Introduction
The signal transducer and activator of transcription
(STAT) 3 is a point of convergence for various oncogenic signaling
pathways through protein tyrosine kinases and is constitutively
activated both in tumor cells and immune cells (1,2).
Furthermore, STAT3 activation promotes the production of
immunosuppressive factors mainly by the tumor itself that induce
immunoregulatory immune cells and a marked immunosuppressive
environment around the tumor tissue (3,4).
Glioblastoma multiforme (GBM) is one of the most
malignant and aggressive tumors and has a very poor prognosis, with
a mean survival time of less than 2 years even with the recent
development of temozolomide (TMZ)-based intensive treatment
(5,6). Once recurrence develops, there are
few therapeutic approaches to control the growth of glioblastoma.
Therefore, in particular, TMZ-resistant GBM is very difficult to
treat, and a novel approach to overcome resistance is eagerly
awaited.
The activation of several signaling pathways,
including receptor tyrosine kinase (7), Akt (8), MAPK (9), Wnt (10) and Notch and Hedgehog (11) pathways, is involved in the
progression of GBM. Importantly, constitutive activation of the
Janus kinase (JAK)/signal transducer and activator of transcription
(STAT) pathway contributes to tumor progression by promoting cell
proliferation and inhibiting apoptosis.
O6-methylguanine-DNA-methyltransferase
(MGMT) is well known to remove methylation from the O6
position of guanine and contribute to TMZ resistance induction
(12). It is generally accepted
that high MGMT expression through the methylation of MGMT promoter
is one of the mechanisms responsible for TMZ resistance.
On the other hand, it is increasingly suggested that
a mechanism other than MGMT can trigger TMZ resistance based on
multiomics analysis. Several novel biomarkers linked to MGMT
expression and the methylation status such as the HOX signature and
EGFR expression (13), somatic
mutation of mismatch repair gene MSH6 (14), prolyl 4-hydroxylase, β-polypeptide
(P4HB), EGFR mutation (EGFRvIII) (15) and CD74, have been reported.
Considering that STAT3 might be another target for
chemo-resistance treatment, several therapeutic agents, including
small molecules acting as STAT3 activity inhibitors, have been
reported to show antitumor effects through the control of
chemo-resistance. Previously, we identified a novel inhibitor of
STAT3 dimerization, STX-0119, which exhibited a potent antitumor
effect on a human lymphoma cell line with highly activated STAT3
and GB-stem cell lines (16). In
the present study, we investigated the effect of STX-0119 on a
TMZ-resistant glioblastoma cell line in vitro and in
vivo, and demonstrated that STX-0119 molecule significantly
inhibited the proliferation of even a TMZ-resistant cell line and
prolonged the survival of tumor-transplanted mice.
Materials and methods
Establishment of TMZ-resistant U87 cell
line
U87 cells were purchased from the American Type
Culture Collection (ATCC, Manassas, VA) and cultured in Dulbecco’s
modified Eagle’s medium (DMEM) (Sigma-Aldrich, St. Louis, MO)
supplemented with 10% fetal bovine serum (FBS; Life Technology,
Carlsbad, CA), penicillin and streptomycin. The U87 parental cell
line, which is sensitive to TMZ, was first maintained at a low dose
of TMZ (5 μM) and then successively exposed to incremental
doses of TMZ (up to 150 μM). After killing the majority of
cells, surviving cells were maintained until a normal rate of
growth was obtained. The IC50 of TMZ was evaluated by
the WST-1 assay. TMZ-resistant (R) U87 cells with more than 20
passages were maintained at a dose of 100 μM TMZ and used
for in vitro and in vivo experiments.
Antibodies and reagents
Antibodies against STAT3, phosphospecific STAT3
(Tyr705), cleaved caspase-3, MGMT and β-actin were purchased from
Cell Signaling Technology, Inc. (Danvers, MA) and Becton-Dickinson
(BD) Biosciences (Franklin Lakes, NJ) for western blot analysis
(WB). Mouse anti-human YKL-40 antibody was purchased from Abcam
(Cambridge, MA, USA).
Chemicals
STX-0119 was supplied by the Center for Drug
Discovery, University of Shizuoka (Shizuoka, Japan). These
compounds were suspended and diluted in a sterile 0.5 w/v% methyl
cellulose 400 cp solution (Wako, Tokyo, Japan) for use in animal
experiments. TMZ was purchased from Sigma-Aldrich.
Cell proliferation assay
Cell proliferation was examined using the WST-1
assay (Dojin Kagaku Corp., Kumamoto, Japan) described previously
(16). Briefly, 1×104
parental U87 or TMZ-R U87 cells were seeded into each well of a
96-well micro-culture plate (Corning Inc., Corning, NY) and
compounds ranging from 0.25 to 500 μM were added. After 4
days, the WST-1 substrate was added to the culture and optical
density (OD) was measured at 450 and 620 nm using an immunoreader
(Immuno Mini NJ-2300; Nalge Nunc International, Roskilde, Denmark).
The IC50 value was defined as the dose needed for a 50%
reduction in OD calculated from the survival curve. Percent
survival was calculated as follows: (mean OD of test wells - mean
OD of background wells)/(mean OD of control wells - mean OD of
background wells) ×100.
Cell invasion assay
The invasion assay using TMZ-R U87 cells was
performed on Matrigel-coated (0.33 mg/ml) Transwell inserts with 8
μm pore size (BD Biosciences). A total of 500 μl
cells at 2×104/ml were added to Transwells in
triplicate, and 750 μl DMEM containing 10% FBS was added to
the lower wells. After 12-18 h incubation, cells that passed
through the membrane were fixed and stained with Diff-Quik II
solution (Siemens AG, Erlangen, Germany). Migrating cells were
counted under a microscope. To analyze the effect of STX-0119 on
invasion activity, TMZ-R U87 cells pre-treated with various doses
of STX-0119 (0, 25, 50 and 100 μM) for 48 h were utilized in
the assay.
Quantitative polymerase chain reaction
(qPCR) analysis
Real-time PCR analysis of stem cell and neuronal
markers, and STAT3 target genes using the 7500 Real-Time PCR System
(Applied Biosystems, Foster, CA) was performed as described
previously. Briefly, all PCR primers (CD24, YKL-40, GDF15,
HLA-DQA1, MAGEC1, MGMT, MMP1 as TMZ-R U87 cell-specific genes;
ALDH1A1, EGFR, GFAP, NANOG, NES, Oct3/4, SOX2 as GB stem cell
markers; FN1, FOXC2, MMP2, SNAIL1, SNAIL2, TCF4, TWIST1, SMAD2 as
EMT-associated genes; STAT3, FOSL2, C/EBP as GB mesenchymal marker
gene; BCL2, Survivin, c-Myc, CXCL10, TGFB1, P53, HIF-1α as STAT3
target genes) and TaqMan probes were purchased from Applied
Biosystems. Parental U87 or TMZ-R U87 cells were treated with
STX-0119 or DMSO for 24 h, and total RNA was extracted.
Complementary DNA was synthesized from 100 ng total RNA and
quantitative PCR was carried using a TaqMan RNA-to-Ct 1-Step kit
(Applied Biosystems).
ELISA for human YKL-40
YKL-40 levels in the supernatant of parental U87 or
TMZ-R U87 cells treated with STX-0119 were measured using human
YKL-40-specific ELISA. Cells were plated in 96-well microplates
(Corning) at 4×104 cells (200 μl cells at
2×105/ml) per well. After cells were treated with
STX-0119, WP1066 or DMSO for 24 h, supernatants were collected and
YKL-40 levels were measured.
Western blot analysis (WB)
TMZ-R U87 cells were treated with STX-0119 or DMSO
at various doses for 24 h. Cells were lysed using RIPA buffer
(Thermo Fisher Scientific Inc., Rockford, IL) containing protease
inhibitors and phosphatase inhibitors and used for western blot
analysis as described previously. Briefly, cell lysate was
subjected to SDS-PAGE with a 7.5% polyacrylamide separating gel,
and then transferred to PVDF membranes. After blocking, the
membranes were incubated at 4°C overnight with the primary antibody
against STAT3, phosphospecific STAT3, YKL-40 and β-actin
(1:200–1:2,000) in blocking solution. After washing, the membranes
were incubated for 1 h with horseradish peroxidase (HRP)-conjugated
anti-mouse IgG (1:5,000). Membranes were treated with ECL plus
reagent (GE Healthcare, Piscataway, NJ) and analyzed on a
chemiluminescence scanner (LAS-3000; Fujifilm, Tokyo, Japan).
Apoptosis induction in TMZ-R U87 cells treated with STX-0119 for 24
h was investigated using a Caspase-3 Western detection kit
including the primary antibody against cleaved caspase-3 (Cell
Signaling).
Animal experiments
Male nude mice (BALB/cA-nu/nu, 5–6 weeks old)
were obtained from Nippon Clea (Tokyo, Japan). All animals were
cared for and used humanely according to the guidelines for the
welfare and use of animals in cancer research, and the procedures
were approved by the Animal Care and Use Committee of Shizuoka
Cancer Center Research Institute.
Parental U87 cells (1×106) and TMZ-R U87
cells (1×106) were inoculated into the flank of
BALB/cA-nu/nu mice. To evaluate the antitumor activity
against subcutaneous (s.c.) inoculated tumors, tumor volume was
calculated based on the National Cancer Institute formula as
follows: tumor volume (mm3) = length (mm) × [width
(mm)]2 × 1/2.
STX-0119 at doses of 40 and 80 mg/kg was
administered orally daily from day 0 to day 4 followed by 2 days of
rest, which was repeated three times over 21 days. TMZ at a dose of
5 mg/kg was administered orally daily from day 0 to day 4. The
efficacy of compounds against human tumor cells inoculated into
nude mice was expressed as the mean V/V0 value
and evaluated as reported previously (16).
Statistical analysis
Statistical difference was analyzed using Student’s
t-test. Values of P<0.05 were considered statistically
significant. For the in vivo experiment, statistical
analysis was performed with corrected P-values to compare with the
untreated control using Mann-Whitney rank-sum test, and to
determine the effect of compounds on survival of tumor-bearing mice
the log-rank test was used for Kaplan-Meier survival curves.
Results
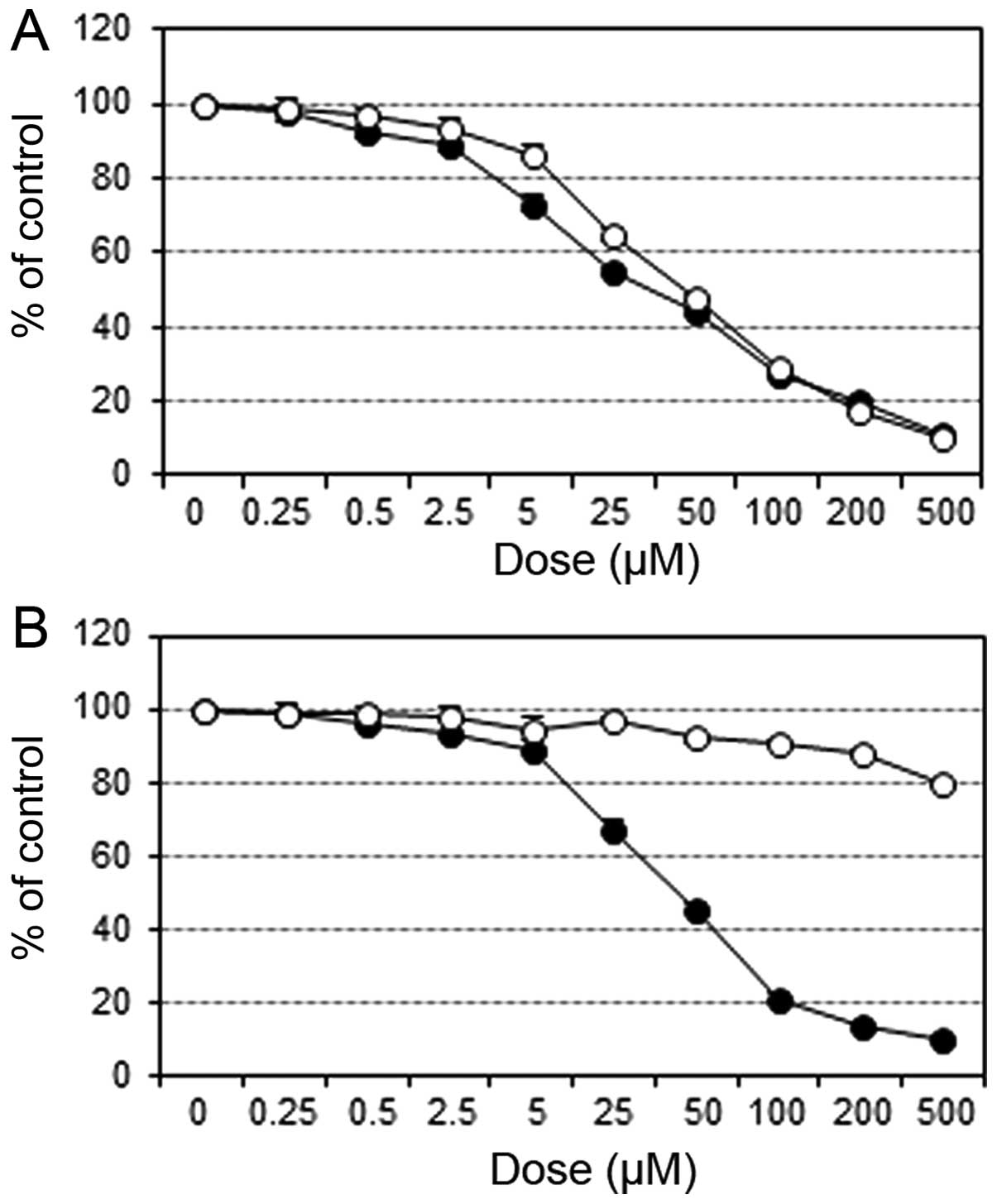
Cell proliferation assay
The growth inhibitory effects of STX-0119 and TMZ on
parental U87 cells were similar (STX-0119 IC50, 34
μM; TMZ IC50, 45 μM for U87 cells).
Whereas, the effect of STX-0119 on TNZ-R U87 was not so different
from U87 cells; however, TMZ had no inhibitory effect on TMZ-R U87
cells (STX-0119 IC50, 43 μM; TMZ IC50,
>500 μM for TMZ-R U87 cells) (Fig. 1).
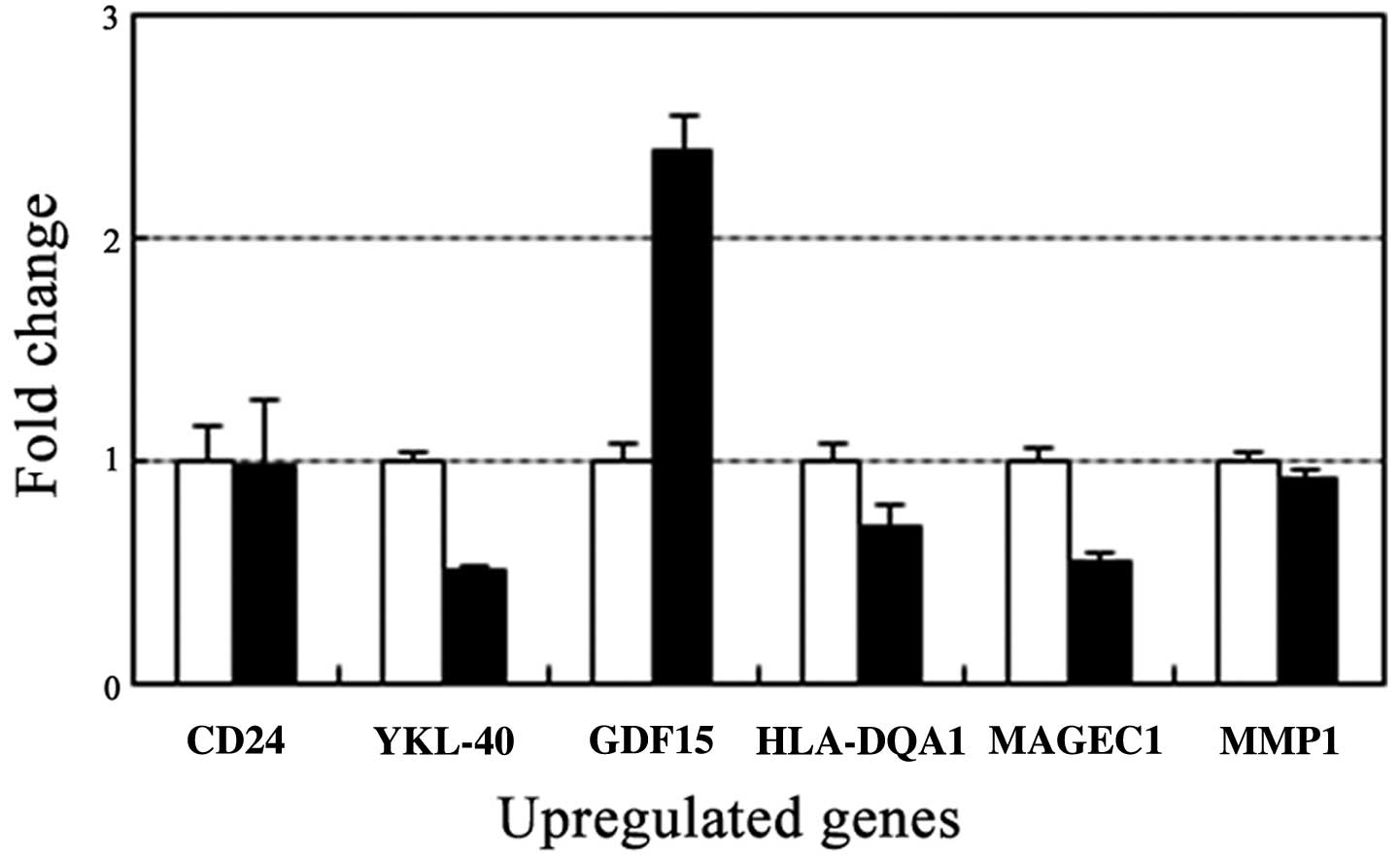
Effect of STX-0119 on upregulated genes
specific for TMZ-R U87 cell line
Based on differential gene expression profiling
(unpublished data) between parental U87 and TMZ-R U87 cells, the
upregulated genes specific for TMZ-R U87 cells were CD24, YKL-40,
GDF15, HLA-DQA1, MAGEC1, MMP1 and MGMT were upregulated more than
10-fold and 5-fold, respectively, in TMZ-R U87 cells compared to
parental U87 cells using real-time PCR. YKL-40 and MAGEC1 gene
expressions were significantly decreased after treatment with
STX-0119 at 100 μM (Fig.
2).
Effect of STX-0119 on STAT3 and YKL-40
protein expressions in TMZ-R U87 cell line
The activation (phosphorylation) of STAT3 and
upregulation of YKL-40 were identified in the TMZ-R U87 cell line
compared to the U87 cell line (data not shown). YKL-40 mRNA was
decreased by STX-0119 in a dose-dependent manner. YKL-40 protein
was markedly downregulated in 100 μM STX-0119-treated cells,
and even in the supernatant. However, the impact of STX-0119 on
STAT3 and phosphorylated STAT3 expression was marginal after
treatment.
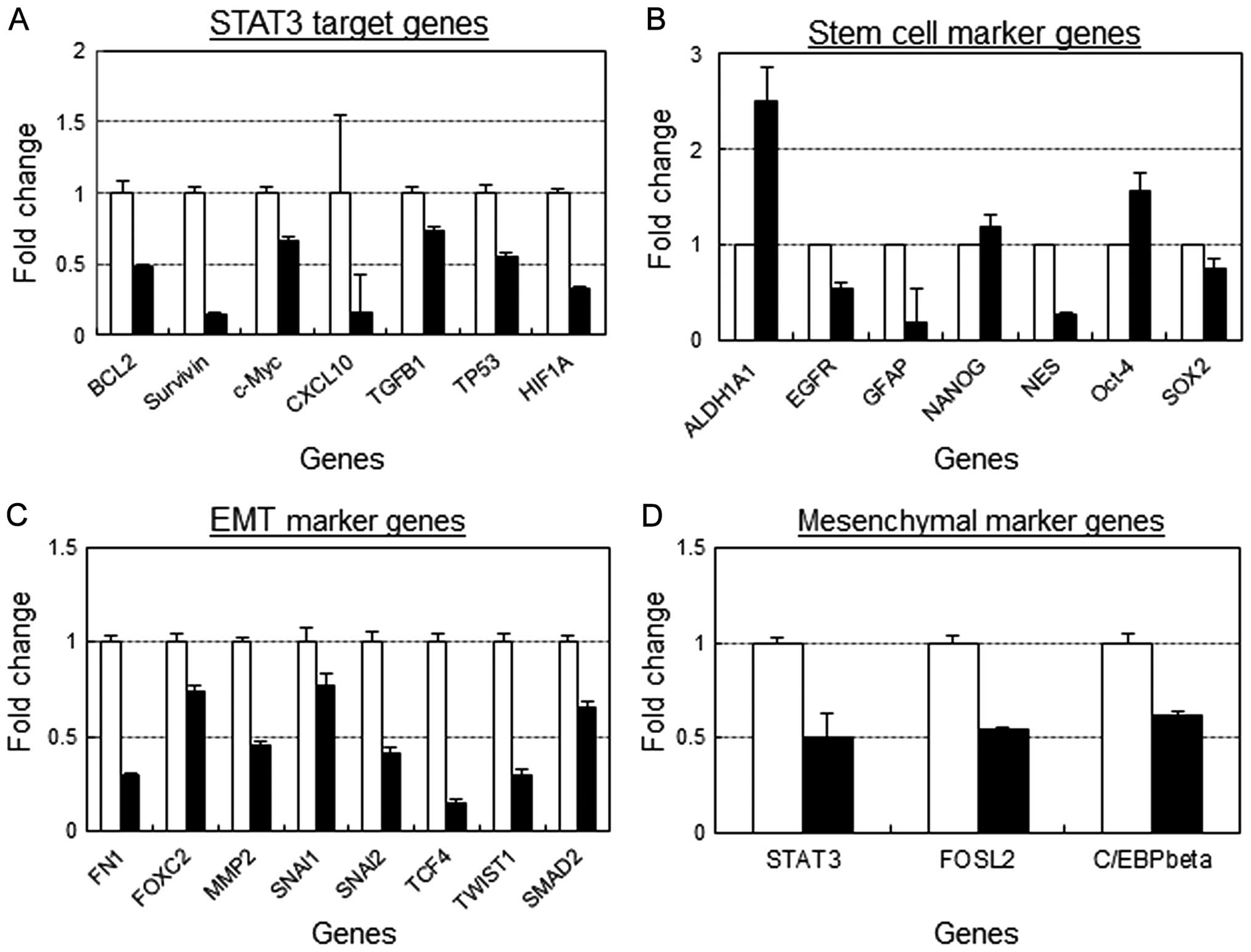
Effect of STX-0119 on upregulated genes
from various marker groups in TMZ-R U87 cell line
Real-time PCR analysis demonstrated that many marker
gene expressions were upregulated more than 2-fold in TMZ-R U87
cells compared to parental U87 cells as follows: BCL2, Survivin,
c-Myc, CXCL10, TGFB1, P53, HIF-1α as STAT3 target genes; ALDH1A1,
EGFR, GFAP, NANOG, NES, Oct3/4, SOX2 as GB stem cell markers; FN1,
FOXC2, MMP2, SNAIL1, SNAIL2, TCF4, TWIST1, SMAD2 as EMT-associated
genes; STAT3, FOSL2, C/EBP as GB mesenchymal marker gene (data not
shown). Several genes, including BCL2, Survivin, CXCL10, HIF1A,
GFAP, NES, FN1, MMP2, SNAI2, TCF4 TWIST1, showed decreased
expression by STX-0119 (Fig.
4A–C). Additionally, the effect of STX-0119 on mesenchymal
markers was moderate (Fig.
4D).
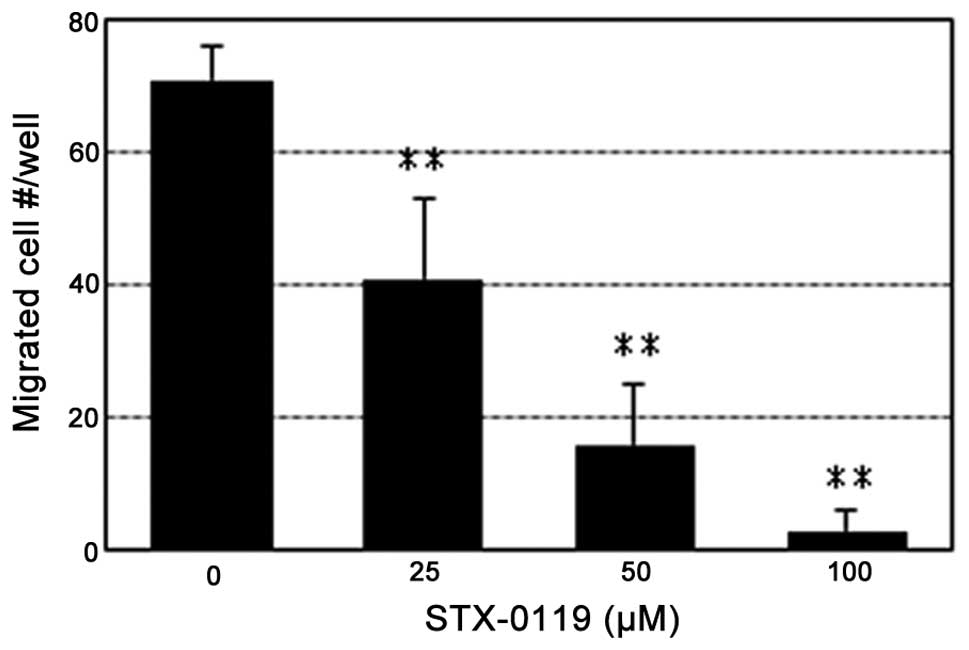
Impact of STX-0119 on invasion activity
of TMZ-R U87 cells
TMZ-R U87 cells were shown to possess greater
invasion activity than parental U87 (data not shown). The invasion
activity of TMZ-R U87 cells was reduced STX-0119 dose-dependently.
Pre-treatment of TMZ-R U87 cells with STX-0119 at 100 μM
suppressed invasion activity by more than 90% compared to the
without compound (Fig. 5).
Apoptosis induction by STX-0119 in TMZ-R
U87 cell line
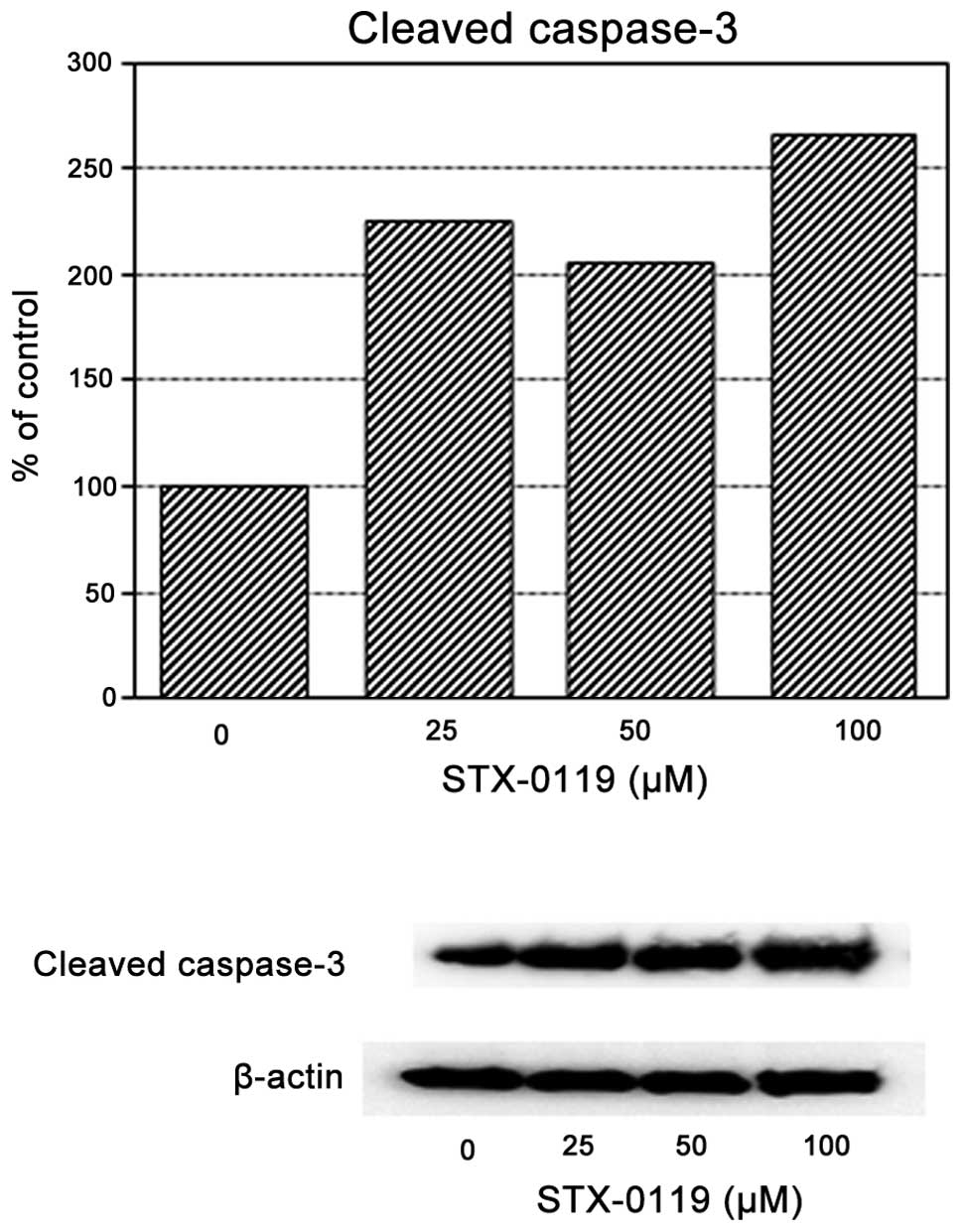
With regard to the cleaved caspase-3 level, its
expression increased at >50 μM STX-0119. TMZ-R U87 cells
treated with STX-0119 at 100 μM demonstrated an increase of
the cleaved caspase-3 expression in TMZ-R U87 cells (Fig. 6).
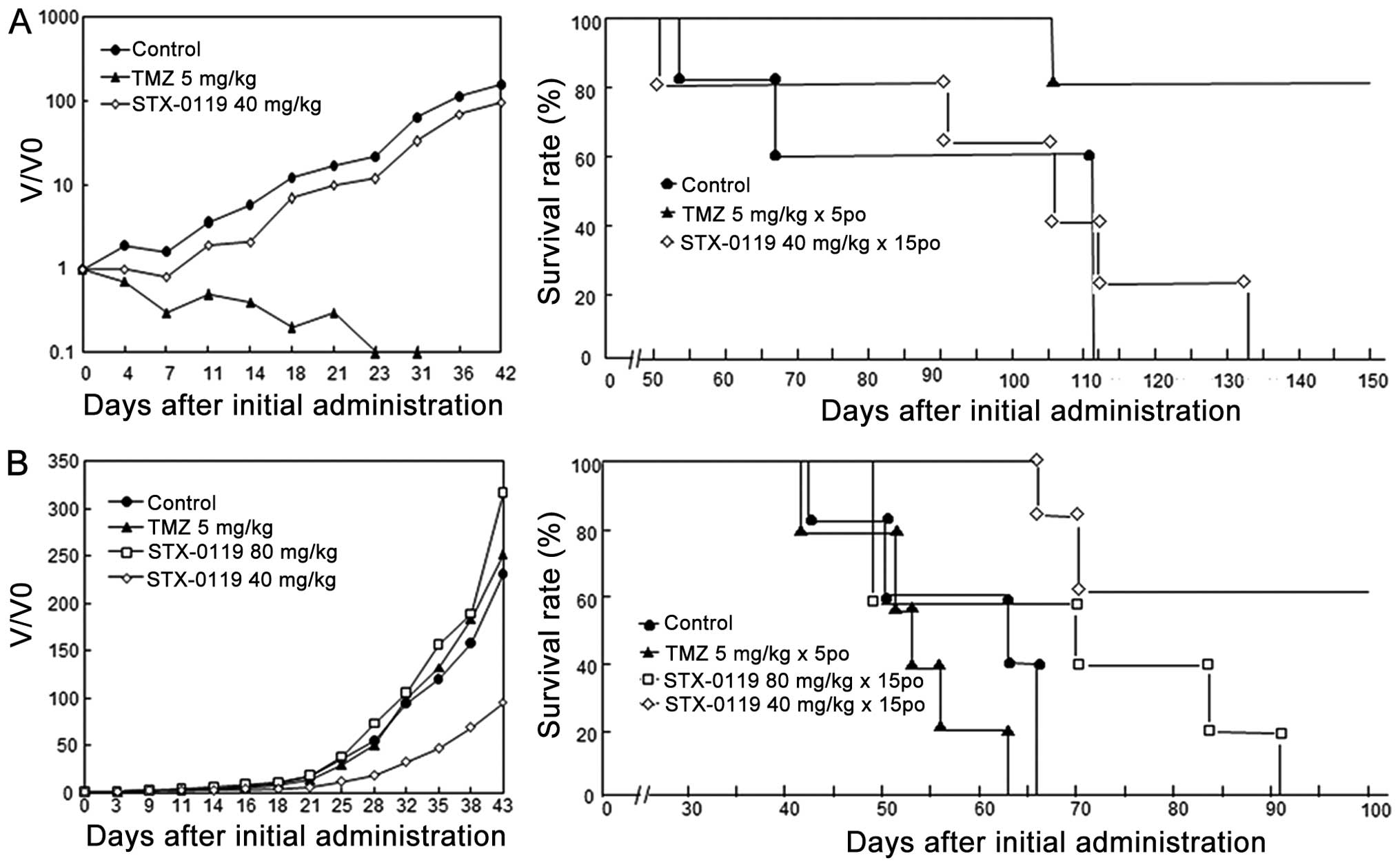
STX-0119 inhibits tumor growth of TMZ-R
U87 cells in vivo
TMZ-R U87 cell-transplanted mice showed significant
resistance to TMZ and a shorter survival time in vivo, while
parental U87 cell-transplanted mice exhibited obvious sensitivity
to TMZ. The inhibitory effect of STX-0119 on parental U87 cells was
marginal and did not show any survival benefit compared to the
control. In contrast, the growth inhibition by TMZ on U87 cells was
marked and showed no tumor recurrence until day 42 and obvious
prolongation of survival (Fig. 7,
Table I).
 | Table I.Survival analysis of U87 parental and
TMZ-R cell-derived tumors. |
Table I.
Survival analysis of U87 parental and
TMZ-R cell-derived tumors.
| Compound | Dose (mg/kg) | Frequency | Mean survival
(days) | ILS (%) | Tumor-free at 150
days |
|---|
| U87 parental cell
tumor | | | | | |
| Control | | | 91.4±28.6 | 0 | 0/5 |
| TMZ | 5 | 5p.o. QD5 | >141.2±19.7 | >54 | 4/5 (P=0.026) |
| STX-0119 | 40 | 15p.o.
(QD5/wk) | 98.6±30.6 | 8 | 0/5 (P=0.934) |
| U87 TMZ-R cell
tumor | | | | | |
| Control | | | 57.6±10.7 | 0 | 0/5 |
| TMZ | 5 | 5p.o. QD5 | 53.0±7.6 | −8 | 0/5 |
| STX-0119 | 40 | 15p.o.
(QD5/wk) | 87.2±17.6 | 51 | 3/5 (P=0.021) |
| STX-0119 | 80 | 15p.o.
(QD5/wk) | 68.6±19.4 | 19 | 0/5 (P=0.145) |
On the other hand, STX-0119 demonstrated a greater
growth-inhibitory effect on TMZ-R U87 cells with more than 50%
inhibition compared to the control. Additionally, STX-0119 showed
an obvious and more efficient prolongation of survival in TMZ-R
U87-bearing mice compared to the TMZ group, which exhibited no
growth inhibition. These effects of STX-0119 were identified in the
40 mg/kg group, but not at 80 mg/kg (Fig. 7, Table
I). In addition, STX-0119 showed no adverse effects on
tumor-bearing mice.
Discussion
High-grade gliomas including glioblastoma
multi-forme (GBM) are the most malignant and aggressive of tumors,
and have a very poor prognosis and a high recurrence rate, with a
mean survival time of less than 2 years even with the recent
development of an intensive temozolomide (TMZ)-based treatment
protocol. Additionally, frequent recurrence even after
chemo-radiation treatment is a crucial problem in the clinical
field, which should be overcome to extend the overall survival of
GBM.
O6-methylguanine-DNA-methyltransferase
(MGMT) is well known to remove methylation from the O6
position of guanine and contribute to TMZ resistance induction.
Accumulated evidence demonstrated that high MGMT expression through
the methylation of MGMT promoter is one of the mechanisms
responsible for TMZ resistance. However, recently, several
researchers demonstrated that major mechanisms other than MGMT are
involved in TMZ resistance. Several biomarkers linking to MGMT
expression and the methylation status, such as HOX signature and
EGFR expression, somatic mutation of mismatch repair gene MSH6,
prolyl 4-hydroxylase, β-polypeptide (P4HB), EGFR mutation
(EGFRvIII) and CD74, have been demonstrated. With regard to novel
approaches to overcome MGMT-related resistance to TMZ, bortezomib
(BZ) as a proteasome inhibitor (17), and even inactivation of MGMT by
gene therapy (18) have been
demonstrated to show moderate effects on MGMT downregulation and
tumor cell death.
On the other hand, a significant association between
STAT3 signaling and GBM or GBM stem cell development and
maintenance has been demonstrated in recent studies (19–23),
which emphasizes the potential of STAT3 as a therapeutic target.
Some studies demonstrated that STAT3 is required for the
proliferation and maintenance of multipotency in GBM stem cells
through siRNA-mediated STAT3 inhibition (19).
Previously, we reported that STX-0119, a novel small
molecule inhibitor of STAT3 dimerization, showed antitumor activity
in vitro and in vivo against a highly STAT3 activated
lymphoma cell line and primary GBM stem cells (24). In particular, STX-0119 showed a
growth-suppressive effect on even highly TMZ-resistant GBM stem
cells derived from a recurrent GB tumor. With regard to the
correlation of STAT3 and TMZ resistance, novel observations have
been reported. Kohsaka et al (25) reported that STAT3 inhibition
down-regulated MGMT expression, and there was a significant
positive correlation between the expression level of MGMT and
phosphorylated STAT3 in high-grade glioma tumors. Villalva et
al (26) demonstrated that a
STAT3 inhibitor, Stattic-mediated STAT3 inhibition, sensitized GBM
stem cells to the inhibitory action of TMZ with a synergistic
effect. Interestingly, intratumoral hypoxia might be a promoting
factor localized in immature GBM stem cells with high MGMT
expression, with strong resistance to TMZ and STAT3 activation in
the inner core of the tumor mass (27). This observation suggested that a
combination of TMZ and HIF-1 inhibitors may have additional
antitumor effects against GBM stem cells with TMZ resistance.
Another approach to overcoming TMZ resistance is the combination of
TMZ and interferon (IFN)-β, which is supposed to show a synergistic
antitumor effect against TMZ-resistant GBM cell lines, such as T98G
cells, through the downregulation of MGMT and STAT3 (28,29).
Motomura et al (30)
reported the benefits of IFN-β and TMZ combination therapy for
newly diagnosed GBM patients in a multicentered clinical trial; the
median survival time (MST) of the combination therapy patients was
significantly greater than the TMZ-alone group (19.9 versus 12.7
months). STAT3 inhibitor compounds including AG490, WP1066 and
Curcumin have been reported to exhibit antitumor activity through
suppression of the invasive and migratory activity of malignant
glioma cells (31–33). In particular, WP1066 was shown to
reduce intratumoral JAK2/STAT3 activity and prolong the survival of
animals given a GBM stem cell xenograft. However, these compounds
have not yet been reported to show antitumor activity against
TMZ-resistant GBM tumors.
Gene expression profiling, which can predict the
response to TMZ in high-grade gliomas, has also been performed.
Yoshino et al (34)
reported that differential gene sets identified using a DNA
microarray between TMZ-sensitive and -resistant GBM cell lines were
not causal factors in the TMZ response besides MGMT. Our similar
analysis using a DNA microarray between TMZ-sensitive and
-resistant U87 cells demonstrated that YKL-40 and MAGEC1 genes,
besides MGMT, were identified as TMZ-resistance-specific genes
(unpublished data). The precise mechanism of YKL-40 involvement in
TMZ resistance is yet to be clarified; however, considering the
downregulation of YKL-40 expression after STX-0119 treatment,
YKL-40 might be a possible surrogate marker for STAT3 targeting
TMZ-R glioblastoma.
A recent study using next-generation genome
sequencing for recurrent glioma after TMZ treatment demonstrated
that newly identified gene mutations in the recurrent tumor,
probably induced by TMZ, might be driver mutations promoting tumor
growth (35). This result strongly
suggests that novel mutations are new causal factors involved in
the TMZ resistance mechanism in recurrent glioma after TMZ
therapy.
Finally, in the current study, we established a
solid TMZ-resistant U87 cell line which showed obvious resistance
in an in vivo model and demonstrated that STX-0119 showed a
potent antitumor effect on TMZ-resistant tumors where STX-0119
significantly prolonged MST in tumor-bearing mice. These results
suggest that STX-0119 is a potential therapeutic to overcome TMZ
resistance in recurrent GBM tumors, and could be the next promising
compound leading to survival prolongation. This is the first report
that a STAT3 inhibitor small molecule compound clearly showed an
anti-tumor effect against TMZ-resistant GBM cells. Based on our new
observations regarding the efficacy mechanism of STX-0119, more
efficient compound development is expected.
Abbreviations:
|
GBM
|
glioblastoma multiforme;
|
|
TMZ
|
temozolomide;
|
|
MGMT
|
O6-methylguanine-DNA-methyltransferase;
|
|
STAT
|
signal transducer and activator of
transcription;
|
|
shRNA
|
small hairpin RNA
|
Acknowledgements
This study was supported by a grant
from a regional innovation strategy support program of the Ministry
of Education, Culture, Sports, Science and Technology, Japan.
References
|
1.
|
Zhong Z, Wen L and Darnell JE Jr: Stat3
and Stat4: members of the family of signal transducers and
activators of transcription. Proc Natl Acad Sci USA. 91:4806–4810.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Bromberg J and Darnell JE Jr: The role of
STATs in transcriptional control and their impact on cellular
functions. Oncogene. 19:2468–2473. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Yu H, Kortylewski M and Pardoll D:
Crosstalk between cancer and immune cells: role of STAT3 in the
tumor microenvironment. Nat Rev Immunol. 7:41–51. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Wang T, Niu G, Kortylewski M, et al:
Regulation of the innate and adaptive immune responses by Stat-3
signaling in tumor cells. Nat Med. 10:48–54. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Stupp R, Mason WP, van den Bent MJ, et al:
Radiotherapy plus concomitant and adjuvant temozolomide for
glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Mirimanoff RO, Gorlia T, Mason W, et al:
Radiotherapy and temozolomide for newly diagnosed glioblastoma:
recursive partitioning analysis of the EORTC 26981/22981-NCIC CE3
phase III randomized trial. J Clin Oncol. 24:2563–2569. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Brennan C, Momota H, Hambardzumyan D,
Ozawa T, Tandon A, Pedraza A and Holland E: Glioblastoma subclasses
can be defined by activity among signal transduction pathways and
associated genomic alterations. PLoS One. 4:e77522009. View Article : Google Scholar
|
|
8.
|
Cheng CK, Fan QW and Weiss WA: PI3K
signaling in glioma-animal models and therapeutic challenges. Brain
Pathol. 19:112–120. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Demuth T, Reavie LB, Rennert JL, et al:
MAP-ing glioma invasion: mitogen-activated protein kinase 3 and p38
drive glioma invasion and progression and predict patient survival.
Mol Cancer Ther. 6:1212–1222. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Pu P, Zhang Z, Kang C, Jiang R, Jia Z,
Wang G and Jiang H: Downregulation of Wnt2 and beta-catenin by
siRNA suppresses malignant glioma cell growth. Cancer Gene Ther.
16:351–361. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Purow BW, Haque RM, Noel MW, et al:
Expression of Notch-1 and its ligands, Delta-like-1 and Jagged-1,
is critical for glioma cell survival and proliferation. Cancer Res.
65:2353–2363. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Hegi ME, Diserens AC, Gorlia T, et al:
MGMT gene silencing and benefit from temozolomide in glioblastoma.
N Engl J Med. 352:997–1003. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Murat A, Migliavacca E, Gorlia T, et al:
Stem cell-related ‘self-renewal’ signature and high epidermal
growth factor receptor expression associated with resistance to
concomitant chemoradiotherapy in glioblastoma. J Clin Oncol.
26:3015–3024. 2008.
|
|
14.
|
Hunter C, Smith R, Cahill DP, et al: A
hypermutation phenotype and somatic MSH6 mutations in recurrent
human malignant gliomas after alkylator chemotherapy. Cancer Res.
66:3987–3991. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Mukherjee B, McEllin B, Camacho CV, et al:
EGFRvIII and DNA double-strand break repair: a molecular mechanism
for radioresistance in glioblastoma. Cancer Res. 69:4252–4259.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Ashizawa T, Miyata H, Ishii H, et al:
Antitumor activity of a novel small molecules STAT3 inhibitor
against a human lymphoma cell line with high STAT3 activation. Int
J Oncol. 38:1245–1252. 2011.PubMed/NCBI
|
|
17.
|
Vlachostergios PJ, Hatzidaki E, Befani CD,
Liakos P and Papandreau CN: Bortezomib overcomes MGMT-related
resistance of glioblastoma cell lines to temozolomide in a
schedule-dependent manner. Invest New Drugs. 31:1169–1181. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Jiang G, Wei ZP, Pei DS, Xin Y, Liu YQ and
Zheng JN: A novel approach to overcome temozolomide resistance in
glioma and melanoma: Inactivation of MGMT by gene therapy. Biochem
Biophys Res Commun. 406:311–314. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Li GH, Wei H, Lv SQ, Ji H and Wang DL:
Knockdown of STAT3 expression by RNAi suppresses growth and induces
apoptosis and differentiation in glioblastoma stem cells. Int J
Oncol. 37:103–110. 2010.PubMed/NCBI
|
|
20.
|
See AP, Han JE, Phallen J, et al: The role
of STAT3 activation in modulating the immune microenvironment of
GBM. J Neurooncol. 110:359–368. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Sherry MM, Reeves A, Wu JK and Cochran BH:
STAT3 is required for proliferation and maintenance of multipotency
in glioblastoma stem cells. Stem Cells. 27:2383–2392. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Liu Y, Li C and Lin J: STAT3 as a
therapeutic target for glioblastoma. Anticancer Agents Med Chem.
10:512–519. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Jackson C, Ruzevick J, Amin AG and Lim M:
Potential role for STAT3 inhibitors in glioblastoma. Neurosurg Clin
N Am. 23:379–389. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Ashizawa T, Miyata H, Iizuka A, et al:
Effect of the STAT3 inhibitor STX-0119 on the proliferation of
cancer stem-like cells derived from recurrent glioblastoma. Int J
Oncol. 43:219–227. 2013.PubMed/NCBI
|
|
25.
|
Kohsaka S, Wang L, Yachi K, et al: STAT3
inhibition overcomes temozolomide resistance in glioblastoma by
donregulating MGMT expression. Mol Cancer Ther. 11:1289–1299. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Villalva C, Martin-Lanneree S, Cortes U,
et al: STAT3 is essential for the maintenance of
neurosphere-initiating tumor cells in patients with glioblastomas:
a potential for targeted therapy? Int J Cancer. 128:826–838. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Pistollato F, Abbadi S, Rampazzo E, et al:
Intratumoral hypoxic gradient drives stem cells distribution and
MGMT expression in glioblastoma. Stem Cells. 28:851–862.
2010.PubMed/NCBI
|
|
28.
|
Yuki K, Natsume A, Yokoyama H, et al:
Induction of oligodendrogenesis in glioblastoma-initiating cells by
IFN-mediated activation of STAT3 signaling. Cancer Lett. 284:71–79.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Yoshino A, Ogino A, Yachi K, et al: Effect
of IFN-β on human glioma cell lines with temozolomide resistance.
Int J Oncol. 35:139–148. 2009.
|
|
30.
|
Motomura K, Natsume A, Kishida Y, et al:
Benefits of interferon-β and temozolomide combination therapy for
newly diagnosed primary glioblastoma with the unmethylated MGMT
promoter: A multicenter study. Cancer. 117:1721–1730. 2011.
|
|
31.
|
Senft C, Priester M, Polacin M, Schroder
K, Seifert V, Kogel D and Weissenberger J: Inhibition of the
JAK-2/STAT3 signaling pathway impedes the migratory and invasive
potential of human glioblastoma cells. J Neurooncol. 101:393–403.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Stechishin OD, Luchman HA, Ruan Y, et al:
On-target JAK2/STAT3 inhibition slows disease progression in
orthotopic xenografts of human glioblastoma brain tumor stem cells.
Neuro Oncol. 15:198–207. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Senft C, Polacin M, Priester M, Seifert V,
Kogel D and Weissenberger J: The nontoxic natural compound Curcumin
exerts anti-proliferative, anti-migratory, and anti-invasive
properties against malignant gliomas. BMC Cancer. 10:4912010.
View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Yoshino A, Ogino A, Yachi K, et al: Gene
expression profiling predicts response to temozolomide in malignant
gliomas. Int J Oncol. 36:1367–1377. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Johnson BE, Mazor T, Hong C, et al:
Mutational analysis reveals the origin and therapy-driven evolution
of recurrent glioma. Science. 343:189–193. 2013. View Article : Google Scholar : PubMed/NCBI
|