Introduction
The tumor necrosis factor (TNF) superfamily consists
of numerous cytokine ligands and receptors that regulate many
biological processes, including cell proliferation and survival,
and their balance is important in maintaining normal cellular
functions (1,2).
TNF superfamily ligands are mostly expressed as type
II transmembrane proteins that can be processed into smaller
soluble proteins exerting a wide variety of biological effects by
binding their cognate family of TNF receptors (1,2).
Tumor necrosis factor-like weak inducer of apoptosis (TWEAK), a
member of the tumor necrosis factor superfamily, was first
described as an inducer of apoptosis in transformed cell lines
(3). It has later become apparent
that TWEAK is a multifunctional cytokine that regulates survival
(4), proliferation (5–7),
migration (6,8,9),
differentiation (10,11), and induces apoptosis in certain
tumor cell lines (12,13). TWEAK acts on responsive cells via
binding to an inducible high affinity cell surface receptor known
as fibroblast growth factor-inducible 14 (Fn14) (14). It has been demonstrated that TWEAK
binding to Fn14, or constitutive Fn14 overexpression, activates
nuclear factor κB (NF-κB) signaling pathway, which is known to play
a pivotal role in immune- and inflammatory processes, oncogenesis
and cancer therapy resistance (15,16).
The aberrant expression of both TWEAK and Fn14 has
been detected in human tumor samples (7,17),
and the possibility that TWEAK and Fn14 could contribute to tumor
growth has recently been reviewed (16,18,19).
TWEAK activates cellular processes attributed to tumor growth such
as matrix metalloprotease (MMP) secretion (20–22),
migration (6,8,9),
proliferation (5–7), and apoptotic resistance (4). TWEAK is also mitogenic for
endothelial cells and stimulates angiogenesis (5,23).
Neuroblastoma, an embryonic tumor of the sympathetic
nervous system, is the most common and deadly extra-cranial tumor
associated with childhood. Neuroblastoma exhibits heterogeneous
biological and clinical features ranging from spontaneous
regression to highly malignant disease with metastatic spread. More
than 40% of children with neuroblastoma are diagnosed as high-risk
patients and despite intensive treatment modalities; the cure rate
for these patients is less than 50% (24). Hence, there is a great need for new
therapies based on a biological understanding of this disease.
The aim of this study was to assess the expression
of TWEAK and Fn14 in childhood neuroblastoma, particularly how it
relates to the functional importance in neuroblastoma cell growth
and survival.
Materials and methods
Reagents and antibodies
Phospho NF-κB p65 (Ser536; cat. 3031), 65 kDa, NF-κB
p65 (cat. 3034) 65 kDa and Fn14 polyclonal antibody were purchased
from Cell Signaling (Beverly, MA, USA). NF-κB p65 (sc-109), TWEAK
(FL-249), and TWEAK (S-20) antibodies were purchased from Santa
Cruz Biotechnology (Santa Cruz, CA, USA). Secondary anti-goat Alexa
594 and anti-rabbit Alexa 488 antibodies were purchased from
Invitrogen (Carlsbad, CA, USA). Human recombinant IL-1β, TNF-α and
TWEAK were purchased from R&D Systems (Abingdon, UK), and human
TWEAK ELISA was purchased from PeproTech (London, UK).
Human tissue samples
Primary neuroblastoma samples from tumors and
non-malignant adrenals were obtained during surgery, snap-frozen in
liquid nitrogen, and transferred to −80°C for future analysis.
Twenty six neuroblastoma samples derived from children of different
ages and all clinical stages, including different biological
subsets (MYCN amplification, 7 of 27; 1p deletion, 9 of 27;
Table I) were analyzed. Three
childhood ganglioneuromas and three samples of non-malignant
adrenals from children aged 12–25 months were also included.
Ethical approval was obtained from the Karolinska University
Hospital Research Ethics Committee.
 | Table IImmunohistochemical assessment of
TWEAK, Fn14 and pNF-κB expression in neuroblastoma tissue
samples. |
Table I
Immunohistochemical assessment of
TWEAK, Fn14 and pNF-κB expression in neuroblastoma tissue
samples.
Neuroblastoma cell lines
Human neuroblastoma cell lines [SK-N-BE(2), SK-N-DZ, SH-SY5Y, SK-N-SH, SK-N-FI,
IMR-32, SK-N-AS and SHEP-1] were grown in RPMI-1640 medium
supplemented with 10% fetal calf serum (FCS), 2 mM L-glutamine, and
antibiotics. The cultures were kept at 37°C in a humified 5%
CO2 atmosphere.
RNA isolation and RT-PCR
Total RNA was extracted from cultured neuroblastoma
cells using RNeasy Mini kit (cat. no. 74104, Qiagen, Valencia, CA,
USA) according to the manufacturer’s instructions. cDNA was
synthesized using 2.0 μg total RNA that was reverse
transcribed in a final volume of 50 μl using the SuperScript
preamplification kit (Life Technologies, Inc., Gaithersburg, MD,
USA).
Gene specific PCR was performed in 50 μl of
reaction mixture containing 2–10 μl cDNA (from isolated
RNA), 2.5 U of Taq DNA polymerase (Promega, Madison, WI, USA), 10
mM Tris-HCl (pH 8.0), 100 mM KCl, 0.1% Triton® X-100,
2.0 mM MgCl2, 1 mM deoxynucleotide triphosphate mix, and
1 μM of each primer.
PCR for TWEAK and Fn14 was performed as follows:
94°C for 5 min (first denaturation/hot start) and then at 94°C for
1 (denaturation), 52°C for 1.5 min (annealing), and 72°C for 1 min
(extension) for 35 cycles with a 10 min final extension at 72°C.
PCR conditions for β-actin were identical except for a 1 min
annealing at 55°C, and a total of 27 cycles, with a 10 min final
extension.
PCR amplifications were performed in a PTC-200
Peltier Thermal Cycler (MJ Research Inc. Waltham, MA, USA). PCR
products were analyzed by agarose gel (1.5%) electrophoresis and
photographed under UV light. Nucleotide sequences of PCR primers
used were as follows: Fn14, 5′-GAC CTG GAC AAG TGC ATG GAC-3′
(sense) and 5′-AGC TGT TTT GTG TGA GCC AGC- 3′ (antisense); TWEAK
5′-ATC GCA GCC CAT TAT GAA GTT C-3′ (sense) and 5′-GAT GGA AAA CAC
GTG AAC AGG C-3′ (antisense); β-actin, 5′-TGA CGG GGT CAC CCA CAC
TGT GCC CAT CTA-3′ (sense) and 5′-ACT CGT CAT ACT CCT GCT TGC TGA
TCC A-3′ (antisense); PCR fragments of 500 (Fn14), 607 (TWEAK), and
625 bp (β-actin) were expected.
Microarray expression analysis
Raw data files from four European expression
microarray studies generated from two Affymetrix platforms (HU133A
and HU133plus2) were obtained from ArrayExpress (www.ebi.ac.uk/microarray-as/ae/) and the r2 data
base (http://hgserver1.amc.nl/cgi-bin/r2/main.cgi). The
three studies run on the HU133A platform (25–27),
were reanalyzed using gcRMA by Bioconducter for R 2.9.2 (library
BioC 2.4) in two separate groups: i) De Preter data set comprising
preamplified primary neuroblastoma samples (n=17, stages 1–4), ii)
McArdle (n=16) and Wilzén (n=8) data sets comprising
non-preamplified primary neuroblastoma samples. Also, expression
values (log2) from 76 MAS5.0 normalized neuroblastoma samples
(stages 1–4) run on the Affymetrix HU133plus2 platform were
obtained from the r2 database (28) and referred to as the Versteeg data
set.
Neuroblastoma samples from all three data sets
(25–27) were divided into two groups based on
their clinical stage (INSS stage) (29) and investigated for differential
expression of TNFSF12 (TWEAK) and TNFRSF12A (Fn14) between groups.
The significance was tested by Welch’s t-test (2-tailed, 2 sample
comparison, unequal variance).
Treatment of cells with TWEAK
To prevent influence of endogenously produced TWEAK,
neuroblastoma cells were serum starved in an RPMI-1640 medium
containing 0.1% FCS for 24 h prior to the incubation of recombinant
human TWEAK for the indicated concentrations and time-points.
Immunohistochemistry
Formalin-fixed and paraffin-embedded tissue sections
were deparaffinized in xylene and graded alcohols, hydrated and
washed in PBS. After antigen retrieval in sodium citrate buffer (pH
6.0) in a microwave oven, the endogenous peroxidase was blocked by
0.3% H2O2 for 15 min. Sections were incubated
overnight at 4°C with primary antibody (TWEAK; FL-249). As a
secondary antibody, the anti-rabbit horseradish peroxidase (HRP)
SuperPicTure Polymer detection kit was used (Zymed-Invitrogen, San
Francisco, CA, USA). A matched isotype control was used as a
control for non-specific background staining. Routine standard
staining showing a normal histology of neuroblastoma was performed
with hematoxylin and eosin.
For immunofluorescence studies, cells were grown on
fibronectin-coated chamber slides (Nunc, Roskilde, Denmark) for 24
h. Cultures were then washed and fixed with 2% paraformaldehyde for
15 min and 70% cold methanol for 5 min. After washing with PBS
buffer, goat-anti TWEAK (S-20) and rabbit-anti Fn14 antibodies were
incubated with cultures overnight at 4°C. After rinsing in PBS,
cultures were incubated with secondary antibodies conjugated with
Alexa 488 and Alexa 599, respectively. A matched isotype control
was used as a control for non-specific background staining. The
cells were examined in a Zeiss axiophot photomicroscope (Carl
Zeiss, Oberkochen, Germany). Nuclear translocation of NF-κB upon
stimulation with TWEAK was performed by immunofluorescence studies
using anti-NF-κB p65 antibody (sc-109) that recognizes both
non-phosphorylated and phosphorylated forms of NF-κB p65.
TWEAK ELISA
TWEAK ELISA was performed to measure the endogenous
production of TWEAK in neuroblastoma cells upon stimulation by
cytokines IL-1β and TNF-α. Neuroblastoma cells [SK-N-AS,
SK-N-BE(2) and SH-SY5Y] were
seeded in a regular growth medium in 96-well plates and allowed to
attach. Cells were then treated with two concentrations (10 and 50
ng/ml) of IL-1β and TNF-α, respectively, for 12 h. Supernatants
from treated cells were collected and 100 μl medium per well
were analyzed by using the Human TWEAK ELISA Development kit
(PeproTech, Rocky Hill, NJ, USA), following the manufacturer’s
instructions.
Protein isolation and immunoblotting
Proteins from TWEAK-treated cells and control cells
were extracted in RIPA lysis buffer (cat. no. 20–188, Upstate
Biotechnology, USA) containing complete, mini, EDTA-free protease
inhibitor (cat. no. 11 836 170 001, Roche). The protein content was
measured using Bradford reagents (Bio-Rad Laboratories, CA, USA).
Equal amounts of protein were separated by NuPAGE, Novex and
Tris-Acetat Mini Gels (Invitrogen) 4–12% in reduced conditions, and
proteins were transferred to a PVDF (Pierce, Rockford, IL, USA)
membrane and incubated with primary antibodies at 4°C overnight.
Alkaline-phosphatase conjugated antibodies were used as secondary
antibodies. Detection and visualization were performed using Pierce
Super Signaling solutions (Pierce), the Fujifilm Luminescent Image
Analyzer LAS-3000 and the Fujifilm MultiGauge (Ver. 3.0) analysis
software.
NF-κB luciferase reporter gene assay
The NF-κB-responsive reporter plasmid κBcon A-LUC
was provided by E. Sontag (30)
and also described in Johannesen et al(31). SK-N-AS and SK-N-BE cells were
seeded in 6-well plates in triplicates the day before transfection.
Cells were transfected using Lipofectamine 2000 (Invitrogen), 4
μg/μl DNA per well and calf thymus DNA (Amersham
Pharmacia, Sweden), and incubated for 6 h at 37°C in a humidified
5% CO2 atmosphere. Cells were then serum starved for 14
h and subsequently incubated for 6 h with TWEAK (100 ng/ml) or
TNF-α (30 ng/ml). Cells were washed in 1X PBS and lysed in TROPIX
lysis buffer containing 0.5 mM DTT. Luciferase activity was
determined using the Dual-Light Luciferase Gene Assay System
(Applied Biosystems Inc., Foster City, CA, USA) in a Luminoscan RT
(Labsystems, Helsinki, Finland). Luciferase measurements were
corrected to protein concentrations in each cell lysate, and
protein measurements were performed as described above. The
difference between the groups was analyzed on log-transformed
values after normalization to total protein content in the cell
lysates using a repeated measures one-way ANOVA test, followed by a
Bonferroni multiple comparison test.
Fn14 and TWEAK siRNA
SK-N-AS and SK-N-BE(2) cells were seeded in 6-well culture
plates in RPMI medium at a 30-50% confluence. Cells were
transfected with target-specific Fn14 (sc-43764), TWEAK (sc-37522),
control (Fluorescein conjugate-A) (sc-36869) or scrambled control
(sc-37007) siRNA (Santa Cruz Biotechnology), respectively, at a
concentration of 33 nM using Lipofectamine 2000 in OptiMEM. To
evaluate cell viability, western blot analysis of protein extracts
and trypan blue exclusion assay were performed 72 h after the
initial transfection. Transfection efficiency was assessed in
SK-N-AS cells transfected with control (Fluorescein conjugate-A)
using flow cytometry.
Detection of MMP-2 and MMP-9 in cell
conditioned media using gelatin zymography
The presence of MMP-2 and MMP-9 in serum-free media
from cells treated with or without recombinant TWEAK was determined
by SDS-gelatin zymography. Approximately 30,000 cells were seeded
in 96-well plates and left to attach in 10% RPMI-1640 overnight.
Then, SK-N-AS and SK-N-SH cells were starved in 0.1% RPMI-1640 for
24 h prior to incubation with TWEAK (0–1000 ng/ml) for 48 h.
SDS-substrate PAGE was done as previously described (32,33)
with gels containing 0.1% (w/v) gelatin. The gelatin zymograms were
calibrated with a mixture of conditioned serum-free medium from
THP-1 and humans skin fibroblast cells (32). Conditioned medium (10 μl)
was mixed with 2.5 μl of loading buffer (250 mM Tris-HCl, pH
6.8, 10% SDS, 0.03% bromophenol blue and 50% glycerol). Eight
μl of this non-heated mixture was applied to the gel, which
was run at 20 mA/gel at 4°C. Thereafter, the gel was washed twice
in 100 ml of washing buffer [2.5% (v/v) Triton X-100 in water], and
then incubated in 100 ml of assay buffer (50 mM Tris-HCl, pH 7.5, 5
mM CaCl2, 0.2 M NaCl and 0.02% Brij-35) for ∼20 h at
37°C. Gels were stained with 0.2% Coomassie brilliant blue R-250
(30% methanol) and destained in a solution containing 30% methanol
and 10% acetic acid. Gelatinase activity was evident as cleared
regions.
Statistical analyses
All statistical analyses were performed with
GraphPad Prism Software (GraphPad Software, San Diego, CA, USA).
The t-test was used to determine whether the mean of a single
sample differed significantly from control. To compare several
treatment groups, one-way ANOVA with Tukey multiple-comparisons
tests were used. P<0.05 was considered statistically
significant.
Results
TWEAK and Fn14 are expressed in
neuroblastoma primary tumors and cell lines
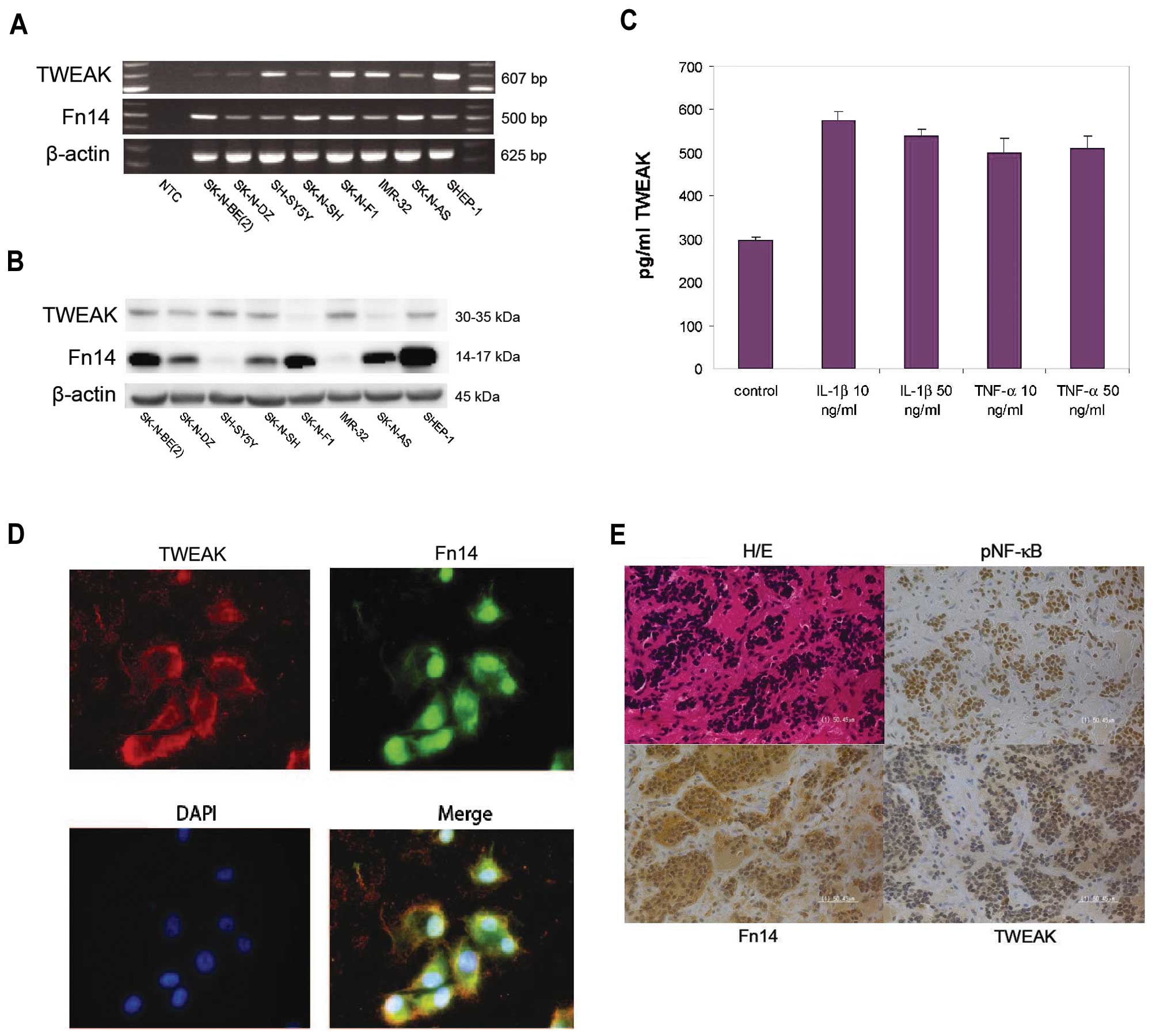
We investigated eight different neuroblastoma cell
lines for the expression of TWEAK and Fn14. All human neuroblastoma
cell lines investigated showed varying degrees of both TWEAK and
Fn14 mRNA and protein expression as detected by RT-PCR and western
blot analysis, respectively (Fig. 1A
and B). The 30–35-kDa band for TWEAK represents the
transmembrane form of TWEAK as the soluble form could not be
detected by western blot analysis. We selected 3 cell lines
SK-N-AS, SK-N-BE(2) and SH-SY5Y
for the in vitro experiments. The rationale behind choosing
these cell lines was based on the genetical and phenotypical
differences between the cell lines. SK-N-AS and SH-SY5Y are typical
non-MYCN-amplified cell lines, whereas the SK-N-BE(2) cell line is MYCN-amplified and
P53-mutated. Furthermore, while the SK-N-BE(2), SK-N-SH and SK-N-AS show multi-drug
resistant (MDR) phenotype, the SH-SY5Y cell line does not. In order
to investigate whether neuroblastoma cells were able to produce and
secrete TWEAK in vitro, we assessed the TWEAK production by
ELISA. The TWEAK levels in cell supernatants from SK-N-AS cells
showed an elevated secretion upon stimulation with pro-inflammatory
cytokines IL-1β and TNF-α (Fig.
1C). Similar results were obtained for SH-SY5Y and
SK-N-BE(2) cell lines (data not
shown).
Immunofluorescence staining of neuroblastoma SK-N-AS
cells stained with antibodies towards TWEAK (red) and Fn14 (green)
revealed the cellular distribution of the ligand and receptor,
demonstrating a distribution of TWEAK and Fn14 in the cytoplasm but
also to a certain degree to the nuclear compartment (Fig. 1D).
The staining of primary neuroblastoma tumor tissue
with antibodies against TWEAK and Fn14 revealed significant
cytoplasmic and nuclear expression of both TWEAK and Fn14 in all
primary tumors that were analyzed (Fig. 1E and Table I). No significant difference in
staining intensity between favorable and non-favorable tumors
(MYCN-amplified vs non-MYCN-amplified) could be
detected by immunohistochemistry and no staining was observed in
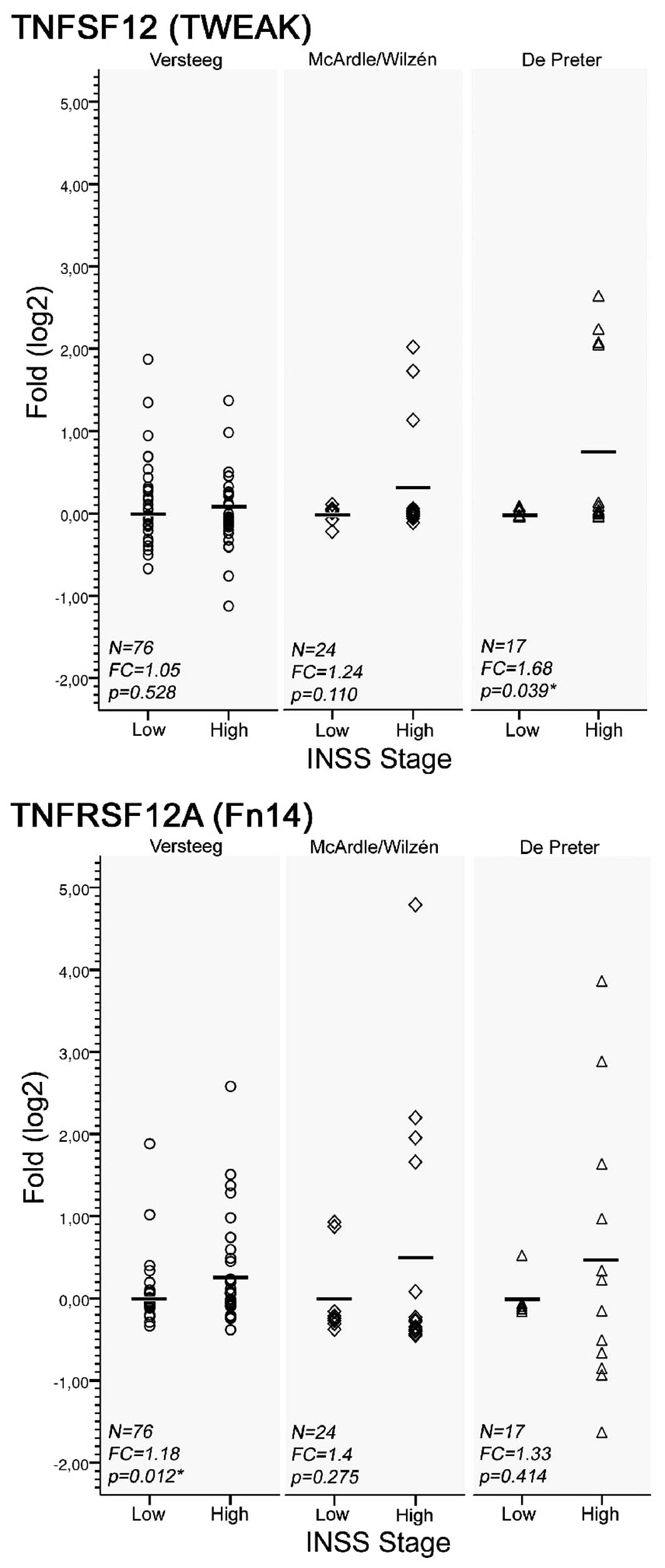
sections incubated with isotype control antibody. Moreover, mRNA
levels of both TWEAK and its receptor Fn14 were found to be
generally higher in primary high stage tumors (stage 3–4) compared
to low stage tumors (stage 1–2), when investigating three European
micro-array data sets (Fig. 2).
The up-regulation was significant for TWEAK and Fn14 in one out of
three data sets respectively, i.e. the De Preter and
Versteeg data sets (p<0.05, Welch’s t-test; Fig. 2). Also, the expression variance of
both TWEAK and Fn14 was considerably higher in the high-stage
group, and the up-regulation seemed to involve a sub-set of tumor
cases. Ten out of 117 cases from all three data sets showed
up-regulation of TWEAK (fold change >2), 13 showed up-regulation
of Fn14 (fold change >2), and 9 showed up-regulation of both
genes (fold change >2) compared to the mean expression levels of
the low-stage group (Fig. 2).
TWEAK promotes NF-κB activation and
nuclear trans localization in neuroblastoma cells
TWEAK treatment has previously been shown to
stimulate NF-κB activation in different cell types (15,34–36).
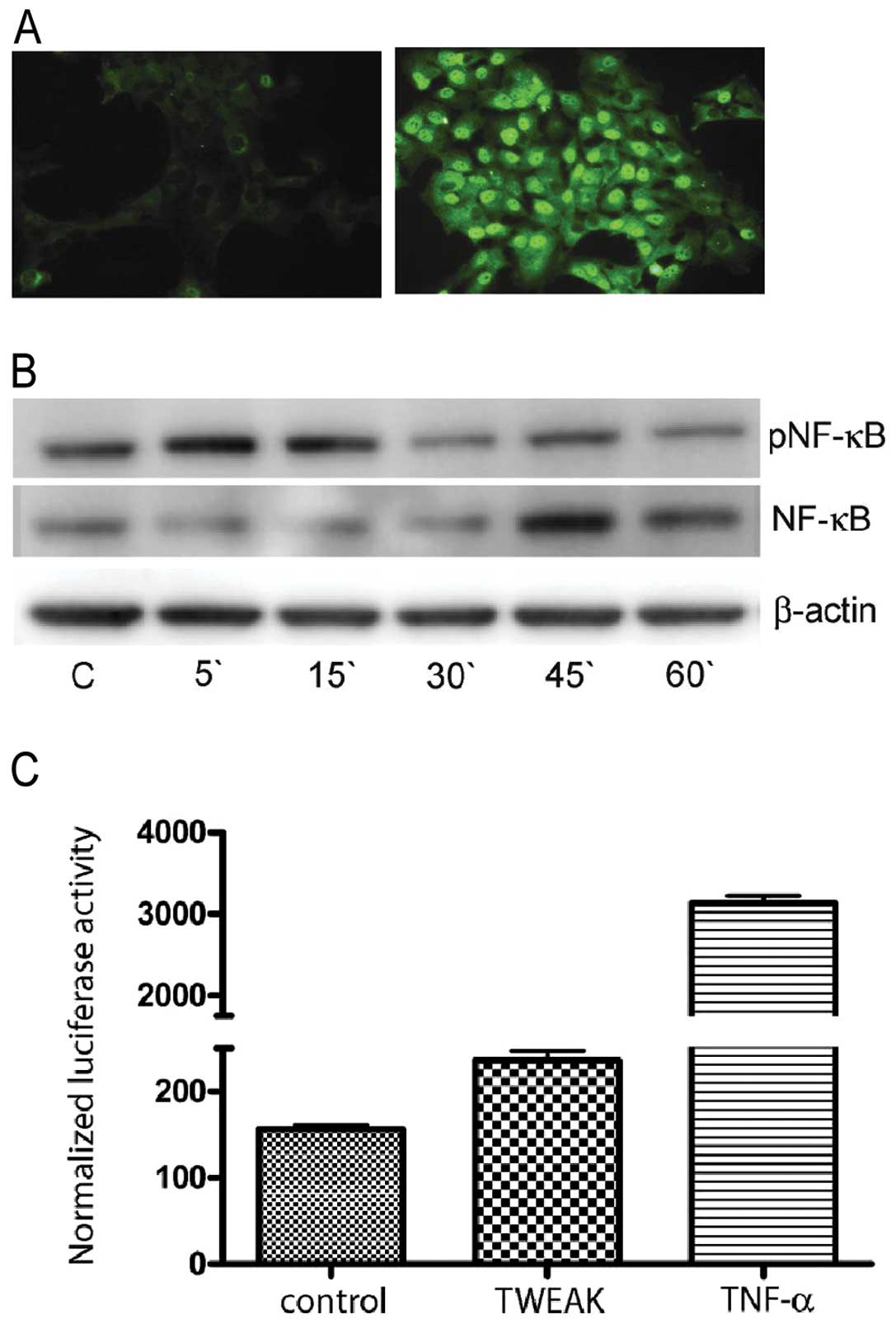
To determine if TWEAK could induce activation of NF-κB in
neuroblastoma cells, we examined the intracellular localization of
NF-κB in TWEAK stimulated SK-N-AS cells. No nuclear staining for
NF-κB was observed in untreated cells, but after 20 min of
stimulation with recombinant TWEAK, nuclear trans-location of NF-κB
was detected (Fig. 3A). To
validate the TWEAK induction of NF-κB in SK-N-AS cells grown in
0.1% FCS, we isolated total protein fractions of cells treated with
TWEAK (100 ng/ml) and immunoblotted for both the NF-κB and the
pNF-κB subunit. Blots revealed an increased level of pNF-κB (p65)
subunit shortly after stimulation by TWEAK at the same time that
NF-κB showed a decreased signal (Fig.
3B). This is consistent with the immunostaining results showing
activation of pNF-κB (Fig. 3A),
and the results of SK-N-AS cells upon stimulation with TWEAK
showing increase of NF-κB transcriptional activity upon transient
transfection using the NF-κB-responsive reporter plasmid κBcon
A-LUC (Fig. 3C).
Furthermore, immunohistochemical analysis of
neuroblastoma primary tumors using phospho-specific NF-κB antibody
(p65) revealed a significant nuclear staining (Fig. 1E).
Silencing of TWEAK and Fn14 reduces
neuroblastoma cell viability
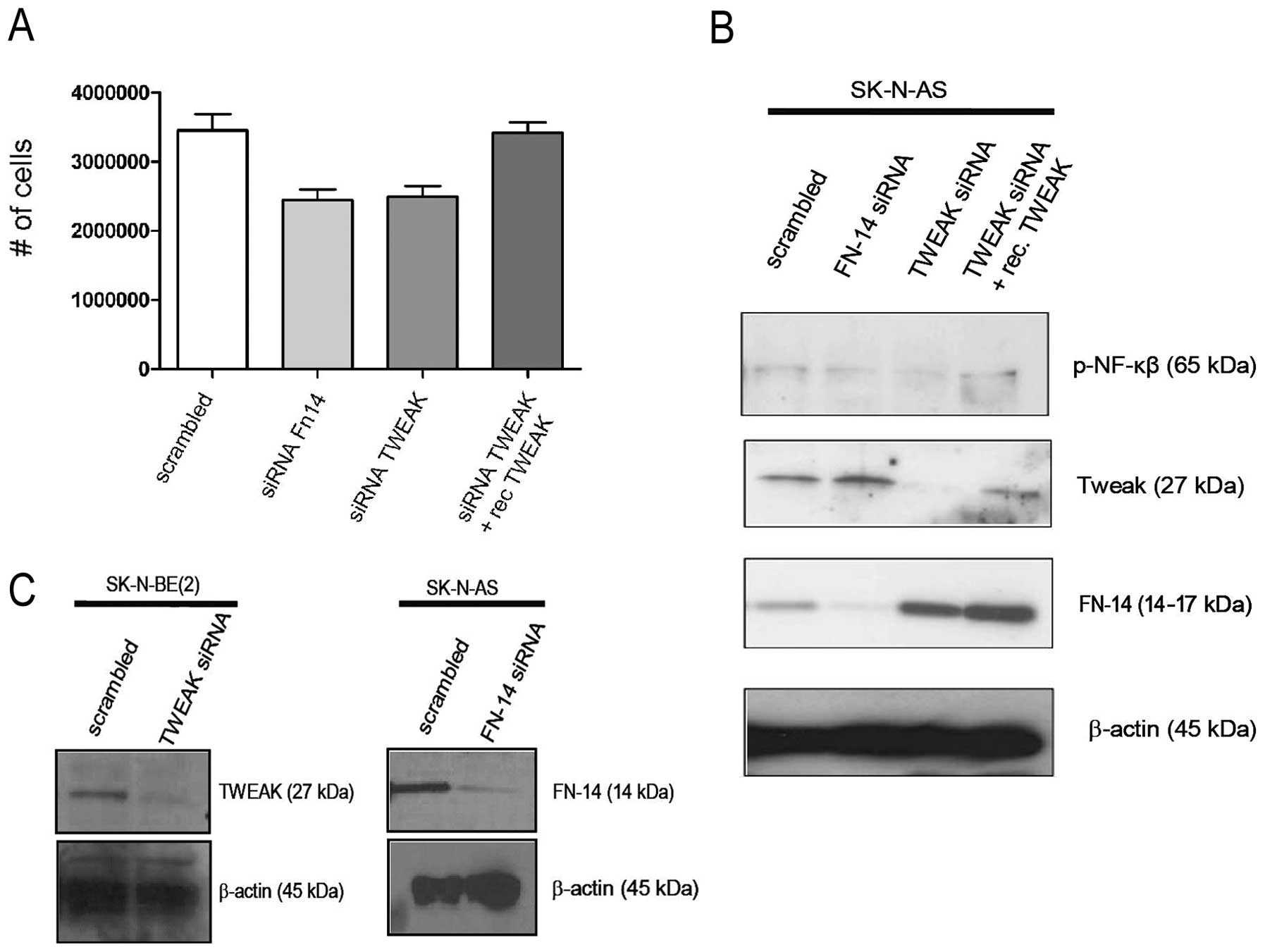
To investigate the influence of TWEAK on
neuroblastoma cell survival, SK-N-AS cells were transfected with
siRNA targeting TWEAK or Fn14. As shown in Fig. 4A, the silencing of both TWEAK and
Fn14 resulted in significant decrease in neuroblastoma cell
survival compared to cells transfected with a scrambled siRNA
construct (p<0.05). Addition of recombinant TWEAK partly
restored TWEAK expression and cell survival in TWEAK siRNA-treated
SK-N-AS cells, underscoring a role for TWEAK in cell survival
(Fig. 4A and B). Increased
expression of Fn14 in TWEAK siRNA-treated cells compared to
scramble- and non-treated cells (Figs.
1B and 4B) is possibly due to
the absence of the ligand resulting in lowered receptor
internalization and degradation of receptor-ligand complex.
Additional western blotting was performed to also confirm the
specific down-regulation of TWEAK or Fn14 expression in SK-N-AS
cells following siRNA transfection (Fig. 4C).
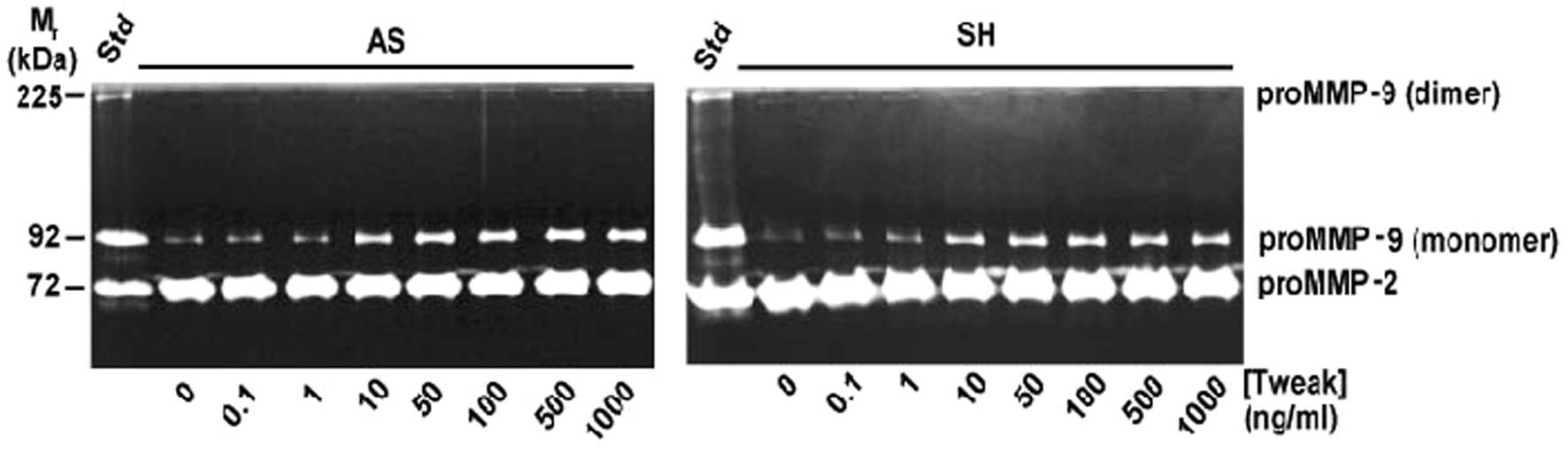
TWEAK induces MMP-9 expression in
neuroblastoma cells
Since MMP-9 is a NF-κB responsive gene and TWEAK has
been shown to induce MMP-9 protein level in other cell systems
(21,22), we investigated the effect of TWEAK
on the secretion of MMP-2 and MMP-9 in four different neuroblastoma
cell lines. Zymography performed on conditioned medium from
neuroblastoma cell lines revealed that MMP-2 was constitutively
expressed, whereas TWEAK induced the release of MMP-9 in a
dose-dependent manner in SK-N-AS and SK-N-SH cells (Fig. 5). Similar results were obtained for
SK-N-BE(2) cells whereas SK-N-SY
cells did not release MMP-9 upon stimulation with TWEAK (data not
shown), indicating a heterogeneity with respect to TWEAK induction
of MMP-9 in neuroblastoma cells.
Discussion
The TNF superfamily of proteins has been implicated
in the regulation of cell survival and proliferation. We have
previously studied the role of tumor necrosis factor-related
apoptosis inducing ligand (TRAIL) in neuroblastoma (37). In the present study, we describe
the expression of TWEAK in neuroblastoma. TWEAK is a member of the
TNF family of cytokines that acts on responsive cells via binding
to a cell surface receptor called Fn14. TWEAK is a multifunctional
cytokine expressed in a variety of normal tissue, but its
expression has also been implicated in certain types of cancer
[reviewed in refs. 16,18]. Equivalent to TWEAK, Fn14 is
expressed in most cells and tissues (16) although the expression level is
normally low. However, Fn14 expression can be induced by several
cytokines, hormones and inflammatory mediators (16). A high expression of Fn14 has been
detected in various adult cancers (7,15,38)
and Fn14 mRNA expression correlates with glioma grade and patient
outcome (15). In addition,
increased Fn14 mRNA and protein have been shown to be associated
with disease progression in esophageal adenocarcinoma (17,39)
and cancer of the mammary (21).
We analyzed neuroblastoma primary tumors from
different biological subsets and clinical stages, and detected
TWEAK and Fn14 in all samples investigated (Table I and Fig. 1E). TWEAK and Fn14 mRNA and protein
were all detected in various degrees in all neuroblastoma cell
lines investigated (Fig. 1A and
B), and soluble TWEAK was detected in supernatants from
neuroblastoma cells (Fig. 1C). The
discrepancy between mRNA and protein levels of TWEAK and Fn14 may
have root in several aspects such as mRNA stability and
post-translational modification. Furthermore, the mRNA analysis was
performed by reversal transcription PCR which is not a quantitative
method. Immunofluorescence images of SK-N-AS cells demonstrate that
the endo genously produced ligand and its receptor are co-localized
to the cytoplasm, and to a certain degree in the nucleus (Fig. 1D). Interestingly, a ‘short’ variant
of endogenously formed TWEAK possessing a nuclear localization
sequence has been shown to colocalize with GSK3β in the nucleus of
human neuroblastoma cells (40).
The significance of TWEAK and Fn14 expression in
neuronal cells is not well understood. TWEAK has been shown to be
expressed in primary murine neurons (41) and in the peripheral nervous system.
TWEAK and Fn14 have been reported to regulate neurite outgrowth and
regeneration (42), and a recent
study has also demonstrated the role of TWEAK/Fn14 in neurite
extension in neural progenitor cells (11).
We observed an up-regulation of both TWEAK and Fn14
mRNA in several primary tumors from three publicly available
European neuroblastoma data sets. It is presently unknown why the
Fn14 gene is elevated in certain solid tumors. Fn14 gene
amplification may possibly occur during the development and
progression of these tumors although this is not frequently
detected in neuroblastomas (43).
Alternatively, the expression of Fn14 might be driven by a range of
cytokines and growth factors produced within the tumor
microenvironment. Additionally, TWEAK produced by tumor cells or
other cells within the tumor microenvironment may activate Fn14 and
NF-κB in cancer cells by use of a positive feedback loop (15). In this study, both TWEAK and Fn14
mRNA were found at significantly higher levels in a sub-set of
neuroblastoma samples. Among 10 cases showing up-regulation of
TWEAK (fold change >2), 8 also show up-regulation of Fn14 (fold
change >2) supporting this hypothesis (Fig. 2).
To examine the effect of TWEAK on the proliferation
of neuroblastoma cells, we stimulated serum-starved cells with an
increasing concentration of TWEAK (1–1000 ng/ml) for 24 and 48 h.
In contrast to other reports using other types of tumor cells
(5,7,23),
we did not detect any significant increased proliferation of
neuroblastoma cells upon stimulation with TWEAK (data not shown).
However, this is in line with results from studies on murine
postnatal neural progenitor cells (11) and human embryonic kidney (HEK 293)
cells (44). TWEAK stimulation
through Fn14 activates the NF-κB signaling pathway in various cells
and induces the expression of pro-inflammatory molecules (16,45).
Inflammatory mediators are critical components of tumor growth and
the possibility that TWEAK produced by tumor cells may act on
stromal cells within the tumor is supported by studies showing that
TWEAK can induce secretion of cytokines from endothelial cells
(7), fibroblasts (46) and macrophages (22). In addition, the activation of
stromal cells may promote the infiltration of innate immune system
cells that in turn may be potential sources for TWEAK and other
pro-inflammatory cytokines. In our study, pro-inflammatory
cytokines were shown to enhance TWEAK secretion by neuroblastoma
cells (Fig. 1C). Therefore,
inflammatory mediators within the tumor microenvironment may
contribute to the further induction of TWEAK secretion by
neuroblastoma cells.
Numerous genes have been described that are
regulated by NF-κB and mediate the survival of cancer cells. These
include genes such as MMP-9(47), VEGF(48), and COX-2(49), which have been closely associated
with invasion and angiogenesis.
In the present study, we detected the nuclear
expression of phospho-NF-κB in all primary neuroblastoma tissue
samples by the use of immunohistochemistry (Fig. 1E). The phosphorylation of NF-κB
(p65) was induced upon TWEAK stimulation of neuroblastoma cells as
shown by western blot analysis (Fig.
3B). Furthermore, we demonstrate the trans-(Fig. 3B). Furthermore, we demonstrate the
trans-location of NF-κB into the nucleus of TWEAK-stimulated
neuroblastoma cells (Fig. 3A). A
recent study demonstrated the the Fn14 expression in gastric cancer
was inversely correlated with patient survival. Furthermore,
expression level of Fn14 was shown to affect cell growth which in
turn was mediated by NF-κB activity (50).
Among proteases involved in tumor invasion and
metastasis are the matrix metalloproteases (MMPs), a large family
of endopeptidases which together can process all extracellular
matrix proteins as well as other non-matrix proteins such as growth
factors, cytokines and receptors (51,52).
Neuroblastoma cells are able to produce extracellular matrix
degrading enzymes such as MMP-2 and MMP-9 (53–55).
TWEAK has been shown to upregulate MMP-9 expression in both normal
(22) and transformed cells
(21,56). Stimulating neuroblastoma cells with
TWEAK induced the expression of MMP-9 in a dose-dependent manner as
observed in gelatin zymograph. Neither of the cell lines responded
to TWEAK stimulation by a change in MMP-2 expression (Fig. 5). These results suggest that TWEAK
may have important functions during the metastasis conversion in
neuroblastoma.
In conclusion, our data demonstrate that TWEAK and
Fn14 are expressed in neuroblastoma, and may play an important role
in pro-survival features of the tumor. Given that targeted antibody
therapy for Fn14 in several human xenografts results in significant
antitumor effects (57,58) a further evaluation of the
mechanisms behind TWEAK and Fn14 expression may reveal new
therapeutic options for neuroblastoma.
Acknowledgements
We wish to thank Anja Inkeri Vepså,
Eli Berg and Kenneth Larsen for their technical assistance. This
study was supported by grants from the Norwegian Cancer Society
(DNK), The Ragnvarda F. Sörvik and Hakon Starheims Foundation,
Norway, The Familien Blix Fond, The Swedish Children Cancer
Foundation, The Swedish Cancer Society, Marta and Gunnar V
Philipson Foundation, The Mary Bevé Foundation, Dammen Foundation,
Karolinska Institutet and the Swedish Research Council.
References
|
1
|
Locksley RM, Killeen N and Lenardo MJ: The
TNF and TNF receptor superfamilies: integrating mammalian biology.
Cell. 104:487–501. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bodmer JL, Schneider P and Tschopp J: The
molecular architecture of the TNF superfamily. Trends Biochem Sci.
27:19–26. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chicheportiche Y, Bourdon PR, Xu H, et al:
TWEAK, a new secreted ligand in the tumor necrosis factor family
that weakly induces apoptosis. J Biol Chem. 272:32401–32410. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tran NL, McDonough WS, Savitch BA, Sawyer
TF, Winkles JA and Berens ME: The tumor necrosis factor-like weak
inducer of apoptosis (TWEAK)-fibroblast growth factor-inducible 14
(Fn14) signaling system regulates glioma cell survival via NFkappaB
pathway activation and BCL-XL/BCL-W expression. J Biol Chem.
280:3483–3492. 2005. View Article : Google Scholar
|
|
5
|
Lynch CN, Wang YC, Lund JK, Chen YW, Leal
JA and Wiley SR: TWEAK induces angiogenesis and proliferation of
endothelial cells. J Biol Chem. 274:8455–8459. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Harada N, Nakayama M, Nakano H, Fukuchi Y,
Yagita H and Okumura K: Pro-inflammatory effect of TWEAK/Fn14
interaction on human umbilical vein endothelial cells. Biochem
Biophys Res Commun. 299:488–493. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kawakita T, Shiraki K, Yamanaka Y, et al:
Functional expression of TWEAK in human hepatocellular carcinoma:
possible implication in cell proliferation and tumor angiogenesis.
Biochem Biophys Res Commun. 318:726–733. 2004. View Article : Google Scholar
|
|
8
|
Donohue PJ, Richards CM, Brown SA, et al:
TWEAK is an endothelial cell growth and chemotactic factor that
also potentiates FGF-2 and VEGF-A mitogenic activity. Arterioscler
Thromb Vasc Biol. 23:594–600. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tran NL, McDonough WS, Donohue PJ, et al:
The human Fn14 receptor gene is up-regulated in migrating glioma
cells in vitro and overexpressed in advanced glial tumors. Am J
Pathol. 162:1313–1321. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Perper SJ, Browning B, Burkly LC, et al:
TWEAK is a novel arthritogenic mediator. J Immunol. 177:2610–2620.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hamill CA, Michaelson JS, Hahm K, Burkly
LC and Kessler A: Age-dependent effects of TWEAK/Fn14 receptor
activation on neural progenitor cells. J Neurosci Res.
85:3535–3544. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Marsters SA, Sheridan JP, Pitti RM, Brush
J, Goddard A and Ashkenazi A: Identification of a ligand for the
death-domain-containing receptor Apo3. Curr Biol. 8:525–528. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nakayama M, Kayagaki N, Yamaguchi N,
Okumura K and Yagita H: Involvement of TWEAK in interferon
gamma-stimulated monocyte cytotoxicity. J Exp Med. 192:1373–1380.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Meighan-Mantha RL, Hsu DK, Guo Y, et al:
The mitogeninducible Fn14 gene encodes a type I transmembrane
protein that modulates fibroblast adhesion and migration. J Biol
Chem. 274:33166–33176. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tran NL, McDonough WS, Savitch BA, et al:
Increased fibroblast growth factor-inducible 14 expression levels
promote glioma cell invasion via Rac1 and nuclear factor-kappaB and
correlate with poor patient outcome. Cancer Res. 66:9535–9542.
2006. View Article : Google Scholar
|
|
16
|
Winkles JA: The TWEAK-Fn14
cytokine-receptor axis: discovery, biology and therapeutic
targeting. Nat Rev Drug Discov. 7:411–425. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Watts GS, Tran NL, Berens ME, et al:
Identification of Fn14/TWEAK receptor as a potential therapeutic
target in esophageal adenocarcinoma. Int J Cancer. 121:2132–2139.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Winkles JA, Tran NL, Brown SA, Stains N,
Cunliffe HE and Berens ME: Role of TWEAK and Fn14 in tumor biology.
Front Biosci. 12:2761–2771. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Winkles JA, Tran NL and Berens ME: TWEAK
and Fn14: new molecular targets for cancer therapy? Cancer Lett.
235:11–17. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li H, Mittal A, Paul PK, et al: Tumor
necrosis factor-related weak inducer of apoptosis augments matrix
metalloproteinase 9 (MMP-9) production in skeletal muscle through
the activation of nuclear factor-kappaB-inducing kinase and p38
mitogen-activated protein kinase: a potential role of MMP-9 in
myopathy. J Biol Chem. 284:4439–4450. 2009.
|
|
21
|
Michaelson JS, Cho S, Browning B, et al:
Tweak induces mammary epithelial branching morphogenesis. Oncogene.
24:2613–2624. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim SH, Kang YJ, Kim WJ, et al: TWEAK can
induce pro-inflammatory cytokines and matrix metalloproteinase-9 in
macrophages. Circ J. 68:396–399. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jakubowski A, Ambrose C, Parr M, et al:
TWEAK induces liver progenitor cell proliferation. J Clin Invest.
115:2330–2340. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maris JM, Hogarty MD, Bagatell R and Cohn
SL: Neuroblastoma. Lancet. 369:2106–2120. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
De Preter K, Vandesompele J, Heimann P, et
al: Human fetal neuroblast and neuroblastoma transcriptome analysis
confirms neuroblast origin and highlights neuroblastoma candidate
genes. Genome Biol. 7:R842006.
|
|
26
|
McArdle L, McDermott M, Purcell R, et al:
Oligonucleotide micro-array analysis of gene expression in
neuroblastoma displaying loss of chromosome 11q. Carcinogenesis.
25:1599–1609. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wilzen A, Nilsson S, Sjoberg RM, Kogner P,
Martinsson T and Abel F: The Phox2 pathway is differentially
expressed in neuroblastoma tumors, but no mutations were found in
the candidate tumor suppressor gene PHOX2A. Int J Oncol.
34:697–705. 2009.
|
|
28
|
Molenaar JJ, Ebus ME, Koster J, et al:
Cyclin D1 and CDK4 activity contribute to the undifferentiated
phenotype in neuroblastoma. Cancer Res. 68:2599–2609. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Brodeur GM, Pritchard J, Berthold F, et
al: Revisions of the international criteria for neuroblastoma
diagnosis, staging, and response to treatment. J Clin Oncol.
11:1466–1477. 1993.PubMed/NCBI
|
|
30
|
Sontag E, Sontag JM and Garcia A: Protein
phosphatase 2A is a critical regulator of protein kinase C zeta
signaling targeted by SV40 small t to promote cell growth and
NF-kappaB activation. EMBO J. 16:5662–5671. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Johannessen M, Olsen PA, Sorensen R,
Johansen B, Seternes OM and Moens U: A role of the TATA box and the
general co-activator hTAF(II)130/135 in promoter-specific
trans-activation by simian virus 40 small t antigen. J Gen Virol.
84:1887–1897. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Malla N, Berg E, Uhlin-Hansen L and
Winberg JO: Interaction of pro-matrix
metalloproteinase-9/proteoglycan heteromer with gelatin and
collagen. J Biol Chem. 283:13652–13665. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Winberg JO, Kolset SO, Berg E and
Uhlin-Hansen L: Macrophages secrete matrix metalloproteinase 9
covalently linked to the core protein of chondroitin sulphate
proteoglycans. J Mol Biol. 304:669–680. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dogra C, Changotra H, Mohan S and Kumar A:
Tumor necrosis factor-like weak inducer of apoptosis inhibits
skeletal myogenesis through sustained activation of nuclear
factor-kappaB and degradation of MyoD protein. J Biol Chem.
281:10327–10336. 2006. View Article : Google Scholar
|
|
35
|
Han S, Yoon K, Lee K, et al: TNF-related
weak inducer of apoptosis receptor, a TNF receptor superfamily
member, activates NF-kappa B through TNF receptor-associated
factors. Biochem Biophys Res Commun. 305:789–796. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Saitoh T, Nakayama M, Nakano H, Yagita H,
Yamamoto N and Yamaoka S: TWEAK induces NF-kappaB2 p100 processing
and long lasting NF-kappaB activation. J Biol Chem.
278:36005–36012. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Johnsen JI, Pettersen I, Ponthan F,
Sveinbjornsson B, Flaegstad T and Kogner P: Synergistic induction
of apoptosis in neuroblastoma cells using a combination of
cytostatic drugs with interferon-gamma and TRAIL. Int J Oncol.
25:1849–1857. 2004.PubMed/NCBI
|
|
38
|
Feng SL, Guo Y, Factor VM, et al: The Fn14
immediate-early response gene is induced during liver regeneration
and highly expressed in both human and murine hepatocellular
carcinomas. Am J Pathol. 156:1253–1261. 2000. View Article : Google Scholar
|
|
39
|
Wang S, Zhan M, Yin J, et al:
Transcriptional profiling suggests that Barrett’s metaplasia is an
early intermediate stage in esophageal adenocarcinogenesis.
Oncogene. 25:3346–3356. 2006.
|
|
40
|
De Ketelaere A, Vermeulen L, Vialard J, et
al: Involvement of GSK-3beta in TWEAK-mediated NF-kappaB
activation. FEBS Lett. 566:60–64. 2004.PubMed/NCBI
|
|
41
|
Yepes M, Brown SA, Moore EG, Smith EP,
Lawrence DA and Winkles JA: A soluble Fn14-Fc decoy receptor
reduces infarct volume in a murine model of cerebral ischemia. Am J
Pathol. 166:511–520. 2005.PubMed/NCBI
|
|
42
|
Tanabe K, Bonilla I, Winkles JA and
Strittmatter SM: Fibroblast growth factor-inducible-14 is induced
in axotomized neurons and promotes neurite outgrowth. J Neurosci.
23:9675–9686. 2003.PubMed/NCBI
|
|
43
|
Caren H, Kryh H, Nethander M, et al:
High-risk neuroblastoma tumors with 11q-deletion display a poor
prognostic, chromosome instability phenotype with later onset. Proc
Natl Acad Sci USA. 107:4323–4328. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ho DH, Vu H, Brown SA, Donohue PJ, Hanscom
HN and Winkles JA: Soluble tumor necrosis factor-like weak inducer
of apoptosis overexpression in HEK293 cells promotes tumor growth
and angiogenesis in athymic nude mice. Cancer Res. 64:8968–8972.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gao HX, Campbell SR, Burkly LC, et al:
TNF-like weak inducer of apoptosis (TWEAK) induces inflammatory and
proliferative effects in human kidney cells. Cytokine. 46:24–35.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chicheportiche Y, Chicheportiche R, Sizing
I, et al: Proinflammatory activity of TWEAK on human dermal
fibroblasts and synoviocytes: blocking and enhancing effects of
anti-TWEAK monoclonal antibodies. Arthritis Res. 4:126–133. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yokoo T and Kitamura M: Antioxidant PDTC
induces stromelysin expression in mesangial cells via a tyrosine
kinase-AP-1 pathway. Am J Physiol. 270:F806–F811. 1996.PubMed/NCBI
|
|
48
|
Huang S, Robinson JB, Deguzman A, Bucana
CD and Fidler IJ: Blockade of nuclear factor-kappaB signaling
inhibits angiogenesis and tumorigenicity of human ovarian cancer
cells by suppressing expression of vascular endothelial growth
factor and interleukin 8. Cancer Res. 60:5334–5339. 2000.
|
|
49
|
Schmedtje JF Jr, Ji YS, Liu WL, DuBois RN
and Runge MS: Hypoxia induces cyclooxygenase-2 via the NF-kappaB
p65 transcription factor in human vascular endothelial cells. J
Biol Chem. 272:601–608. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kwon OH, Park SJ, Kang TW, et al: Elevated
fibroblast growth factor-inducible 14 expression promotes gastric
cancer growth via nuclear factor-kappaB and is associated with poor
patient outcome. Cancer Lett. 314:73–81. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tonti GA, Mannello F, Cacci E and Biagioni
S: Neural stem cells at the crossroads: MMPs may tell the way. Int
J Dev Biol. 53:1–17. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hadler-Olsen E, Fadnes B, Sylte I,
Uhlin-Hansen L and Winberg JO: Regulation of matrix
metalloproteinase activity in health and disease. FEBS J.
278:28–45. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Farina AR, Tiberio A, Tacconelli A,
Cappabianca L, Gulino A and Mackay AR: Identification of
plasminogen in Matrigel and its activation by reconstitution of
this basement membrane extract. Biotechniques. 21:904–909.
1996.PubMed/NCBI
|
|
54
|
Ara T, Fukuzawa M, Kusafuka T, et al:
Immunohistochemical expression of MMP-2, MMP-9, and TIMP-2 in
neuroblastoma: association with tumor progression and clinical
outcome. J Pediatr Surg. 33:1272–1278. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ribatti D, Surico G, Vacca A, et al:
Angiogenesis extent and expression of matrix metalloproteinase-2
and -9 correlate with progression in human neuroblastoma. Life Sci.
68:1161–1168. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Huang M, Narita S, Tsuchiya N, et al:
Overexpression of Fn14 promotes androgen-independent prostate
cancer progression through MMP-9 and correlates with poor treatment
outcome. Carcinogenesis. 32:1589–1596. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Culp PA, Choi D, Zhang Y, et al:
Antibodies to TWEAK receptor inhibit human tumor growth through
dual mechanisms. Clin Cancer Res. 16:497–508. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Michaelson JS, Kelly R, Yang L, Zhang X,
Wortham K and JosepH IB: The anti-Fn14 antibody BIIB036 inhibits
tumor growth in xenografts and patient derived primary tumor models
and enhances efficacy of chemotherapeutic agents in multiple
xenograft models. Cancer Biol Ther. 13:812–821. 2012. View Article : Google Scholar : PubMed/NCBI
|