Introduction
Primary open angle glaucoma (POAG) leads to
irreversible blindness (1),
which is characterized by the gradual apoptosis of retinal ganglion
cells, resulting in visual field defects (2). Intraocular pressure (IOP) elevation
is the major risk factor for the progression of POAG (3). However, the underlying pathogenesis
of POAG remains inconclusive. Considering that ~75% of aqueous
humor outflow occurs through the trabecular meshwork (TM) pathway
(4), increased IOP-associated
POAG is considered to be mainly due to elevated resistance to the
TM outflow (5).
Most of the increased resistance to aqueous humor
outflow is due to an increase in extracellular matrix (ECM)
deposition (6-8). ECM contains elastin, collagens and
fibronectin (FN). Considering the involvement of the ECM in
increasing the resistance of the TM to aqueous humor outflow,
pathological dysregulation of the ECM is strongly implicated in
POAG, in part mediated via the matricellular proteins, which are
important regulatory factors-extracellular proteins, including
tenascin-C, secreted protein and rich in cysteine (SPARC) and
connective tissue growth factor (CTGF), and communicate with each
other and regulate their surrounding ECM (9,10). CTGF and SPARC may promote tissue
fibrosis and abnormal tissue remodelling and disrupt ECM
homeostasis, which eventually contributes to the pathogenesis of
glaucoma (11,12). In addition, aging, actomyosin
contractile systems (e.g. ciliary muscle contractions) and
inflammation also contribute to the TM resistance to outflow
(13,14)
FYVE, RhoGEF and PH domain containing 4 (FGD4) is a
guanine nucleotide exchange factor specific for activation of cell
division control protein 42 homolog (CDC42) (GTP bound) (15). The FGD4/CDC42 pathway is involved
in the regulation of cell morphology, cytoskeleton organization,
intercellular adhesion and ECM synthesis of non-ocular cells
(16-18). Thus, the FGD4/CDC42 pathway may
also contribute to the regulation of cell morphology, cytoskeletal
organization, cell adhesion and ECM synthesis in human trabecular
meshwork (HTM) cells.
There are several therapeutic agents available for
reducing IOP in glaucoma (19).
However, few of them are designed specifically for controlling the
pathogenesis of POAG, e.g. targeting ECM accumulation in the TM.
Moreover, the currently available anti-glaucoma agents can only
decrease IOP, but not prevent the progression of, nor cure POAG
(20,21).
Statins reduce serum cholesterol via selectively
inhibiting HMG-CoA reductase for the biosynthesis of cholesterol
(22-24). Statins are well known for their
'pleiotropic effects', e.g., anti-fibrotic, anti-inflammatory and
immunomodulatory (25-29). Interestingly, statins also reduce
the progression of POAG in specific populations (30-32), suggesting that statins minimize
the risk of the development of glaucoma, as well as delay the
progression of visual field defect (32). Because statins suppress ECM
synthesis in non-ocular tissues, we hypothesized that statins can
also decrease ECM dysregulation in human trabecular meshwork (HTM),
constituting a potential anti-glaucoma mechanism (33,34). We demonstrated previously that
atorvastatin improved aqueous humor outflow in a porcine whole-eye
perfusion model ex vivo (35), but it is unclear whether
atorvastatin is able to reduce IOP in POAG in vivo.
Transforming growth factor (TGF)-β2, expressed in TM
cells, contributes to ECM generation (36). An adenoviral TGF-β2-induced
ocular hypertension murine model was found to display a high
glaucoma risk via decreasing aqueous outflow facility (36,37). Such an animal model is expected
to be a useful model to investigate the pathogenesis of glaucoma
(38).
Our present study aimed to investigate whether
atorvastatin was able to reduce TGF-β2-induced intraocular
hypertension in vivo, as well as to investigate the possible
molecular expression in the dysregulation of the ECM in primary HTM
cells following atorvastatin administration in vitro.
Materials and methods
Animals
All animals used in this research followed the ARVO
statement for the Use of Animals (https://www.arvo.org/About/policies/statement-for-the-use-of-animals-in-ophthalmic-and-visual-research/).
BALB/cJ mice (8 weeks; 3 females and 3 males per group, 4 groups
and 24 mice in this study; 22~25 g), purchased from the Lingchang
Animal Facility (Shanghai, China), were offered food and water
ad libitum on a 12 h light/12 h dark cycle at 20°C room
temperature. At the end of the experiment the animals were
euthanized by cervical dislocation. The study was conducted
according to the guidelines of the Declaration of Helsinki and
approved by the Institutional Animal Care and Use Committee of the
Huashan Hospital, Fudan University (Protocol #: JS-194, Approval
Date: 19 February 2019).
Induction of ocular hypertension in
vivo
The ocular hypertension model was induced by
injecting an adenoviral vector of human TGF-β2
(Ad.hTGF-β2226/228) in 2 µl (6×107
pfu) intravitreally in the right eye, as described previously
(36). The left eye was given
carrier Ad with an empty vector (6×107 pfu in 2
µl), as a control. There was also a blank, no-injection
control group. The Ad.hTGF-β2226/228-injected animals
were further divided into groups with and without atorvastatin
administration (20 mg/kg/day, gavage), starting at day 15. Thus,
there were totally four groups of experimental mice:
Ad.hTGF-β2226/228-induced (n=6);
Ad.hTGF-β2226/228-induced with atorvastatin from day 15
(n=6); Ad. empty-injected (n=6); and non-injection (n=6). All IOP
measurements were implemented using a Tonolab rebound tonometer
(Icare), as described (39).
Trabecular meshwork cell culture
All experiments were carried out according to the
guidelines of the Human Ethics Committee of Huashan Hospital, Fudan
University (protocol #: 2020-611, approval date: 31 March 2020)
(40). Primary HTM cells were
isolated and characterized as described previously (41). There were 4 donors from Huashan
Hospital, a Red Cross Hospital (Shanghai, China), aged 30, 35, 48
and 50 years, including 2 females and 2 males, respectively. Formal
written consent was obtained from each of the donors in this study.
All the donors had no previous diagnosis or history of eye disease.
A more detailed description of the procedures for generating
primary HTM cells were as follows. Human TM tissues were explanted
from the corneal edge and were further minced in culture solution.
The minced tissues were further cultured for 4 h at 37°C under 5%
CO2 until the cells were adherent to the bottom of the
culture flask. The culture flashes were washed with fresh culture
medium to remove dead unadhered cells. The culture medium consisted
of DMEM containing 15% fetal bovine serum (FBS) (Gibco; Thermo
Fisher Scientific, Inc.). HTM cells were sub-cultured when the
cells reached 80% confluence. The HTM cells were passaged at a
ratio of 1:3 to maintain the same conditions. The third passage of
HTM cells was identified and validated as HTM cells
immunohistochemically for the specific proteins FN, vimentin and
laminin (LN). The cultured HTM cells were stimulated with 250 and
500 nM dexamethasone (DEX) (Sigma-Aldrich; Merck KGaA) with fresh
medium every 2 days until the 6th day. The level of myocilin
(MYOC), which correlates with the trabecular meshwork
glucocorticoid response, was assessed with western blot analysis.
These HTM cells were passaged for 5 cultures in this study.
Atorvastatin treatment
The HTM cells were serum-deprived for 12 h until
they reached 90% confluency. These cells were subsequently treated
with atorvastatin (Sigma-Aldrich; Merck KGaA) at different dosages
(0, 50, 100, 150 and 200 µM), and for different times (12,
24, 36 and 48 h). The morphological changes in the HTM cells were
captured under phase contrast microscopy (ECLIPSE Ni-U; Nikon). The
effect of atorvastatin on the viability of HTM cells was determined
using a CCK-8 assay kit (Dojindo Molecular Technologies, Inc.)
following atorvastatin treatment (0, 50, 100, 150, 200 µM),
as previously described (35).
Real-time PCR
Whole cell RNA from HTM cells was extracted, using
Trizol reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
Prime-Script RT reagent kit (RR036A; Takara Biotechnology Co.) was
used for cDNA synthesis from 1 µg RNA. Real-time PCR was
conducted using a qPCR kit (RR820A; Takara Biotechnology Co.), and
the primers used are shown in Table
I. The procedures have been described in detail previously
(35).
 | Table ISequences of the specific
primers. |
Table I
Sequences of the specific
primers.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| Collagen
I |
ACGTCCTGGTGAAGTTGGT |
CAGGGAAGCCTCTCTCTCCT |
| FN |
AATATCTCGGTGCCATTTGC | AAAGGCAT
GAAGCACTCAA |
| SPARC |
ATGACGACGGCACCTACAG |
TCGCGTTGGGGTAACTTTTCA |
| CTGF |
GCAGGCTAGAGAAGCAGAGC |
ATGTCTTCATGCTGGTGCAG |
| GAPDH |
ACAGTCAGCCGCATCTTC |
CTCCGACCTTCACCTTCC |
Western blot analysis
Western blot analysis was performed, as described
previously (35,42). The concentration of each protein
sample was determined using the Bradford protein-detection method
(Bio-Rad, Laboratories, Inc.) and separated by SDS-polyacrylamide
gel electrophoresis (PAGE) (10% acrylamide), then transferred onto
a polyvinylidene fluoride (PVDF) membrane. The blots were incubated
with the primary antibodies: rabbit anti-collagen I antibody
(14695-1-AP, 1:1,000 dilution; Proteintech), rabbit
anti-fibronectin antibody (15613-1-AP, 1:1,500 dilution;
Proteintech), rabbit anti-CTGF antibody (ab6992, 1:1,000 dilution;
Abcam), rabbit anti-SPARC antibody (15274-1-AP, 1:1,000 dilution,
Proteintech), GAPDH (60004-1-Ig, 1:500 dilution, Proteintech) in
blocking solution overnight at 4°C. Then the membranes were
incubated with goat anti-rabbit secondary antibody (SE134, 1:1,000
dilution, Solarbio) at room temperature for 1 h, followed by
washing with TBST for 3 times. Then, typically enhanced
chemiluminescent (ECL) kit was used according to the manufacturer's
instructions (POO18FS, Beyotime). A particular band of each sample
was visualized by the Odyssey infrared imaging system (LI-COR,
Inc.). The bands were analyzed using ImageJ software and normalized
to each GAPDH band (version 1.48v; National Institutes of Health).
The protein density of every band was analyzed with Image J
software (http://imagej.nih.gov/ij/).
Immunofluorescence
Glass coverslips seeded with HTM cells were fixed
with 4% paraformaldehyde. After 3 times washing with PBS, the
primary antibody was bound at 4°C overnight: rabbit anti-collagen I
antibody (14695-1-AP, 1:1,000 dilution; Proteintech), rabbit
anti-fibronectin antibody (15613-1-AP, 1:1,500 dilution;
Proteintech), rabbit anti-CTGF antibody (ab6992, 1:1,000 dilution;
Abcam), rabbit anti-SPARC antibody (15274-1-AP, 1:1,000 dilution;
Proteintech), rabbit anti-vimentin antibody (10366-1-AP, 1:1,000
dilution; Proteintech), rabbit anti-laminin (LN) antibody
(23498-1-AP, 1:1,000 dilution; Proteintech). The procedures have
been described in detail previously (35).
Array hybridization and data
acquisition
Following 24 h atorvastatin or vehicle treatment
(10, 25, 50, 100, 200 µM) in HTM cells, total RNA was
extracted and purified using Trizol reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). Array hybridization was performed, using
GeneChip® Hybridization, Wash and Stain kit (cat
#900720, Affymetrix; Thermo Fisher Scientific, Inc.) in
Hybridization Oven 645 (cat #00-0331-220V, Affymetrix; Thermo
Fisher Scientific, Inc.) and Fluidics Station 450 (cat #00-0079,
Affymetrix; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. The slide was scanned with the default
settings of the GeneChip® Scanner 3000 (Cat #00-00212,
Affymetrix) and Command Console® Software 4.0
(Affymetrix). Raw data were normalized using the MAS 5.0 algorithm
(Affy packages in R).
GO and KEGG analysis
The differentially expressed genes were analyzed by
Gene Oncology (GO) enrichment, and were implemented, using the top
GO R packages based on the Fisher's exact test (43). Clusters of Orthologous Groups and
KEGG databases were used to classify and analyze differentially
expressed genes (44), and
P<0.05 was used as the significance enrichment standard.
Statistical analysis
Data were analyzed using SPSS17.0 version 12.0
software (SPSS, Inc.). All data are represented as the mean ±
standard deviation (SD). Analysis of variance (ANOVA) was used to
compare results among two or more groups, followed by least
significant difference (LSD) post hoc analysis; P<0.05 was
deemed to be a statistically significant difference.
Results
Atorvastatin reduces intraocular pressure
in a mouse model of ocular hypertension
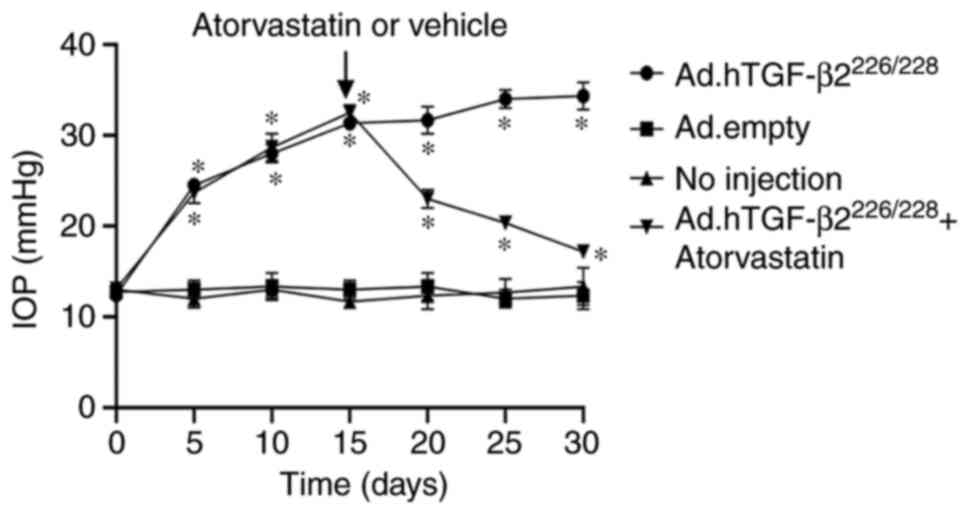
IOP was increased 2-fold following intravitreal
injection of Ad.hTGF-β2226/228 (12±1.6 vs. 24±1.7 mmHg,
P<0.01) at day 5 (Fig. 1). A
significantly elevated IOP of 32.3±0.7 mmHg was observed by day 15
following vector intravitreal injection, which plateaued until day
30, which was ~1.6-fold higher than the non-injection eyes
(12.8±0.4 mmHg) or the vehicle (Ad. Empty) eyes (12.6±0.8 mmHg)
(P<0.001). By day 20 (5 days after the commencement of the
gavage of atorvastatin), IOP was significantly decreased in the
hypertensive IOP group treated with atorvastatin compared to the
non-atorvastatin-treated group (21.5±0.7 vs. 32.5±0.7 mmHg,
P<0.01). By day 30, the IOP of the atorvastatin treatment
subgroup of the Ad.hTGF-β2226/228-induced hypertensive
IOP group continued to become significantly lower than that of the
non-atorvastatin-treated group (15.4±1.1 vs. 32.3±0.7 mmHg,
P<0.01). There were no adverse events during the period of the
experiment.
Atorvastatin toxicity and HTM cell
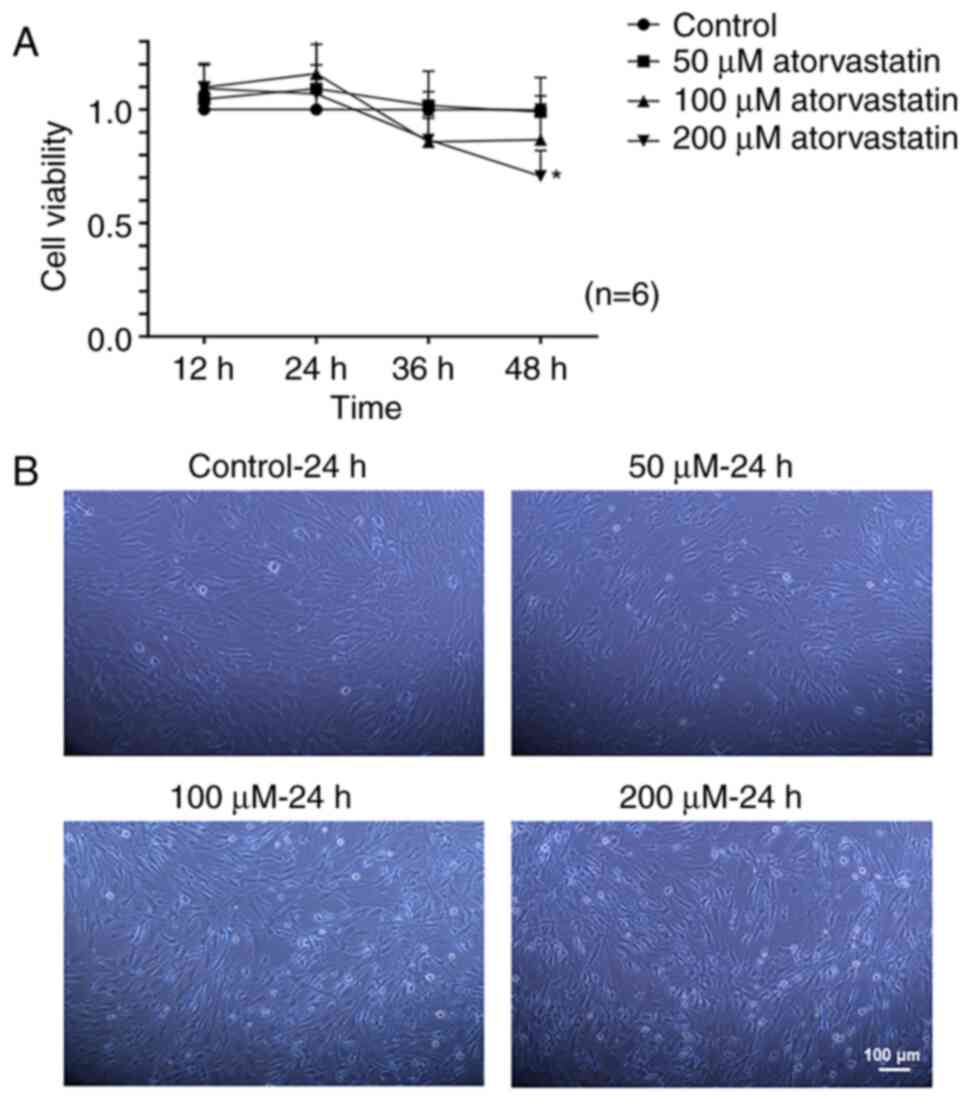
morphology
The isolated primary HTM cells grew out from the HTM
tissues (Fig. 2A) and expressed
FN, vimentin and LN (Fig. 2B),
and MYOC was increased after stimulation with DEX in a
dose-dependent manner (Fig. 2C).
There was no significant difference in viability of the HTM cells
treated with and without atorvastatin (50-100 µM), except
with atorvastatin at a dosage of 200 µM for 48 h (Fig. 3A). Thus, the time and the dose of
atorvastatin were selected to be no more than 24 h nor over 200
µM in the subsequent experiments (Fig. 3A). There were morphological
changes in the HTM cells following 24 h of statin treatment. The
shape of the HTM cells was altered from an elongated and adherent
cell morphology into rounded and detached cells with reduced
elongation in a dose-dependent manner following statin treatment
(Fig. 3B).
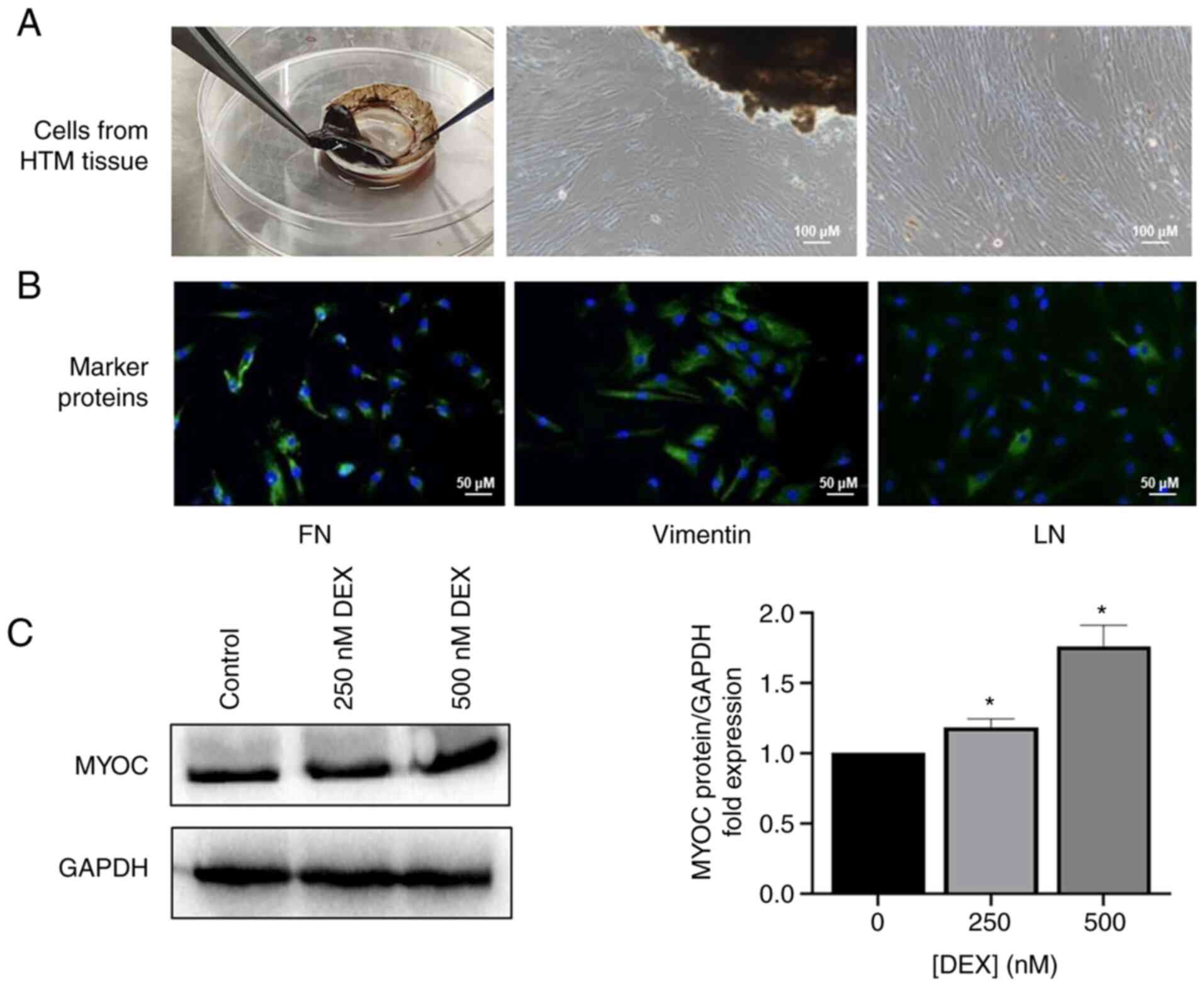 | Figure 2Characterization of HTM cells. (A)
HTM cells that grew out from the HTM tissues; (scale bar, 100
µm). (B) The expression of FN, vimentin, and LN of primary
HTM cells was evaluated immunocytochemically; (scale bar, 50
µm). (C) The level of MYOC in HTM cells following treatment
of DEX (250, 500 nM) for 6 days. *P<0.05. HTM, human
trabecular meshwork; FN, fibronectin; LN, laminin; MYOC, myocilin;
DEX, dexamethasone. |
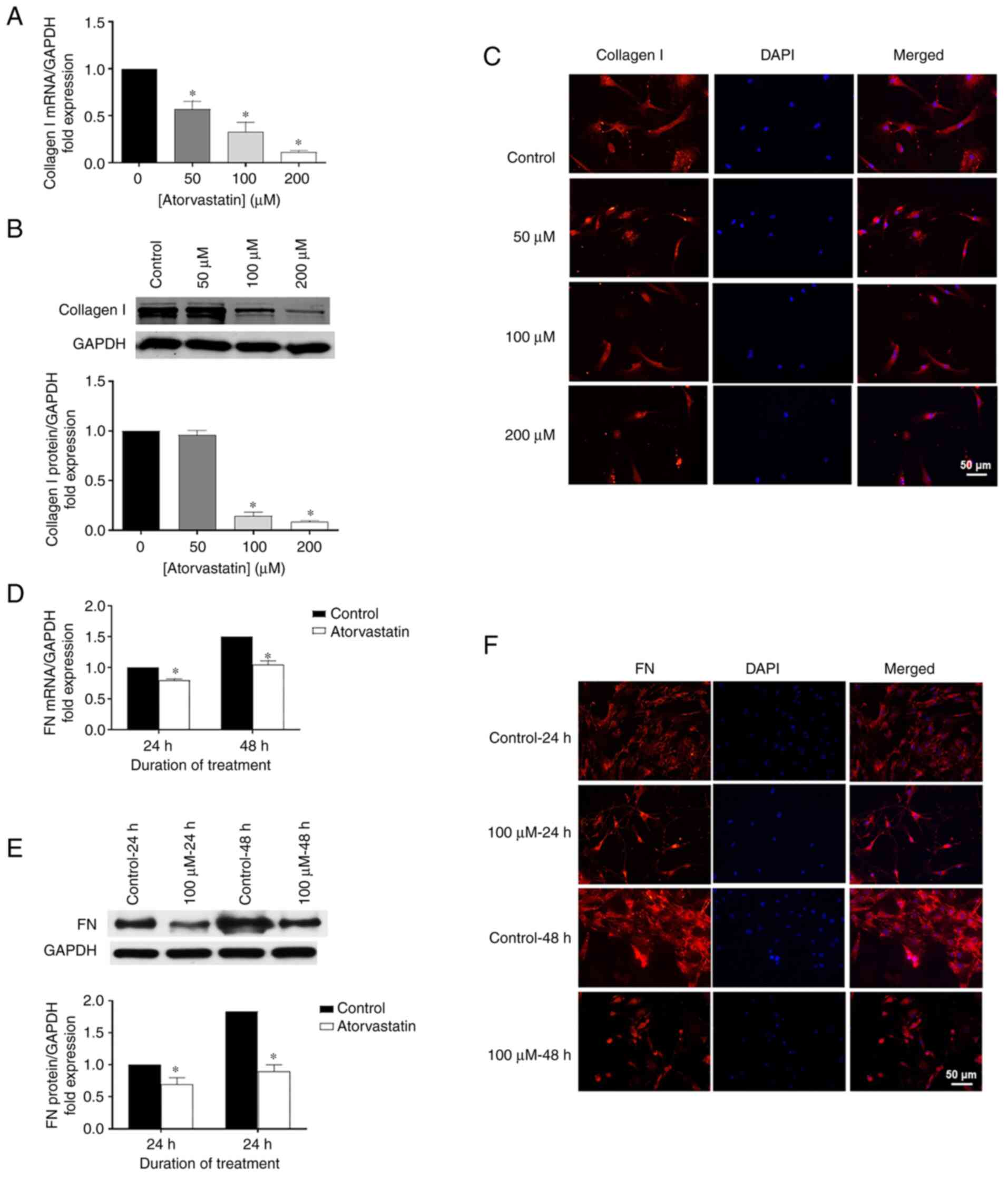
Effects of atorvastatin on expression
levels of collagen I and FN
Using RT-qPCR, western blot analysis and
immunohistochemistry, the role of atorvastatin on ECM expression
was explored, particularly for collagen I and FN. Collagen I mRNA
was significantly decreased in the HTM cells following atorvastatin
treatment for 24 h (Fig. 4A)
related to the change of dose, compared with control. Similar
patterns were observed for FN mRNA in the HTM cells treated with
100 µM atorvastatin for 24 or 48 h, showing significantly
decreased FN mRNA compared to the control (Fig. 4D). Consistent data were also
obtained for protein expression using western blot analysis
(Fig. 4B and E) and
immunohistochemistry (Fig. 4C and
F) in HTM cells following atorvastatin treatment.
Effects of atorvastatin on CTGF and
SPARC
In addition to ECM, atorvastatin inhibited CTGF and
SPARC expression, which are two important matricellular proteins.
To confirm the results obtained by RT-qPCR, Western blot analysis
and immunohistochemistry were performed. CTGF and SPARC mRNA were
significantly decreased in HTM cells followed atorvastatin
treatment for 24 h in a dose-dependent manner, comparing with
control (Fig. 5A and D). Similar
trends were obtained by western blot analysis (Fig. 5B and E) and immunohistochemistry
(Fig. 5C and F), showing that
the protein products of CTGF and SPARC were reduced following
atorvastatin treatment.
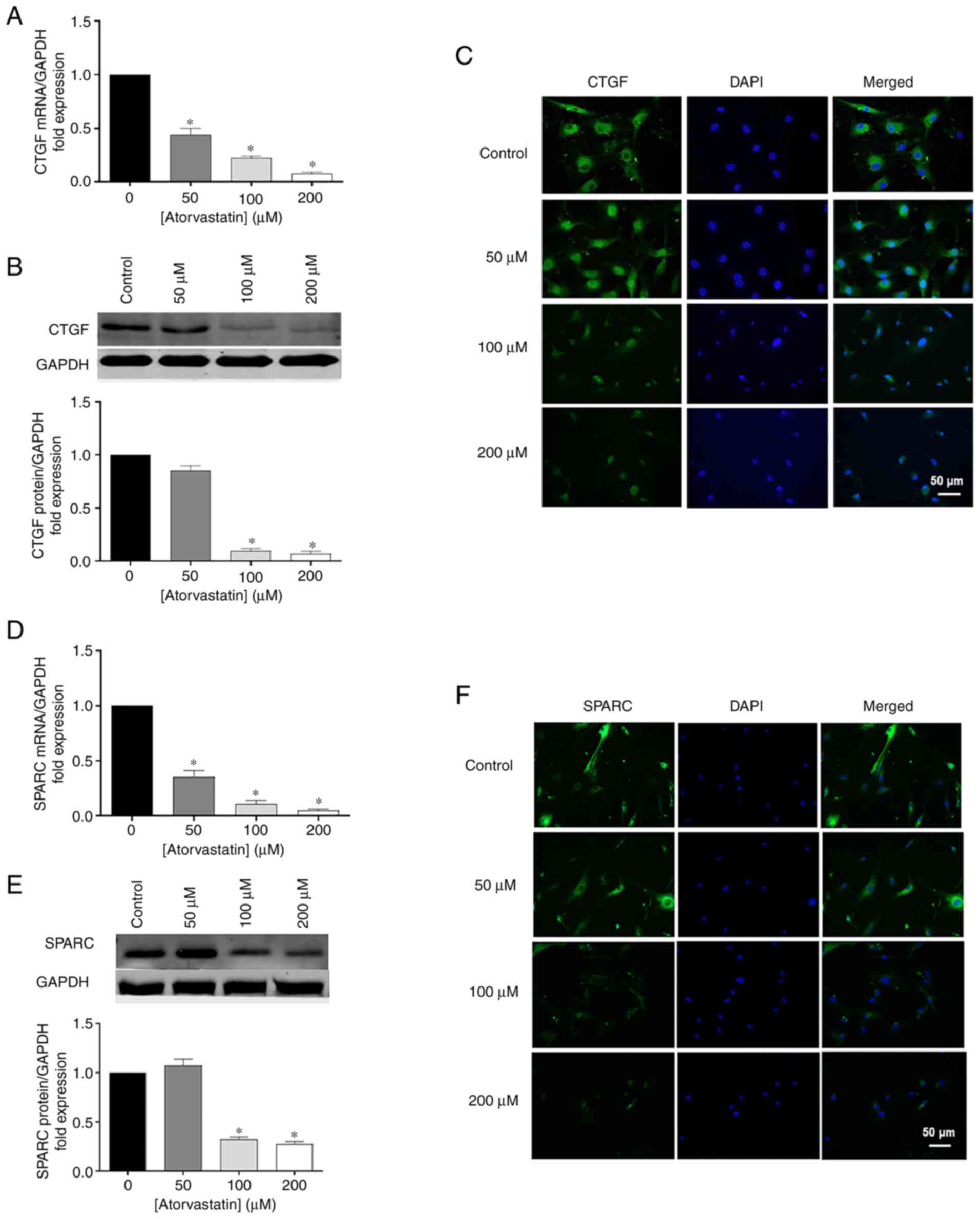 | Figure 5Effects of atorvastatin on decreasing
(A) mRNA, (B) protein levels, and (C) nuclear localization of CTGF
in HTM cells. Effects of atorvastatin on decreasing (D) mRNA, (E)
protein levels, and (F) nuclear localization of SPARC in HTM cells
(*P<0.05; n=4, n represents an independent experiment
number, using different major HTM cell lines; scale bar, 50
µm). HTM, human trabecular meshwork; CTGF, connective tissue
growth factor; SPARC, secreted protein and rich in cysteine. |
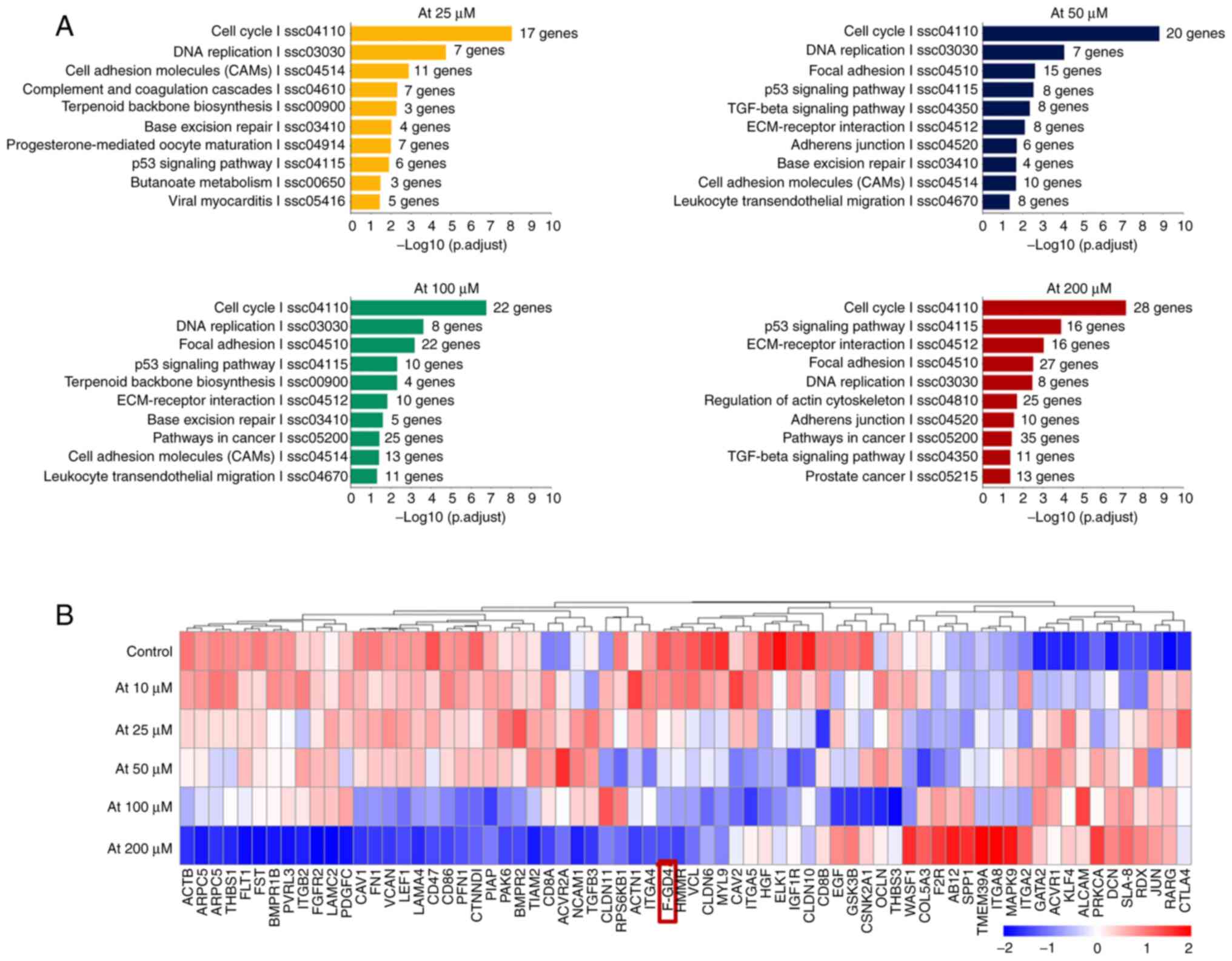
GO and KEGG analysis
Gene expression profile microarray was utilized for
detection of a correlation of differential gene expression
(Fig. 6). GO and KEGG gene
enrichment analysis showed that in addition to lipid metabolism and
cell cycle-related pathways (Fig.
6A), the differentially expressed genes mainly included 'cell
adhesion molecules', 'focal adhesion', the 'TGF-β signaling
pathway', 'ECM receptor interaction', 'adhesions junction',
'regulation of actin cytoskeleton and other extracellular matrix
synthesis pathways', 'cell-cell adhesion pathways', and
'cytoskeleton related pathways'. These pathways are all related to
POAG pathogenesis. Among these genes, FGD4 was downregulated most
significantly in a concentration-dependent manner (Fig. 6B).
Verification of the expression of FGD
protein isoforms in HTM and the inhibitory effect of atorvastatin
on FGD4 gene expression in trabecular meshwork cells
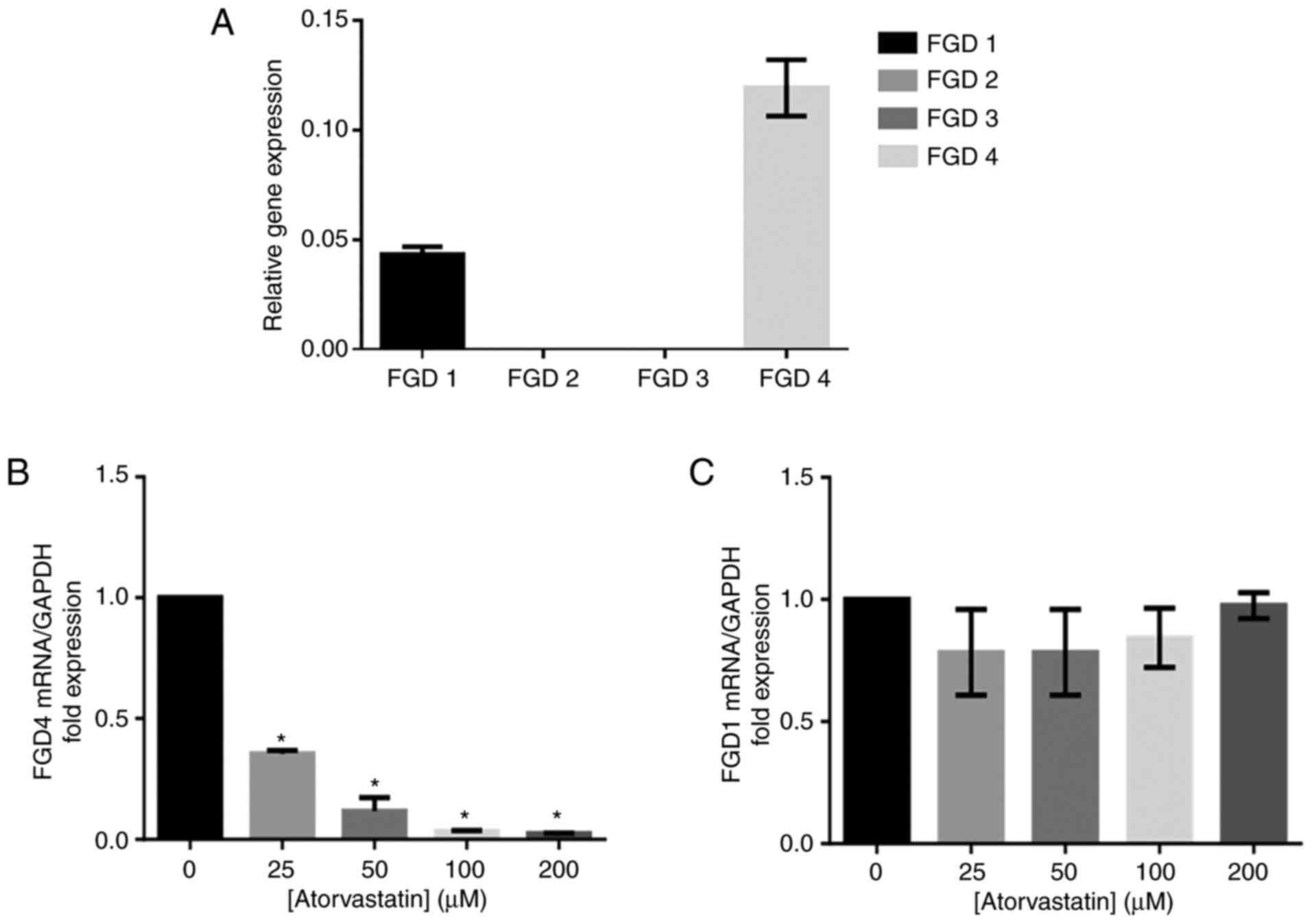
Our study found that the FGD4 gene was the
most highly expressed isoform of FGD in HTM cells, and FGD1 was the
second most highly expressed among the FGD genes (Fig. 7A). The expression of the
FGD4 gene was decreased with increasing atorvastatin
concentration (Fig. 7B). There
was no significant change in FGD1 gene expression following
atorvastatin treatment (Fig.
7C).
Discussion
In the present study, we observed that atorvastatin
reduced intraocular pressure (IOP) of ocular hypertension in
vivo and in vitro, consistent with the observed
morphologic change of isolated HTM cells. Atorvastatin also
suppressed two key matricellular proteins [secreted protein and
rich in cysteine (SPARC) and connective tissue growth factor
(CTGF)] and decreased extracellular matrix (ECM) synthesis
[collagen I and fibronectin (FN)] in human trabecular meshwork
(HTM) cells. In addition, our data support the hypothesis that
FYVE, RhoGEF and PH domain containing 4 (FGD4)/cell division
control protein 42 homolog (CDC42) might be the pivotal factors to
trigger these changes, which could be a novel potential therapeutic
target for glaucoma pathogenesis and treatment. All the findings
suggest that atorvastatin may be able to be a useful agent in the
management of glaucoma.
Ocular hypertension induced by
Ad.hTGF-β2226/228 can be reduced following atorvastatin
treatment, suggesting that statins are effective in the management
of ocular hypertension induced via the transforming growth factor
(TGF)-β2 pathway, which is in line with others (37,38), showing hypertensive IOP is
induced via the TGF-β2 route in vivo. However, the precise
underlying mechanism of atorvastatin in reducing IOP is still
unclear and will be clarified in the future, because elevated IOP
is the most pivotal risk factor contributing to the progression of
glaucoma (1,2).
Furthermore, there was no obvious cytotoxicity
observed in the HTM cells in response to atorvastatin treatment,
even at a rather high concentration, suggesting that the dosage of
atorvastatin used in the present study is safe and reliable.
Intercellular space between HTM cells was increased and HTM cell
size was reduced following atorvastatin treatment in a
dose-response manner, suggesting that there is a potential role for
statins in reducing IOP, which may also be associated with a
decrease in actin stress fibers and focal adhesion (45). The increased intercellular space
and smaller size of HTM cells could improve permeability of the TM
and reduce resistance of the TM to outflow. The permeability of the
TM is acutely modulated by the combined action of contractile and
volume-regulatory properties (46). Taken together, this could explain
why atorvastatin reduced IOP via decreasing cell-cell adhesion and
increasing intercellular space.
It has been reported that statins suppress ECM
synthesis in various non-ocular tissues (33,34). Furthermore, there is a
correlation between IOP and synthesis of ECM of the TM, where
reducing ECM contributes to a decrease in outflow resistance
(47), which is in line with our
finding that atorvastatin decreased collagen I and FN expression in
HTM cells. Thus, these data suggest that atorvastatin acts as a
novel anti-glaucoma medicine, targeting the pathogenesis of primary
open angle glaucoma (POAG). Moreover, we also found that
atorvastatin inhibited matricellular protein expression (SPARC and
CTGF), which enhance fibrosis and increase ECM deposition (10), providing a possible explanation
for the involvement of a reduction in IOP at the molecular
level.
FGD4 is closely related to the interaction among ECM
receptors, adhesion junctions, and the regulation of the actin
cytoskeleton, as demonstrated following GO and KEGG gene enrichment
analysis. We found that FGD4 was significantly inhibited in
response to atorvastatin treatment, suggesting the importance of
FGD4 during the development of POAG. FGD4 activates CDC42 via GTP.
CDC42 is one of the three members of the Rho protein guanosine
triphosphatase (GTPase) family (48), which is closely related to the
pharmacological effects of statins (49). The FGD4/CDC42 signaling pathway
influences cytoskeleton remodelling and ECM generation (50,51), and thus might also play a pivotal
role in glaucoma pathogenesis. Considering data from whole genome
microarray, we believe that atorvastatin reduces IOP via the
FGD4/CDC42 signaling pathway, which suppresses ECM expression and
cytoskeleton remodelling, which will be verified in a future
study.
In conclusion, the present study demonstrated that
atorvastatin reduced IOP, accompanied by an anti-ECM synthesis
effect, improved cellular spacing, and decreased focal adhesion
proteins. Statins may be good candidates as a novel treatment for
glaucoma. In future research, the precise underlying mechanisms of
the FGD4/CDC42 signaling pathway involvement in the anti-glaucoma
effect of atorvastatin will be investigated.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XYS and YYZ completed all experimental components
and contributed to the interpretation of the results. YYC, WTL, LC,
JLZ and YZ performed the statistical analysis and contributed to
the interpretation of the results. YYC and WTL participated in the
revision of this paper. YYZ contributed to the experimental design.
XYS and YYZ wrote the paper. All authors read and approved the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work (including all data) are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The animal study was conducted according to the
guidelines of the Declaration of Helsinki and approved by the
Institutional Animal Care and Use Committee of the Huashan
Hospital, Fudan University (Protocol #: JS-194, Approval Date: 19
February 2019). The primary HTM cell experiments were carried out
according to the guidelines of the Human Ethics Committee of
Huashan Hospital, Fudan University (protocol #: 2020-611, approval
date: 31 March 2020). Formal written consent was obtained from each
of the donors in this study.
Patient consent for publication
Not applicable.
Competing interests
All the authors declare no competing interests. This
research is original and has not been published elsewhere.
Acknowledgments
Not applicable.
Funding
This study was supported by the Science and Technology
Commission of Shanghai Municipality (Technology project
18441901400)
References
|
1
|
Weinreb RN, Aung T and Medeiros FA: The
pathophysiology and treatment of glaucoma: A review. JAMA.
311:1901–1911. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Weinreb RN and Khaw PT: Primary open-angle
glaucoma. Lancet. 363:1711–1720. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
De Moraes CG, Liebmann JM and Levin LA:
Detection and measurement of clinically meaningful visual field
progression in clinical trials for glaucoma. Prog Retin Eye Res.
56:107–147. 2017. View Article : Google Scholar :
|
|
4
|
Pederson JE, Gaasterland DE and MacLellan
HM: Uveoscleral aqueous outflow in the rhesus monkey: Importance of
uveal reabsorption. Invest Ophthalmol Vis Sci. 16:1008–1017.
1977.PubMed/NCBI
|
|
5
|
Larsson LI, Rettig ES and Brubaker RF:
Aqueous flow in open-angle glaucoma. Arch Ophthalmol. 113:283–286.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tam LC, Reina-Torres E, Sherwood JM,
Cassidy PS, Crosbie DE, Lütjen-Drecoll E, Flügel-Koch C, Perkumas
K, Humphries MM, Kiang AS, et al: Enhancement of outflow facility
in the murine eye by targeting selected tight-junctions of
Schlemm's canal endothelia. Sci Rep. 7:407172017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bradley J, Vranka J, Colvis CM, Conger DM,
Alexander JP, Fisk AS, Samples JR and Acott TS: Effect of matrix
metalloproteinases activity on outflow in perfused human organ
culture. Invest Ophthalmol Vis Sci. 39:2649–2658. 1998.PubMed/NCBI
|
|
8
|
Vranka JA, Kelley MJ, Acott TS and Keller
KE: Extracellular matrix in the trabecular meshwork: Intraocular
pressure regulation and dysregulation in glaucoma. Exp Eye Res.
133:112–125. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wight TN and Potter-Perigo S: The
extracellular matrix: An active or passive player in fibrosis? Am J
Physiol Gastrointest Liver Physiol. 301:G950–G955. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wallace DM, Murphy-Ullrich JE, Downs JC
and O'Brien CJ: The role of matricellular proteins in glaucoma.
Matrix Biol. 37:174–182. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brekken RA and Sage EH: SPARC, a
matricellular protein: At the crossroads of cell-matrix. Matrix
Biol. 19:569–580. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kuespert S, Junglas B, Braunger BM, Tamm
ER and Fuchshofer R: The regulation of connective tissue growth
factor expression influences the viability of human trabecular
meshwork cells. J Cell Mol Med. 19:1010–1020. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tamm ER: The trabecular meshwork outflow
pathways: Structural and functional aspects. Exp Eye Res.
88:648–655. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cela D, Brignole-Baudouin F, Labbé A and
Baudouin C: The trabecular meshwork in glaucoma: An inflammatory
trabeculopathy? J Fr Ophtalmol. 44:e497–e517. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shahid M, George TB, Saller J, Haija M,
Sayegh Z, Boulware D, Strosberg J, Chakrabarti R and Coppola D:
FGD4 (Frabin) overexpression in pancreatic neuroendocrine
neoplasms. Pancreas. 48:1307–1311. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sundararaman A and Mellor H: A functional
antagonism between RhoJ and Cdc42 regulates fibronectin remodelling
during angiogenesis. Small GTPases. 12:241–245. 2021. View Article : Google Scholar :
|
|
17
|
Ge J, Burnier L, Adamopoulou M, Kwa MQ,
Schaks M, Rottner K and Brakebusch C: RhoA, Rac1, and Cdc42
differentially regulate αSMA and collagen I expression in
mesenchymal stem cells. J Biol Chem. 293:9358–9369. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin J, Huang H, Lin L, Li W and Huang J:
MiR-23a induced the activation of CDC42/PAK1 pathway and cell cycle
arrest in human cov434 cells by targeting FGD4. J Ovarian Res.
13:902020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheema A, Chang RT, Shrivastava A and
Singh K: Update on the medical treatment of primary open-angle
glaucoma. Asia Pac J Ophthalmol (Phila). 5:51–58. 2016. View Article : Google Scholar
|
|
20
|
Rennie G, Wilkinson A, White A, Ruospo M,
Teixeira-Pinto A and Strippoli G: Topical medical therapy and
ocular perfusion pressure in open angle glaucoma: A systematic
review and meta-analysis. Curr Med Res Opin. 35:1421–1431. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Adams CM, Stacy R, Rangaswamy N, Bigelow
C, Grosskreutz CL and Prasanna G: Glaucoma-next generation
therapeutics: Impossible to possible. Pharm Res. 36:252018.
View Article : Google Scholar
|
|
22
|
Vaughan CJ and Delanty N: Neuroprotective
properties of statins in cerebral ischemia and stroke. Stroke.
30:1969–1973. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zacco A, Togo J, Spence K, Ellis A, Lloyd
D, Furlong S and Piser T: 3-hydroxy-3-methylglutaryl coenzyme A
reductase inhibitors protect cortical neurons from excitotoxicity.
J Neurosci. 23:11104–11111. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schmeer C, Kretz A and Isenmann S:
Statin-mediated protective effects in the central nervous system:
General mechanisms and putative role of stress proteins. Restor
Neurol Neurosci. 24:79–95. 2006.PubMed/NCBI
|
|
25
|
Song J, Deng PF, Stinnett SS, Epstein DL
and Rao PV: Effects of cholesterol-lowering statins on the aqueous
humor outflow pathway. Invest Ophthalmol Vis Sci. 46:2424–2432.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nakagami H, Jensen KS and Liao JK: A novel
pleiotropic effect of statins: Prevention of cardiac hypertrophy by
cholesterol-independent mechanisms. Ann Med. 35:398–403. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wilson HL, Schwartz DM, Bhatt HR,
McCulloch CE and Duncan JL: Statin and aspirin therapy are
associated with decreased rates of choroidal neovascularization
among patients with age-related macular degeneration. Am J
Ophthalmol. 137:615–624. 2004.PubMed/NCBI
|
|
28
|
Polman CH and Killestein J: Statins for
the treatment of multiple sclerosis: Cautious hope. Lancet.
363:15702004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Endo A: The discovery and development of
HMG-CoA reductase inhibitors. J Lipid Res. 33:1569–1582. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Stein JD, Newman-Casey PA, Talwar N, Nan
B, Richards JE and Musch DC: The relationship between statin use
and open-angle glaucoma. Ophthalmology. 119:2074–2081. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
McGwin G, McNeal S, Owsley C, Girkin C,
Epstein D and Lee PP: Statins and other cholesterol-lowering
medications and the presence of glaucoma. Arch Ophthalmol.
122:822–826. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Talwar N, Musch DC and Stein JD:
Association of daily dosage and type of statin agent with risk of
open-angle glaucoma. JAMA Ophthalmol. 135:263–267. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu H, Zeng L, Peng H, Chen S, Jones J,
Chew TL, Sadeghi MM, Kanwar YS and Danesh FR: HMG-CoA reductase
inhibitor simvastatin mitigates VEGF-induced 'inside-out' signaling
to extracellular matrix by preventing RhoA activation. Am J Physiol
Physiol. 291:F995–F1004. 2006. View Article : Google Scholar
|
|
34
|
Schaafsma D, Dueck G, Ghavami S, Kroeker
A, Mutawe MM, Hauff K, Xu FY, McNeill KD, Unruh H, Hatch GM and
Halayko AJ: The mevalonate cascade as a target to suppress
extracellular matrix synthesis by human airway smooth muscle. Am J
Respir Cell Mol Biol. 44:394–403. 2011. View Article : Google Scholar
|
|
35
|
Cong L, Fu S, Zhang J, Zhao J and Zhang Y:
Effects of atorvastatin on porcine aqueous humour outflow and
trabecular meshwork cells. Exp Ther Med. 15:210–216. 2018.
|
|
36
|
Shepard AR, Millar JC, Pang IH, Jacobson
N, Wang WH and Clark AF: Adenoviral gene transfer of active human
transforming growth factor-β2 elevates intraocular pressure and
reduces outflow facility in rodent eyes. Invest Ophthalmol Vis Sci.
51:2067–2076. 2010. View Article : Google Scholar
|
|
37
|
Fuchshofer R and Tamm ER: The role of
TGF-β in the pathogenesis of primary open-angle glaucoma. Cell
Tissue Res. 347:279–290. 2012. View Article : Google Scholar
|
|
38
|
Swaminathan SS, Oh DJ, Kang MH, Shepard
AR, Pang IH and Rhee DJ: TGF-β2-mediated ocular hypertension is
attenuated in SPARC-null mice. Invest Ophthalmol Vis Sci.
55:4084–4097. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang WH, Millar JC, Pang IH, Wax MB and
Clark AF: Noninvasive measurement of rodent intraocular pressure
with a rebound tonometer. Invest Ophthalmol Vis Sci. 46:4617–4621.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Stamer WD, Roberts BC, Epstein DL and
Allingham RR: Isolation of primary open-angle glaucomatous
trabecular meshwork cells from whole eye tissue. Curr Eye Res.
20:347–350. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Stamer WD and Clark AF: The many faces of
the trabecular meshwork cell. Exp Eye Res. 158:112–123. 2017.
View Article : Google Scholar :
|
|
42
|
Jurisic V, Srdic-Rajic T, Konjevic G,
Bogdanovic G and Colic M: TNF-α induced apoptosis is accompanied
with rapid CD30 and slower CD45 shedding from K-562 cells. J Membr
Biol. 239:115–122. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gracey AY, Fraser EJ, Li W, Fang Y, Taylor
RR, Rogers J, Brass A and Cossins AR: Coping with cold: An
integrative, multitissue analysis of the transcriptome of a
poikilothermic vertebrate. Proc Natl Acad Sci. 101:16970–16975.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kanehisa M, Goto S, Kawashima S, Okuno Y
and Hattori M: The KEGG resource for deciphering the genome.
Nucleic Acids Res. 32:D277–D280. 2004. View Article : Google Scholar :
|
|
45
|
O'Brien ET, Kinch M, Harding TW and
Epstein DL: A mechanism for trabecular meshwork cell retraction:
ethacrynic acid initiates the dephosphorylation of focal adhesion
proteins. Exp Eye Res. 65:471–483. 1997. View Article : Google Scholar
|
|
46
|
Honjo M and Tanihara H: Impact of the
clinical use of ROCK inhibitor on the pathogenesis and treatment of
glaucoma. Jpn J Ophthalmol. 62:109–126. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Faralli JA, Schwinn MK, Gonzalez JM Jr,
Filla MS and Peters DM: Functional properties of fibronectin in the
trabecular meshwork. Exp Eye Res. 88:689–693. 2009. View Article : Google Scholar :
|
|
48
|
Eliáš M and Klimeš V: Rho GTPases:
Deciphering the evolutionary history of a complex protein family.
Methods Mol Biol. 827:13–34. 2012. View Article : Google Scholar
|
|
49
|
Wang CY, Liu PY and Liao JK: Pleiotropic
effects of statin therapy: Molecular mechanisms and clinical
results. Trends Mol Med. 14:37–44. 2008. View Article : Google Scholar
|
|
50
|
Liu HP, Chen CC, Wu CC, Huang YC, Liu SC,
Liang Y, Chang KP and Chang YS: Epstein-Barr virus-encoded LMP1
interacts with FGD4 to activate Cdc42 and thereby promote migration
of nasopharyngeal carcinoma cells. PLoS Pathog. 8:e10026902012.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Bhoopathi P, Gondi CS, Gujrati M, Dinh DH
and Lakka SS: SPARC mediates Src-induced disruption of actin
cytoskeleton via inactivation of small GTPases Rho-Rac-Cdc42. Cell
Signal. 23:1978–1987. 2011. View Article : Google Scholar : PubMed/NCBI
|