Introduction
Colon cancer is one of the most common causes of
cancer related mortality worldwide (1). Its overall incidence is ~5% and the
5-year survival rate ranges from 40 to 60% (2). The mechanisms of colon cancer
carcinogenesis and development have drawn much attention in recent
years for its high incidence and poor prognosis. However, the
etiology and pathogenesis of colon cancer remain unclear, although
heterogeneous genetic alterations have been reported to be
responsible. The most common genetic alterations include mutations
and loss of heterozygosity of tumor suppressors, such as
adenomatous polyposis coli (APC) (3). Several studies have demonstrated that
colon cancers were involved in the activation of Wnt, Notch, BMP
and Hedgehog signaling pathways, which accompany establishment of
tumorigenic state (4). Msi1 has
been found to activate the Notch and Wnt signaling pathways in
several types of normal and cancerous cells (5,6).
Therefore, we hypothesized that Msi1 maybe have association with
the progression of colon cancer.
Msi1, located at chromosome 12q24, is a member of
the Musashi (MSI) family of the RNA binding proteins, which are
characterized by RNA recognition motif. It can be regulated by ELAV
or HuR by maintaining the stabilization of its mRNA or accumulation
of mRNA translation (7,8), as well as by tumor suppressor
microRNAs (9). It was first found
in Drosophila as a determinant of sensory organ development
(10) and highly conserved across
species (11) with essential roles
in the stem cell maintenance, nervous system development and
tumorigenesis (12,13). In mammals, Msi1 is highly expressed
in the neural progenitor and stem cells (13) for maintenance of self-renewal and
differentiation. Recent studies have demonstrated that Msi1 exists
in many adult cancers, including gliomas (14), gastric cancer (15), hepatoma (16) and colorectal adenoma (17). Consistently, high level of Msi1
expression is considered to be associated with many malignancies
(18) and predicts poor prognosis
(19,20). All these finding suggest that Msi1
function as an oncogene. This notion is further supported by the
findings that ablation of Msi1 in gliomas (6) or bladder carcinoma cells (21) suppressed the cell cycle, and
induced apoptosis and showed severe decline in cell numbers.
However, Msi1 has also been proposed to have no significant effect
on cell proliferation in human intestinal epithelial cells
(22).
We performed detailed analyses on the role of Msi1
in colon carcinoma, and found that Msi1 was upregulated. Knockdown
of endogenous Msi1 protein inhibited the growth and tumor formation
by activating the cell cycle suppressor
p21cip. Our findings support the
hypothesis that Msi1 functions as an oncogene in colon cancer.
Materials and methods
Cell lines
Human colon cancer cell lines SW480, HCT116, SW620
and cervical cancer cell line HeLa were purchased from the American
Type Culture Collection (ATCC), cultured in Dulbecco’s modified
Eagle’s medium (DMEM; Sigma-Aldrich, St. Louis, MO, USA)
supplemented with 10% fetal bovine serum (FBS; Invitrogen,
Carlsbad, CA, USA) and 1% penicillin-streptomycin. All cell lines
were maintained at 37°C in an atmosphere of 5% CO2.
Tissue samples
Fresh frozen specimens of matched colon cancer
tissues and adjacent non-tumor tissues were from 20 patients in the
Department of General Surgery of the Fourth Hospital of Hebei
Medical University, obtained between March 2011 and January 2013,
and stored at −80°C for further use. All of the tissues were from
untreated patients who were undergoing surgery. Written informed
consent was obtained from each patient before the surgery, and the
study protocol was approved by the Institutional Research Ethics
Committee.
Semi-quantitative real-time PCR
analysis
Total RNA was isolated from colon cancer cell lines
using TRIzol reagent (Invitrogen). Total cDNA was synthesized using
the M-MLV Reverse Transcriptase (Fermentas, Vilnius, Lithuania).
The mRNA expression levels were measured by qRT-PCR using the 7500
Real-Time PCR detection system (Applied Biosystems, Foster City,
CA, USA). Amplification was performed with the SYBR Premix Ex Taq™
II kit (code: DRR081A; Takara, Dalian, China) according to a 2-step
cycle procedure consisting of 45 cycles of denaturation at 95°C for
10 sec and annealing/elongation at 60°C for 30 sec. We measured
mRNA levels semi-quantitatively by the Δ/Δ threshold cycle (Ct)
method. GAPDH was used as internal control. Fold-changes were
calculated and normalized as relative to the parental cells. The
primers were used as follows: p21cip1 F,
5′-AAATCGTCCAGCGACCTTCC-3′ and p21cip1 R,
5′-GCCCTGTCCATAGCCTCTACT-3′; GAPDH F, 5′-CAAG GGCATCCTGGGCTACA-3′,
GAPDH R, 5′-AAGTGGTCG TTGAGGGCAAT-3′.
Western blot analysis
Cells and clinical tissues were lysed on ice in
lysis buffer (50 mM Tris-HCl, pH 7.4; 150 mM NaCl; 2 mM EDTA; 1%
NP-40; and 0.1% sodium dodecyl sulfate SDS) containing freshly
added protease inhibitor cocktail (Complete Mini; Roche
Diagnostics, Branchburg, NJ, USA). Aliquots of samples with the
same amount of protein were determined using the Bradford assay
(Bio-Rad Laboratories, Hercules, CA, USA), and mixed with loading
buffer (final concentrations of 62.5 mM Tris-HCl, pH 6.8, 2.3% SDS,
100 mM dithiothreitol and 0.005% bromophenol blue). Then the
protein extracts were boiled and separated by SDS-PAGE and blotted
to activated polyvinylidene difluoride (PVDF) membranes (Millipore,
Billerica, MA, USA). Then the membranes were blocked with 5%
fat-free milk and probed with first antibodies overnight. The first
antibodies included the following: anti-Msi1 (1:500, sc-98845;
Santa Cruz Biotechnology, Santa Cruz, CA, USA), anti-p21 (1:500,
sc-397; Santa Cruz Biotechnology), and anti-β-actin (1:500,
sc-47778; Santa Cruz Biotechnology). The membranes were then washed
four times in PBST and incubated with a secondary antibody coupled
to horseradish peroxidase (Thermo Fisher Scientific, Inc., New
York, NY, USA), followed by ECL detection (Millipore) and
visualization on X-ray film. Relative quantitation was determined
with the AlphaView system (Cell Biosciences, Santa Clara, CA, USA)
using β-actin as the loading control.
Plasmid construction
To generate plasmids that express Msi1-specific
small interfering RNA (siRNA) which targeting MSI1, the
oligonucleotide inserts were designed using an online siRNA design
tool (Ambion, Austin, TX, USA). The double oligonucleotides were
annealed and cloned into the lentivirus GV248-GFP shRNA vector
(Shanghai Genechem Co., Ltd., Shanghai, China) as per the
manufacturer’s introductions. Lentivirus was produced after
GV248-GFP shRNA vector and two package plasmids were co-transfected
to 293T cells with Lipofectamine 2000 transfection reagent
(Invitrogen). Then colon cancer cells were infected by recombined
lentivirus, and cell clones were selected to the amplification
culture. Stably transfected clones were extracted and identified by
western blotting.
A luciferase reporter vector (pMIR-REPORT; Ambion)
was used to generate reporter constructs. Fragment of the 3′ UTR of
human p21cip1 was amplified from HCT116
cDNA by PCR using primers p21cip1 WT F/R
and cloned into the pMIR-REPORT luciferase plasmid. The mutant
p21cip1 3′ UTR was generated by using the
p21cip1 WT F/MU R and
p21cip1 MU F/WT R primers. The following
primers were used: shMsi1-1F, 5′-GATCCTGTTACATGGTGTTTCGAATTCAAGA
GATTCGAAACACCATGTAACATCA-3′ and shMsi1-1R,
5′-GACAATGTACCACAAAGCTTAAGTTGAGAAAGCT TTGTGGTACATTGTAGTTCGA-3′;
shMsi1-2F, 5′-GATCC TCCTGTATCATATGTAAATTTCAAGAGAATTTACATA
TGATACTGGACGA-3′ and shMsi1-2R, 5′-AGGACATAGT
ATACATTTAAAGTTCTCTTAAATGTATACTATGACCT GCTTTCGA-3′;
p21cip1 WT F, 5′-ATTGAGCTCTAATCCGC
CCACAGGAAG-3′ and p21cip1 WT R,
5′-CTCAAGCTTACA AGTAAAGTCACTAAG-3′;
p21cip1 MU F, 5′-TGGGAAGCA
GTGTCTTTCCTGGCACTAACGTT-3′ and p21cip1 MU
R, 5′-AGACACTGCTTCCCAGCCCCATATGAGCCCAC-3′.
Cell proliferation and cell viability
assays
Cells (5×104) were cultured in triplicate
in 35-mm cell culture dishes. The cells were harvested
longitudinally, and counted every day for one week using a
hemocytometer under light microscopy.
Cell viability was assessed every other day using
3-(4,5-dimethylthiazol-yl)-2,5-diphenyl tetrazolium bromide (MTT;
Sigma-Aldrich) dye according to the standard protocol.
Approximately 1,000 cells/well were seeded in a 96-well plate. MTT
solution (20 μl) was added to 200 μl of culture media and incubated
for 4 h, and then cell growth was determined by measuring the
absorbance at 490 nm.
Colony formation assay
Cells (1×103) of each type were cultured
in 10-cm culture dishes and exposed to fresh media every 3 days.
Colonies with diameters >0.2 mm were counted at day 14.
Tumorsphere formation assay
One hundred Msi1-KD or -control cells were seeded in
6-well plates in 2 ml of serum-free stem cell medium as described
above. Fresh medium were added to each well every 3 days. The
tumorspheres were analyzed on day 14 for tumorsphere forming
ability.
Animal and tumor xenograft assay
Exponentially growing cells collected from stable
transfectants were bilaterally inoculated into subcutaneous sites
of 4- to 6-week-old Balb/c nude mice. Tumor dimensions were
measured with calipers once every week, and volumes
(cm3) were calculated according to the standard formula,
length × width2/2. At the end of the experiment, tumors
were dissected out, and the net weight per mouse was measured. The
experimental protocols used herein were evaluated and approved by
the Animal Care and Use Committee of the Fourth Hospital of Hebei
Medical University.
Cell cycle analysis
Cells (1×106) were harvested and washed
twice with PBS, followed by fixation with 75% cold ethanol at 4°C
overnight. After the cells were washed twice in PBS, the cells were
suspended in PBS with 50 μg/ml propidium iodide (PI; Sigma) and 10
μg/ml RNaseA (Sigma). They were then incubated at 4°C for 30 min in
the dark. Cells were analyzed for DNA content by FACSCalibur (BD
Biosciences) and the results of the cell cycle were analyzed by
FlowJo software.
Luciferase assay
Cells were lysed in 100 μl of passive lysis buffer
(Promega, Madison, WI, USA) at 48 h after transfection and assayed
with a Dual-luciferase report assay kit according to the
manufacturer’s instructions. For each assay, cell extract (20 μl)
was used and the reaction was started by injection of 50 μl of
luciferase substrate. Each reaction was measured for 10 sec in the
Luminometer. Luciferase activities were expressed as the ratio of
firefly to Renilla luciferase activity.
Statistical analysis
Data are presented as the mean ± standard deviation
(SD). The Student’s t-test was performed using the SPSS 16.0 (SPSS,
Inc., Chicago, IL, USA). P<0.05 was regarded as statistically
significant.
Results
Msi1 expression in human colon cancer
tissues and cell lines
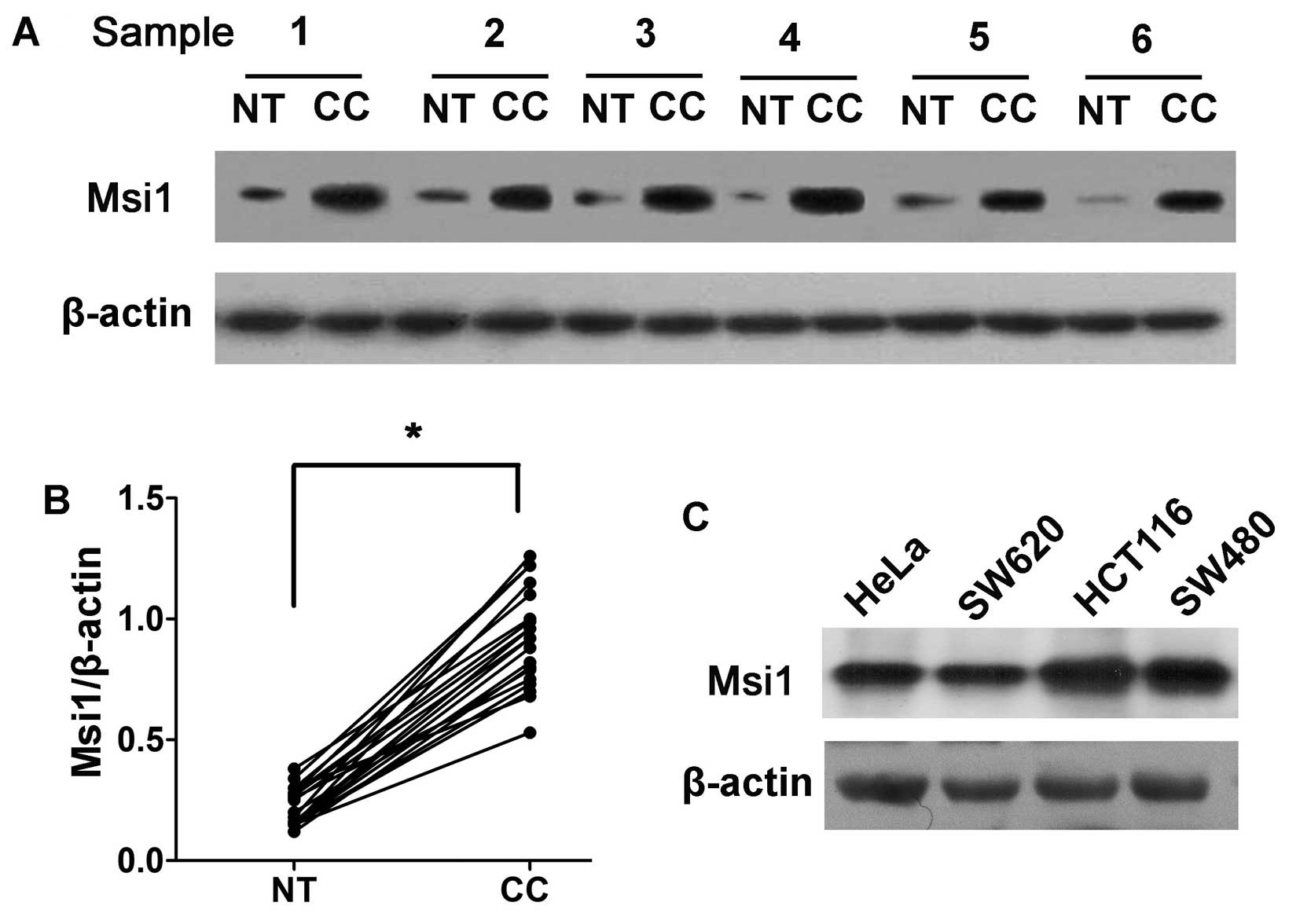
To assess Msi1 expression in clinical patients,
western blot analysis was conducted on 20 pairs of colon cancer
tissues and matched adjacent non-tumor tissues (Fig. 1A). Msi1 expression in all 20 cancer
tissues was markedly higher than that in the corresponding
non-tumor tissues (Student’s t-test, P<0.01; Fig. 1B), suggesting that the activation
of Msi1 may contribute to the development and progression of colon
cancer.
We also evaluated the expression of Msi1 in colon
cancer cell lines by western blotting. We detected a high level of
Msi1 expression in SW620, HCT116 and SW480 cells (Fig. 1C).
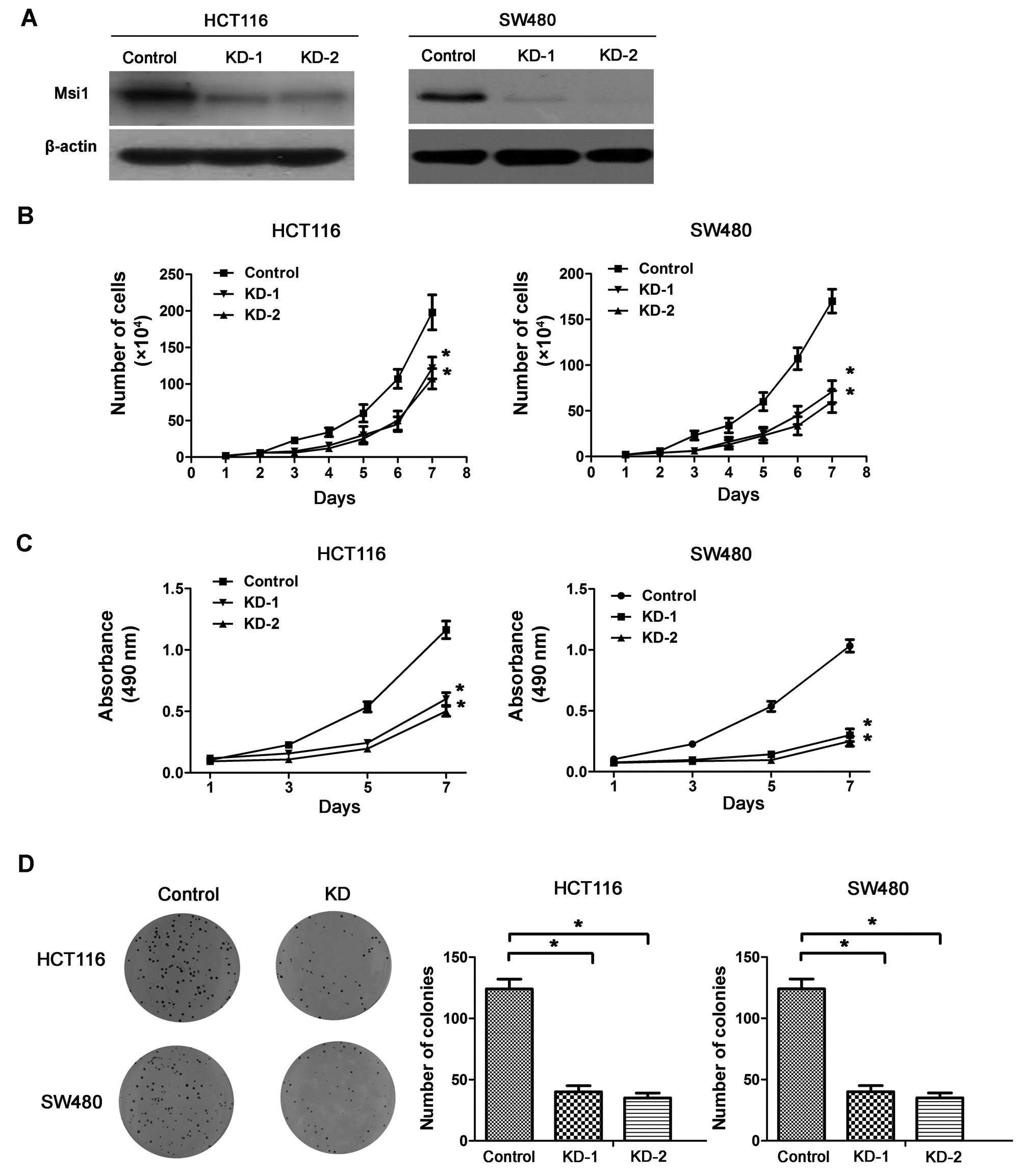
The knockdown of Msi1 inhibits the
proliferation of colon cancer cells in vitro
To determine whether Msi1 regulated the
proliferation of colon cancer cell lines, the endogenous Msi1 was
knocked down by short hairpin RNA (shRNA) in HCT116 and SW480 cells
(Fig. 2A). The Msi1 knockdown
HCT116 and SW480 cells (HCT116-KD and SW480-KD) had much lower
proliferation ability than the corresponding control cells
(HCT116-control and SW480-control), as measured by both cell growth
curve assay (P<0.01; Fig. 2B)
and cell viability assay (P<0.01; Fig. 2C), indicating that the knockdown of
Msi1 suppressed the growth of colon cancer cells in
vitro.
Furthermore, colony formation assays showed that the
efficiency of foci formation was dramatically decreased in Msi1
knockdown clones compared with control clones (P<0.01; Fig. 2D). These findings suggested that
enforced inhibition of Msi1 led to retardation of colon cancer cell
growth in vitro.
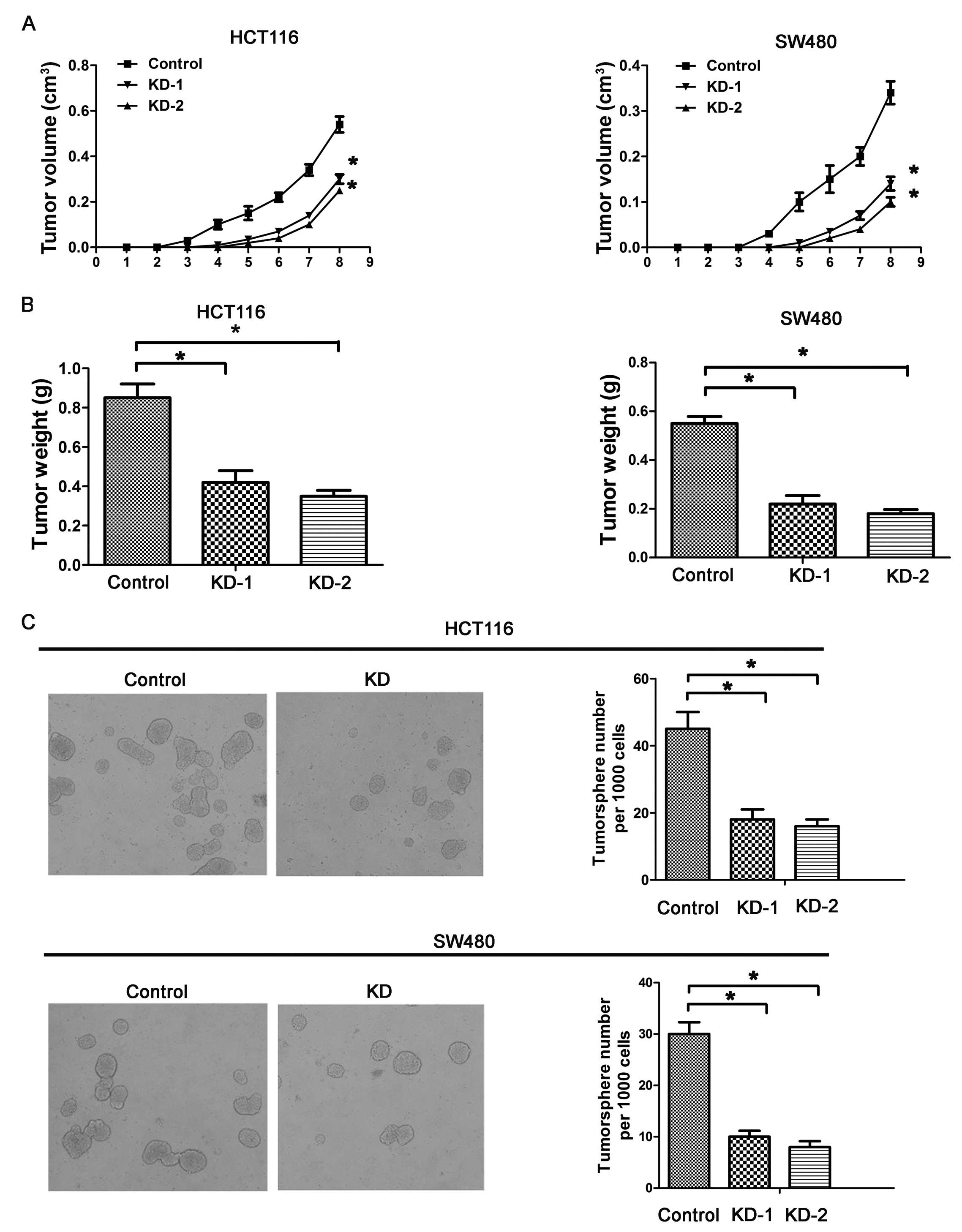
Silencing of Msi1 inhibits the tumor
formation of colon cancer cells in vivo and in vitro
As Msi1 modulates the proliferation in vitro,
we explored whether it alters tumorigenic potential of colon cancer
cells in vivo. Xenograft assays were created with nude mice,
and the development and growth of solid tumors were monitored every
week. The palpable tumor formation of HCT116-KD and HCT116-control
cells occurred at the same time after inoculation. However, tumor
development with HCT116-KD cells was significantly delayed
(P<0.01; Fig. 3A). The tumor
weights generated from HCT116-KD were reduced significantly
(P<0.01; Fig. 3B). Similar
results were observed from SW480 cells (Fig. 3A and B), suggesting that the
silencing of Msi1 induces the suppression of the tumor formation
and development of colon cancer, and this inhibition may have a
close relationship with the decreased cell proliferation. Together,
these data indicated that knockdown of Msi1 expression inhibited
the growth of colon cancers.
Several studies have recently indicated the
existence of CSCs (cancer stem cells) in various cancer types
(23–25), and CSCs are critical for the
maintenance of tumor growth, progression and resistance to
chemotherapy or radiotherapy, as well as recurrence and metastasis
(26). It has been demonstrated
that Msi1 regulates the proliferation and differentiation of CSCs.
In the present study, we used a tumorsphere culture system to
conduct the latent role of Msi1 on tumorsphere formation. Knockdown
of Msi1 inhibited the tumorsphere formation and growth of HCT116-KD
and SW480-KD cell lines, as demonstrated by the significantly
reduced numbers of tumorspheres compared with the numbers in
control cells (P<0.01; Fig.
3C). Our results indicate that Msi1 is a key regulator of the
growth of tumorspheres.
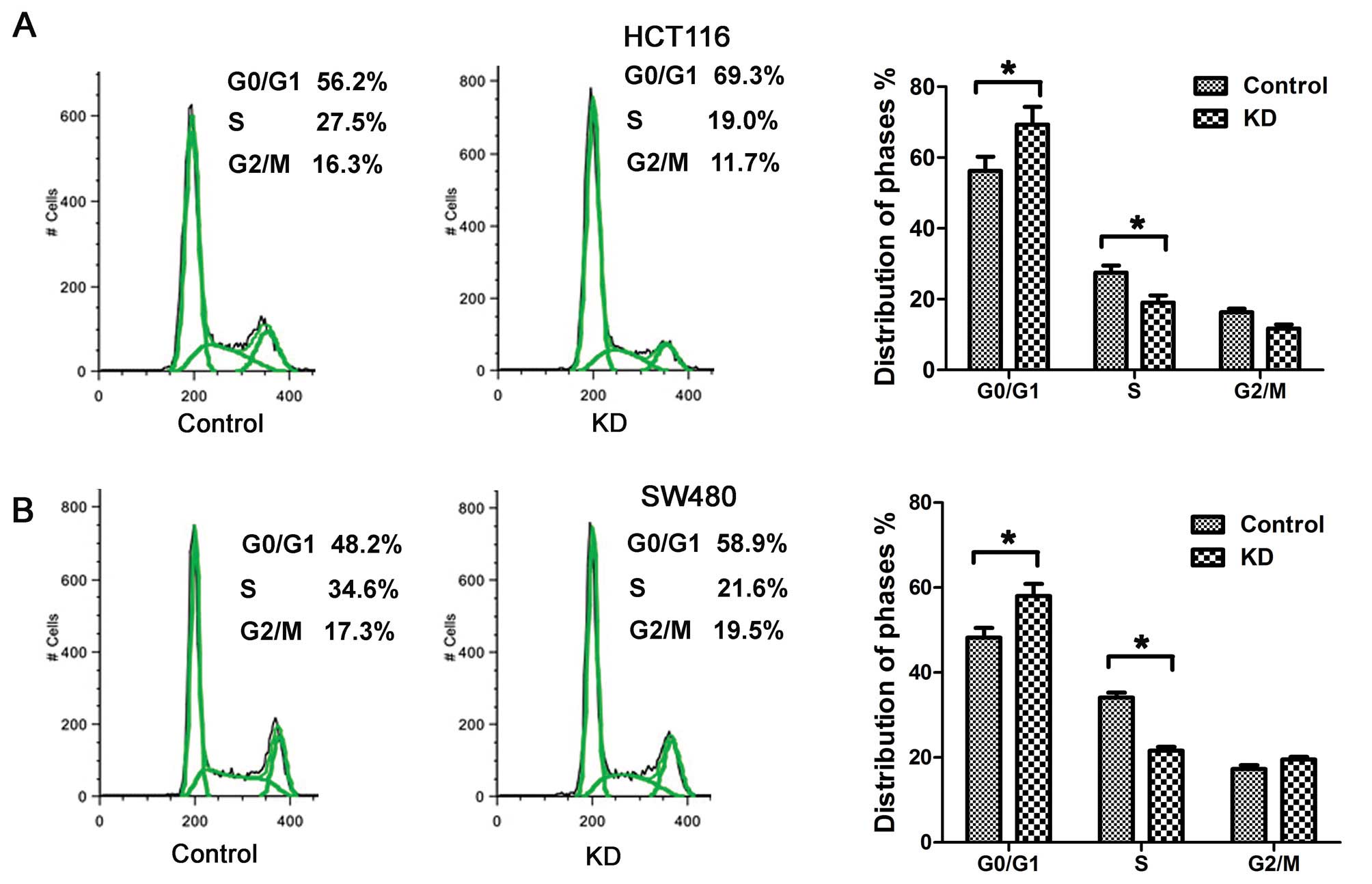
Msi1 regulates the cell cycle of colon
cancer cells in vitro
To further investigate the potential mechanism by
which silence of Msi1 regulates growth of colon cancer cells, we
characterized cell cycles by FACS analysis. As shown in Fig. 4A, the proportion of HCT116-KD cells
in G0/G1 phase increased markedly to 69.3%,
while proportion in S phase decreased to 19.0%. The ratio of
G1 phase to S phase of HCT116-KD cells was much higher
than that of HCT116-control cells. A similar effect was observed in
SW480-KD cells (P<0.01; Fig.
4B), indicating that the knockdown of Msi1 expression blocked
G1/S phase transition of cell cycle.
Msi1 negatively regulates
p21cip1 expression
It has been reported that Msi1 blocked the
G1/S phase transition of cell cycle by negative
regulation of cyclin-dependent kinase (CDK) inhibitor
p21cip1 in bladder carcinoma (21) and breast cancer (19). To test whether it also occurs in
colon cancer cells, p21cip1 mRNA and
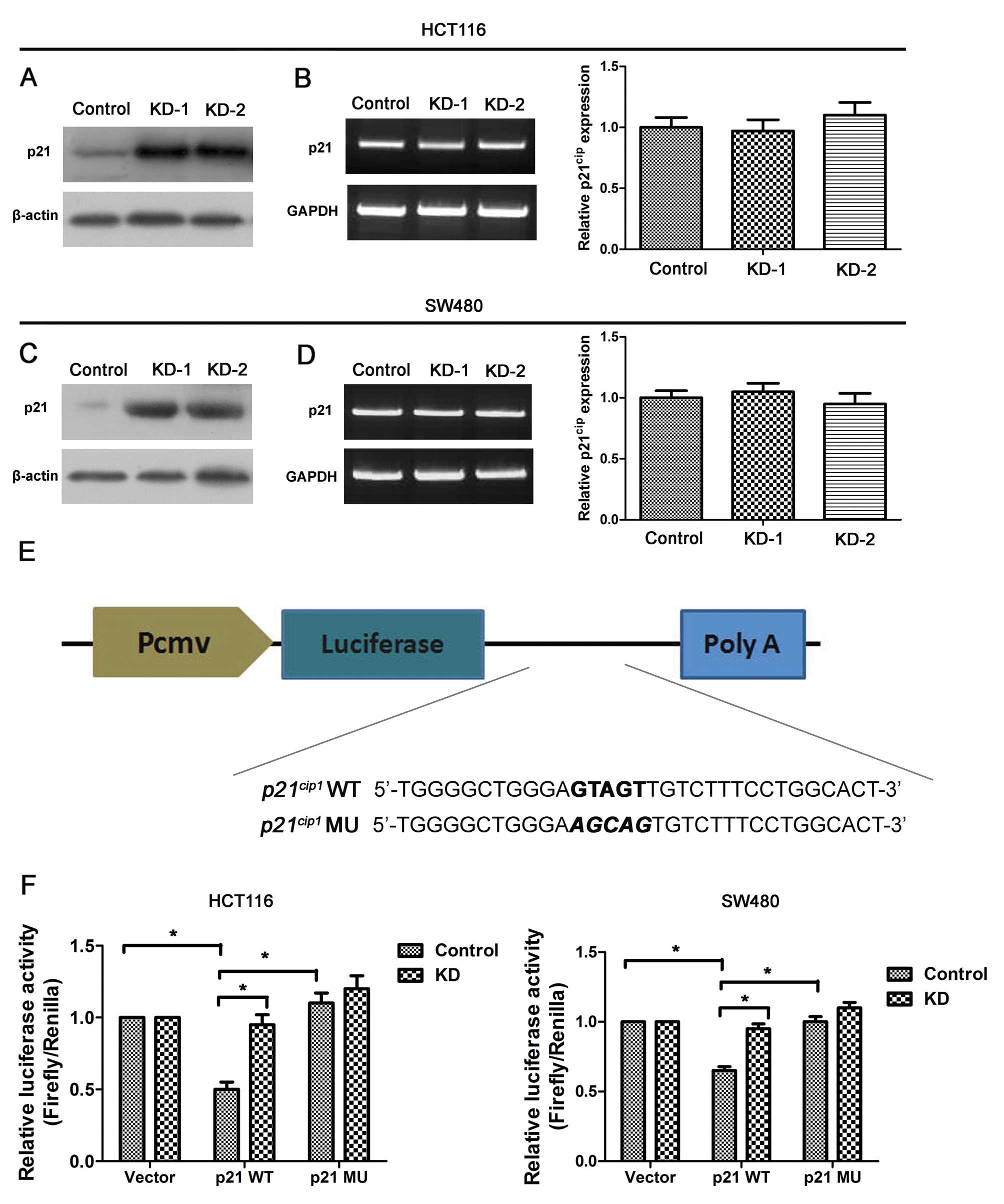
protein levels were examined by qRT-PCR and western blot analysis.
Significantly, increased levels of p21cip1
protein were observed in the lysates of both HCT116-KD and SW480-KD
cells, but there was little change in
p21cip1 mRNA levels, suggesting that Msi1
may reduce p21cip1 protein levels by
inhibiting translation.
To confirm the direct interaction between Msi1 and
its putative target site, the human
p21cip1 3′ UTR containing the wild-type
and mutant Msi1 binding sequence was cloned downstream from the
luciferase report element (Fig.
5E). The relative luciferase activity of wild-type
p21cip1 3′ UTR reporter was increased by
48% in HCT116-KD cells relative to controls, while the augmentation
was diminished in cells transfected with the mutant reporter
(P<0.01; Fig. 5F). Similarly,
knockdown of Msi1 enhanced the relative luciferase activity of the
wild-type in SW480 cells (P<0.01; Fig. 5F). Mutation of the Msi1 binding
site abrogated the elevate effect, demonstrating the specificity of
the Msi1 target sequence.
Discussion
Msi1, a multifunction RNA binding protein of MSI
family, is present in many types of normal cells (27–29)
and is involved in CNS stem/progenitor cell fate (13,30),
inner ear development (31),
repair of small-intestinal and stomach injury (32,33),
and maturation of photoreceptor and oocyte (34,35),
intestinal metaplasia (36),
atherosclerotic arteries (37) and
Alzheimer disease and Pick disease (38). Its overexpression in tumor cells
promotes tumor growth, invasion and metastasis, inhibits apoptosis
and predicts poor prognosis. The pathophysiological functions
involved have been found in several types of cancers, such as
glioma (6), medulloblastoma
(39) and breast cancer (19).
In the present study, we detected the expression of
Msi1 in colon cancer tissues. Msi1 protein was abundant in colon
cancer tissues. Low levels of Msi1 can be detected in matched
adjacent non-tumor tissues. Nishimura and co-workers (40) found that Msi1 were located in the
human normal colon crypt cells, indicating that it may be a
possible stem cell marker of human colon epithelium. The low levels
of Msi1 in normal colon tissues suggests that Msi1 may participate
in the physiological function of the colon, and may be involved in
the proliferation and initiation of the cells. In addition, colon
cancer tissues have a significantly higher expression than normal
tissues, indicating Msi1 plays important roles in colon cancers.
Thus, we hypothesized that Msi1 was associated with colon cancer
stem cells.
To further explore how Msi1 is involved in colon
carcinogenesis, functional characterization was conducted by
downregulation of Msi1 in colon cancer cell lines. We found that
knockdown of Msi1 significantly inhibited cell proliferation and
colony formation, induced cell cycle arrest in
G0/G1 phase, demonstrating that Msi1 related
signaling is a crucial regulator of the development and growth of
colon cancers. Our data are in agreement with previous findings in
other types of tumors (6,19,39),
suggesting that Msi1 related signaling may have a similar
regulatory effect on the growth of different types of human
malignancies.
Several studies have recently suggested the
existence of CSC in colon cancers (41,42).
Cancer stem cells are a rare cell population, which have the
ability for self-renewal, differentiation and tumorigenesis
(43). Recent research has found
that cancer stem cells are rich in tumorspheres and give rise to
tumors (44). Indeed, it has been
reported that Msi1 related signaling regulates the proliferation of
normal and cancer stem cells (12,30).
We characterized the role of Msi1 in the proliferation of colon CSC
by using tumor xenograft and tumorsphere formation assays and
observed that the knockdown of Msi1 expression significantly
suppressed the tumor formation in vivo and remarkably
decreased the number of tumorspheres formed in culture conditions
in vitro that allowed the proliferation of only CSC.
Therefore, it is possible that Msi1 related signaling may regulate
the growth of human colon cancers by promoting the proliferation of
both CSC and cancer cells.
Several lines of evidence suggest that Msi1 is a
positive regulator of cell proliferation (45). Msi1 promotes cell proliferation
through downregulation of the inhibitors of proliferation (6). In the present study, we also found
that downregulation of Msi1 inhibited the cell cycle by blocking
the G1/S transition in 2 colon cancer cell lines.
Furthermore, we found that knockdown of Msi1 upregulated the
expression of p21cip1. Luciferase assay
revealed that Msi1 suppressed p21cip1
expression by directly binding to the consensus sequence of
p21cip1 3′UTR in colon cancer cells. These
results are consistent with the findings in bladder carcinoma
(21) and breast cancer (19). p21cip1 is
a universal cyclin dependent kinase (CDK) inhibitor that directly
inhibits the activity of cyclin-CDK complexes, resulting in cell
cycle arrest at G0/G1 phase. Therefore, our
data suggested that cell growth retardation induced by
downregulation of Msi1 in colon cancer is probably through
activation of p21cip1.
In summary, we showed that Msi1 was upregulated in
colon cancers, and knockdown of Msi1 inhibited the proliferation
involved the coordinated direct activation of CDK inhibitor
p21cip1. The present study indicates that
downregulating the expression or restoring the expression of
p21cip1 is a possible method to treat
certain types of colon cancers.
Acknowledgements
The present study was partially supported by a grant
from the National Natural Science Foundation of China (grant no.
31271223), and the Hebei Provincial Medical Foundation (grant no.
ZL20140107). We gratefully thank Professor Depei Li for providing
invaluable assistance.
References
|
1
|
Weitz J, Koch M, Debus J, Hohler T, Galle
PR and Buchler MW: Colorectal cancer. Lancet. 365:153–165. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Siegel R, Ward E, Murray T, Xu J
and Thun MJ: Cancer statistics, 2007. CA Cancer J Clin. 57:43–66.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fodde R, Smits R and Clevers H: APC,
signal transduction and genetic instability in colorectal cancer.
Nat Rev Cancer. 1:55–67. 2001. View
Article : Google Scholar
|
|
4
|
Bertrand FE, Angus CW, Partis WJ and
Sigounas G: Developmental pathways in colon cancer: crosstalk
between WNT, BMP, Hedgehog and Notch. Cell Cycle. 11:4344–4351.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang XY, Yin Y, Yuan H, Sakamaki T, Okano
H and Glazer RI: Musashi1 modulates mammary progenitor cell
expansion through proliferin-mediated activation of the Wnt and
Notch pathways. Mol Cell Biol. 28:3589–3599. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Muto J, Imai T, Ogawa D, et al:
RNA-binding protein Musashi1 modulates glioma cell growth through
the post-transcriptional regulation of Notch and PI3 kinase/Akt
signaling pathways. PLoS One. 7:e334312012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ratti A, Fallini C, Cova L, et al: A role
for the ELAV RNA-binding proteins in neural stem cells:
stabilization of Msi1 mRNA. J Cell Sci. 119:1442–1452. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vo DT, Abdelmohsen K, Martindale JL, et
al: The oncogenic RNA-binding protein Musashi1 is regulated by HuR
via mRNA translation and stability in glioblastoma cells. Mol
Cancer Res. 10:143–155. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vo DT, Qiao M, Smith AD, Burns SC, Brenner
AJ and Penalva LO: The oncogenic RNA-binding protein Musashi1 is
regulated by tumor suppressor miRNAs. RNA Biol. 8:817–828. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nakamura M, Okano H, Blendy JA and Montell
C: Musashi, a neural RNA-binding protein required for Drosophila
adult external sensory organ development. Neuron. 13:67–81. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sakakibara S, Nakamura Y, Yoshida T, et
al: RNA-binding protein Musashi family: roles for CNS stem cells
and a subpopulation of ependymal cells revealed by targeted
disruption and antisense ablation. Proc Natl Acad Sci USA.
99:15194–15199. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Glazer RI, Vo DT and Penalva LO: Musashi1:
an RBP with versatile functions in normal and cancer stem cells.
Front Biosci. 17:54–64. 2012. View
Article : Google Scholar
|
|
13
|
Kaneko Y, Sakakibara S, Imai T, et al:
Musashi1: an evolutionally conserved marker for CNS progenitor
cells including neural stem cells. Dev Neurosci. 22:139–153. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Toda M, Iizuka Y, Yu W, et al: Expression
of the neural RNA-binding protein Musashi1 in human gliomas. Glia.
34:1–7. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nikpour P, Emadi-Baygi M, Mohhamad-Hashem
F, Maracy MR and Haghjooy-Javanmard S: MSI1 overexpression in
diffuse type of gastric cancer. Pathol Res Pract. 209:10–13. 2013.
View Article : Google Scholar
|
|
16
|
Shu HJ, Saito T, Watanabe H, et al:
Expression of the Musashi1 gene encoding the RNA-binding protein in
human hepatoma cell lines. Biochem Biophys Res Commun. 293:150–154.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schulenburg A, Cech P, Herbacek I, et al:
CD44-positive colorectal adenoma cells express the potential stem
cell markers musashi antigen (msi1) and ephrin B2 receptor (EphB2).
J Pathol. 213:152–160. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rezza A, Skah S, Roche C, Nadjar J,
Samarut J and Plateroti M: The overexpression of the putative gut
stem cell marker Musashi-1 induces tumorigenesis through Wnt and
Notch activation. J Cell Sci. 123:3256–3265. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang XY, Penalva LO, Yuan H, et al:
Musashi1 regulates breast tumor cell proliferation and is a
prognostic indicator of poor survival. Mol Cancer. 9:2212010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu DC, Yang ZL and Jiang S:
Identification of musashi-1 and ALDH1 as carcinogenesis,
progression, and poor-prognosis related biomarkers for gallbladder
adenocarcinoma. Cancer Biomark. 8:113–121. 2010.PubMed/NCBI
|
|
21
|
Nikpour P, Baygi ME, Steinhoff C, et al:
The RNA binding protein Musashi1 regulates apoptosis, gene
expression and stress granule formation in urothelial carcinoma
cells. J Cell Mol Med. 15:1210–1224. 2011. View Article : Google Scholar
|
|
22
|
Murayama M, Okamoto R, Tsuchiya K, et al:
Musashi-1 suppresses expression of Paneth cell-specific genes in
human intestinal epithelial cells. J Gastroenterol. 44:173–182.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Margaritescu C, Pirici D, Simionescu C and
Stepan A: The utility of CD44, CD117 and CD133 in identification of
cancer stem cells (CSC) in oral squamous cell carcinomas (OSCC).
Rom J Morphol Embryol. 52:985–993. 2011.PubMed/NCBI
|
|
24
|
Janikova M, Skarda J, Dziechciarkova M, et
al: Identification of CD133+/nestin+ putative
cancer stem cells in non-small cell lung cancer. Biomed Pap Med Fac
Univ Palacky Olomouc Czech Repub. 154:321–326. 2010. View Article : Google Scholar
|
|
25
|
Eramo A, Lotti F, Sette G, et al:
Identification and expansion of the tumorigenic lung cancer stem
cell population. Cell Death Differ. 15:504–514. 2008. View Article : Google Scholar
|
|
26
|
Chinn SB, Darr OA, Peters RD and Prince
ME: The role of head and neck squamous cell carcinoma cancer stem
cells in tumorigenesis, metastasis, and treatment failure. Front
Endocrinol (Lausanne). 3:902012.
|
|
27
|
Kayahara T, Sawada M, Takaishi S, et al:
Candidate markers for stem and early progenitor cells, Musashi-1
and Hes1, are expressed in crypt base columnar cells of mouse small
intestine. FEBS Lett. 535:131–135. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu X, Lin F, Fang H, Yang X and Qin L:
Expression of a putative stem cell marker Musashi-1 in endometrium.
Histol Histopathol. 26:1127–1133. 2011.PubMed/NCBI
|
|
29
|
Raji B, Dansault A, Leemput J, et al: The
RNA-binding protein Musashi-1 is produced in the developing and
adult mouse eye. Mol Vis. 13:1412–1427. 2007.PubMed/NCBI
|
|
30
|
Okano H, Kawahara H, Toriya M, Nakao K,
Shibata S and Imai T: Function of RNA-binding protein Musashi-1 in
stem cells. Exp Cell Res. 306:349–356. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sakaguchi H, Yaoi T, Suzuki T, Okano H,
Hisa Y and Fushiki S: Spatiotemporal patterns of Musashi1
expression during inner ear development. Neuroreport. 15:997–1001.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu T, Lan SY, Wu B, et al: Musashi1 and
hairy and enhancer of split 1 high expression cells derived from
embryonic stem cells enhance the repair of small-intestinal injury
in the mouse. Dig Dis Sci. 56:1354–1368. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nagata H, Akiba Y, Suzuki H, Okano H and
Hibi T: Expression of Musashi-1 in the rat stomach and changes
during mucosal injury and restitution. FEBS Lett. 580:27–33. 2006.
View Article : Google Scholar
|
|
34
|
Susaki K, Kaneko J, Yamano Y, et al:
Musashi-1, an RNA-binding protein, is indispensable for survival of
photoreceptors. Exp Eye Res. 88:347–355. 2009. View Article : Google Scholar
|
|
35
|
Arumugam K, Macnicol MC and Macnicol AM:
Autoregulation of Musashi1 mRNA translation during Xenopus oocyte
maturation. Mol Reprod Dev. 79:553–563. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Akasaka Y, Saikawa Y, Fujita K, et al:
Expression of a candidate marker for progenitor cells, Musashi-1,
in the proliferative regions of human antrum and its decreased
expression in intestinal metaplasia. Histopathology. 47:348–356.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bobryshev YV, Tran D, Botelho NK, Lord RV
and Orekhov AN: Musashi-1 expression in atherosclerotic arteries
and its relevance to the origin of arterial smooth muscle cells:
histopathological findings and speculations. Atherosclerosis.
215:355–365. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lovell MA and Markesbery WR: Ectopic
expression of Musashi-1 in Alzheimer disease and Pick disease. J
Neuropathol Exp Neurol. 64:675–680. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vo DT, Subramaniam D, Remke M, et al: The
RNA-binding protein Musashi1 affects medulloblastoma growth via a
network of cancer-related genes and is an indicator of poor
prognosis. Am J Pathol. 181:1762–1772. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nishimura S, Wakabayashi N, Toyoda K,
Kashima K and Mitsufuji S: Expression of Musashi-1 in human normal
colon crypt cells: a possible stem cell marker of human colon
epithelium. Dig Dis Sci. 48:1523–1529. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ke J, Wu X, He X, et al: A subpopulation
of CD24+ cells in colon cancer cell lines possess stem
cell characteristics. Neoplasma. 59:282–288. 2012. View Article : Google Scholar
|
|
42
|
Schneider M, Huber J, Hadaschik B, Siegers
GM, Fiebig HH and Schuler J: Characterization of colon cancer
cells: a functional approach characterizing CD133 as a potential
stem cell marker. BMC Cancer. 12:962012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lin L, Fuchs J, Li C, Olson V, Bekaii-Saab
T and Lin J: STAT3 signaling pathway is necessary for cell survival
and tumorsphere forming capacity in
ALDH+/CD133+ stem cell-like human colon
cancer cells. Biochem Biophys Res Commun. 416:246–251. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kanemura Y, Mori K, Sakakibara S, et al:
Musashi1, an evolutionarily conserved neural RNA-binding protein,
is a versatile marker of human glioma cells in determining their
cellular origin, malignancy, and proliferative activity.
Differentiation. 68:141–152. 2001. View Article : Google Scholar : PubMed/NCBI
|