for 24 or 48 h at
37°C, MTT (#M2128, Sigma-Aldrich; Merck KGaA; 20 µl/well)
solution was added, and the cells were incubated for 6 h at 37°C.
The medium in the plate was then discarded, and 150 µl
dimethylsulfoxide (DMSO) were added to each well. The absorbance
was then measured at 570 nm using the PowerWave HT microplate
spectrophotometer (BioTek Instruments, Inc.).
Post-surgical residual tumor xenograft
models
Male athymic BALB/c nude mice (n=20, 5-6 weeks old,
weighing 20-25 g) were purchased from Beijing Vital River
Laboratory Animal Technology Co. Ltd. [Certificate no. SCXK (Jing)
2016-0011 (no. 11400700161156)]. All animal experiments were
performed in accordance with the NIH guidelines (Guide for the Care
and Use of Laboratory Animals) and approved by the Ethics Committee
of the Second Affiliated Hospital of Harbin Medical University. The
Ethics Committee of the Second Affiliated Hospital of Harbin
Medical University approved the present study (permit no.
KY2016-032). Mice were kept in a controlled SPF environment at
23±2°C, 40-70% humidity under a 12 h dark/light cycle with free
access to irradiated food and sterile water. They were housed in
individually ventilated cages: 5 per cage, with 4-6 mm corncob
bedding following disinfection with 60Co radiation. A
murine model was established with a post-surgical residual tumor to
mimic the recurrence of a solid tumor. The AGS tumor cells
(>5×107/ml) were harvested and washed with saline. A
cell suspension of 1×106 in 0.2 ml was injected
subcutaneously into the right axillary of the mice to establish the
traditional tumor xenograft nude mouse model (n=5). When the tumors
reached an average volume of 1,000-1,500 mm3, the tumors
were removed from the mice aseptically and diced into small cubes
(2.0×2.0×2.0 mm). These tumor chunks were then injected
subcutaneously into the right axillary of the new mice (n=15) to
establish a murine model with a post-surgical residual tumor. When
the tumors had reached an average volume of 300-350 mm3,
the mice were anesthetized by intraperitoneal injection of 1.2%
avertin (2, 2, 2-tribromoethanol) solution (260 mg/kg). A small
incision was made to remove tumor tissues around the tumor edge
aseptically, leaving a residual tumor volume of 60-100
mm3. After the surgery, the mice were divided into 3
groups (n=5) based on the tumor volume. Body weight was measured
twice a week. The mice in the NETs inhibition treatment groups were
administered daily with DNAse-1 or GSK484 by an intraperitoneal
injection at a dose of 15,000 units/kg DNAse-1 or 20 mg/kg GSK484
for 14 days. The control mice were treated with 0.1% DMSO in 100
µl saline. Tumor diameters was measured in two dimensions
every 2-3 days. Tumor volume was calculated according to the
formula: Volume (mm3)=(width2 × length)/2.
The mice were sacrificed at 14 days after the injection of DNAse-1
or GSK484. At the end of the experiments, the mice were euthanized
by cervical dislocation. Tumors were excised, weighed, and fixed in
4% paraformaldehyde at room temperature for the paraffin sections
or stored at -80°C.
Western blot analysis
Whole cell lysates were extracted using RIPA lysis
buffer (#P0013C, Beyotime Institute of Biotechnology, Inc.) with
protease inhibitor (Roche Diagnostics). The quantities of protein
were determined using a bicinchoninic acid kit (#P0012, Beyotime
Institute of Biotechnology). Protein samples of 100 µg per
lane were loaded onto 10% SDS-PAGE gels and transferred onto
nitrocellulose membranes (PALL) for 90 min. For immunodetection,
the membranes were incubated with the following primary antibodies
at 4°C overnight: E-cadherin (#ab76055, Abcam) at a 1:1,000
dilution, vimentin (#HPA001762, Sigma-Aldrich; Merck KGaA) at a
1:1,000 dilution, and cit-h3 (#ab5103, Abcam), GAPDH (#60004-1-Ig,
Proteintech) at a 1:1,000 dilution. The fluorescence-conjugated
secondary IRDye800 mouse (#962-32210) and rabbit antibodies
(#962-32211) were purchased from LI-COR. Following incubation with
secondary antibody at 1:7,500 dilution with gentle shaking at room
temperature for 60 min, the membranes were scanned by Odyssey
Infrared Imaging System (LI-COR Biosciences). Protein expression
was quantified using Image-Pro® Plus software (version
6.0; Media cybernetics, Inc.). All experiments were performed at
least 5 times.
Immunohistochemistry
For immunohistochemistry, heat-induced epitope
retrieval was performed by Citrate Antigen Retrieval solution
(#C1031, Beijing Solarbio Science & Technology Co., Ltd.) for
40 min for vimentin, or by EDTA Antigen Retrieval solution
(#ZLI-9079, ZSGB-Bio) for E-cadherin. The tissue preparations were
incubated with the primary antibodies for E-cadherin (#20874-1-AP,
ProteinTech Group, Inc.,) at a 1:500 dilution, vimentin
(#HPA001762, Sigma-Aldrich; Merck KGaA) at a 1:250 dilution at 4°C
overnight, followed by incubation with the secondary antibody
EnVision™+/HRP rabbit polymer (#P0448, Dako; Agilent Technologies,
Inc.) at a 1:200 dilution at room temperature for 30 min. Secondary
antibody detection was performed by using the SIGMAFAST™
3,3′-diaminobenzidine tablets (DAB Peroxidase Substrate Tablet Set,
#D4168, Sigma-Aldrich; Merck KGaA). The slides were counterstained
with hematoxylin for 2 min following color separation by 1% acetic
acid at room temperature for 30 sec. The Samples were visualized
under a light microscope (Nikon, model Eclipse E400) and images
were captured using a Nikon Digital Camera (ACT-1 Nikon
software).
Immunofluorescence in tissues
To detect NETs by immunofluorescence, sections were
stained as previously described (33). Non-specific binding sites were
blocked with 2% goat serum in 2% BSA-PBS. In human tissue staining,
NETs were counterstained with a rabbit anti-citrullinated histone
H3 polyclonal antibody (#ab5103, Abcam) at 1:200 dilution, a mouse
anti-NE monoclonal antibody (#sc-55548, Santa Cruz Biotechnology,
Inc.) at 1:200 dilution as the primary antibodies at 4°C overnight.
The samples were incubated with goat anti-mouse IgG (H+L) Highly
Cross-Adsorbed Secondary Antibody, Alexa Fluor 488 (#A-11032,
Invitrogen™, Thermo Fisher Scientific, Inc.), or goat anti-rabbit
IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor 594
(#A32740, Invitrogen™, Thermo Fisher Scientific, Inc.) at a 1:200
dilution at room temperature for 1 h. Nuclei were stained with
4′,6-diamidino-2-phenylindole (#C1002, Beyotime Institute of
Biotechnology) at 1:5 dilution at room temperature for 5 min. The
NETs staining methods for the mouse tumor tissues were consistent
with those in human tissues, except that antibody species were
different. For briefly, NETs were counterstained with a rabbit
anti-citrullinated histone H3 antibody (#ab5103, Abcam) at 1:200
dilution, a rabbit anti-NE antibody (#AF-0010, Affinity
Biosciences) at a 1:200 dilution as the primary antibodies at 4°C
overnight. F(ab′)2-goat anti-rabbit IgG (H+L) Alexa Fluor 488
antibody (#A11070, Invitrogen™, Thermo Fisher Scientific, Inc.) and
F(ab′)2-goat anti-rabbit IgG (H+L) Alexa Fluor 594 antibody
(#A-11072, Invitrogen™, Thermo Fisher Scientific, Inc.) were used
at a 1:500 dilution at room temperature for 45 min. Mouse sections
staining followed the method as previously reported (34). NETs were defined as co-localized
DNA, NE and citrullinated histone H3(cit-h3) after acquiring images
with a laser scanning confocal microscope (LSCM; Olympus
Corp.).
Statistical analysis
Statistical analysis was performed using GraphPad
Software 5.0 (GraphPad Software, Inc.). Comparisons between 2
groups were conducted using a Student's t-test. Differences between
multiple groups were determined by one-way ANOVA with Tukey's post
hoc test. A probability value of P<0.05 was considered to
indicate a statistically significant difference.
Results
Comparison of the clinicopathological
characteristics of patients with different stages of GC
A total of 51 patients with GC, including 12 cases
at stage I, 12 cases at stage II, 14 cases at stage III and 13
cases at stage IV, along with 16 healthy controls were recruited in
this prospective study. The patient clinical information is
presented in Table I and no
significant differences were observed in age, sex, and leukocyte
and neutrophil counts among the different groups (P>0.05).
Peritoneal metastasis was present in 46.1% of patients with stage
IV GC, liver metastasis was found in 38.5% (2 cases of liver
metastasis were accompanied by peritoneal metastasis), and
metastasis in other organs was present in 23% of cases. In the
present study, the overall incidence of lymphovascular invasion
(LVI) in all enrolled patients was 45%, with a positive rate of
64.3% in patients with stage III and 69.2% in patients with stage
IV GC.
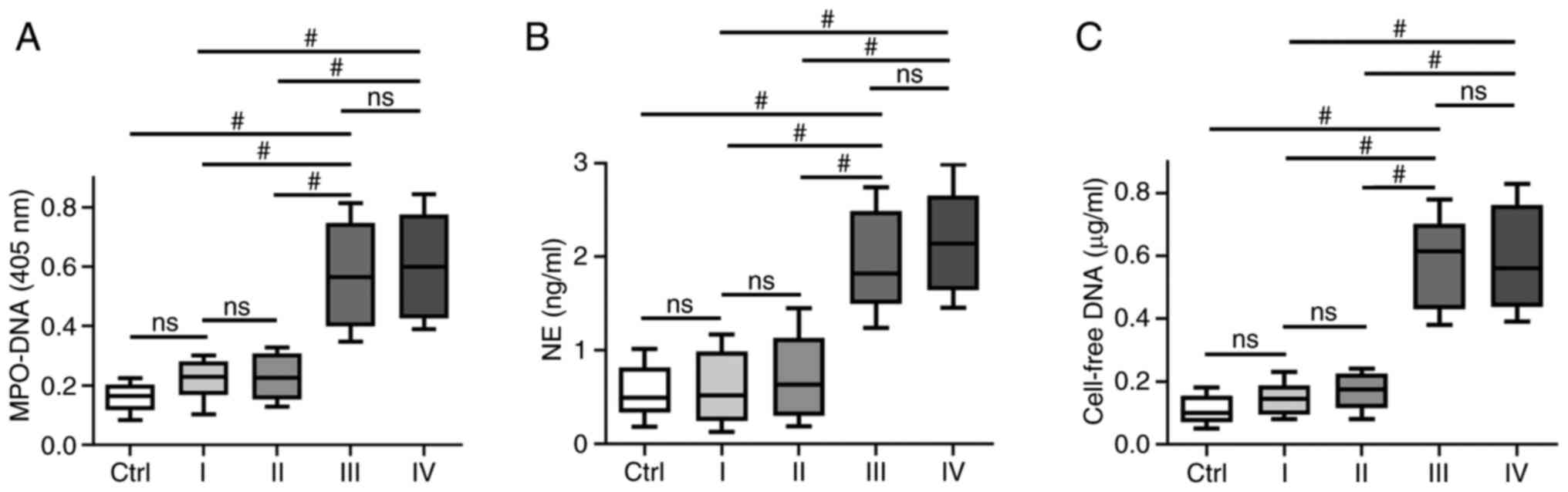
Association between circulating NETs and
clinical TNM stage in GC
NETs are extracellular web-like structures of DNA
decorated with various antimicrobial proteins, such as
neutrophil-derived NE and myeloperoxidase (MPO). To identify NETs
formation in plasma, the levels of different NETs markers, such as
circulating MPO-DNA complexes, cf-DNA and NE were assessed. It was
identified that the levels of MPO-DNA complexes, cf-DNA and NE were
higher in patients with stage III/IV GC (P<0.001) compared with
patients with stage I/II GC or the healthy controls; however, no
significant difference was observed between patients with stage
I/II GC and the healthy controls. In addition, no significant
difference was identified in NETs markers, such as MPO-DNA complex,
cf-DNA and NE in patients GC with stage III and IV disease
(Fig. 1).
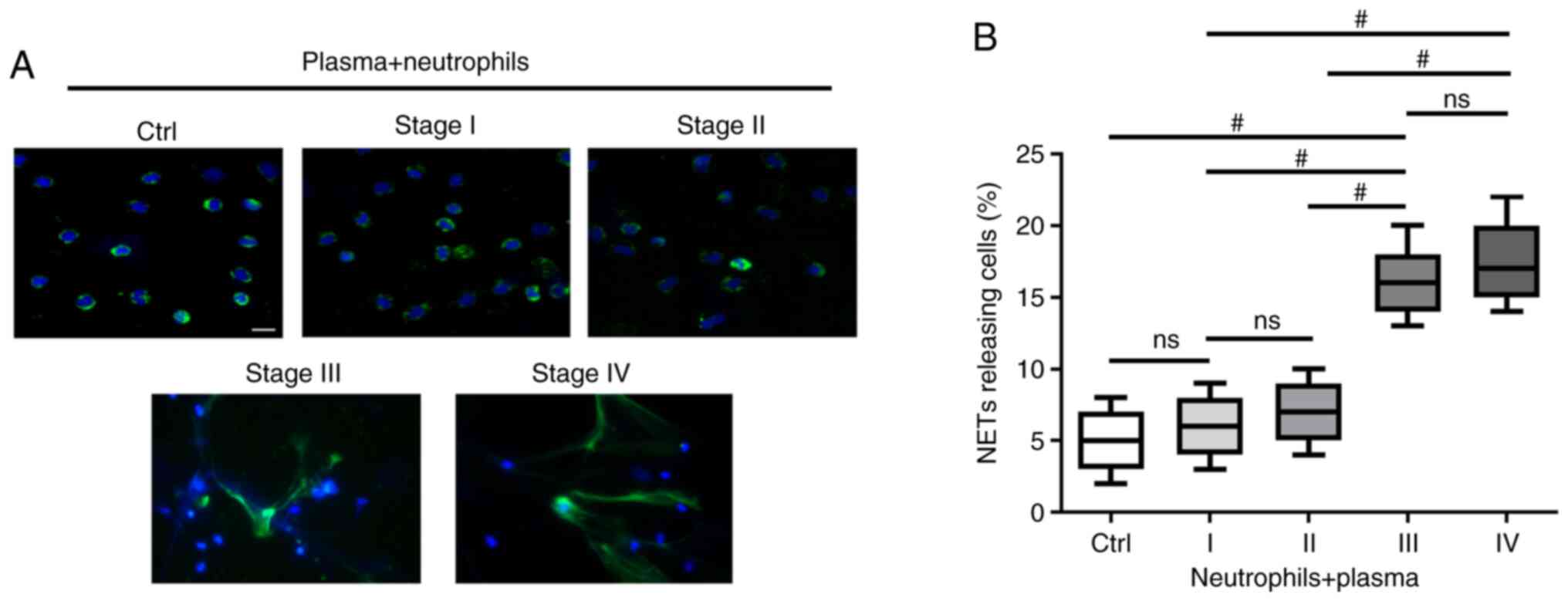
TME in patients with advanced GC primes
neutrophils to form NETs
To investigate whether the TME of GC induces
neutrophils to form NETs, the extracellular co-localization of DNA
and granule proteins were observed by staining with Hoechst 33342
and NE. NETs exhibited an extracellular fiber-like structure
protruding from neutrophils stimulated with plasma from patients
with stage III and IV GC (Fig.
2A). However, the effect was not significant when compared with
neutrophils stimulated with plasma from healthy individuals or
patients with stage I/II GC (Fig.
2A). The percentage of NETs-releasing cells following
stimulation with plasma from patients with stage III/IV GC was
higher compared with those stimulated with plasma from heathy
controls and patients with stage I/II GC (P<0.001; Fig. 2B).
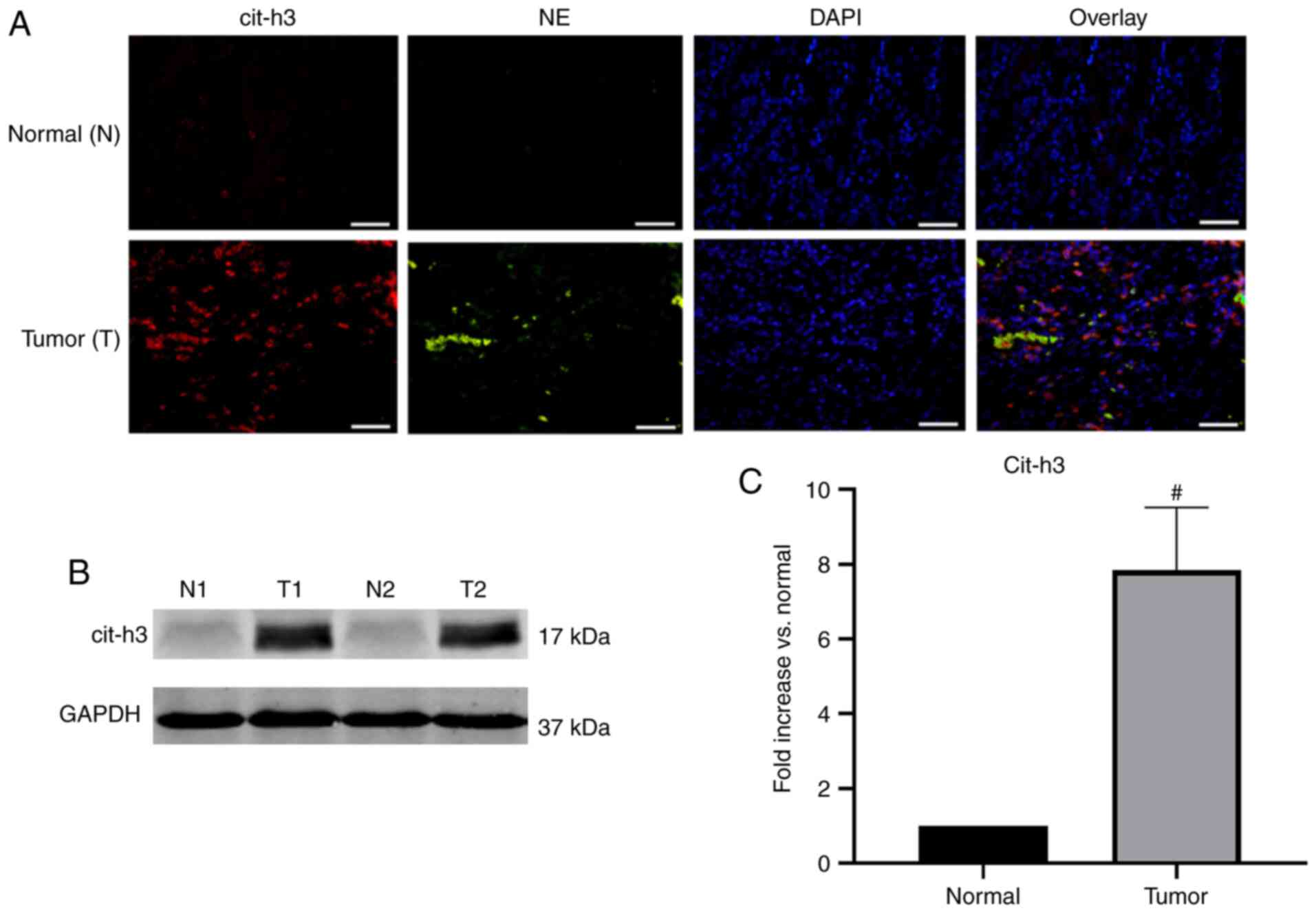
NETs deposited in human GC tissue
Surgical resection specimens of patients undergoing
D2 radical gastrectomy for GC (n=10) were collected. The
pathological type of the GC samples was adenocarcinoma. Confocal
microscopy revealed that NE and cit-h3 co-localized inside the
web-structure of NETs of GC samples, with almost no expression in
the control group (Fig. 3A).
Western blot analysis further confirmed that the expression of
cit-h3, an important biomarker of NETs, was significantly higher in
GC tissues compared with normal resection margins (P<0.001;
Fig. 3B and C). The abundant
deposition of NETs in tumor tissues was found to be consistent with
the above-mentioned immunofluorescence results.
NETs promote the migration of GC
cells
To investigate the effects of NETs on GC cells,
PMA-stimulated neutrophils were used to mimic NETs formation. NETs
formation was identified by evaluating the co-localization of
extracellular DNA and NE by immunofluorescence microscopy (Fig. 4A). The effects of NETs on cell
proliferation were assessed by measuring the viability of AGS cells
by MTT assay. It was demonstrated that the cell proliferation rate
did not differ significantly among the 3 groups (P>0.05;
Fig. 4B). Subsequently, flow
cytometric analysis revealed a similar percentage of cell cycle
arrest at the G2/M-phase in NETs-treated AGS cells compared with
the control and NETs-suppressed groups (P>0.05; Fig. 4C and D). These results indicate
that NETs did not exert any effects on cell proliferation or cell
cycle in the models investigated. Furthermore, the effects of NETs
on the migration of AGS cells were evaluated by Transwell assay. It
was demonstrated that the culture media (CM) of PMA-stimulated
neutrophils significantly improved the migratory ability of AGS
cells, and the pro-migratory effect was impaired by
degrading/inhibiting NETs (P<0.001; Fig. 4E and F).
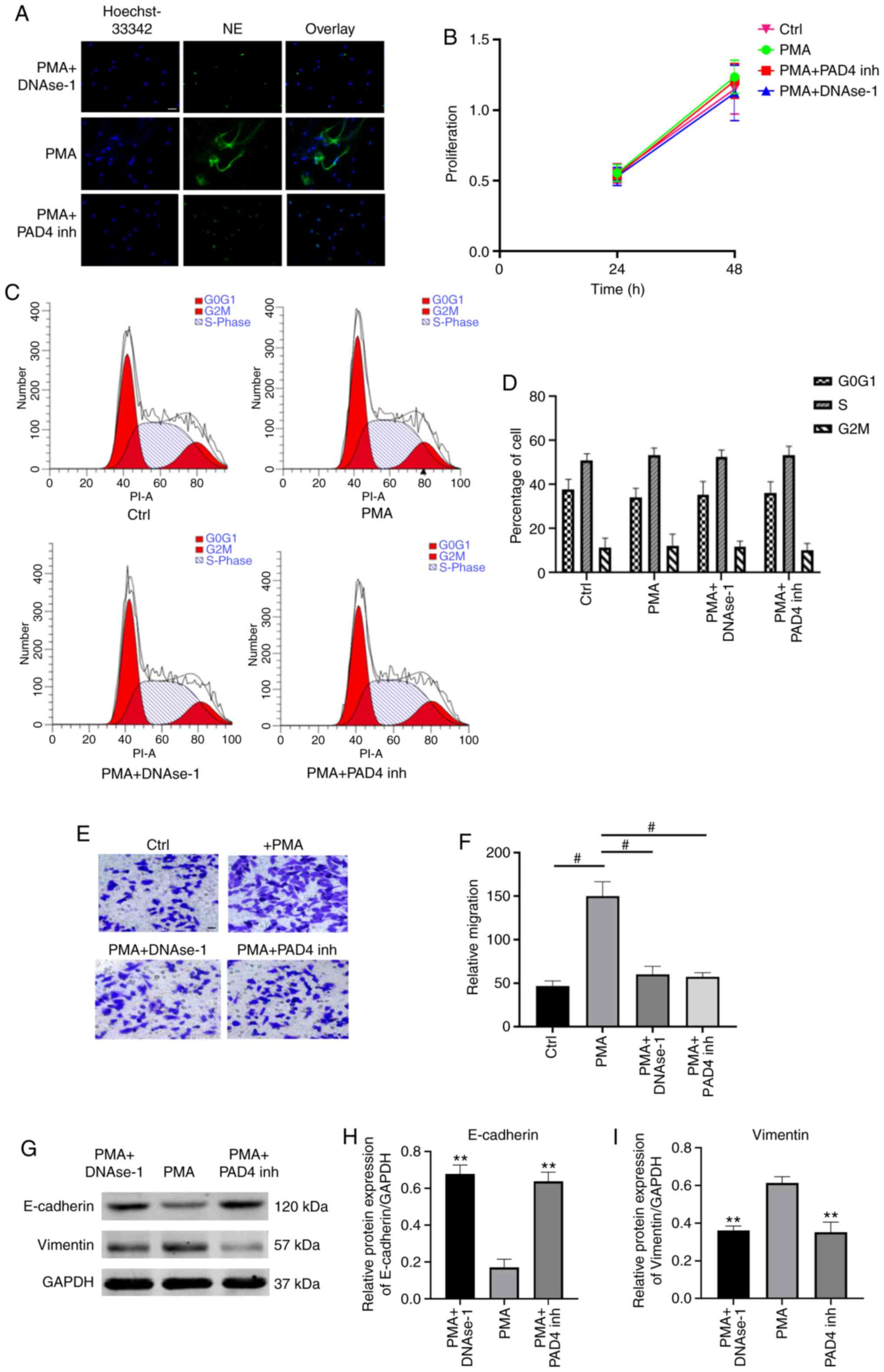 | Figure 4NETs promote tumor cell migration
without altering cell cycle and proliferation. (A) Representative
immunofluorescence microphotographs showing NETs (Hoechst-44432,
blue; NE, green). Scale bar, 50 µm; magnification, ×200. (B)
MTT assay indicated that there was no significant change in the
proliferation rate of AGS cells for 48 h in each group. Data are
presented as the means ± SEM from n=3 separate experiments. (C) The
effects of NETs on cell cycle was assessed by flow cytometry, AGS
cells were stimulated by NETs or inhibiting NET CM for 24 h. (D)
The percentage of cell population at each phase in different group.
(E) The effects of NETs on the migration ability of GC cells was
assessed by Transwell assay; scale bar, 20 µm;
magnification, ×400. (F) Statistical graphs are presented as the
means ± SEM, #P<0.001; n=5. (G) The effects of NETs
on the EMT markers in AGS cells and GAPDH were examined by western
blot analysis. (H and I) Statistical analysis of the expression of
E-cadherin and vimentin was showed. Data were normalized to the
GAPDH protein level and are expressed as the mean ± SEM,
**P<0.01 vs. PMA group; n=3. NET, neutrophil
extracellular trap; GC, gastric cancer. |
NETs promote EMT in GC cells
EMT is a critical process for aggressive metastatic
dissemination of cancer, which undergoes multiple and dynamic
transitional states from epithelial to mesenchymal phenotypes. The
present study investigated whether EMT plays a role in mediating
the pro-metastatic properties of NETs by measuring the changes in
the expression of theses biomarkers by western blot analysis.
Following treatment with NETs CM for 24 h, the AGS cells exhibited
a decreased expression of the epithelial marker, E-cadherin, and an
enhanced expression of the mesenchymal marker, vimentin. Notably,
the EMT-promoting effect of the NETs were eliminated by
DNAse-1/PAD4 inhibitor (P<0.01; Fig. 4G-I)
Effects of NETs inhibitors on tumor
progression and EMT in vivo
A post-operative recurrent tumor model was used to
evaluate the antitumor activity of DNAse-1/GSK-484 (Fig. 5A). The body weight of the nude
mice remained stable following NETs inhibition therapy (P>0.05;
Fig. 5B). By comparing the
volume and weight of tumors in each group, neither DNAse-1 nor PAD4
inhibitor significantly inhibited the growth of residual tumors
(P>0.05; Fig. 5C and D). A
significant deposition of NETs was observed in the tumor sections
of the NC group, as extracellular colocalization of NE with
citrullinated H3; however, by contrast, a lack of deposition was
observed in the DNAse-1/PAD 4 inhibitor group (Fig. 5E). To verify this observation,
the expression of cit-h3 in tumor tissues was compared in each
group and the results were consistent with the results of tissue
fluorescence analysis (P<0.01; Fig. 5G and J). In immunohistochemistry
and western blot assays, the epithelial marker, E-cadherin, was
found to be upregulated, while the mesenchymal marker, vimentin,
was downregulated by DNAse-1/GSK-484 (P<0.01; Fig. 5F-J).
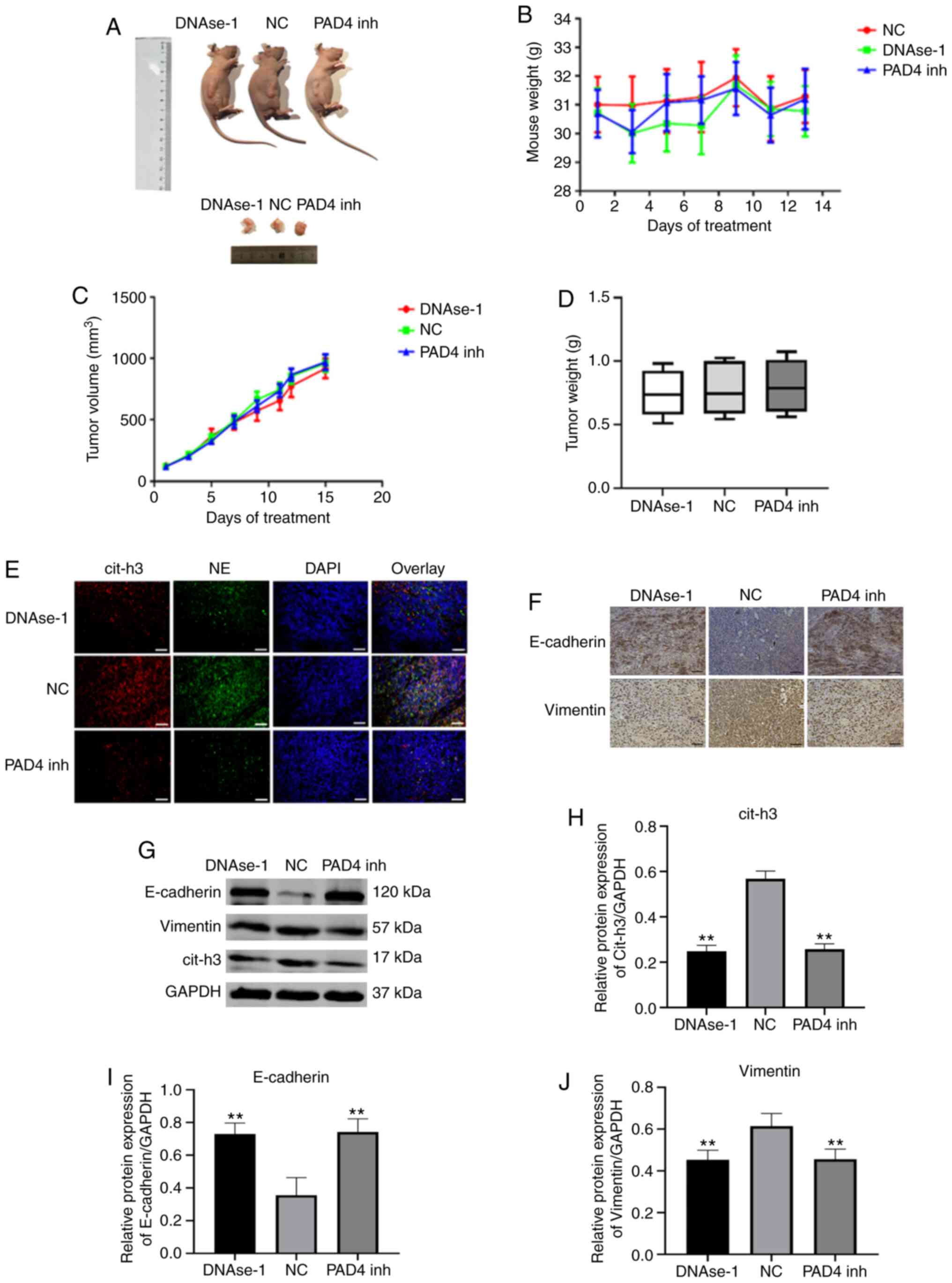 | Figure 5Effects of NET inhibitors on solid
tumor growth in nude mice. (A) Images of AGS residual tumor
xenografts treated with Dnase I or GSK-484; scale bar, 1 cm. (B-D)
The graphs of mouse weight, tumor growth curve, average tumor
weight. AGS cells induced NETs deposition in the tumors. (E) Images
showing representative immunostaining for neutrophil elastase
(green), citrullinated histone H3 (red), and DAPI (blue) in the
tumors of mice treated as indicated; scale bar, 50 µm;
magnification, ×400. (F) Representative IHC images of murine tumor
sections stained for the EMT-related markers; n=5. scale bar, 50
µm; magnification, ×400. (G) The effects of NETs inhibitors
on EMT process in murine tumor tissues, protein levels of
E-cadherin, vimentin and citrullinated histone H3 were detected by
western blot analysis. (H-J) Statistical analysis of the expression
of E-cadherin, vimentin and citrullinated histone H3. Data were
normalized to the GAPDH protein level and expressed as the means ±
SEM, **P<0.01 vs. NC group, n=5. NET, neutrophil
extracellular trap; GC, gastric cancer; cit-h3, citrullinated
histone H3. |
Discussion
In response to various stimuli, such as infections
and inflammatory cytokines, neutrophils release cloudy-like
structures that consist of their DNA-histone complexes decorated
with antimicrobial peptides, which are termed NETs (35-37). NETs have been demonstrated to
increase hypercoagulability by promoting thrombin and fibrin
generation (38). However, the
detailed mechanisms through which NETs cause tumor cell progression
and metastasis remains unclear. The present study investigated
whether NETs can promote the ability of tumor cells to migrate and
if their effect can be specifically blocked by NETs inhibitors. It
was identified that the tumor-promoting effect of NETs was closely
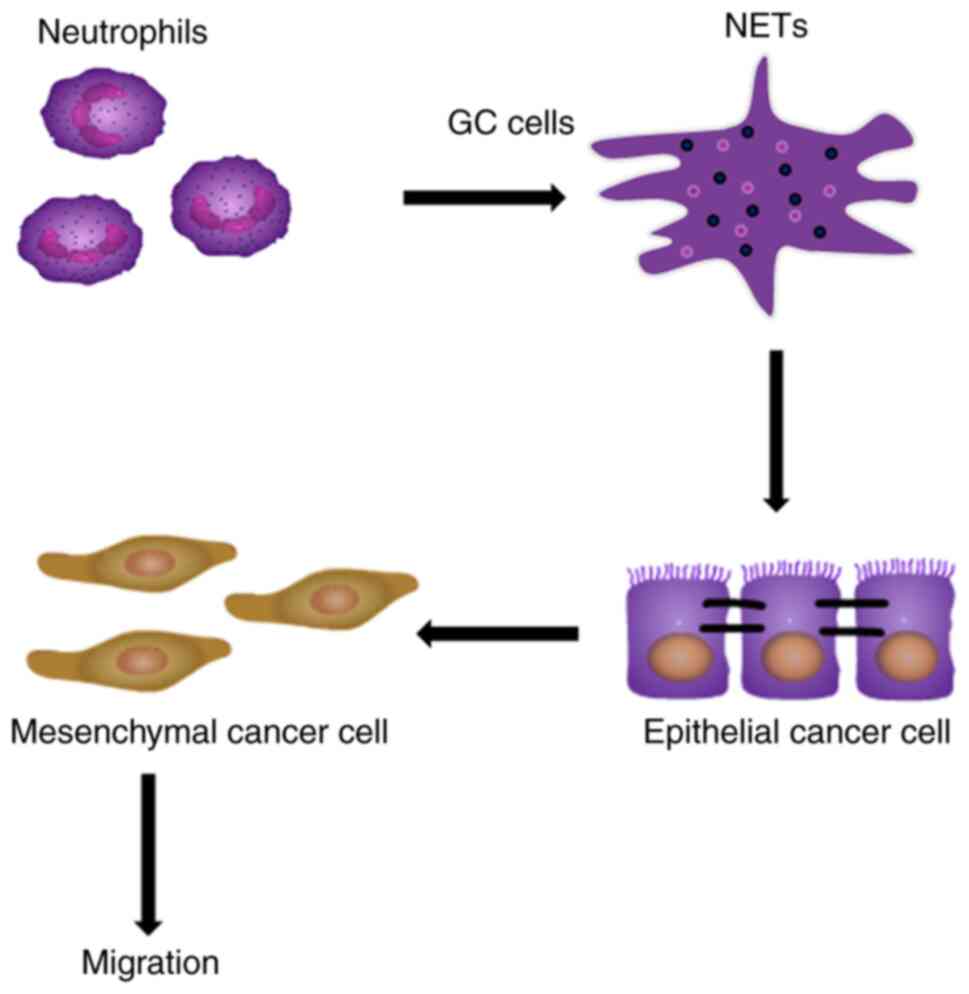
associated with EMT, a key process in GC metastasis (Fig. 6).
Circulating NETs markers are significantly elevated
in various types of cancer and are associated with a poor prognosis
of patients (39,40). It has previously been
demonstrated that NETs can lead to the formation of hypercoagulable
tumor states; however, the association between NETs and tumor
metastasis remains unclear (41). Circulating MPO-DNA complex,
cell-free DNA and NE have been proposed as NETs markers (42). The NETs levels in the peripheral
blood of patients with different TNM stages GC were detected and
analyzed retrospectively. It was identified that the formation
capacity of NETs in GC was closely associated with the progression
of the disease. It is understood that patients with advanced tumors
have a higher risk of metastasis compared with patients with
early-stage disease. The present study found that the level of NETs
in patients with stage III/IV GC was significantly higher compared
with that in healthy controls and patients with stage I/II GC,
while no significant difference was found between patients with
stage III and IV GC. Metastasis is more likely to occur in patients
with stage III/IV GC, suggesting that the level of NETs may be
associated with metastasis. Stage IV GC is notorious for distant
metastasis in patients and high mortality rates, whereas no
metastasis is present in patients with stage III disease. As
regards the hypothesis of whether NETs play a role in tumor
metastasis, the present study revealed a notable phenomenon. LVI is
considered to be an independent prognostic factor for patients with
GC (43). Consistent with the
finding of no significant differences observed in the NETs levels
in patients with stage III and IV GC, there was also no significant
difference in the incidence of LVI in patients with stage III and
IV GC; however, it was significantly higher compared with the other
groups. These results suggest that NETs may play an important role
in tumor metastasis. However, certain limitations of the present
study should be acknowledged: The clinical sample size was small
and insufficient to evaluate survival and determine the diagnostic
significance of NETs. However, due to the limited literature in
this area, it was considered important to report these results in
order to encourage further studies to evaluate the effects of NETs
on tumor progression.
Previous studies have demonstrated that the
production of NETs in tumor patients is significantly higher
compared with that in healthy controls (44,45). Microenvironmental changes in
tumor patients are more likely to stimulate the production of NETs,
and tumor cells can release inflammatory factors, such as IL-6,
IL-8 and G-CSF, to promote the release of NETs from neutrophils
(46,47). The present study revealed that
the total number of neutrophils and the percentage of neutrophils
in patients with GC did not differ significantly compared with the
healthy controls; however, the level of NETs differed between the 2
groups. The TME plays an important role in tumor progression
(48). We speculated that the
release of NETs may be associated with the TME. Therefore, first,
neutrophils were stimulated with normal plasma in the current
study; however, no NETs formation was detected. Subsequently,
neutrophils were stimulated with plasma from patients with
different TNM stages of GC, and it was identified that plasma from
patients with stage III and IV GC significantly induced the
production of NETs. This was consistent with the analysis of
peripheral blood. The results suggested that the level of NETs in
tumor patients is associated with TME, and that NETs are an
important part of the TME. Therefore, NETs may be considered as
prognostic markers in patients.
Neutrophils and NETs are highly infiltrated in
tumors (49). In a study on
breast cancer, a high expression of NETs was identified in high
metastatic triple-negative breast cancer tissues, with a lower
expression in HER-2 type breast cancer, and almost no NETs were
observed in luminal breast cancer. It was also revealed that this
differential expression of NETs was positively associated with the
degree of metastasis and closely associated with the prognosis of
patients (13). The present
study first reported the high expression of NETs in GC tissues.
Subsequently, a comparison was made between GC tissues and the
normal resection margins. It was identified that the expression of
NETs in tumor tissues was significantly increased, while NETs could
hardly be detected in normal tissues. A large extent of NETs
deposition in tumors may serve as a promoter of tumor progression
and metastasis. NETs, whose main components are cell-free DNA and
histones, have been reported to induce tumor metastasis by
capturing circulating free tumor cells (15). In addition to promoting
metastasis, NETs are also considered to play an antitumor role in
certain tumors, and have been reported to inhibit the proliferation
and promote the apoptosis of colon cancer cells (19). The antitumor effect of NETs was
also reported in a study on melanoma (50). It was hypothesized that NETs play
a dual role in tumor progression. On the one hand, NETs can inhibit
tumor progression as a continuation of initial inflammatory
response. On the other hand, NETs may play an adverse role in tumor
progression offering a scaffold with many molecules with biological
activity, which may promote malignant cells progression and
migration (15). When the
melanoma cell line A375 was stimulated with NETs, their ability to
metastasize and proliferate declined. However, this antitumor
effect can be reversed by DNase I (20). Moreover, the web-like structure
promotes the adhesion of melanoma cells similar to the mechanism
for capturing microbes (36). In
addition, NETs were found to inhibit cancer cell growth by inducing
apoptosis of Caco-2 and AML cells (19). However, current studies on the
antitumor effects of NETs lack in-depth studies on the mechanisms,
which are presented in terms of phenotypes. Therefore, the present
study investigated the effects of NETs on GC cells. By stimulating
tumor cells with NETs-containing culture medium or a NETs inhibitor
to observe the effects of NETs on cell migration, it was identified
that the specific tumor-promoting effect of NETs were blocked by
NET inhibitor. DNase-1 is a powerful inhibitor that degrades NETs
by degrading the DNA structure. PAD4 is a key enzyme involved in
the release of NETs into the extracellular domain, and inhibition
of PAD4 has been reported to inhibit chromatin depolymerization and
thereby inhibit the the release of NETs into the extracellular
domain. The present study confirmed through in vitro
experiments that both inhibitors can effectively inhibit the
formation of NETs and the migration of tumor cells, to play a
specific antitumor role. However, the limitations of NETs on other
aspects of tumor cells need to be further verified. The current
results suggested that NETs inhibitors may have some practical
application value in the treatment and prevention of tumor
metastasis.
EMT is a prerequisite and important process for the
metastasis of GC (51).
Following the loss of the epithelial phenotype and the gain of
mesenchymal features, tumor cells acquire a potent ability to
migrate and invade which then leads to distant metastasis.
Neutrophils have been reported to induce EMT in tumor cells. When
tumor cells were stimulated by neutrophils, the expression of
classical EMT markers, such as slug and vimentin, were upregulated
(52). Hu et al (53) reported that the number of
infiltrated neutrophils in tumor cells was negatively associated
with the expression of the epithelial marker E-cadherin in lung
cancer tissues. The present study found a large number of NETs in
patients with advanced GC with high risk of metastasis. Stimulation
of GC cells with NETs-containing medium inhibited the expression of
epithelial marker, E-cadherin, enhanced the expression of the
stromal marker, vimentin, and promoted the migration of GC cells.
This pro-EMT effect was attenuated by the NETs inhibitors, and this
specific effect may provide novel strategies for the treatment of
tumor metastasis. The occurrence of neutrophil-induced EMT is
associated with numerous signaling pathways, such as the AKT
signaling pathway, the TGF-β/Smad signaling pathway and the ERK
signaling pathway (53-55). Whether NETs can be activated via
relevant signaling pathways to promote EMT in GC cells remains to
be further verified.
In addition, the NETs promoted cell migration
through EMT is probability mediated by NETs-associated proteases.
The NETs DNA bound to the extracellular matrix (ECM), thus bringing
NETs associated proteases to their substrate (27). The NETs-mediated proteolytic
remodeling of ECM may cause many epitope exposures that affected
proliferation or cell cycle change of cancer cells through various
signaling pathway, such as integrin and Ras/Raf/MAPK signaling
(27,56). In addition, STAT1/STAT3/STAT5 and
cyclin-dependent-kinase (CDKs) are both involved in the regulation
of cell cycle and cell proliferation (57,58). Whether NETs induce cell
proliferation and cell cycle changes in these ways remains to be
tested. It may be a potent strategy to prevent cancer recurrence
and broadly serve as treatment for other NETs associated
pathological processes, such as EMT. If such an association exists,
it may be possible that NETs and their downstream effectors could
be targeted to reduce the risk of tumor metastasis. The mechanisms
of the effects of NETs on tumor progression will be the focus of
our future research.
In conclusion, the present study revealed a novel
pro-tumor activity of neutrophils mediated by NETs, which may
explain the poor prognosis of patients with GC associated with
neutrophil aggregation within the tumor environment. As presented
in Fig. 6, GC primes neutrophils
towards NETosis, then NETs deposit in GC tissues or adhere to
epithelial cancer cells, notably without affecting cell
proliferation and cell cycle, and NETs promote cancer cell
migration via EMT in vitro and in vivo.
The number of NETs in peripheral blood was increased
significantly in patients with advanced GC and was positively
associated with the increased risk of tumor staging and metastasis.
NETs are expected to become a novel tumor biomarker and provide a
novel therapeutic target for the comprehensive treatment of GC. In
the future, NETs inhibitors may become a promising drug which may
be used to prevent tumor recurrence.
Availability of data and materials
The datasets generated and analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
TZ designed the study, performed some experiments,
analyzed the data, prepared the figures and wrote the manuscript.
XZ obtained funding, designed the study, performed some experiments
and revised the manuscript. LL, BW, HL and ZX conducted the mouse
experiments, cell culture and cell treatments. CY, RL, DH and QW
performed some experiments and analyzed some of the results. All
authors were substantially involved in the research, acquisition of
the data, analysis and manuscript preparation. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All procedures performed in studies involving human
participants were in accordance with the standards upheld by the
Ethics Committee of the Second Affiliated Hospital of Harbin
Medical University and with those of the 1964 Helsinki Declaration
and its later amendments for ethical research involving human
subjects. All subjects manually signed the informed consent. All
animal experiments were performed in accordance with the NIH
guidelines (Guide for the Care and Use of Laboratory Animals) and
approved by the Ethics Committee of the Second Affiliated Hospital
of Harbin Medical University, P.R. China. The Ethics Committee of
the Second Affiliated Hospital of Harbin Medical University
approved the present study (permit no. KY2016-032).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Professor Chu
Wenfeng of Harbin Medical University for providing guidance on the
experimental techniques and for the provision of the experimental
instruments.
Funding
The present study was supported by the National Science
Foundation of China (grant no. 81672355).
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Arneth B: Tumor microenvironment. Medicina
(Kaunas). 56:152019. View Article : Google Scholar
|
|
4
|
Quail DF and Joyce JA: Microenvironmental
regulation of tumor progression and metastasis. Nat Med.
19:1423–1437. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gonzalez H, Hagerling C and Werb Z: Roles
of the immune system in cancer: From tumor initiation to metastatic
progression. Genes Dev. 32:1267–1284. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang X, Shi H, Yuan X, Jiang P, Qian H
and Xu W: Tumor-derived exosomes induce N2 polarization of
neutrophils to promote gastric cancer cell migration. Mol Cancer.
17:1462018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shaul ME and Fridlender ZG:
Tumour-associated neutrophils in patients with cancer. Nat Rev Clin
Oncol. 16:601–620. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Coffelt SB, Wellenstein MD and de Visser
KE: Neutrophils in cancer: Neutral no more. Nat Rev Cancer.
16:4314462016. View Article : Google Scholar
|
|
9
|
Liu Y and Liu L: The pro-tumor effect and
the anti-tumor effect of neutrophils extracellular traps. Biosci
Trends. 13:469–475. 2020. View Article : Google Scholar
|
|
10
|
Demers M, Krause DS, Schatzberg D,
Martinod K, Voorhees JR, Fuchs TA, Scadden DT and Wagner DD:
Cancers predispose neutrophils to release extracellular DNA traps
that contribute to cancer-associated thrombosis. Proc Natl Acad Sci
USA. 109:13076–13081. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cedervall J, Hamidi A and Olsson AK:
Platelets, NETs and cancer. Thromb Res. 164(Suppl 1): S148–S152.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Berger-Achituv S, Brinkmann V, Abed UA,
Kühn LI, Ben-Ezra J, Elhasid R and Zychlinsky A: A proposed role
for neutrophil extracellular traps in cancer immunoediting. Front
Immunol. 4:482013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Park J, Wysocki RW, Amoozgar Z, Maiorino
L, Fein MR, Jorns J, Schott AF, Kinugasa-Katayama Y, Lee Y, Won NH,
et al: Cancer cells induce metastasis-supporting neutrophil
extracellular DNA traps. Sci Transl Med. 8:361ra1382016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Najmeh S, Cools-Lartigue J, Rayes RF,
Gowing S, Vourtzoumis P, Bourdeau F, Giannias B, Berube J, Rousseau
S, Ferri LE and Spicer JD: Neutrophil extracellular traps sequester
circulating tumor cells via β1-integrin mediated interactions. Int
J Cancer. 140:2321–2330. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cools-Lartigue J, Spicer J, McDonald B,
Gowing S, Chow S, Giannias B, Bourdeau F, Kubes P and Ferri L:
Neutrophil extracellular traps sequester circulating tumor cells
and promote metastasis. J Clin Invest. 123:3446–3458. 2013.
View Article : Google Scholar :
|
|
16
|
Grilz E, Mauracher LM, Posch F,
Königsbrügge O, Zöchbauer-Müller S, Marosi C, Lang I, Pabinger I
and Ay C: Citrullinated histone H3, a biomarker for neutrophil
extracellular trap formation, predicts the risk of mortality in
patients with cancer. Br J Haematol. 186:311–320. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Erpenbeck L and Schön MP: Neutrophil
extracellular traps: Protagonists of cancer progression? Oncogene.
36:2483–2490. 2017. View Article : Google Scholar
|
|
18
|
Claushuis TA, de Stoppelaar SF, Stroo I,
Roelofs JJ, Ottenhoff R, van der Poll T and Van't Veer C: Thrombin
contributes to protective immunity in pneumonia-derived sepsis via
fibrin polymerization and platelet-neutrophil interactions. J
Thromb Haemost. 15:744–757. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Arelaki S, Arampatzioglou A, Kambas K,
Papagoras C, Miltiades P, Angelidou I, Mitsios A, Kotsianidis I,
Skendros P, Sivridis E, et al: Gradient infiltration of neutrophil
extracellular traps in colon cancer and evidence for their
involvement in tumour growth. PLoS One. 11:e01544842016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schedel F, Mayer-Hain S, Pappelbaum KI,
Metze D, Stock M, Goerge T, Loser K, Sunderkötter C, Luger TA and
Weishaupt C: Evidence and impact of neutrophil extracellular traps
in malignant melanoma. Pigment Cell Melanoma Res. 33:63–73. 2020.
View Article : Google Scholar
|
|
21
|
Gupta GP and Massagué J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pastushenko I and Blanpain C: EMT
transition states during tumor progression and metastasis. Trends
Cell Biol. 29:212–226. 2019. View Article : Google Scholar
|
|
23
|
Zhang W, Gu J, Chen J, Zhang P, Ji R, Qian
H, Xu W and Zhang X: Interaction with neutrophils promotes gastric
cancer cell migration and invasion by inducing
epithelial-mesenchymal transition. Oncol Rep. 38:2959–2966. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li S, Cong X, Gao H, Lan X, Li Z, Wang W,
Song S, Wang Y, Li C, Zhang H, et al: Correction to:
Tumor-associated neutrophils induce EMT by IL-17a to promote
migration and invasion in gastric cancer cells. J Exp Clin Cancer
Res. 38:1772019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rayes RF, Vourtzoumis P, Bou Rjeily M,
Seth R, Bourdeau F, Giannias B, Berube J, Huang YH, Rousseau S,
Camilleri-Broet S, et al: Neutrophil extracellular trap-associated
CEACAM1 as a putative therapeutic target to prevent metastatic
progression of colon carcinoma. J Immunol. 204:2285–2294. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
American Joint Committee on Cancer (AJCC):
Cancer Staging Manual. AJCC; Chicago, IL: 2016, https://cancerstaging.org/references-tools/deskreferences/pages/default.aspx.
|
|
27
|
Albrengues J, Shields MA, Ng D, Park CG,
Ambrico A, Poindexter ME, Upadhyay P, Uyeminami DL, Pommier A,
Küttner V, et al: Neutrophil extracellular traps produced during
inflammation awaken dormant cancer cells in mice. Science.
361:eaao42272018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li B, Liu Y, Hu T, Zhang Y, Zhang C, Li T,
Wang C, Dong Z, Novakovic VA, Hu T and Shi J: Neutrophil
extracellular traps enhance procoagulant activity in patients with
oral squamous cell carcinoma. J Cancer Res Clin Oncol.
145:1695–1707. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu Y, Li H, Yan R, Wang C, Wang Y, Zhang
C, Liu M, Zhou T, Zhu W, Zhang H, et al: Increased neutrophil
activation and plasma DNA levels in patients with pre-eclampsia.
Thromb Haemost. 118:2064–2073. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schimmel M, Nur E, Biemond BJ, van Mierlo
GJ, Solati S, Brandjes DP, Otten HM, Schnog JJ, Zeerleder S and
Curama Study Group: Nucleosomes and neutrophil activation in sickle
cell disease painful crisis. Haematologica. 98:1797–1803. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yoo DG, Floyd M, Winn M, Moskowitz SM and
Rada B: NET formation induced by Pseudomonas aeruginosa cystic
fibrosis isolates measured as release of myeloperoxidase-DNA and
neutrophil elastase-DNA complexes. Immunol Lett. 160:186–194. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stakos DA, Kambas K, Konstantinidis T,
Mitroulis I, Apostolidou E, Arelaki S, Tsironidou V, Giatromanolaki
A, Skendros P, Konstantinides S and Ritis K: Expression of
functional tissue factor by neutrophil extracellular traps in
culprit artery of acute myocardial infarction. Eur Heart J.
36:1405–1414. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Abed AU and Brinkmann V:
Immunofluorescence labelling of human and murine neutrophil
extracellular traps in paraffin-embedded tissue. J Vis Exp.
151:e601152019.
|
|
34
|
Owen GR, Häkkinen L, Wu C and Larjava H: A
reproducible technique for specific labeling of antigens using
preformed fluorescent molecular IgG-F(ab′)2 complexes from primary
anti-bodies of the same species. Microsc Res Tech. 73:623–630.
2010.
|
|
35
|
Hu Z, Murakami T, Tamura H, Reich J,
Kuwahara-Arai K, Iba T, Tabe Y and Nagaoka I: Neutrophil
extracellular traps induce IL-1β production by macrophages in
combination with lipopolysaccharide. Int J Mol Med. 39:549–558.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Brinkmann V, Reichard U, Goosmann C,
Fauler B, Uhlemann Y, Weiss DS, Weinrauch Y and Zychlinsky A:
Neutrophil extracellular traps kill bacteria. Science.
303:1532–1535. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nauseef WM and Borregaard N: Neutrophils
at work. Nat Immunol. 15:602–611. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang C, Sun W, Cui W, Li X, Yao J, Jia X,
Li C, Wu H, Hu Z and Zou X: Procoagulant role of neutrophil
extracellular traps in patients with gastric cancer. Int J Clin Exp
Pathol. 8:14075–14086. 2015.
|
|
39
|
Marin Oyarzún CP, Carestia A, Lev PR,
Glembotsky AC, Castro Ríos MA, Moiraghi B, Molinas FC, Marta RF,
Schattner M and Heller PG: Neutrophil extracellular trap formation
and circulating nucleosomes in patients with chronic
myeloproliferative neoplasms. Sci Rep. 6:387382016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Oklu R, Sheth RA, Wong KHK, Jahromi AH and
Albadawi H: Neutrophil extracellular traps are increased in cancer
patients but does not associate with venous thrombosis. Cardiovasc
Diagn Ther. 7(Suppl 3): S140–S149. 2017. View Article : Google Scholar
|
|
41
|
Guglietta S and Rescigno M:
Hypercoagulation and complement: Connected players in tumor
development and metastases. Semin Immunol. 28:578–586. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Brinkmann V: Neutrophil extracellular
traps in the second decade. J Innate Immun. 10:414–421. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wu L, Liang Y, Zhang C, Wang X, Ding X,
Huang C and Liang H: Prognostic significance of lymphovascular
infiltration in overall survival of gastric cancer patients after
surgery with curative intent. Chin J Cancer Res. 31:785–796. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Abdol Razak N, Elaskalani O and Metharom
P: Pancreatic cancer-induced neutrophil extracellular traps: A
potential contributor to cancer-associated thrombosis. Int J Mol
Sci. 18:4872017. View Article : Google Scholar :
|
|
45
|
Richardson JJR, Hendrickse C, Gao-Smith F
and Thickett DR: Neutrophil extracellular trap production in
patients with colorectal cancer in vitro. Int J Inflam.
2017:49150622017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang Y, Wang C, Yu M, Zhao X, Du J, Li Y,
Jing H, Dong Z, Kou J, Bi Y, et al: Neutrophil extracellular traps
induced by activated platelets contribute to procoagulant activity
in patients with colorectal cancer. Thromb Res. 180:87–97. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Arpinati L, Shaul ME, Kaisar-Iluz N, Mali
S, Mahroum S and Fridlender ZG: NETosis in cancer: A critical
analysis of the impact of cancer on neutrophil extracellular trap
(NET) release in lung cancer patients vs. mice Cancer Immunol
Immunother. 69:199–213. 2020. View Article : Google Scholar
|
|
48
|
Wu L, Saxena S and Singh RK: Neutrophils
in the tumor microenvironment. Adv Exp Med Biol. 1224:1–20. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hisada Y, Grover SP, Maqsood A, Houston R,
Ay C, Noubouossie DF, Cooley BC, Wallén H, Key NS, Thålin C, et al:
Neutrophils and neutrophil extracellular traps enhance venous
thrombosis in mice bearing human pancreatic tumors. Haematologica.
105:218–225. 2020. View Article : Google Scholar :
|
|
50
|
Andzinski L, Kasnitz N, Stahnke S, Wu CF,
Gereke M, von Köckritz-Blickwede M, Schilling B, Brandau S, Weiss S
and Jablonska J: Type I IFNs induce anti-tumor polarization of
tumor associated neutrophils in mice and human. Int J Cancer.
138:1982–1993. 2016. View Article : Google Scholar
|
|
51
|
Natalwala A, Spychal R and Tselepis C:
Epithelial-mesenchymal transition mediated tumourigenesis in the
gastrointestinal tract. World J Gastroenterol. 14:3792–3797. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang Y, Chen J, Yang L, Li J, Wu W, Huang
M, Lin L and Su S: Tumor-contacted neutrophils promote metastasis
by a CD90-TIMP-1 Juxtacrine-paracrine loop. Clin Cancer Res.
25:1957–1969. 2019. View Article : Google Scholar
|
|
53
|
Hu P, Shen M, Zhang P, Zheng C, Pang Z,
Zhu L and Du J: Intratumoral neutrophil granulocytes contribute to
epithelial-mesenchymal transition in lung adenocarcinoma cells.
Tumour Biol. 36:7789–7796. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wu Y, Zhao Q, Peng C, Sun L, Li XF and
Kuang DM: Neutrophils promote motility of cancer cells via a
hyaluronan-mediatedTLR4/PI3K activation loop. J Pathol.
225:438–447. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhou Q, Wang X, Yu Z, Wu X, Chen X, Li J,
Zhu Z, Liu B and Su L: Transducin (β)-like 1 X-linked receptor 1
promotes gastric cancer progression via the ERK1/2 pathway.
Oncogene. 36:1873–1886. 2017. View Article : Google Scholar
|
|
56
|
Molina JR and Adjei AA: The Ras/Raf/MAPK
pathway. J Thorac Oncol. 1:7–9. 2006. View Article : Google Scholar
|
|
57
|
Groner B and von Manstein V: Jak Stat
signaling and cancer: Opportunities, benefits and side effects of
targeted inhibition. Mol Cell Endocrinol. 451:1–14. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lim S and Kaldis P: Cdks, cyclins and
CKIs: Roles beyond cell cycle regulation. Development.
140:3079–3093. 2013. View Article : Google Scholar : PubMed/NCBI
|