Introduction
Porphyromonas gingivalis (Pg) is an anaerobic
Gram-negative asaccharolytic bacteria that is well-characterized
among the 'red-complex' perio-pathogens known to play a critical
role in the development of periodontitis (1) and systemic inflammatory diseases.
Pg produces several known bacterial toxins, such as
lipopolysaccharide (LPS), gingipains and fimbriae by local
secretion, as well as incorporation into and the release of complex
outer-membrane vesicles (OMVs) (2,3).
Over time, these factors are destructive to the local periodontium,
ultimately leading to additional dysbiotic changes, including
periodontitis and the loss of clinical epithelial attachment,
alveolar bone and pro-inflammatory mediators.
Among these, Pg LPS is known to cause
inflammation by triggering innate immune responses via a unique
Toll-like receptor interaction (4,5).
Indeed, the authors have previously demonstrated that the local
oral delivery of Pg LPS induced periodontitis and alveolar
bone loss in ApoE-deficient mice with increased amounts of
pro-inflammatory cytokines both locally and systemically (6).
On the other hand, Pg gingipains are cysteine
endo-proteases that exert their virulent effects by degrading the
extracellular matrix (7–9), cleaving numerous anti-bacterial
proteins in saliva and inducing systemic inflammation via the
interleukin (IL)-1β/NLR family pyrin domain containing 3
inflammasome pathway (10).
Pg produces three different types of gingipains:
Arginine-gingipain A (RgpA), arginine-gingipain B (RgpB) and
lysine-gingipain (Kgp) (7).
Together, these gingipains not only degrade and cause epithelial
cell detachment in the gingival tissues, but also degrade other
proteins such as complement system, cytokines and collagen
(11). Gingipains, as already
mentioned, are known to be secreted, and transported to the
extracellular bacterial environment in soluble and OMV-associated
forms (12,13). They are also essential for the
survival and pathogenicity of Pg, playing critical roles in
the bacterial colonization, inactivation of host defenses and
tissue destruction (13,14).
To date, a number of studies have developed and
utilized murine models in attempts to examine the effects of
Pg on both local and systemic alterations in various tissues
(15,16). Among these, the inoculation of
live bacteria into the oral cavity is frequently used (17). This model requires the
application of repeated, large concentrations/volumes of live
bacteria into the oral cavity (3–4 times per week over a long
period of time, such as 8–12 weeks) without causing efficient
pathologic outcomes, such as alveolar bone loss, the key outcome
measurement of periodontitis (18,19). The injection of LPS into the
gingival tissues has also been used (20,21); however, it does not account for
the effects of other bacterial pathogens. Experimental
periodontitis has also been induced with the placement of
Pg-adhered ligatures into the gingival sulcus in mice
(22,23). However, a concern associated with
this model is that the effect of bacteria may be masked by the
mechanistic trauma from the ligature. Collectively, these models do
not faithfully mimic the true clinical settings in which Pg
colonizes around the tooth to assert local and systemic
effects.
The main aim of the present study was to establish a
mouse model in which a small volume of Pg is topically
applied directly into the gingival pocket to allow bacterial
colonization and asserts both local and systemic effects. The
wild-type mouse model developed herein demonstrated the
establishment of a chronic active infection of Pg into the
oral cavity/gingival pockets similar to that observed in a human
infection/colonization with Pg. Topical Pg
application into the gingival pocket resulted in local chronic
colonization, as well as local and systemic inflammation, alveolar
bone loss, and the accumulation of Pg LPS and gingipain
aggregates in gingival tissue and aortic walls in mice.
Materials and methods
Animals and animal welfare
considerations
A total of 50 4-week-old mice (C57BL6 background)
were purchased from Jackson Laboratory. All mice were housed in a
pathogen-free animal experimental facility at the University of
California, Los Angeles University, under a 12-h light/dark cycle.
All mice were fed normal chow and had free access to drinking water
and food. The health and behavior of the mice were monitored three
times a week throughout the whole duration of the experiment (12
weeks). Isoflurane (2%) and a mixture of ketamine (100 mg/kg) and
xylazine (5 mg/kg) were used as anesthetics during ligature
placement and phosphate-buffered saline (PBS) or bacteria
inoculation. Carprofen (3 mg/kg), a pain relief drug, was also used
after ligature placement to minimize the pain of the mice. The
ketamine/xylazine mixture and carprofen were administered via
intraperitoneal (i.p.) injection, and isoflurane was administered
via inhalation. All mice were administered ketamine/xylazine prior
to euthanasia to minimize suffering. Euthanasia was performed via
cardiac perfusion, and the heartbeat of the mice was assessed for 5
min to verify death. All mice were euthanized as designed, apart
from 1 mouse that died during the bacteria inoculation via
isoflurane inhalation. All procedures were performed in compliance
with the institution's policy and applicable provisions of the
United States Department of Agriculture (USDA) Animal Welfare Act
Regulations and the Public Health Service (PHS) Policy. The
experimental protocols were approved by the Animal Research
Committee (ARC) of the University of California, Los Angeles (UCLA)
under ARC# 2019-057.
Creation of gingival pocket to retain
topically applied PBS or bacteria in the pocket
To create a gingival pocket that allows for
retaining topically applied PBS or Pg W83 (Pg)
(obtained from Dr Gena D. Tribble, University of Texas School of
Dentistry, Houston, TX, USA) directly into the gingival pocket, a
6-0 silk ligature was placed around the second molars for 1 week
under general anesthesia using ketamine/xylazine (100 and 5 mg/kg,
respectively) as previously described (6,24). After the ligature placement, all
mice were administered 2 mg ampicillin and 2 mg neomycin daily by
gavage for 4 days out of the 7 days in total of the ligature to
facilitate the subsequent Pg application into the pocket and
to enhance Pg bacterial colonization in the pocket.
Following the removal of the ligature 1 week after placement, the
mice were divided into 4 groups as follows: Group 1 (n=10), PBS
application for 5 weeks and sacrifice at the 4th week after the
final inoculation; group 2 (n=15), Pg application for 5
weeks and sacrifice at the 4th week after the final administration;
group 3 (n=10), PBS application for 5 weeks and sacrifice at the
8th week after the final administration; and group 4 (n=15),
Pg application for 5 weeks and sacrifice at the 8th week
after the final application.
Culture of Pg and topical application of
PBS or Pg directly into the gingival pocket
Pg was cultured based on the recommended
protocol with some modifications (25). Briefly, the Pg culture was
grown for 3–5 days on a tryptic soy blood agar plate (TSB
containing 1.5% agar and 5% defibrinated sheep blood, Hemostat
Laboratories) supplemented with 5 μg/ml of hemin, 0.5
μg/ml vitamin K1 (Difco; BD Biosciences) and
0.05% L-cysteine (Sigma-Aldrich; Merck KGaA) in an anaerobic
chamber at 37°C until OD600 of ~1.5 [~7.3×109
colony forming U/ml (CFU/ml)]. The Pg culture was
concentrated by centrifugation at 10,000 × g at 20°C for 15 min and
washed once with PBS before the bacterial pellet was suspended in
sterile 1% methyl cellulose solution to yield a final concentration
of 5×1011 CFU/ml. The fresh Pg culture
preparations were conducted three times a week for 5 consecutive
weeks. Pg (10 μl) in methyl cellulose solution
(5×109 CFU per gingival pocket) or 10 μl PBS
prepared in methyl cellulose solution were topically applied
directly onto the lingual side of the maxillary second molar into
the subgingival area using the microvolume micropipette (0.1–10
μl) under the BM-LED microscope (Meiji Techno) (Fig. S1).
Sample and tissue collection
Whole blood was collected from mice by cardiac
puncture under general anesthesia with isoflurane (Abbott
Pharmaceutical Co. Ltd.). Following blood collection, swab samples
were obtained from the gingival pocket and gingival tissue using
sterile endodontic absorbent paper points (Dentsply) for
semi-quantitative PCR analysis to determine the presence of
Pg (Pg colonies) in the pocket. The mice were then
perfused and fixed with 4% paraformaldehyde in PBS via the left
ventricle for 5 min. Following perfusion, the heart and a short
section of the aorta root were removed for cryosection. The
maxillae of the mice were then excised and fixed with 4%
paraformaldehyde in PBS, pH 7.4, at 4°C over-night and stored in
70% ethanol solution for micro-computed tomography (μCT)
analysis.
Frozen sectioning and staining of aortic
root
The heart samples were embedded in cyromolds with
Tissue-Tek O.C.T. compound (Sakura Finetek), and stayed frozen at
−80°C until cryo-sectioning. Frozen heart blocks were sectioned at
a thickness of 20 μm at −20°C using a CM3050 S cryostat
(Leica Microsystems, Inc.) and a Cyrostar NX70 cyrostat (Thermo
Fisher Scientific, Inc.) for hematoxylin and eosin (H&E;
Sigma-Aldrich; Merck KGaA) and Oil Red O (Sigma-Aldrich; Merck
KGaA) staining of the aortic root region for 15 min at room
temperature.
Detection of Pg colonies from the
gingival pocket
Swab samples taken from the gingival pocket were
stored in 200 μl PBS. Genomic DNA was extracted using the
Qiagen QIAamp DNA Micro kit (Qiagen, Inc.) from the sample
following proteinase K (Thermo Fisher Scientific, Inc.) treatment.
The protein-free purified DNA was then dissolved in 20 μl
distilled water. An aliquot (5 μl) of the dissolved DNA was
diluted 10-fold (to yield 50 μl diluted DNA samples). The
diluted DNA sample was boiled for 5 min. The resulting lysate (4
μl) was then used directly as a template in a PCR reaction:
The amount of DNA used for the PCR analysis was 1/50 of original
samples taken from each swab. All PCR reactions were performed as
previously described (26) using
the following primers, which detect Pg W83-specific 16S
rDNA: Forward, 5′-AGG CAG CTT GCC ATA CTG CG-3′; and reverse,
5′-ACT GTT AGC AAC TAC CGA TGT-3′. Each PCR reaction was composed
of a denaturation cycle of 95°C for 8 min, followed by 50 cycles
composed of one step of 95°C for 30 sec, an annealing step of 56°C
for 30 sec, and an extension step at 72°C for 40 sec. PCR was
performed using a thermal cycler (SimpliAmp, Applied Biosystems;
Thermo Fisher Scientific, Inc.). Amplified products were detected
by electrophoresis. Each gel run was performed at 1% ethidium
bromide on a 1% agarose gel at 110 V for 20 min. The DNA ladder was
1 Kb plus DNA ladder from Thermo Fisher Scientific, Inc. Purified
Pg DNA (1,000 CFU) for PCR was included as a positive
control to quantify the CFU of Pg from each pocket swab, and
PCR without template DNA served as a negative control. The density
of the amplified DNA was analyzed using ImageJ software Version
1.52 (National Institute of Health).
μCT and histological analysis of
maxillae
The fixed maxillae were subjected to μCT
scanning (Skyscan1275, Bruker-microCT Systems) using a voxel size
of 20 μm3 and a 0.5 mm aluminum filter.
Two-dimensional slices from each maxilla were combined using NRecon
and CTAn/CTVol programs (Bruker-microCT Systems) to form a
three-dimensional reconstruction. The level of bone resorption was
calculated as the distance from the palatal and mesiobuccal
cement-enamel junction (CEJ) to the alveolar bone crest (ABC) of
the second molars by an investigator (YB). The reading was
confirmed in a blinded-manner by another individual (SK).
Following μCT scanning, the maxillae were
decalcified with 5% EDTA (Sigma-Aldrich; Merck KGaA) and 4% sucrose
(Sigma-Aldrich; Merck KGaA) in PBS (pH 7.4) for 3 weeks at 4°C. The
decalcification solution was changed daily. Decalcified maxillae
were processed for paraffin embedding the blocks at the UCLA
Translational Procurement Core Laboratory (TPCL). Blocks were
sectioned at 5-μm using a microtome (Thermo Fisher
Scientific, Inc.). After dewaxing with xylene, the sections were
stained with hematoxylin and eosin (H&E, Sigma-Aldrich; Merck
KGaA) at room temperature for 30 sec. Digital images of the stained
sections were obtained using the DP72 microscope (Olympus
Corporation). The clinical attachment loss (CAL) was obtained under
the microscope by measuring the CEJ to the base of the pocket depth
by an investigator (SK). The reading was confirmed in a
blinded-manner by another individual (YB).
Antibody production for Pg lysine
gingipain (Kgp) and arginine gingipain B (RgpB)
As anti-Pg (strain W83) gingipain antibodies
were of limited availability, antibodies against Pg
gingipains were generated. Briefly, two sets of primers were
designed to amplify the sequence encoding Kgp amino acids A22 to
I400 and RgpB N401 to K736 from Pg W83 (Table SI). The amplified DNA fragments
were cloned into pMCSG7 using a ligation-independent cloning (LIC)
method as previously described (27), and the gingipain coding sequences
were confirmed by DNA sequencing. Generated gingipain-expressing
plasmids were introduced into E. coli BL21 (DE3) (Thermo
Fisher Scientific, Inc.). Both cultures of 500 ml Luria-Bertani
(LB) broth in the presence of 100 μM ampicillin were induced
by the addition of 0.5 mM Isopropyl ß-D-1-thiogalactopyranoside
(IPTG) at OD600 of 0.7–0.8 at 18°C overnight. The
induced cell cultures were pelleted by centrifugation at 6,500 × g
for 10 min at 4°C and washed with 50 mM Tris·Cl and 150 mM NaCl, pH
7.5 prior to suspension in sample buffer containing 1 mM PMSF, 20
mM β-mercaptoethanol and 20 mM imidazole. The cell suspensions were
subjected to French Press (Glen Mills) to lyse the cells. The
lysates were centrifuged at 6,500 × g for 20 min at 4°C to separate
the unlysed cells or insoluble proteins from the soluble proteins.
Recombinant proteins were purified using Ni-NTA agarose affinity
chromatography and purified proteins were analyzed by 0.1%
Coomassie blue R250 (Fisher Bioreagents) in 50% methanol and 10%
acetic acid solution to confirm the successful purification. The
purified proteins, Kgp (7 mg/ml) and RgpB (3.8 mg/ml), were shipped
to Cocalico Biologicals, Inc. for raising custom polyclonal
antibody in rabbits. The produced antiserums were used for the
detection of gingipains (or antibody against gingipains) in
gingival tissue, blood and arterial walls.
Determination of pro-inflammatory
cytokines in mouse serum
The levels of pro-inflammatory cytokines were
detected as follows: Briefly, whole mouse blood was collected at 4
or 8 weeks after the final PBS or Pg inoculation with
cardiac puncture. The serum was separated from the blood for the
detection of pro-inflammatory cytokines using the Quantibody Mouse
Cytokine Array kit (RayBiotech, Inc.) which allows for the
determination of low levels (<20–30 pg/ml) of cytokines [e.g.,
granulocyte-macrophage colony-stimulating factor (GM-CSF),
interferon (IFN)-γ, IL-1α, IL-1β, IL-6, IL-17, tumor necrosis
factor (TNF)-α, vascular endothelial growth factor (VEGF),
macrophage colony-stimulating factor (M-CSF), keratinocyte
chemoattractant (KC)] from the serum samples.
Reverse transcription-quantitative PCR
(RT-qPCR) for determining the expression levels of pro-inflammatory
cytokines from gingival and aortic tissue
Total RNA was extracted from mouse gingival and
aortic tissue using the RNeasy micro kit (Qiagen GmbH) and reverse
transcribed for 5 min at 65°C, 2 min at 25°C and for 50 min at 45°C
cycles using the SuperScript® III Reverse Transcriptase
Synthesis kit (Thermo Fisher Scientific, Inc.). Subsequently, qPCR
was performed using PowerUp™ SYBR-Green Master Mix (Thermo Fisher
Scientific, Inc.) or TaqMan primers (Applied Biosystems; Thermo
Fisher Scientific, Inc.) for 2 min at 95°C (one step denaturation)
and amplification of DNA with 15 sec at 95°C and 1 min at 60°for 40
cycles as suggested by the manufacturer (Thermo Fisher Scientific,
Inc.). The sequences of the primers used for RT-qPCR are presented
in Table SI. Glyceraldehyde
3-phosphate dehydrogenase (GAPDH) served as a control and the fold
induction was calculated using the comparative ΔCq method and are
presented as relative transcript levels (2-ΔΔCq)
(28).
Detection of anti-Pg gingipains (Kgp and
RgpB) and anti-Pg LPS antibodies from mouse serum
ELISA was performed on collected mouse sera to
determine Pg Kgp-, RgpB- and LPS-specific antibodies. ELISA
plates (Corning, Inc.) were coated with 100 ng Kgp or RgpB in
carbonate-bicarbonate buffer (pH 9.6). ELISA plate kit pre-coated
with 100 ng of Pg LPS were obtained from Chondrex, Inc. for
the detection of anti-LPS antibody from serum (cat. no. 6222;
Chondrex, Inc.). The coated wells of each plate were blocked with
2% bovine serum albumin (BSA) (Chondrex, Inc.), and various
dilutions of mouse sera (1:100, 1:200, 1:400, 1:800, 1:1,600 and
1:3,200) were added to the wells and incubated for 2 h at room
temperature. Anti-mouse horse radish peroxidase (HRP)-conjugated
IgG antibody (cat. no. 7076, Cell Signaling Technology, Inc.) was
then added at a 1:500 ratio for 1 h, followed by TMB substrate
solution (eBioscience; Thermo Fisher Scientific, Inc.), and after 6
min, the resultant color intensity was recorded at 450 nm. Antibody
levels as the absorbance OD value was measured using the Infinite
M1000 microplate reader (Tencan).
Detection of Pg gingipain (Kgp and RgpB)
and LPS aggregates in the gingiva and arterial wall of mice
For immunofluorescence analysis, formalin-fixed
paraffin-embedded sections of gingival tissues and frozen sections
of hearts were incubated with primary antibodies against Pg
Kgp (rabbit polyclonal antibody), Pg RgpB (rabbit polyclonal
antibody) and Pg LPS (mouse monoclonal antibody from
Millipore Sigma, followed by fluorometric detection with Alexa
Fluor 488-conjugated secondary antibodies (Thermo Fisher
Scientific, Inc.). Sequentially, the sections were mounted on
slides with VECTASHIELD® anti-fade mounting medium with
4′,6-diamidino-2-phenylindole dihydrochloride (DAPI; Vector
Laboratories, Inc.). The slides were then examined under a Fluoview
FV200i confocal fluorescent microscope (Olympus Corporation).
Digital images of the stained sections were obtained using a
microscope (DP72; Olympus Corporation).
Statistical analysis
All graphs were created and statistical analyses
were performed using GraphPad Prism 9.3.1 (GraphPad Software,
Inc.). An unpaired Student's t-test was used for two-group
comparisons, and for multiple comparisons, one-way ANOVA with
Turkey's post hoc test was used. A P-value <0.05 was considered
to indicate a statistically significant difference. All results
from in vitro experiments were confirmed by at least three
independent experiments. Error bars represent the mean ± SEM.
Results
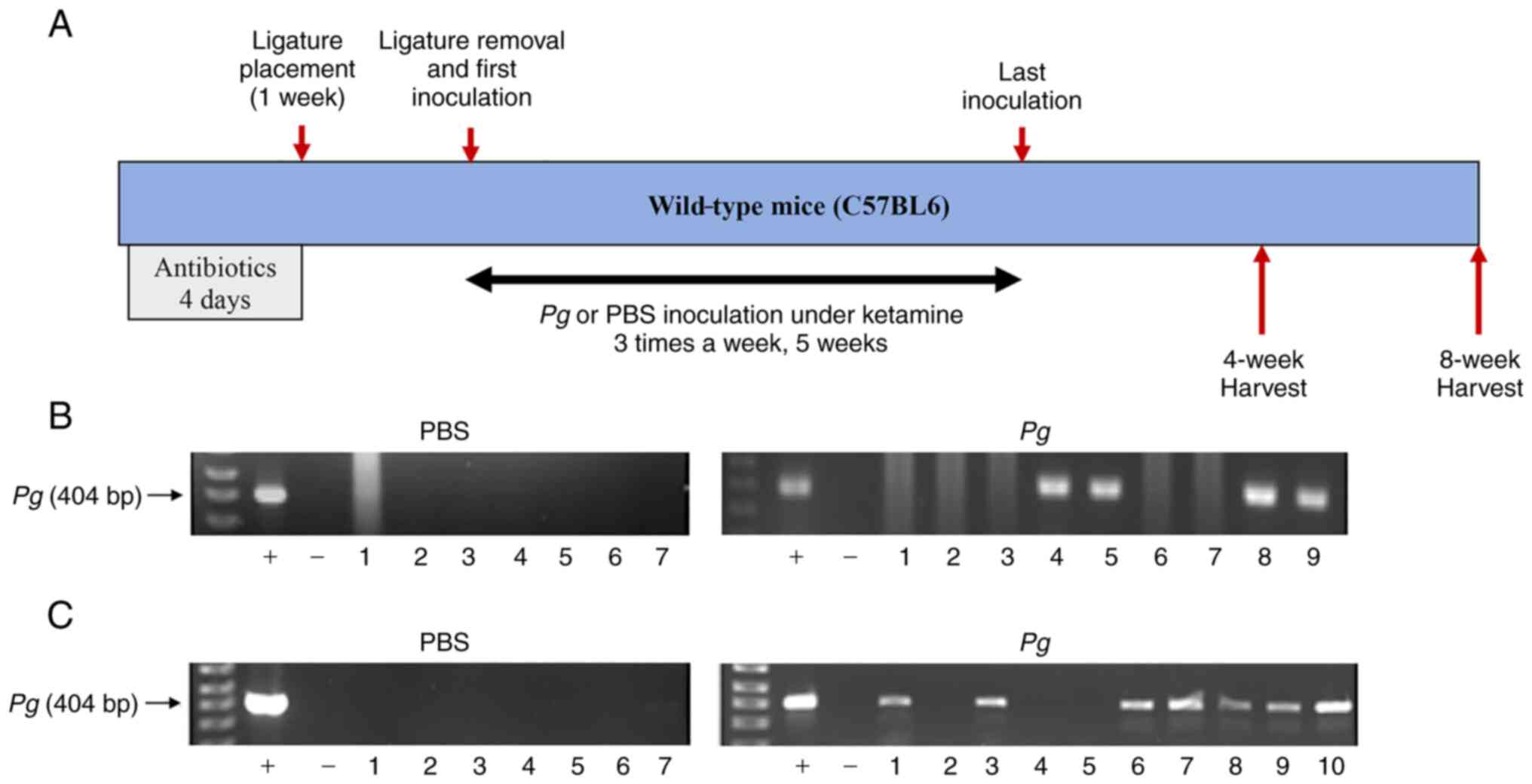
Topical Pg application into the gingival
pocket induces Pg colony formation in the majority of gingival
pockets in mice
To retain PBS and Pg in the gingival pockets
following the topical application, artificial pockets were created
around the maxillary second molars of mice by placing silk ligature
around the molars. At 1 week after the placement, the ligatures
were removed and PBS or freshly cultured Pg were topically
applied into the gingival pocket three times per week for 5 weeks.
After completing the course of PBS or Pg application, the
mice were left for 4 or 8 weeks in cages, after which they were
sacrificed (Fig. 1A). As shown
in Fig. 1, the swab samples from
the mice receiving the topical application of PBS directly into the
gingival pocket did not exhibit any presence of amplified bacterial
DNA. However, the swab samples from the mice receiving the topical
application of Pg exhibited amplified bacterial DNA bands in
4 samples (out of 9) and 7 samples (out of 10) when analyzed at 4
or 8 weeks, respectively after the final Pg inoculation
(Fig. 1B and C). The amount of
detected Pg/swab/mouse was ~150,000–210,000 CFU at 4 weeks
and ~15,000–55,000 CFU at 8 weeks after the final Pg
inoculation. These data indicate that the repeated topical
application of Pg into the gingival pocket can induce and
establish bacterial colony formation in the mouse gingival
pocket.
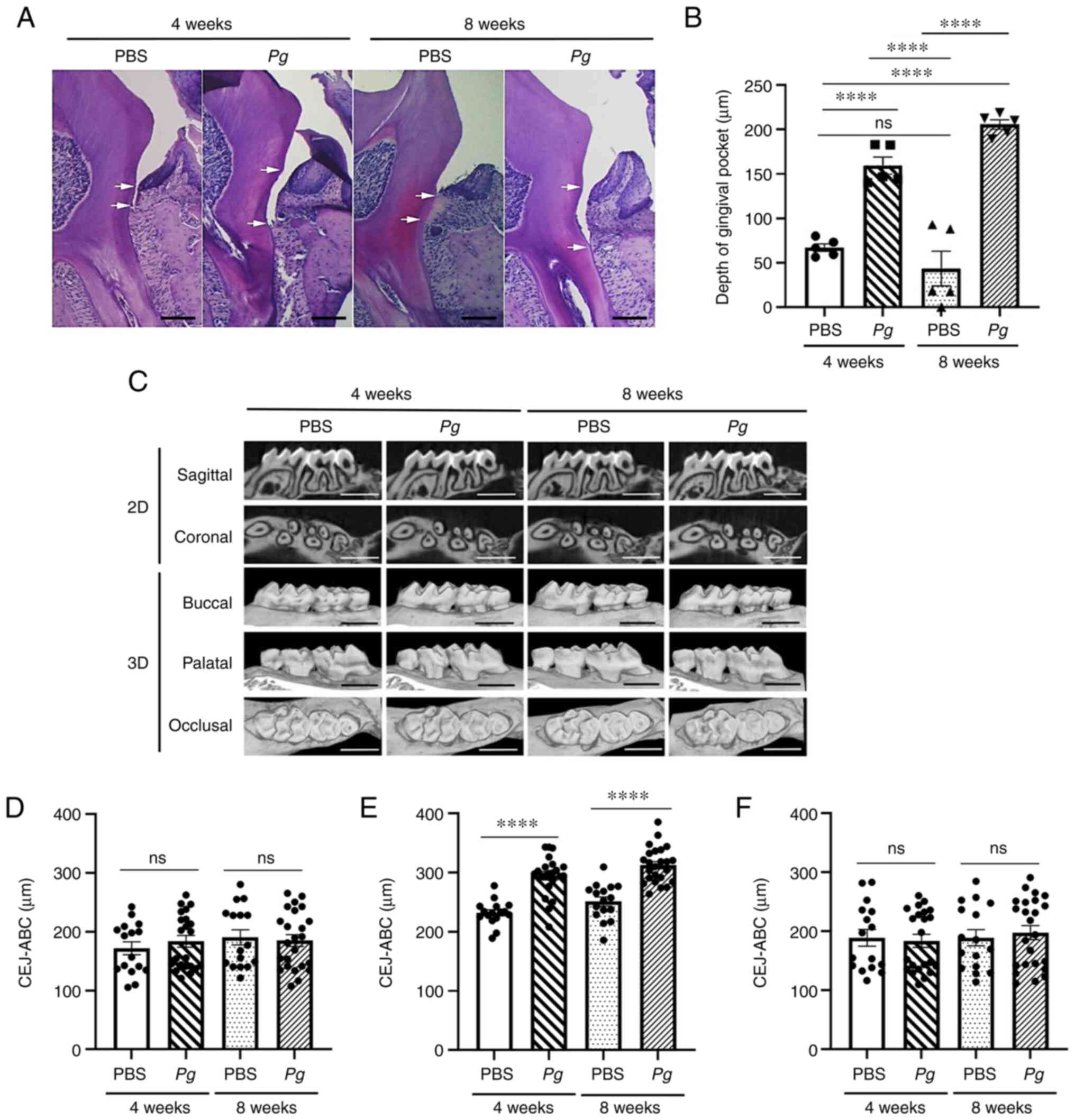
Topical Pg application induces the loss
of the clinical epithelial attachment and alveolar bone
The histological examination revealed that the
clinical epithelial attachment loss measured from the CEJ to the
base of the gingival pocket was significantly increased in the
Pg-inoculated site, and the clinical attachment loss was
more significant in mice at 8 weeks when compared to that in mice
at 4 weeks (Fig. 2A and B),
suggesting ongoing active inflammation after 4 weeks of the
epithelial junction. The μCT analysis also revealed similar
patterns in bone loss (Fig. 2C).
When alveolar bone loss was measured from the CEJ to the crest of
the alveolar bone, only the site at which Pg was applied
(second molars) exhibited bone loss, while the sites at which
Pg was not applied were unaltered (Fig. 2D-F), indicating the specificity
of Pg-induced bone loss.
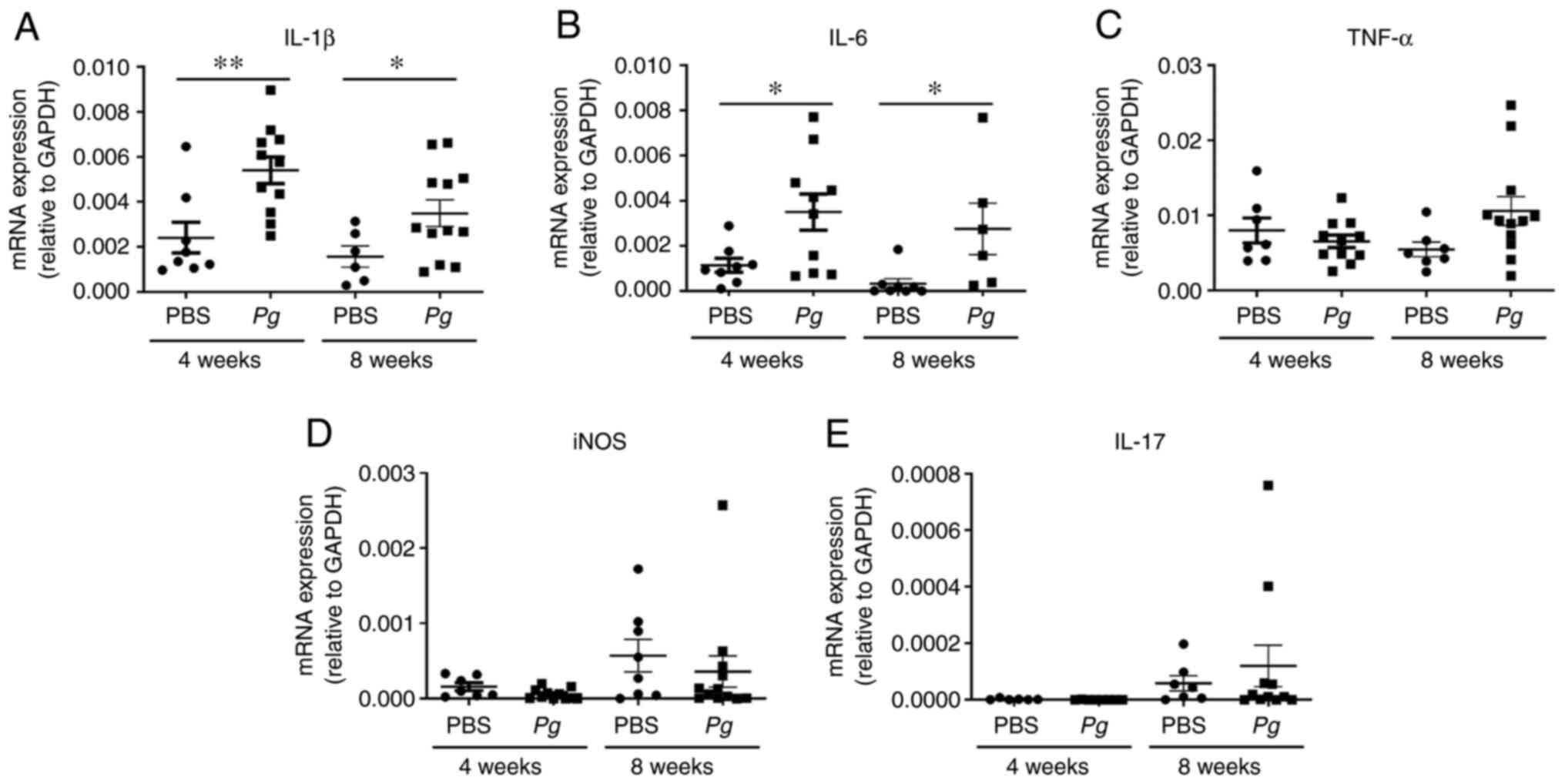
Topical Pg application increases the
expression of pro-inflammatory cytokines in the gingival
tissue
To further examine the inflammatory status around
the Pg-inoculated soft tissue, gingival tissues around the
second molars were isolated and subjected to RT-qPCR analysis for
determining the expression levels of IL-1β, IL-6, TNF-α, inducible
nitric oxide synthase (iNOS) and IL-17. The expression levels of
pro-inflammatory cytokines, including IL-1β and IL-6 were
upregulated by the 4th and 8th week when compared with the controls
with PBS treatment (Fig. 3A and
B). The other screened cytokines, such as TNF-α, iNOS and IL-17
were also upregulated, although the increases were not
statistically significant (Fig.
3C-E). These data indicated that the topical Pg
application into the gingival pocket induced local inflammation
around the tooth.
Topical Pg application directly into the
gingival pocket induces systemic inflammation
As periodontitis embodies systemic inflammation, the
levels of inflammatory cytokines in serum were examined to
determine whether the topical Pg application can induce
systemic inflammation. In mice receiving the topical Pg
application, significantly higher levels of pro-inflammatory
cytokines, such as TNF-α, IL-13, IL-1β, IL-17, KC and IFN-γ, were
detected in serum compared to those in mice receiving PBS when
measured at the 4th week after the final Pg or PBS
inoculation (Fig. 4). However,
higher levels of IL-1β and KC were detected only in the serum of
mice receiving Pg compared to those receiving PBS when
measured at the 8th week after the final Pg or PBS
inoculation (Fig. 4). The serum
levels of IL-1α, IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, IL-12,
M-CSF, GM-CSF, VEGF, monocyte chemotactic protein-1 (MCP-1) or
regulated upon activation, normal T cell expressed and secreted
(RANTES) were not markedly altered by the Pg inoculation
regardless of the time point (4th or 8th week) after the final
Pg inoculation (Fig.
S2).
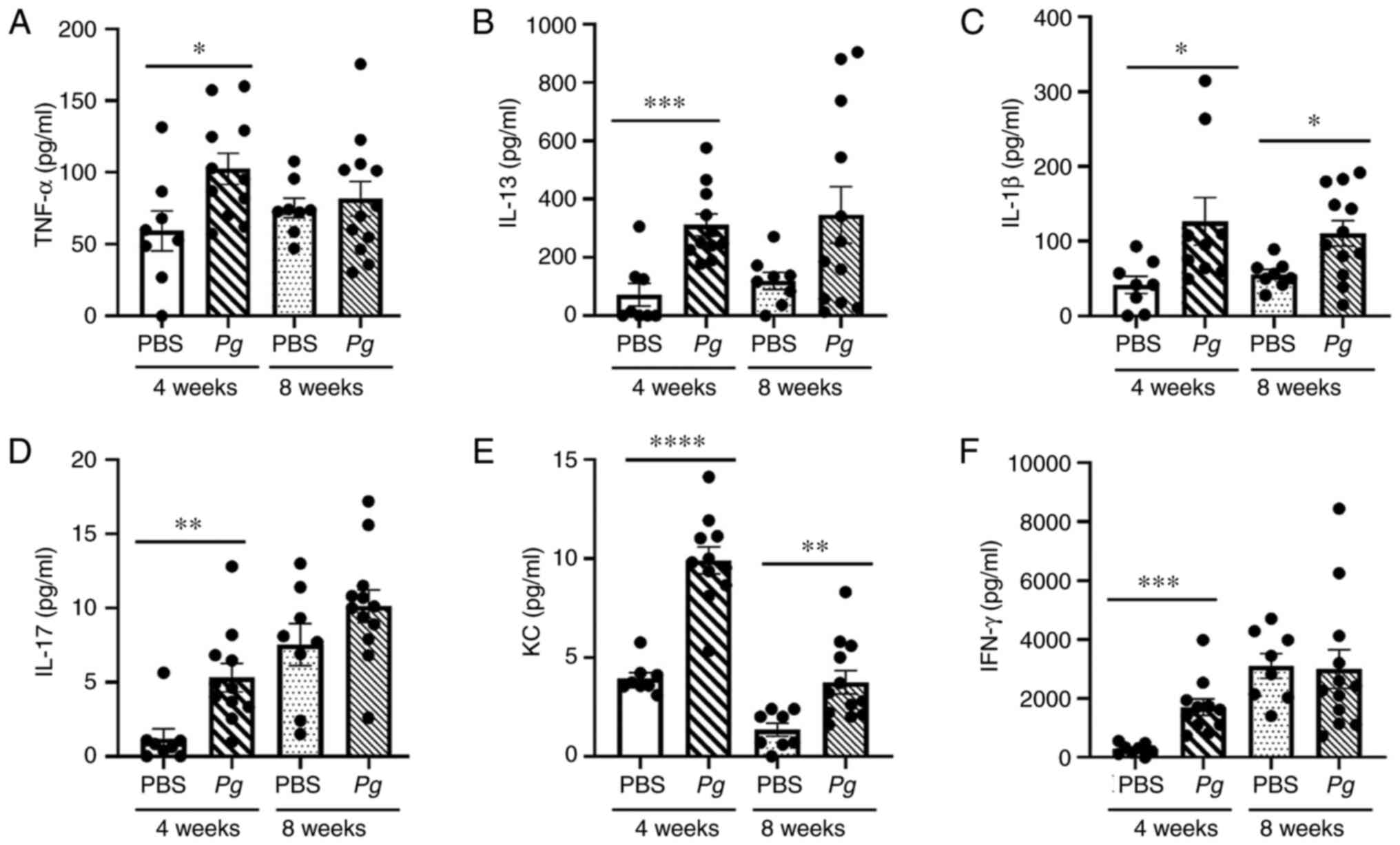 | Figure 4Topical Pg application
directly into the gingival pocket induces systemic inflammation.
ELISA measuring the levels (pg/ml) of (A) TNF-α, (B) IL-13, (C)
IL-1β, (D) IL-17, (E) KC, and (F) IFN-γ from mouse sera at 4 and 8
weeks after the final PBS or Pg inoculation.
*P<0.05, **P<0.01,
***P<0.005 and ****P<0.001 determined
using one-way ANOVA. Results represent the mean ± SEM; n=8–12 mice
per group. Each dot represents a result from 1 mouse. Pg,
Porphyromonas gingivalis; IL, interleukin; TNF-α, tumor
necrosis factor α; KC, keratinocyte chemoattractant; IFN-γ,
interferon γ. |
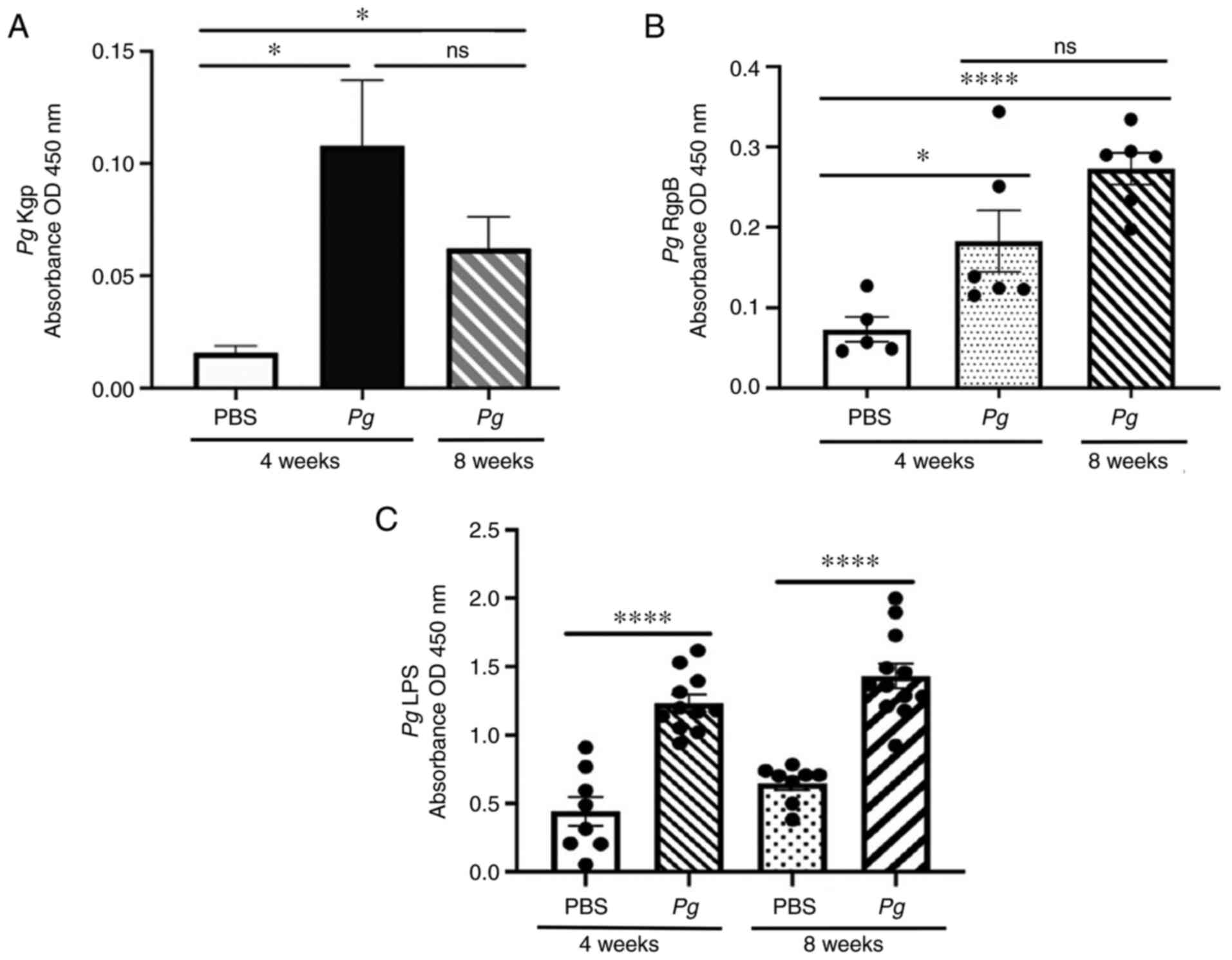
Anti-Pg gingipain and anti-Pg LPS
antibodies detected in the sera of mice receiving the topical Pg
application into the gingival pocket
Anti-Pg gingipain and Pg LPS
antibodies in the blood were measured using Pg gingipain
(Kgp and RgpB) and Pg LPS as probes using an ELISA-based
assay. Both anti-gingipain and anti-LPS antibodies were detected in
sera of mice receiving the topical Pg application; however,
no such antibodies (or basal level) were detected in mice receiving
the topical PBS application (Fig.
5).
Pg gingipain and LPS are identified in
the gingival tissues of mice receiving the topical application of
Pg into the gingival pocket
Gingipains and LPS are crucial virulent factors
released in both soluble mediators and in OMVs by Pg, and
they are frequently found in tissues (2,3).
In the present study, to detect whether these proteins are present
in the gingival tissue of mice receiving the topical Pg
application, recombinant proteins for gingipain (Kgp and RgpB) were
generated and used to raise antibodies against them (Fig. S3). For LPS, the commercially
available antibody was used. Immunofluorescence staining using
these antibodies revealed that both gingipain and LPS were detected
at the site at which Pg was inoculated (Fig. 6). These data indicate that local
Pg LPS and gingipains may, in part, be responsible for the
gingival inflammation.
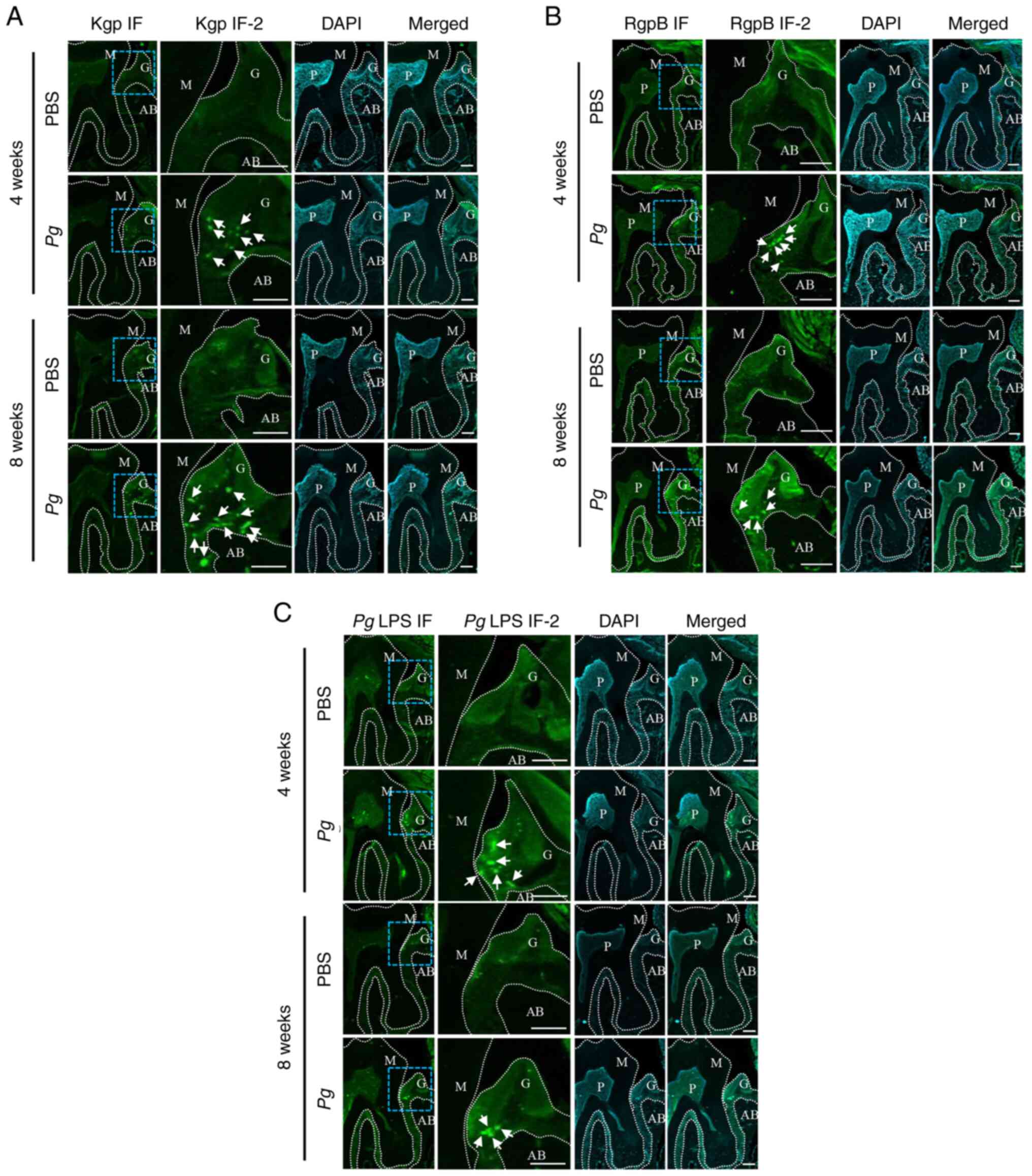 | Figure 6Pg gingipains and LPS
aggregates are present in the gingival tissues of mice receiving
the topical Pg inoculation into the gingival pocket. (A) IF
analysis using Kgp. PBS and Pg were from the mice sacrificed
at 4 or 8 weeks after the final PBS or Pg inoculation. Kgp
IF-2 is the higher-magnified image of blue-dotted square in Kgp IF.
Kgp (bright green color dots, arrows) was present only in the
gingival tissue of mouse receiving topical Pg not in those
receiving PBS. (B) IF analysis using RgpB. PBS and Pg were
from the mice sacrificed at 4 or 8 weeks after the final PBS or
Pg inoculation. RgpB IF-2 is the higher-magnified image of
blue-dotted square in RgpB IF. The RgpB (bright green color dots,
arrows) was present only in the gingival tissue of mouse receiving
topical Pg not in those receiving PBS. (C) IF analysis using
Pg LPS. PBS and Pg were from the mice sacrificed at 4
or 8 weeks after the final PBS or Pg inoculation. Pg
LPS -2 is the higher-magnified image of blue-dotted square in
Pg LPS IF. The Pg LPS (bright green color dots,
arrows) was present only in the gingival tissue of mouse receiving
topical Pg not in those receiving PBS. DAPI was heavily
stained in pulpal and gingival tissues. Scale bars, 100 μm.
M, upper second molar; G, gingival tissue; AB, alveolar bone; P,
pulpal tissue; Pg, Porphyromonas gingivalis; IF,
immunofluorescence; LPS, lipopolysaccharide; Kgp, lysine-gingipain;
RgpB, arginine-gingipain B. |
Pg gingipain and Pg LPS are found in the
aortic roots of mice receiving the topical Pg application directly
into the gingival pocket
The authors have previously reported that
ligature-induced periodontitis causes the exacerbation of
atherosclerotic lesions in ApoE-deficient mice (6,24). In the present study, to evaluate
the status of the atherosclerosis in these wild-type mice, the
hearts and the aortic roots were harvested together. No changes
were observed in the histological features of lipid deposition that
are indicative of atherosclerosis (Fig. S4). Subsequently, the aortic
roots were further examined for evidence of Pg gingipains
and Pg LPS aggregates. Immunofluorescence staining revealed
distinct focal signals of both Pg gingipains and Pg
LPS in the aortic walls of mice receiving the topical Pg
application (Fig. 7; the second
and fourth rows in each panel). By contrast, there were no signals
in the aortic roots of mice receiving the topical PBS application
(Fig. 7; the first and third
rows in each panel).
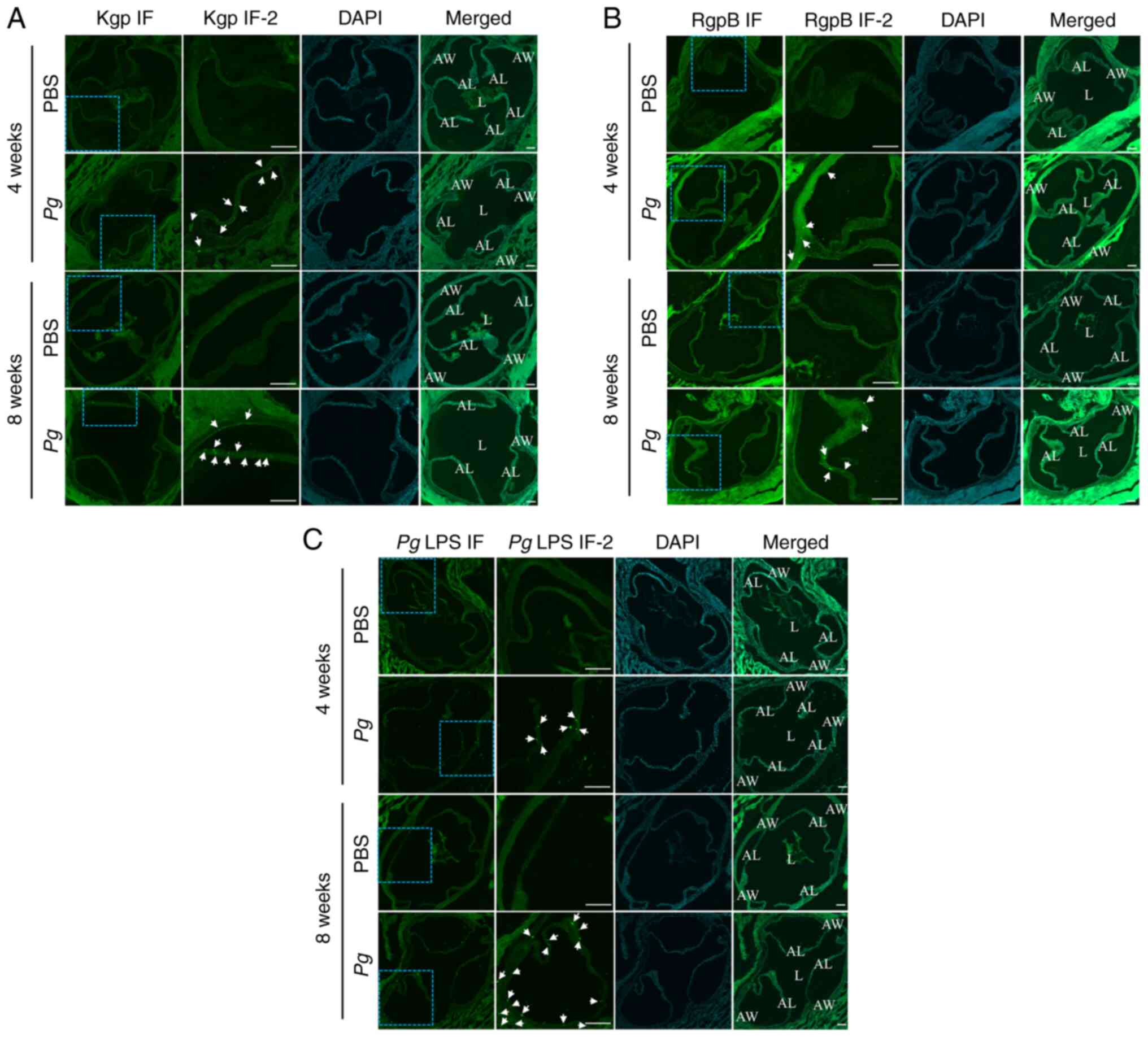 | Figure 7Pg gingipain and Pg LPS
aggregates are found in the aortic roots of mice receiving
Pg inoculation into the gingival pocket. (A) IF analysis
using Kgp. PBS and Pg were from the mice sacrificed at 4 or
8 weeks after the final PBS or Pg inoculation. Kgp IF-2 is
the higher-magnified image of blue-dotted square in Kgp IF. The
RgpB (bright green color dots, arrows) was present only in the
aortic leaflet and aortic wall of mouse receiving topical
Pg, not in those receiving PBS. (B) IF analysis using RgpB.
PBS and Pg were from the mice sacrificed at 4 or 8 weeks
after the final PBS or Pg inoculation. RgpB IF-2 is the
higher-magnified image of blue-dotted square in RgpB IF. The RgpB
(bright green color dots, arrows) was present only in the aortic
leaflet and aortic wall of mouse receiving topical Pg, not
in those receiving PBS. (C) IF analysis using Pg LPS. PBS
and Pg were from the mice sacrificed at 4 or 8 weeks after
the final PBS or Pg inoculation. Pg LPS -2 is the
higher-magnified image of blue-dotted square in Pg LPS IF.
The LPS (bright green color dots, arrows) was present only in the
aortic leaflet and aortic wall of mouse receiving topical
Pg, not in those receiving PBS. Nuclei of cells were
localized with DAPI staining. Scale bars, 100 μm. L, lumen;
AW, aortic wall; AL, aortic leaflet; Pg, Porphyromonas
gingivalis; IF, immunofluorescence; LPS, lipopolysaccharide;
Kgp, lysine-gingipain; RgpB, arginine-gingipain B. |
Discussion
To date, to the best of our knowledge, no animal
models have been reported that faithfully mimic the clinical
settings in which perio-pathogens, including Pg colonize
around a tooth. In the present study, a mouse model of Pg
colonization was successfully established, in which Pg was
topically and directly applied into an artificially created
gingival pocket. This was able to induce more chronic active
periodontitis and other systemic effects, in part due to the
prolonged colonization of Pg around the tooth.
Mice are not the natural host of Pg (29). For this reason, several methods
have been developed to mimic a human Pg associated
periodontal disease state in mice, including the inoculation of
live bacteria into the oral cavity (17). Although this method has been
widely used to study and demonstrate the local and systemic effects
of Pg (30,31), its general application to the
whole oral cavity and the use of the high concentrations and
exhaustive repeated applications (3–4x/week over a period of 8–12
weeks) without causing drastic alveolar bone loss, renders it
difficult to examine the sole effects of bacterial colonization
around the tooth to the local and systemic outcomes. The model
established herein is distinct from previously reported models, in
that the ligature was only used for 1 week to create gingival
pocket, after which the ligature was removed and Pg was
locally and topically applied on one tooth for 5 weeks. Within this
relatively short period of time, bacterial colonization was
efficiently established and conferred local effects by causing
alveolar bone loss on the applied tooth only, as well as systemic
effects, as demonstrated by the presence of Pg pathogen in
the vascular system.
One of the major concerns with the conventional
periodontitis model (e.g., oral inoculation) is a potential
systemic effect via the gastrointestinal (GI) tract by swallowing
the inoculated bacteria. In the model in the present study,
although an attempt was made to apply Pg locally in a small
volume, this potential still exists as locally administered
Pg can be subsequently swallowed by the mouse. On the other
hand, knowing this shortcoming, this mouse model was developed
strategically by creating a gingival pocket with a ligature (to
maximize the administration and colonization of Pg locally)
and topically applying Pg for only 5 weeks (to allow the
colonization of bacteria around the tooth), and leaving the mice
for an additional 4 and 8 weeks without any further Pg
application (to allow the clearance of bacteria from the GI tract
and systemic circulation as a direct result of Pg
inoculation). It was reasoned that terminating the oral inoculation
of bacteria and leaving the mice for an additional 4 or 8 weeks
would allow a sufficient amount of time for bacterial ingestion via
the GI tract to be reduced and to maximize bacterial colonization
on the tooth surface. Indeed, it has been reported that the average
half-life of bacteria clearance from blood is 2–10 min in mice
(32). Therefore, it is
conceivable that the systemic effects that were observed in the
present study may primarily be derived from the colonized Pg
around the tooth through the gingival tissues, rather than by the
accidental swallowing of the inoculated bacteria via the GI
tract.
It is noteworthy that not all mice inoculated with
Pg topically developed colonies around the second molar
tooth. Indeed, the present study demonstrated that 45–70% of mice
established Pg colonies around the second molars (Fig. 1). As this was a pilot study, it
is conceivable that increasing the numbers of adjacent molar teeth
would increase the number of positive animals and may reflect even
better the human oral condition. The detected amount of
Pg/swab/mouse was higher at 4 weeks (~150,000–210,000 CFU)
when compared to that at 8 weeks (~15,000–55,000 CFU). The higher
numbers observed at 4 weeks may be the result of carryover from the
final inoculation of the bacterial colonies. Alternatively, the
lower numbers observed at 8 weeks may be due to increasing host
defense responses (e.g., anti-Pg antibodies toward Pg
due to the chronic infection status). Collectively, these data
indicate that the establishment of Pg colonies even in one
second molar appears to be directly linked to the ongoing more
chronic colonization and resulting systemic virulent effects of
Pg in the mice.
It is noteworthy that even though only a small
number of teeth were inoculated and colonized, systemic
manifestations were readily observed, suggesting a relatively
potent inflammatory capacity of Pg for the murine host as
observed herein. Moreover, given the relatively limited number of
total bacteria in the oral cavity, the observance of LPS and
gingipains at distant tissue sites (aortic tissues) may suggest
again that these bacterial toxins are readily transported
presumably via OMVs originating in the gums. Multiple studies have
concluded similar findings. Recently, He et al (33) demonstrated in a mouse model that
released OMVs from Pg exerted potent cytotoxic effects on
lung epithelial cells; Pg OMVs revealed their ability to
induce the apoptosis of lung epithelial cells and disrupt the
epithelial barrier system. They concluded from their experimental
design that these observations suggest that OMVs deliver their
pathogenic factors from the oral cavity to respiratory organs
without the direct translocation of Pg itself (33). O'Brien-Simpson et al
(34) similarly demonstrated
that Pg-infected cells and the RgpA-Kgp OMV complexes at low
concentrations stimulated secretory intercellular adhesion molecule
1, IL-8, IL-6 and MCP secretion from cultured human epithelial (KB)
and fibroblast (MRC-5) cells. However, at high concentrations, a
reduction in the level of these mediators was observed (34). By contrast, macrophage
inflammatory protein 1 and IL-1 were stimulated only at high
Pg cell concentrations. Pg-infected cells and the
RgpA-Kgp OMV complexes were shown to induce the apoptosis of KB and
MRC-5 cells in a time- and concentration-dependent manner (34). These data suggest that the
RgpA-Kgp complexes penetrate the gingival connective tissue; at low
concentrations distal from the plaque, the complexes stimulate the
secretion of pro-inflammatory mediators, while at high
concentrations proximal to the plaque, they induce apoptosis and
attenuate the secretion of pro-inflammatory mediators.
Previously, it was demonstrated that ligature
placement in the gingival pocket enhanced expression of
pro-inflammatory cytokines in both the local gingiva, aortic tissue
and serum, and that Pg. LPS further exacerbated these
expression levels in ApoE-deficient mice (6,24). Pg LPS, the major
glycolipids present at the surface of the Pg, is one of the
virulent factors in Pg. It has recently been demonstrated
that Pg LPS weakly induces pro-inflammatory cytokine
production in mice by activating Toll-like receptor 4 (35). Similarly, the present study found
that the local inoculation of Pg induced the expression of
pro-inflammatory cytokines even in wild-type mice (Figs. 3 and 4). This is also in line with a previous
report in which both wild-type and ApoE-deficient mice
exhibited an increased expression of pro-inflammatory cytokines in
serum when challenged by Pg via the oral cavity (36).
Macrophages are important immune cells that are
known to be associated with atherosclerosis. In particular,
macrophages produce TNF-α that alters the phenotypes of vascular
smooth muscle cells and contributes to the development of
atherosclerosis (37).
TNF-α-producing M1 macrophages are crucial producers of
pro-inflammatory cytokines and are activated by stimuli, such as
LPS (38). Therefore, the
prolonged production of LPS from locally accumulated Pg may promote
macrophages to undergo M1 polarization, affecting the local
diseases (e.g., periodontitis) and the systemic diseases (e.g.,
atherosclerosis).
In the mouse model in the present study, Pg
LPS and gingipains were detected in the gingival tissues (Fig. 6C) and the aortic walls (Fig. 7C). Such findings as discussed
above suggest that Pg OMVs containing these toxins may have
penetrated through the epithelial barriers and reside in the
gingival tissues as well as in the blood stream. Indeed, a previous
study demonstrated that Pg OMVs and occasionally PG DNA was
found to be localized into squamous epithelium and capillary
endothelium in patients with periodontitis (39). Pg DNA was also detected in
the aortic walls (40–42). These results however, are
complicated by the fact that Pg, similar to several other
Gram-negative bacterial OMVs carry numerous forms of both
functional RNA and DNA transcripts. Without controlling for a more
direct means of detecting the whole bacterial cell, one cannot
conclude what the source of the DNA is in these tissues. It is
becoming clearer that Pg OMV, as well as other species OMVs,
such H. Pylori in which a cell modulating and transforming
cell factor CagA are found circulating in human serum samples of
infected patients' chronic gastritis (43).
The rather limited topical application of Pg
into the gingival pocket of the second molars appears to have led
to a more chronic active colonization, possibly inducing the
secretion of free bacterial toxins or in the form of OMVs
containing gingipains and LPS into the blood or lymphatics. It
cannot be discounted that a planktonic form of Pg is
released from the bacterial colony in the sulcus with Pg
penetration/transmigration through the gingival epithelial tissues
and the aortic endothelial layers and/or through a 'Trojan Horse'
cellular macrophage mechanism.
Pg gingipains are cysteine proteases that
assert their virulent effects by degrading the extracellular matrix
(7–9). It has been demonstrated that
Pg gingipains disrupt epithelial barrier functions by
degrading JAM1, a tight junction-associated protein, leading to
Pg penetration into the tissues (44). Pg produces three different
types of gingipains: Arginine-gingipain A (RgpA),
arginine-gingipain B (RgpB) and lysine-gingipain (Kgp) (7). Herein, by generating recombinant
proteins, antibodies against RgpB and Kgp were developed (Fig. S3). Using these antibodies,
Pg gingipains were detected in the gingival tissues
(Fig. 6A and B). Of note,
anti-Pg gingipain and LPS antibodies were detected in the
serum (Fig. 5A and B), while
Pg gingipains and LPS were also found in the aortic walls
(Fig. 7). Such finding suggests
that the effects of Pg gingipains and LPS are at both local
and systemic levels, asserting ECM-degrading functions and
assisting in Pg penetrations into different parts of the
body, including the aorta. Alternatively, it is possible that the
OMV containing LPS can be released from the Pg,
transmigrating from the gingival sulcus through the blood to
proximal tissue organs, such as the aorta.
Wild-type mice do not develop lipid deposition at
the intercostal arteries, at the junction of the aorta to the
heart, unless fed a high-fat diet for prolonged period of time
(45). Even with the high-fat
diet in mice with an ApoE-deficient background, the
development of atherosclerosis is attenuated in the presence of
statins (24,46–49). As such, it is highly probable
that additional factors, such as high serum lipid levels are
required in order to create an inflammatory environment that can
drive atherosclerosis development in the aortic areas.
Additionally, it is known that pro-inflammatory responses are
highly locally regulated and thus may be inhibited to assure
intra-cellular survival.
As demonstrated herein, while anti-Pg LPS
and anti-Pg RgpB antibodies were progressively present at
higher levels at 8 weeks as compared with 4 weeks, the
anti-Pg Kgp antibody level was decreased at 8 weeks,
although the difference was not statistically significant (Fig. 5). However, in a murine lesion
model, O'Brien-Simpson et al (8) demonstrated that the virulence of
Kgp was more significant when compared to that of RgpB, suggesting
that the potency of these two gingipains was functionally
different. RgpB and Kgp are encoded from different genes, and they
are also post-translationally regulated (50–52). As such, it would of interest to
further examine their regulations and functions as Pg
colonizes around the tooth in a time-dependent manner; this may
more easily studied using the model presented herein.
The accumulated Pg gingipain/LPS aggregates
in the arterial wall are known to be associated with the
development of atherosclerosis (53). It is highly probable that
anti-gingipain antibodies are associated with human periodontitis
and may be important for the control of periodontitis. While
targeting gingipains may provide some therapeutic improvement,
targeting the whole bacteria with a precision biological such as
monoclonal antibody would be significantly more efficacious, as it
would result in the complete cessation of all bacterial toxins from
the local and systemic circulation and would possibly allow for the
re-establishment of a more normal oral microbiome.
In conclusion, the present study established the
development in vivo of a chronic Pg periodontal
infection/colonization of a wild-type mouse model that is more
bacteriologically similar to that of the human condition. The
advantages of this animal model include: i) Demonstration of the
proof-of-principle that topically applied Pg bacteria into
the small artificially created gingival pocket leads to a
pro-longed bacterial colonization around the tooth and induces
subsequent local and systemic inflammatory responses; ii) the
establishment of a mouse model that can provide a strategy with
which to evaluate the local effects of specific strains of
bacteria, such as Pg; and iii) the utilization of this mouse
model to further study other systemic diseases. In particular, as
elevated cholesterol levels are closely associated with the
development of atherosclerosis and Alzheimer's disease (54,55), the further utilization of this
novel Pg periodontal model applied in ApoE-deficient
mice may prove to be useful for the further examination of the
pathogenesis of these periodontitis-related toxins and systemic
diseases, as well as of the therapeutic efficacy of different
treatment modalities, including Pg monoclonal antibodies and
vaccines.
Supplementary Data
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
NHP, RK and PLN were involved in the
conceptualization of the study. SK, YB, JK, BT and DK performed the
experiments and participated in data analysis. CC and HTT were
responsible for THE Pg culture and gingipain antibody
production. SHL performed the RT-qPCR analyses. SK, YB, RK and NHP
were involved in the discussion and interpretation of the results.
SK, RK, PN, and NHP drafted the manuscript. SK, YB, JK and CC
confirm the authenticity of all the raw data. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
All procedures were performed in compliance with
the institution's policy and applicable provisions of the United
States Department of Agriculture (USDA) Animal Welfare Act
Regulations and the Public Health Service (PHS) Policy. The
experimental protocols were approved by the Animal Research
Committee (ARC) of the University of California, Los Angeles (UCLA)
under ARC# 2019-057.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
The authors would like to thank Dr Marc Penn and Dr
Dan Sindelar from Keystone Bio Inc for their critical reading of
the manuscript and for providing constructive criticisms.
Funding
The present study was supported in part by the research funds
awarded from the UCLA Chancellor's Office and Keystone Bio Inc. and
the National Institute of Dental and Craniofacial Research
(NIDCR)/NIH under the Award Number DE026758.
References
|
1
|
Darveau RP: Periodontitis: A polymicrobial
disruption of host homeostasis. Nat Rev Microbiol. 8:481–490. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hajishengallis G, Wang M and Liang S:
Induction of distinct TLR2-mediated proinflammatory and proadhesive
signaling pathways in response to porphyromonas gingivalis
fimbriae. J Immunol. 182:6690–6696. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Takii R, Kadowaki T, Baba A, Tsukuba T and
Yamamoto K: A functional virulence complex composed of gingipains,
adhesins, and lipopolysaccharide shows high affinity to host cells
and matrix proteins and escapes recognition by host immune systems.
Infect Immun. 73:883–893. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hiyari S, Atti E, Camargo PM, Eskin E,
Lusis AJ, Tetradis S and Pirih FQ: Heritability of periodontal bone
loss in mice. J Periodontal Res. 50:730–736. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu R, Desta T, Raptis M, Darveau RP and
Graves DT: P. gingivalis and E. coli lipopolysaccharides exhibit
different systemic but similar local induction of inflammatory
markers. J Periodontol. 79:1241–1247. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Suh JS, Kim S, Boström KI, Wang CY, Kim RH
and Park NH: Periodontitis-induced systemic inflammation
exacerbates atherosclerosis partly via endothelial-mesenchymal
transition in mice. Int J Oral Sci. 11:212019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Imamura T: The role of gingipains in the
pathogenesis of periodontal disease. J Periodontol. 74:111–118.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
O'Brien-Simpson NM, Paolini RA, Hoffmann
B, Slakeski N, Dashper SG and Reynolds EC: Role of RgpA, RgpB, and
Kgp proteinases in virulence of Porphyromonas gingivalis W50 in a
murine lesion model. Infect Immun. 69:7527–7534. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pike RN, Potempa J, McGraw W, Coetzer TH
and Travis J: Characterization of the binding activities of
proteinase-adhesin complexes from Porphyromonas gingivalis. J
Bacteriol. 178:2876–2882. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ding PH, Yang MX, Wang NN, Jin LJ, Dong Y,
Cai X and Chen LL: Porphyromonas gingivalis-induced NLRP3
inflammasome activation and its downstream interleukin-1beta
release depend on caspase-4. Front Microbiol. 11:18812020.
View Article : Google Scholar
|
|
11
|
Fitzpatrick RE, Wijeyewickrema LC and Pike
RN: The gingipains: Scissors and glue of the periodontal pathogen,
Porphyromonas gingivalis. Future Microbiol. 4:471–487. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gui MJ, Dashper SG, Slakeski N, Chen YY
and Reynolds EC: Spheres of influence: Porphyromonas gingivalis
outer membrane vesicles. Mol Oral Microbiol. 31:365–378. 2016.
View Article : Google Scholar
|
|
13
|
Guo Y, Nguyen KA and Potempa J: Dichotomy
of gingipains action as virulence factors: From cleaving substrates
with the precision of a surgeon's knife to a meat chopper-like
brutal degradation of proteins. Periodontol 2000. 54:15–44. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Grenier D, Roy S, Chandad F, Plamondon P,
Yoshioka M, Nakayama K and Mayrand D: Effect of inactivation of the
Arg- and/or Lys-gingipain gene on selected virulence and
physiological properties of porphyromonas gingivalis. Infect Immun.
71:4742–4748. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lalla E, Lamster IB, Feit M, Huang L and
Schmidt AM: A murine model of accelerated periodontal disease in
diabetes. J Periodontal Res. 33:387–399. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Graves DT, Kang J, Andriankaja O, Wada K
and Rossa C Jr: Animal models to study host-bacteria interactions
involved in periodontitis. Front Oral Biol. 15:117–132. 2012.
View Article : Google Scholar
|
|
17
|
Baker PJ, Evans RT and Roopenian DC: Oral
infection with Porphyromonas gingivalis and induced alveolar bone
loss in immunocompetent and severe combined immunodeficient mice.
Arch Oral Biol. 39:1035–1040. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bainbridge B, Verma RK, Eastman C, Yehia
B, Rivera M, Moffatt C, Bhattacharyya I, Lamont RJ and Kesavalu L:
Role of Porphyromonas gingivalis phosphoserine phosphatase enzyme
SerB in inflammation, immune response, and induction of alveolar
bone resorption in rats. Infect Immun. 78:4560–4569. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Baker PJ, Dixon M, Evans RT and Roopenian
DC: Heterogeneity of Porphyromonas gingivalis strains in the
induction of alveolar bone loss in mice. Oral Microbiol Immunol.
15:27–32. 2000. View Article : Google Scholar
|
|
20
|
Dumitrescu AL, Abd-El-Aleem S, Morales-Aza
B and Donaldson LF: A model of periodontitis in the rat: Effect of
lipopolysaccharide on bone resorption, osteoclast activity, and
local peptidergic innervation. J Clin Periodontol. 31:596–603.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nishida E, Hara Y, Kaneko T, Ikeda Y, Ukai
T and Kato I: Bone resorption and local interleukin-1alpha and
interleukin-1beta synthesis induced by Actinobacillus
actinomycetemcomitans and Porphyromonas gingivalis
lipopolysaccharide. J Periodontal Res. 36:1–8. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Marchesan J, Girnary MS, Jing L, Miao MZ,
Zhang S, Sun L, Morelli T, Schoenfisch MH, Inohara N, Offenbacher S
and Jiao Y: An experimental murine model to study periodontitis.
Nat Protoc. 13:2247–2267. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kimura S, Nagai A, Onitsuka T, Koga T,
Fujiwara T, Kaya H and Hamada S: Induction of experimental
periodontitis in mice with Porphyromonas gingivalis-adhered
ligatures. J Periodontol. 71:1167–1173. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Suh JS, Lee SH, Fouladian Z, Lee JY, Kim
T, Kang MK, Lusis AJ, Boström KI, Kim RH and Park NH: Rosuvastatin
prevents the exacerbation of atherosclerosis in ligature-induced
periodontal disease mouse model. Sci Rep. 10:63832020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Belanger M, Rodrigues P and Progulske-Fox
A: Genetic manipulation of porphyromonas gingivalis. Curr Protoc
Microbiol. Chapter 13: Unit13C.2. 2007. View Article : Google Scholar
|
|
26
|
Velsko IM, Chukkapalli SS, Rivera MF, Lee
JY, Chen H, Zheng D, Bhattacharyya I, Gangula PR, Lucas AR and
Kesavalu L: Active invasion of oral and aortic tissues by
Porphyromonas gingivalis in mice causally links periodontitis and
atherosclerosis. PLoS One. 9:e978112014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Siegel SD, Amer BR, Wu C, Sawaya MR,
Gosschalk JE, Clubb RT and Ton-That H: Structure and mechanism of
LcpA, a phosphotransferase that mediates glycosylation of a
gram-positive bacterial cell wall-anchored protein. mBio.
10:e01580–01518. 2019.PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
29
|
Hayashi C, Gudino CV, Gibson FC III and
Genco CA: Review: Pathogen-induced inflammation at sites distant
from oral infection: bacterial persistence and induction of
cell-specific innate immune inflammatory pathways. Mol Oral
Microbiol. 25:305–316. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gibson FC III, Hong C, Chou HH, Yumoto H,
Chen J, Lien E, Wong J and Genco CA: Innate immune recognition of
invasive bacteria accelerates atherosclerosis in apolipoprotein
E-deficient mice. Circulation. 109:2801–2806. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lalla E, Lamster IB, Hofmann MA,
Bucciarelli L, Jerud AP, Tucker S, Lu Y, Papapanou PN and Schmidt
AM: Oral infection with a periodontal pathogen accelerates early
atherosclerosis in apolipoprotein E-null mice. Arterioscler Thromb
Vasc Biol. 23:1405–1411. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Harrington WN, Nolan J, Nedosekin DA,
Smeltzer MS and Zharov VP: Real-time monitoring of bacteria
clearance from blood in a murine model. Cytometry A. 97:706–712.
2020. View Article : Google Scholar
|
|
33
|
He Y, Shiotsu N, Uchida-Fukuhara Y, Guo J,
Weng Y, Ikegame M, Wang Z, Ono K, Kamioka H, Torii Y, et al: Outer
membrane vesicles derived from Porphyromonas gingivalis induced
cell death with disruption of tight junctions in human lung
epithelial cells. Arch Oral Biol. 118:1048412020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
O'Brien-Simpson NM, Pathirana RD, Walker
GD and Reynolds EC: Porphyromonas gingivalis RgpA-Kgp
proteinase-adhesin complexes penetrate gingival tissue and induce
proinflammatory cytokines or apoptosis in a concentration-dependent
manner. Infect Immun. 77:1246–1261. 2009. View Article : Google Scholar :
|
|
35
|
Nativel B, Couret D, Giraud P, Meilhac O,
d'Hellencourt CL, Viranaïcken W and Silva CRD: Porphyromonas
gingivalis lipopolysaccharides act exclusively through TLR4 with a
resilience between mouse and human. Sci Rep. 7:157892017.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Maekawa T, Takahashi N, Tabeta K, Aoki Y,
Miyashita H, Miyauchi S, Miyazawa H, Nakajima T and Yamazaki K:
Chronic oral infection with Porphyromonas gingivalis accelerates
atheroma formation by shifting the lipid profile. PLoS One.
6:e202402011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Parameswaran N and Patial S: Tumor
necrosis factor-α signaling in macrophages. Crit Rev Eukaryot Gene
Expr. 20:87–103. 2010. View Article : Google Scholar
|
|
38
|
Yunna C, Mengru H, Lei W and Weidong C:
Macrophage M1/M2 polarization. Eur J Pharmacol. 877:1730902020.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rajakaruna GA, Negi M, Uchida K, Sekine M,
Furukawa A, Ito T, Kobayashi D, Suzuki Y, Akashi T, Umeda M, et al:
Localization and density of porphyromonas gingivalis and tannerella
forsythia in gingival and subgingival granulation tissues affected
by chronic or aggressive periodontitis. Sci Rep. 8:95072018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Delbosc S, Alsac JM, Journe C, Louedec L,
Castier Y, Bonnaure-Mallet M, Ruimy R, Rossignol P, Bouchard P,
Michel JB and Meilhac O: Porphyromonas gingivalis participates in
pathogenesis of human abdominal aortic aneurysm by neutrophil
activation. Proof of concept in rats. PLoS One. 6:e186792011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nakano K, Nemoto H, Nomura R, Inaba H,
Yoshioka H, Taniguchi K, Amano A and Ooshima T: Detection of oral
bacteria in cardiovascular specimens. Oral Microbiol Immunol.
24:64–68. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kurihara N, Inoue Y, Iwai T, Umeda M,
Huang Y and Ishikawa I: Detection and localization of
periodontopathic bacteria in abdominal aortic aneurysms. Eur J Vasc
Endovasc Surg. 28:553–558. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shimoda A, Ueda K, Nishiumi S,
Murata-Kamiya N, Mukai SA, Sawada SI, Azuma T, Hatakeyama M and
Akiyoshi K: Exosomes as nanocarriers for systemic delivery of the
helicobacter pylori virulence factor CagA. Sci Rep. 6:183462016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Takeuchi H, Sasaki N, Yamaga S, Kuboniwa
M, Matsusaki M and Amano A: Porphyromonas gingivalis induces
penetration of lipopolysaccharide and peptidoglycan through the
gingival epithelium via degradation of junctional adhesion molecule
1. PLoS Pathog. 15:e10081242019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Paigen B, Morrow A, Holmes PA, Mitchell D
and Williams RA: Quantitative assessment of atherosclerotic lesions
in mice. Atherosclerosis. 68:231–240. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Johnston TP, Baker JC, Hall D, Jamal S,
Palmer WK and Emeson EE: Regression of poloxamer 407-induced
atherosclerotic lesions in C57BL/6 mice using atorvastatin.
Atherosclerosis. 149:303–313. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Johnston TP, Nguyen LB, Chu WA and Shefer
S: Potency of select statin drugs in a new mouse model of
hyperlipidemia and atherosclerosis. Int J Pharm. 229:75–86. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kleemann R, Princen HM, Emeis JJ, Jukema
JW, Fontijn RD, Horrevoets AJG, Kooistra T and Havekes LM:
Rosuvastatin reduces atherosclerosis development beyond and
independent of its plasma cholesterol-lowering effect in
APOE*3-Leiden transgenic mice: Evidence for antiinflammatory
effects of rosuvastatin. Circulation. 108:1368–1374. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Park KY and Heo TH: Combination therapy
with cilostazol and pravastatin improves antiatherogenic effects in
low-density lipoprotein receptor knockout mice. Cardiovasc Ther.
36:e124762018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Potempa J, Banbula A and Travis J: Role of
bacterial proteinases in matrix destruction and modulation of host
responses. Periodontol 2000. 24:153–192. 2000. View Article : Google Scholar
|
|
51
|
Pavloff N, Pemberton PA, Potempa J, Chen
WC, Pike RN, Prochazka V, Kiefer MC, Travis J and Barr PJ:
Molecular cloning and characterization of Porphyromonas gingivalis
lysine-specific gingipain. A new member of an emerging family of
pathogenic bacterial cysteine proteinases. J Biol Chem.
272:1595–1600. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pavloff N, Potempa J, Pike RN, Prochazka
V, Kiefer MC, Travis J and Barr PJ: Molecular cloning and
structural characterization of the Arg-gingipain proteinase of
Porphyromonas gingivalis. Biosynthesis as a proteinase-adhesin
polyprotein. J Biol Chem. 270:1007–1010. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Hashimoto M, Kadowaki T, Tsukuba T and
Yamamoto K: Selective proteolysis of apolipoprotein B-100 by
Arg-gingipain mediates atherosclerosis progression accelerated by
bacterial exposure. J Biochem. 140:713–723. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Shepardson NE, Shankar GM and Selkoe DJ:
Cholesterol level and statin use in Alzheimer disease: I. Review of
epidemiological and preclinical studies. Arch Neurol. 68:1239–1244.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Vaya J and Schipper HM: Oxysterols,
cholesterol homeostasis, and Alzheimer disease. J Neurochem.
102:1727–1737. 2007. View Article : Google Scholar : PubMed/NCBI
|